Embed Size (px)
Citation preview

Directed Differentiation and Transplantation of Human EmbryonicStem Cell-Derived Motoneurons
HYOJIN LEE,a,b GEORGE AL SHAMY,b YECHIEL ELKABETZ,a,b CLAUDE M. SCHOFIELD,d NEIL L. HARRSION,d
GEORGIA PANAGIOTAKOS,b NICHOLAS D. SOCCI,c VIVIANE TABAR,b LORENZ STUDERa,b
aDevelopmental Biology Program, bDivision of Neurosurgery, and cComputational Biology Center, Sloan-KetteringInstitute, New York, New York, USA; dDepartments of Anesthesiology and Pharmacology, Weill Medical Collegeof Cornell University, New York, New York, USA
Key Words. Embryonic stem cells • Neural differentiation • Transplantation • Human embryonic stem cells
ABSTRACT
Motoneurons represent a specialized class of neurons essen-tial for the control of body movement. Motoneuron loss isthe cause of a wide range of neurological disorders includingamyotrophic lateral sclerosis and spinal muscular atrophy.Embryonic stem cells are a promising cell source for thestudy and potential treatment of motoneuron diseases. Here,we present a novel in vitro protocol of the directed differ-entiation of human embryonic stem cells (hESCs) into en-graftable motoneurons. Neural induction of hESCs was in-duced on MS5 stromal feeders, resulting in the formation ofneural rosettes. In response to sonic hedgehog and retinoicacid, neural rosettes were efficiently directed into spinalmotoneurons with appropriate in vitro morphological, phys-
iological, and biochemical properties. Global gene expres-sion analysis was used as an unbiased measure to confirmmotoneuron identity and type. Transplantation of motoneu-ron progeny into the developing chick embryo resulted inrobust engraftment, maintenance of motoneuron phenotype,and long-distance axonal projections into peripheral hosttissues. Transplantation into the adult rat spinal cordyielded neural grafts comprising a large number of humanmotoneurons with outgrowth of choline acetyltransferasepositive fibers. These data provide evidence for in vivosurvival of hESC-derived motoneurons, a key requirementin the development of hESC-based cell therapy in motoneu-ron disease. STEM CELLS 2007;25:1931–1939
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Motoneurons are the key effector cell type for control of motorfunction, and loss of motoneurons is associated with a numberof debilitating diseases such as amyotrophic lateral sclerosis(ALS) and spinal muscular atrophy [1, 2]. Motoneurons are alsoregarded as a great model for probing mechanisms of vertebratecentral nervous system (CNS) development, and the transcrip-tional pathways that guide motoneuronal specification are wellcharacterized [3]. Recent studies have demonstrated the in vitroderivation of motoneurons from mouse [4, 5] and human em-bryonic stem cells (hESCs) [6, 7]. Current hESC-derived mo-toneuron differentiation protocols are based on embryoid bodymediated neural induction followed by exposure to definedmorphogens [6, 7] such as sonic hedgehog (SHH) and retinoicacid (RA) acting as ventralizing and caudalizing factors, respec-tively. It has been suggested that the ability to undergo mo-toneuron specification under these conditions is temporally re-stricted to the earliest stages of neural induction [6, 7].Characterization of these cells in vitro and in vivo has beenlimited. Furthermore, there are currently no published data onthe ability of hESC-derived motoneurons to secrete acetylcho-line, the key neurotransmitter of spinal motoneurons, and tosurvive and maintain motoneuron characteristics in the devel-oping or adult cord. In vivo survival and the ability for ortho-
topic integration are key requirements for the future applicationsin animal models of motoneuron disease. Here, we present anovel in vitro protocol for the efficient generation of hESC-derived motoneurons. Furthermore, we demonstrate in vivosurvival and maintenance of motoneuron phenotype upon trans-plantation into the developing chick and adult rat spinal cord.
MATERIALS AND METHODS
Culture of Undifferentiated hESC and NonhumanPrimate ESC LinesH1 (WA-01, passages [P]36–52), H9 (WA-09, P44–60), RUES1-EGFP (P39–45), and the cynomolgus parthenogenetic line (Cyno1,P42–59) were cultured on mitotically inactivated mouse embry-onic fibroblasts (Specialty Media, Phillipsburg, NJ, http://www.specialtymedia.com). Undifferentiated hESCs and monkey ESCswere maintained under growth conditions and passaging techniquesdescribed previously for each given line: H1, H9, and RUES1-eGFP[8]; Cyno1 [9].
Neural Induction and Motoneuron DifferentiationMS5 stromal cells were maintained in �-minimal essential mediumcontaining 10% fetal bovine serum and 2 mM L-glutamine asdescribed previously [5]. Neural differentiation of hESCs was in-duced via coculture on MS5, and hESCs were plated at 5–20 � 103
Correspondence: Lorenz Studer, M.D., Developmental Biology & Neurosurgery, Memorial Sloan-Kettering Cancer Center, 1275 York Ave.,Box 256, New York, New York 10021, USA. Telephone: 212-639-6126; Fax: 212-717-3642; e-mail: [email protected] ReceivedFebruary 5, 2007; accepted for publication April 20, 2007; first published online in STEM CELLS EXPRESS May 3, 2007. ©AlphaMed Press1066-5099/2007/$30.00/0 doi: 10.1634/stemcells.2007-0097
EMBRYONIC STEM CELLS
STEM CELLS 2007;25:1931–1939 www.StemCells.com

cells on a confluent layer of irradiated (50 Gy) stromal cells in 6-cmcell culture plates in serum replacement medium containing Dul-becco’s modified Eagle’s medium, 15% knockout serum replace-ment (Invitrogen, Carlsbad, CA, http://www.invitrogen.com), 2 mML-glutamine, and 10 �M �-mercaptoethanol. At day 16, cultureswere switched to N2 medium modified according to Johe et al. [10].In some of the experiments, Noggin was added to the culturemedium during neural induction on MS5. Neural rosettes weremechanically isolated around day 28 of differentiation. Isolatedrosettes were gently replated on polyornithine (15 �g/ml)/laminin(1 �g/ml) coated culture dishes and maintained in N2 mediumsupplemented with ascorbic acid (AA) and brain-derived neurotro-phic factor (BDNF) in the presence of RA/SHH (termed passage 1[P1]). After 7–10 days (�80% confluence), cells were mechanicallypassaged after exposure to Ca2/Mg2-free Hanks’ balanced salt so-lution (HBSS) for 1 hour at room temperature and spun at 200g for5 minutes. Cells were resuspended in N2 medium supplementedwith BDNF and AA and replated again onto polyornithine/laminincoated culture dishes (50–100 � 103 cells per cm2) in the presenceof RA/SHH (passage 2 [P2]). After an additional 10–15 days ofculture, cells were differentiated in the absence of RA/SHH but inthe presence of AA, BDNF, and glial cell line-derived neurotrophicfactor (GDNF) (R&D Systems Inc., Minneapolis, http://www.rndsystems.com). Medium was changed every 2–3 days, and growthfactors were added in various combinations and at various timepoints as described in the main text: SHH (500 ng/ml), BDNF (20ng/ml), GDNF (20 ng/ml), Noggin (500 ng/ml) (all from R&DSystems), all-trans-Retinoic acid (1 �M), and AA (0.2 mM) (bothfrom Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com). Dif-ferentiation of hESCs into midbrain dopamine neurons for Af-fymetrix (Santa Clara, California, http://www.affymetrix.com) geneexpression analysis followed our previously published protocol[11].
ImmunocytochemistryCells were fixed in 4% paraformaldehyde/0.15% picric acid andstained with the following primary antibodies: rabbit polyclonalantibodies: Sox1 (United States Biological, Swampscott, Massachu-setts, http://www.usbio.net; 1:200), Pax6 and �-III Tubulin (Co-vance, Princeton, NJ, http://www.covance.com; 1:75, 1:2,000),Otx2 (F. Vaccarino; 1:1,000), Nkx6.1 (S. Heller; 1:5,000), BF1(1:1,000); goat polyclonal antibodies: choline acetyl transferase(ChAT) (Chemicon, Temecula, CA, http://www.chemicon.com;1:200); mouse monoclonal antibodies (IgG): Pax6, HB9 (MNR2),Isl1, Pax7, Lhx3 (Lim3), Nkx2.2 and HoxB4 (Developmental Stud-ies Hybridoma Bank, Iowa City, IA, http://www.uiowa.edu/�dshbwww; l:75), Tuj1 and HoxC8 (Covance; 1:500, 1:200), Olig2(Qing Richard Lu; 1:1,000), human neural cell adhesion molecule(hNCAM, clone Eric1; Santa Cruz Biotechnology Inc., Santa Cruz,CA, http://www.scbt.com; 1:200), human nuclear antigen (Chemi-con; 1:200); guinea pig: HoxC6 (T. Jessell, 1:8,000). AppropriateAlexa488 and Alexa555 or Alexa568 labeled secondary antibodies(Molecular Probes, Eugene, OR, http://probes.invitrogen.com), bi-otinylated rabbit anti-goat followed by Alexa555 or Alexa568, andAlexa488-StreptAvidin conjugate and 4,6-diamidino-2-phenylin-dole (DAPI) counterstain were used for visualization. Image acqui-sition for cultured cells was performed using an inverted OlympusIX71 epifluorescence microscope with appropriate filter sets(Chroma, Rockingham, VT, http://www.chroma.com) using singlechannel acquisition on a Hamamatsu Orca CCD camera. Histolog-ical sections were imaged on an upright Olympus epifluorescencemicroscope (AX70) linked to a Photometrics CoolSNAP fx, ac-quired as single channel images, and merged using IPLab version3.6. Confocal images such as Figure 5C were obtained via opticalsectioning at 0.8 �m on a Leica (Heerbrugg, Switzerland, http://www.leica.com) TCS SO2 AOBS set-up and reconstructed using aLeica Confocal Software Lite package.
Reverse Transcription-Polymerase Chain ReactionTotal RNA was extracted using the RNeasy kit and DNase Itreatment (Qiagen, Hilden, Germany, http://www1.qiagen.com) toavoid genomic contamination. Undifferentiated hESCs and hESCprogeny at day 28 of differentiation were manually dissected off the
feeder layer to avoid contamination with stromal cells. Total RNA(1–2 �g each) was reverse transcribed (SuperScript; Invitrogen).Polymerase chain reaction (PCR) conditions were optimized, andlinear amplification range was determined for each primer by vary-ing annealing temperature and cycle number. PCR products wereidentified by size. All samples were treated with DNase to eliminatepotential contaminating genomic DNA. Primer sequences, cyclenumbers, and annealing temperatures are provided as supplementaldata (supplemental online Table 1). All reverse transcription-PCRdata shown are from H9; similar data were derived from H1 andCyno1.
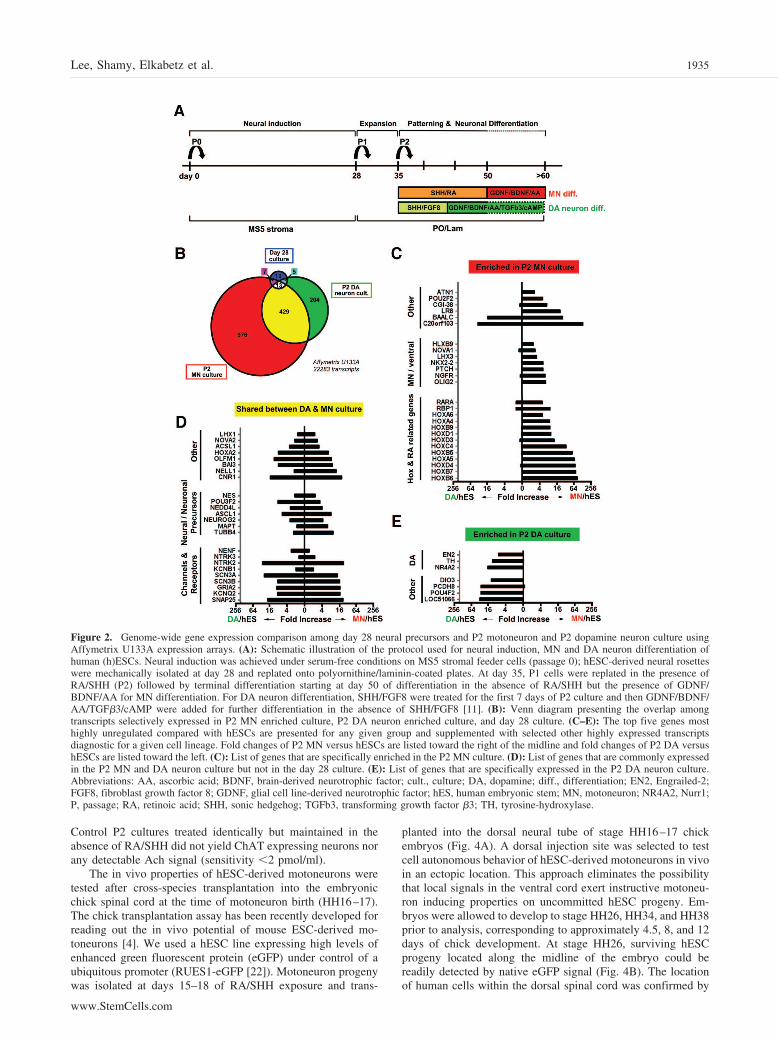
Affymetrix AnalysisTotal RNA (5 �g) from day 15 of P2 motoneuron (MN) culture, day14 of P2 dopamine (DA) culture, three samples of day 28 neuralprecursors cocultured on MS5 stroma, and three samples of undif-ferentiated hESCs (H1; passages 42–46) were processed by theMemorial Sloan-Kettering Cancer Center Genomics Core Facilityand hybridized on Affymetrix U133A human oligonucleotide ar-rays. All analyses were done using the R statistical system and theBioConductor (http://www.bioconductor.org) microarray analysispackages. The raw Affymetrix cell files were normalized and quan-titated using the GC-RMA package, and the logarithm (base 2) ofthe expression levels was used in the subsequent analysis. Todetermine genes that were differentially expressed among each ofthe three differentiated samples (P2 MN, P2 DA neuron, and day 28neural precursors) and the pooled data from the three undifferenti-ated samples of hESCs, a variant of the t test was used as imple-mented in the LIMMA package from BioConductor. In this method,the variance is corrected by adding a term that is computed from theglobal noise levels in the samples. This term improves robustness ofthe variance estimate, especially when dealing with small samplesizes. To account for multiple testing, we used the false discoveryrate (FDR) method and applied a cutoff of 0.05 in the FDR to eachlist. This gave us three lists of sizes 1,426 (P2 MN), 651 (P2 DAneurons), and 44 (day 28 neural precursors). The larger number oftranscripts in P2 MN and P2 DA versus day 28 neural precursor(group) indicates that these two groups are more distinct fromundifferentiated hESCs than day 28 neural precursors.
ElectrophysiologyFor electrophysiological recordings, we mechanically transferred P2cells at day 15 of differentiation onto astrocyte feeders that werederived in an autologous fashion from the respective hESC line orCyno1 following protocols described previously [11]. Recordingswere performed 10–15 days after replating in differentiation me-dium containing AA, BDNF, and GDNF as described above. Cellswere placed on the stage of an inverted microscope (Axiovert 200;Carl Zeiss, Jena, Germany, http://www.zeiss.com), and the culturemedium was replaced by a physiological saline solution containing145 mM NaCl, 3 mM KCl, 1.5 mM CaCl2, 1 mM MgCl2, 5 mMglucose, and 10 mM HEPES (pH 7.4) and having 320–330 mOsmosmolarity. Recordings were performed at room temperature usingthe whole-cell patch-clamp technique. Patch pipettes were of resis-tance 4–5 M� and contained 140 mM potassium gluconate, 10 mMNaCl, 1 mM CaCl2, 10 mM HEPES, 0.2 mM EGTA, 3 mMATP-Mg, and 0.4 mM GTP (pH 7.3), and osmolarity was adjustedto 295–305 mOsm with sucrose. Pipette-to-bath resistance wastypically 5–10 M�. Data were acquired in current-clamp modeusing the HEKA EPC 10 amplifier and acquisition system (HEKA,Lambrecht, Germany, http://www.heka.com).
High-Performance Liquid Chromatography AnalysisHigh-performance liquid chromatography (HPLC) analysis wasused to quantify acetylcholine (Ach) secreted from the hESC- orCyno1-derived motoneurons. Ach was identified by electrochemicaldetection after enzymatic conversion of Ach into H2O2 by a solid-phase reactor (containing immobilized choline oxidase and acetyl-cholinesterase enzymes) [12]. Conditioned medium was collected at50–65 days of differentiation for H9 (WA-09) cells and 40–55 daysof differentiation for Cyno1 cells and stored at �80°C prior toanalysis. Separation of the injected samples (25 �l per injection;
1932 Human Embryonic Stem Cell-Derived Motoneurons

ESA [Chelmsford, MA, http://www.esainc.com] Autosampler 540)was achieved by isocratic elution in a mobile phase containing 100mM Na2HPO4 (Sigma) (pH 8.0), 2 mM 1-Octanesulfonic acidsodium salt (Fluka, Buchs, Switzerland, http://www.sigmaaldrich.com/Brands/Fluka_Riedel_Home.html), and 0.005% reagent MB(ESA) at 0.3 ml/minute. The oxidative potential of the analyticalcell (ESA model 5040 with platinum target, Coulochem II) was setat 300 mV. Results were validated by coelution with acetylcholinestandards under varying buffer conditions and detector settings. Thelimit of detection in the system was 50 fmol per injection.
Cell Counts and Statistical AnalysesData for percentage of HB9/DAPI or HB9/�-III Tubulin cells werederived from a total of three independent experiments for each H9(WA-09) and Cyno1 line. Cells were selected for quantification inuniform random fashion (fractionator). Each field was scored firstfor DAPI positive nuclei or �-III Tubulin positive cells and subse-quently HB9/DAPI or HB9/�-III Tubulin colocalization. For eachexperiment, an average of greater than 8,000 cells were scored. Thetotal number of cells analyzed in this study exceeded 25,000 cells.Percentages of HB9/DAPI or HB9/�-III Tubulin cells were com-pared with analysis of variance and Newman-Keuls post hoc anal-ysis (Statistica 5.5; StatSoft, Tulsa, OK, http://www.statsoft.com).Data are presented as mean � SEM.
In Ovo TransplantationFertile eggs (CBT Farms, Chestertown, MD) were incubated at37°C in a humidified incubator. hESC-derived motoneurons at days15–18 in P2 stage were dissociated and transplanted into stage16-17 chick spinal cord. Graft site extended over 1–2 somites. Asmall mechanical lesion was performed to generate space for graft.Eggs were placed back into the incubator for up to 9 days post-transplantation. Embryos were harvested and fixed with 4% para-formaldehyde for 1 day, incubated in 30% sucrose for 1–2 days, andembedded in optimum cutting temperature compound and cryosec-tioned for further immunohistochemical analysis.
In Vivo Transplantation into Adult Rat Spinal CordThree-month-old Sprague-Dawley rats were anesthetized with amixture of ketamine/xylazine and placed in a stereotactic frame(Kopf, Tujunga, California, http://www.kopfinstruments.com). Theskin over the cervical area was shaved and prepped in Betadinesolution. An incision was performed through skin and fascia downto the C2 spinous process. The paraspinal muscles were dissectedoff the spinous process and laminae and the posterior bone elementswere removed using a dental drill and small bone rongeurs. Themedian dorsal groove and vessel were used as the midline reference.A pulled glass microcapillary and pump were used to deliver thecells. Cells were harvested at day 15 of P2 differentiation (SHH/RAstage) and mechanically triturated after incubation in Ca2/Mg2-freeHBSS for 1 hour at room temperature. Cells were resuspended at100,000 cells per microliter for transplantation. Bilateral injectionsdelivering 40,000 cells each were performed at the following coor-dinates: 0.5 mm lateral to the midline and 1.2 mm in depth fromsurface [13]. The dura was punctured with a 25-gauge needle tofacilitate smooth entry of the microcapillary, which remained inplace postinjection for a period of 10 minutes. The muscles wereapproximated with a 000 Vicryl suture and the skin with staples.The animals did not experience any obvious neurological deficitpost operation. All animals received 20 mg/kg cyclosporin intra-peritoneal 1 day before surgery and for the duration of the experi-ment. All animals received Sulfatrim medicated feeds as of the dayof surgery. Crede maneuver (massaging of the bladder) was per-formed on all animals as a precautionary measure on day 1. Animalswere sacrificed on day 1 (two animals) and 2 weeks (two animals)and 6 weeks (six animals) after transplantation.
RESULTS
We have previously described MS5 stromal feeder based cocul-ture strategies for the directed differentiation of mouse and
hESCs toward defined neural progeny [5, 11, 14]. The MS5protocol belongs to the category of “stromal cell-derived induc-ing activity” based neural induction strategies [15, 16] and canbe adapted to the generation of a wide range of neuronal andglial cell types in mouse ESCs [5]. During hESC differentiation,MS5 based protocols yield large numbers of columnar neurepi-thelial structures termed “rosettes,” which can be further pat-terned and differentiated into various neuronal and glial cellfates such as midbrain dopamine neurons [11] or multipotentneural precursor cells [14]. Exposure of hESC-derived neuronalrosettes to fibroblast growth factor 8 (FGF8) and SHH enrichesfor ventral midbrain precursor cells that can be further differ-entiated into postmitotic dopamine neuron progeny. These datasuggest that neural rosettes appropriately respond to develop-mental cues that specify anterior-posterior (AP) and dorsoven-tral (DV) identity during neural development. Here, we testedwhether MS5 based neural induction of hESCs can be adapted forthe efficient derivation of functional and engraftable motoneurons.
Neural induction of hESCs resulted in the formation ofneural rosettes, as described previously [11], expressing Pax6and Sox1 (Fig. 1A). These two transcription factors are knownmarkers of the neural plate stage in mouse development [17–19]. At day 28 of differentiation on MS5, most cells in neuralrosettes coexpressed Pax6 and Sox1 (Fig. 1A) and were stronglypositive for the markers associated with anterior neural identitysuch as BF1 (FOXG1B) [20] and Otx2 (data not shown). Incontrast, there was a lack of expression of posterior neuralmarkers such as Gbx2 (anterior hindbrain) and HoxB4 (poste-
Figure 1. Neural patterning and motoneuron derivation from human(h)ESCs. (A): Pax6 and Sox1 expression in neural rosettes at day 28 ofdifferentiation on MS5 stroma. (B–C): Representative anterior-posterior(B) and dorsoventral (C) markers expressed in P2 rosette progeny (day45 of differentiation) in the presence or absence (inset) of RA/SHH.(D–G): Immunocytochemical analysis of motoneuron markers at day 15(D, E) and day 25 (F, G) of differentiation during P2 stage. (H):Percentage of HB9/DAPI cells for hESC and Cyno1 progeny at P2 after15 days of RA/SHH treatment or under control conditions in the absenceof RA/SHH. (I): Gene expression analysis of motoneuron markersduring differentiation; day 0 (undifferentiated hESCs) and day 28 ofdifferentiation on MS5; days 5, 10, and 15 during P2 differentiation. Forschematic culture protocols, see Figure 2A. Scale bar in (A) correspondsto 200 �m for (A–C); 100 �m for (D, E); 25 �m for (F); 30 �m for (G).Abbreviations: ChAT, choline acetyl transferase; DAPI, 4,6-diamidino-2-phenylindole; GAPDH, glyceraldehyde-3-phosphate dehydrogenase;hES, human embryonic stem; MN, motoneuron; P2, passage 2; RA,retinoic acid; SHH, sonic hedgehog; �Tub, �-III Tubulin; VAChT,vesicular acetylcholine transporter.
1933Lee, Shamy, Elkabetz et al.
www.StemCells.com

rior hindbrain and spinal cord) (data not shown). These datasuggest that, under default conditions, neural rosettes inducedon MS5 progress toward an anterior neural patterning state.
We next tested whether progressive expression of anteriorneural markers in rosettes can be modulated by appropriateextrinsic cues. Neural rosettes were mechanically isolated andmaintained under feeder-free conditions on polyornithine/lami-nin-coated plates (P1 cultures). After 7 days of P1 culture, cellswere replated to the P2 stage. Immunocytochemical analysisrevealed maintenance of BF1 and Otx2 (Fig. 1B, inset) expres-sion at day 10 of P2 cultures in the absence of extrinsicallyadded patterning cues. However, exposure of P2 cultures toRA/SHH led to a dramatic decrease in BF1 with a correspond-ing increase in hindbrain and spinal cord markers such asHoxB4 and HoxC8 (Fig. 1B and supplemental online Fig. 1).
These data indicate that RA/SHH exposure is sufficient tosuppress BF1 expression and to induce caudal identity markers.Progeny derived from human neural rosettes also showed ap-propriate modulation of DV fate markers in response to SHHexposure. Expression of ventral identity genes characteristic ofthe caudal CNS such as Nkx6.1 and Nkx2.2 required exposureof both RA and SHH (Fig. 1C and supplemental online Fig. 1).
We next tested whether RA/SHH-mediated induction ofcaudal and ventral patterning markers is associated with mo-toneuron specification and terminal differentiation into postmi-totic motoneuron progeny. P1 cells were passaged and main-tained for an additional 5–15 days in the presence of RA/SHH(P2 cultures). Gene expression analysis (Fig. 1I) confirmed theinduction of transcripts related to motoneuron precursor cells(Nkx6.1, Olig2), early postmitotic motoneurons (neurogenin 2,Isl1), and more mature motoneurons (ChAT and vesicular ace-tylcholine transporter). Immunocytochemical studies at P2 con-firmed the presence of large numbers of Olig2� motoneuronprecursors emerging from neuroepithelial structures. Cells ex-pressing the postmitotic motoneuron marker Isl1 were arrangedin the periphery of the Olig2� precursor cell domains (Fig. 1D).After 10 days of P2 exposure to RA/SHH, large numbers of cellsexpressing the somatic motoneuron marker HB9� were de-tected, and the percentage of HB9� cells continued to increasein number until day 15 of RA/SHH treatment (Fig. 1E). Uponfurther differentiation in the absence of RA/SHH but in thepresence of GDNF, BDNF, and AA, expression of more maturemotoneuron markers was observed, including ChAT, the keyenzyme required for the synthesis of Ach. An average of 26% oftotal cells expressed ChAT after 15 days of RA/SHH exposurefollowed by 5 days of differentiation in GDNF, BDNF, and AA.Less than 5% ChAT� cells were present prior to day 10 ofSHH/RA treatment. Cells treated identically but not subjected toSHH/RA exposure had less than 1% of cells positive for ChAT(supplemental online Fig. 2). Many ChAT� cells coexpressedLhx3 (Fig. 1F). Coexpression of ChAT and HB9 confirmedmotoneuron identity of hESC progeny (Fig. 1G). Expression ofLhx3 in motoneuron progeny suggests that a subset of hESC-derived motoneurons expresses medial motor column markers,similar to the data reported recently for mouse embryonic stemcell-derived motoneurons [21]. The protocol was highly repro-ducible and yielded similar results for motoneuron progenyfrom a variety of hESC lines such as H1, H9, and RUES1-eGFP[22] (data not shown). The average number of HB9� cellsderived from hESC line (H9) was 20% out of total (DAPI�)cells (Fig. 1H). We next confirmed the robustness of the mo-toneuron induction protocol in Cyno1, a parthenogenetic mon-key ESC-like cell line [9, 23]. Motoneuron yield in Cyno1 was43% HB9� cells out of the total (DAPI�) population (Fig. 1H)and 65% of all (�-III Tubulin�) neurons.
Our study presents an MS5 based neural induction protocolfor the efficient derivation of hESC-derived motoneurons. Mo-
toneuron differentiation of hESCs in vitro has been character-ized by the expression of a distinct set of markers such as HB9,Lhx3, and Isl1 [6, 7]. However, there has been no previousreport describing unbiased global expression profiles in specifichESC-derived neuronal populations. We performed genome-wide expression analysis using oligonucleotide arrays (Af-fymetrix U133A) to test whether known and novel candidatemarkers of motoneuron identity can be identified in hESCprogeny. We compared expression profiles of hESC-derivedcultures enriched in motoneuron (day 15 in P2 culture) versusmidbrain dopamine neuron progeny (day 14 in P2 culture). Aschematic representation of the experimental conditions is pro-vided in Figure 2A. Transcripts related to hESC-derived mo-toneurons were defined as those with a signal threshold ofgreater than five times the levels detected in three independentsamples of undifferentiated hESCs. The same criteria were usedto define transcripts related to midbrain DA neuron differenti-ation and day 28 early neural precursor cells. A Venn diagramwas drawn among all the transcripts related to at least one of thethree categories of neural progeny. Based on this analysis, 976transcripts were selectively enriched in motoneuron cultures,204 transcripts in DA neuron cultures, and 19 transcripts in day28 neural precursor cultures. A total of 429 transcripts wereshared between P2 motoneuron and P2 DA neuron cultures (Fig.2B). Motoneuron specific markers could be grouped into vari-ous categories including genes associated with ventral neuralpatterning and transcripts related to Hox and RA signaling. Keymotoneuron markers such as HB9 (HLXB9) and Lhx3 were alsogrouped among motoneuron specific transcripts, validating theglobal expression data and independently confirming motoneu-ron identity of hESC progeny (Fig. 2C). Transcripts sharedbetween motoneuron and midbrain DA neuron cultures butabsent in early neural precursors and in undifferentiated hESCsincluded ion channel related markers defining pan-neuronalidentity, a number of neurotrophic factor and neurotrophic fac-tor receptor genes, and multiple neural and neuronal precursorgenes (Fig. 2D). Key DA neuron specific transcripts comprisedmany of the known markers of midbrain DA neuron identitysuch as tyrosine-hydroxylase, Nurr1, and Engrailed-2 (Fig. 2E).
Work with mouse ESCs suggests that RA/SHH treatmentinduces motoneurons of rostral spinal cord identity expressingHoxC5 and HoxC6 [4]. In contrast, work with hESC-derivedmotoneurons suggested the derivation of motoneuron progenywith more caudal characteristics as assessed by the expressionof HoxC8 [6]. However, the gene expression data presentedindicate biased expression toward class 4–6 Hox genes (Fig.2C) comparable with the profile reported for mouse ESC-de-rived motoneurons. Using immunocytochemistry, we observedthat our hESC differentiation protocol yields motoneurons ofboth rostral (HoxC6) and more caudal (HoxC8) identity (Fig.3A–3C). Overlap of HoxC6 and HoxC8 is indicative of poste-rior type brachial motoneuron identity [24, 25]. We next testedin vitro function of motoneurons derived from H9 and Cyno1using patch clamp studies (Fig. 3D). Single cell recordingsdemonstrated the induction of trains of action potentials inresponse to depolarizing currents. Mature electrophysiologicalsignatures required coculture with astrocytes that were derivedin an autologous fashion from the respective hESC line orCyno1 following protocols described previously [11]. Biochem-ical evidence of motoneuron function was obtained by measur-ing Ach release in the conditioned medium of hESC- andCyno1-derived motoneuron cultures after 15 days of RA/SHHexposure followed by 10 days of differentiation in the presenceof AA, BDNF, and GDNF (Fig. 3E). HPLC analysis usingelectrochemical detection after enzymatic conversion of Achinto H2O2 by a solid-phase reactor [12] detected an average of7.5–12 pmol Ach per milliliter medium (24 hours conditioning).
1934 Human Embryonic Stem Cell-Derived Motoneurons
Control P2 cultures treated identically but maintained in theabsence of RA/SHH did not yield ChAT expressing neurons norany detectable Ach signal (sensitivity �2 pmol/ml).
The in vivo properties of hESC-derived motoneurons weretested after cross-species transplantation into the embryonicchick spinal cord at the time of motoneuron birth (HH16–17).The chick transplantation assay has been recently developed forreading out the in vivo potential of mouse ESC-derived mo-toneurons [4]. We used a hESC line expressing high levels ofenhanced green fluorescent protein (eGFP) under control of aubiquitous promoter (RUES1-eGFP [22]). Motoneuron progenywas isolated at days 15–18 of RA/SHH exposure and trans-
planted into the dorsal neural tube of stage HH16–17 chickembryos (Fig. 4A). A dorsal injection site was selected to testcell autonomous behavior of hESC-derived motoneurons in vivoin an ectopic location. This approach eliminates the possibilitythat local signals in the ventral cord exert instructive motoneu-ron inducing properties on uncommitted hESC progeny. Em-bryos were allowed to develop to stage HH26, HH34, and HH38prior to analysis, corresponding to approximately 4.5, 8, and 12days of chick development. At stage HH26, surviving hESCprogeny located along the midline of the embryo could bereadily detected by native eGFP signal (Fig. 4B). The locationof human cells within the dorsal spinal cord was confirmed by
Figure 2. Genome-wide gene expression comparison among day 28 neural precursors and P2 motoneuron and P2 dopamine neuron culture usingAffymetrix U133A expression arrays. (A): Schematic illustration of the protocol used for neural induction, MN and DA neuron differentiation ofhuman (h)ESCs. Neural induction was achieved under serum-free conditions on MS5 stromal feeder cells (passage 0); hESC-derived neural rosetteswere mechanically isolated at day 28 and replated onto polyornithine/laminin-coated plates. At day 35, P1 cells were replated in the presence ofRA/SHH (P2) followed by terminal differentiation starting at day 50 of differentiation in the absence of RA/SHH but the presence of GDNF/BDNF/AA for MN differentiation. For DA neuron differentiation, SHH/FGF8 were treated for the first 7 days of P2 culture and then GDNF/BDNF/AA/TGF�3/cAMP were added for further differentiation in the absence of SHH/FGF8 [11]. (B): Venn diagram presenting the overlap amongtranscripts selectively expressed in P2 MN enriched culture, P2 DA neuron enriched culture, and day 28 culture. (C–E): The top five genes mosthighly unregulated compared with hESCs are presented for any given group and supplemented with selected other highly expressed transcriptsdiagnostic for a given cell lineage. Fold changes of P2 MN versus hESCs are listed toward the right of the midline and fold changes of P2 DA versushESCs are listed toward the left. (C): List of genes that are specifically enriched in the P2 MN culture. (D): List of genes that are commonly expressedin the P2 MN and DA neuron culture but not in the day 28 culture. (E): List of genes that are specifically expressed in the P2 DA neuron culture.Abbreviations: AA, ascorbic acid; BDNF, brain-derived neurotrophic factor; cult., culture; DA, dopamine; diff., differentiation; EN2, Engrailed-2;FGF8, fibroblast growth factor 8; GDNF, glial cell line-derived neurotrophic factor; hES, human embryonic stem; MN, motoneuron; NR4A2, Nurr1;P, passage; RA, retinoic acid; SHH, sonic hedgehog; TGFb3, transforming growth factor �3; TH, tyrosine-hydroxylase.
1935Lee, Shamy, Elkabetz et al.
www.StemCells.com
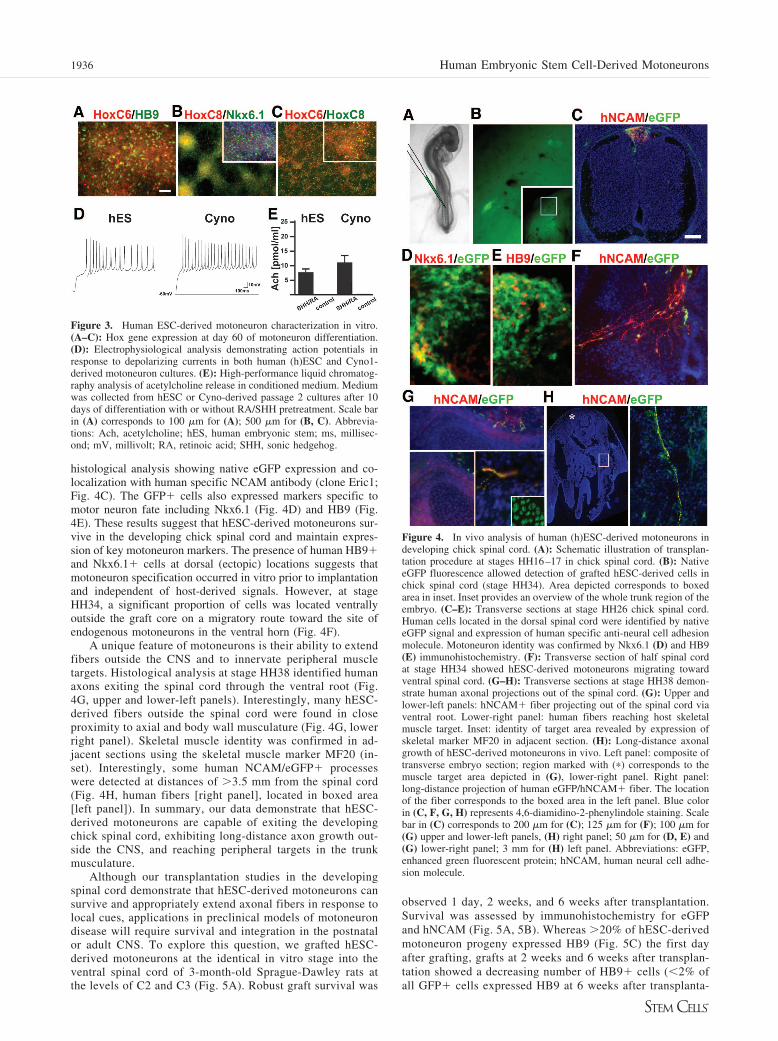
histological analysis showing native eGFP expression and co-localization with human specific NCAM antibody (clone Eric1;Fig. 4C). The GFP� cells also expressed markers specific tomotor neuron fate including Nkx6.1 (Fig. 4D) and HB9 (Fig.4E). These results suggest that hESC-derived motoneurons sur-vive in the developing chick spinal cord and maintain expres-sion of key motoneuron markers. The presence of human HB9�and Nkx6.1� cells at dorsal (ectopic) locations suggests thatmotoneuron specification occurred in vitro prior to implantationand independent of host-derived signals. However, at stageHH34, a significant proportion of cells was located ventrallyoutside the graft core on a migratory route toward the site ofendogenous motoneurons in the ventral horn (Fig. 4F).
A unique feature of motoneurons is their ability to extendfibers outside the CNS and to innervate peripheral muscletargets. Histological analysis at stage HH38 identified humanaxons exiting the spinal cord through the ventral root (Fig.4G, upper and lower-left panels). Interestingly, many hESC-derived fibers outside the spinal cord were found in closeproximity to axial and body wall musculature (Fig. 4G, lowerright panel). Skeletal muscle identity was confirmed in ad-jacent sections using the skeletal muscle marker MF20 (in-set). Interestingly, some human NCAM/eGFP� processeswere detected at distances of �3.5 mm from the spinal cord(Fig. 4H, human fibers [right panel], located in boxed area[left panel]). In summary, our data demonstrate that hESC-derived motoneurons are capable of exiting the developingchick spinal cord, exhibiting long-distance axon growth out-side the CNS, and reaching peripheral targets in the trunkmusculature.
Although our transplantation studies in the developingspinal cord demonstrate that hESC-derived motoneurons cansurvive and appropriately extend axonal fibers in response tolocal cues, applications in preclinical models of motoneurondisease will require survival and integration in the postnatalor adult CNS. To explore this question, we grafted hESC-derived motoneurons at the identical in vitro stage into theventral spinal cord of 3-month-old Sprague-Dawley rats atthe levels of C2 and C3 (Fig. 5A). Robust graft survival was
observed 1 day, 2 weeks, and 6 weeks after transplantation.Survival was assessed by immunohistochemistry for eGFPand hNCAM (Fig. 5A, 5B). Whereas �20% of hESC-derivedmotoneuron progeny expressed HB9 (Fig. 5C) the first dayafter grafting, grafts at 2 weeks and 6 weeks after transplan-tation showed a decreasing number of HB9� cells (�2% ofall GFP� cells expressed HB9 at 6 weeks after transplanta-
Figure 3. Human ESC-derived motoneuron characterization in vitro.(A–C): Hox gene expression at day 60 of motoneuron differentiation.(D): Electrophysiological analysis demonstrating action potentials inresponse to depolarizing currents in both human (h)ESC and Cyno1-derived motoneuron cultures. (E): High-performance liquid chromatog-raphy analysis of acetylcholine release in conditioned medium. Mediumwas collected from hESC or Cyno-derived passage 2 cultures after 10days of differentiation with or without RA/SHH pretreatment. Scale barin (A) corresponds to 100 �m for (A); 500 �m for (B, C). Abbrevia-tions: Ach, acetylcholine; hES, human embryonic stem; ms, millisec-ond; mV, millivolt; RA, retinoic acid; SHH, sonic hedgehog.
Figure 4. In vivo analysis of human (h)ESC-derived motoneurons indeveloping chick spinal cord. (A): Schematic illustration of transplan-tation procedure at stages HH16–17 in chick spinal cord. (B): NativeeGFP fluorescence allowed detection of grafted hESC-derived cells inchick spinal cord (stage HH34). Area depicted corresponds to boxedarea in inset. Inset provides an overview of the whole trunk region of theembryo. (C–E): Transverse sections at stage HH26 chick spinal cord.Human cells located in the dorsal spinal cord were identified by nativeeGFP signal and expression of human specific anti-neural cell adhesionmolecule. Motoneuron identity was confirmed by Nkx6.1 (D) and HB9(E) immunohistochemistry. (F): Transverse section of half spinal cordat stage HH34 showed hESC-derived motoneurons migrating towardventral spinal cord. (G–H): Transverse sections at stage HH38 demon-strate human axonal projections out of the spinal cord. (G): Upper andlower-left panels: hNCAM� fiber projecting out of the spinal cord viaventral root. Lower-right panel: human fibers reaching host skeletalmuscle target. Inset: identity of target area revealed by expression ofskeletal marker MF20 in adjacent section. (H): Long-distance axonalgrowth of hESC-derived motoneurons in vivo. Left panel: composite oftransverse embryo section; region marked with (�) corresponds to themuscle target area depicted in (G), lower-right panel. Right panel:long-distance projection of human eGFP/hNCAM� fiber. The locationof the fiber corresponds to the boxed area in the left panel. Blue colorin (C, F, G, H) represents 4,6-diamidino-2-phenylindole staining. Scalebar in (C) corresponds to 200 �m for (C); 125 �m for (F); 100 �m for(G) upper and lower-left panels, (H) right panel; 50 �m for (D, E) and(G) lower-right panel; 3 mm for (H) left panel. Abbreviations: eGFP,enhanced green fluorescent protein; hNCAM, human neural cell adhe-sion molecule.
1936 Human Embryonic Stem Cell-Derived Motoneurons

tion). The decrease in HB9 and Nkx6.1 expression wasparalleled by an increase in the number of ChAT� neuronsfrom 4% of GFP� cells within the first week after transplan-tation to 14% at 6 weeks. The temporary decrease in thepercentage of ChAT� cells compared with in vitro numberssuggest that ChAT� cells may preferentially die or tempo-rarily lose ChAT expression following grafting. Loss of HB9expression in vivo followed a time course similar to thatobserved in hESC-derived motoneuron cultures in vitro. Fur-thermore, motoneurons in the developing chick spinal cordshowed a similar loss in HB9 over time, and no HB9 expres-sion was observed in endogenous motoneurons of the adultrat spinal cord. These data suggest that the switch from HB9to ChAT expression corresponds to physiological motoneu-ron maturation rather than loss of in vivo phenotype. At 6weeks after transplantation, the graft core was composed of ahigh percentage of eGFP/ChAT� cells (Fig. 5D). We alsoobserved single eGFP/ChAT double-labeled neurons locatedventrally outside of the graft core with characteristic mo-toneuron morphologies (Fig. 5E). Current analysis was lim-ited to the spinal cord and did not address fiber outgrowthoutside the CNS. However, hNCAM staining revealed exten-sive fiber outgrowth and cell migration toward the ventralsurface of the spinal cord (Fig. 5F). This suggests that hESC-derived motoneurons might be able to project toward theventral root in the adult spinal cord, similar to our datapresented in the embryonic chick spinal cord, albeit at adifferent time scale. We did not observe signs of tumorige-nicity in any of the animals grafted. However, studies using
larger groups of animals and assessing long-term survivalbeyond 6 weeks will be required to fully assess in vivo safetyof the cells.
DISCUSSION
This study demonstrates that hESC progeny is developmen-tally competent to respond to morphogens that determine APand DV axis specification during early vertebrate develop-ment. Our protocols yield neural rosettes containing largenumbers of neural precursors coexpressing Pax6 and Sox1and maintained in vitro beyond the stage perviously definedcritical for motoneuron specification. We previously showedthat hESC-derived neural progeny at day 28 (expressing Pax6and Sox1) are capable of midbrain dopaminergic differenti-ation [11]. The current study extends these findings to morecaudal cell types, including motoneurons. Patterning of day28 rosettes toward caudal fates is quite remarkable, given themajority of rosette cells expressing BF1, a marker tradition-ally associated with forebrain commitment. Our data alsosuggest that neural rosettes adopt anterior CNS markers bydefault. An anterior neural default model of CNS develop-ment has been postulated previously based on classic exper-iments in xenopus embryos. These studies showed that ante-rior CNS fates are first established and subsequently undergoa caudal transformation in response to secreted signals [26].Morphogen-induced patterning of neural rosettes provides apowerful strategy to systematically generate neural cell typesof distinct AP identity.
The protocols presented here offer a simple and efficientstrategy to generate human motoneurons. Based on veryconservative calculations, a single hESC plated at day 0 onMS5 for neural induction yields approximately 100 HB9�motoneurons at day 50 of differentiation. These numberssuggest that therapeutically relevant numbers of motoneuronscan be readily achieved. Although spinal motoneurons arederived from a single ventral pMN domain [27, 28], theyfurther acquire many different subtype identities based onpositional identity, axonal projections, and gene expression.For translational applications of ESC-derived motoneurons, itwill be essential to develop motoneuron subtype specificprotocols that match the diseased population. There is evi-dence that most motoneurons derived from mouse ESCsusing the RA/SHH protocol correspond to cervical or bra-chial level motoneurons based on Hoxc5 and Hoxc6 expres-sion [4]. Similarly, many hESC-derived motoneuron in ourprotocol exhibit characteristics of brachial motoneurons.However, there is a slight caudal shift as compared withmouse ESC-derived motoneuron progeny toward HoxC6 andHoxC8 expression.
It is known that RA and FGF have antagonistic rolesduring development of the early chick spinal cord. FGFinhibits differentiation in the caudal stem zone, whereas RAprovided rostrally by cells in the somitic mesoderm is re-quired for neuronal differentiation and patterning of the ven-tral spinal cord. Mutual inhibition between the RA and FGFpathways not only controls neural differentiation but alsounderlies the progressive assignment of rostrocaudal identityby regulating Hox gene availability and activation [29, 30].In addition, recent work has shown that combinatorial actionsof Wnt, RA, and FGF signals specify progenitor cell identi-ties and motoneuron subtypes in the developing spinal cord[31]. Our current motoneuron differentiation protocols arebased on exposure to high concentrations of RA, which mayrestrict AP identity. By decreasing the levels of RA and
Figure 5. In vivo assessment of transplanted human (h)ESC-derivedmotoneurons in the adult rat spinal cord. (A–C): Coronal sections ofspinal cord within the first 2 weeks after transplantation. Human graftwas identified in orthotopic location in the ventral spinal cord by theexpression of eGFP and human nuclear antigen. (B): Higher magnifi-cation image demonstrating colocalization of hNA and GFP�. (C):Expression of the motoneuron markers HB9 and Nkx6.1 in hESC-derived motoneuron grafts after transplantation. (D–F): Coronal sec-tions of grafted adult rat spinal cord 6 weeks after transplantation. (D):hESC-derived eGFP� cells expressed ChAT, a more mature motoneu-ron marker. (E): Reconstruction of confocal z-axis stack showing colo-calization of eGFP and ChAT in a neuron located ventrally outside thegraft core. (F): hNCAM� fibers projecting toward ventral surface. Insome instances, hNCAM� cell bodies were aligned along human fibers(graft core is indicated by dashed line). Blue color in all figures repre-sents 4,6-diamidino-2-phenylindole staining. Scale bar in (A) corre-sponds to 200 �m for (A), 150 �m for (F), 100 �m for (C) right panel;60 �m for (C) left panel, 50 �m for (B, E), and 35 �m for (D).Abbreviations: ChAT, choline acetyl transferase; eGFP, enhanced greenfluorescent protein; hNA, human nuclear antigen; hNCAM, humanneural cell adhesion molecule.
1937Lee, Shamy, Elkabetz et al.
www.StemCells.com

titrating FGF levels, hESC-derived rosette progeny may bedirected toward more caudal motoneuron fates. Pretreatmentwith Wnt agonistic molecules may be another strategy toaffect AP identity in motoneuron progeny.
We also successfully demonstrated that the transplantationof hESC-derived motoneurons in the developing chick embryoresulted in motoneuron survival and axonal outgrowth towardperipheral targets, despite the cross-species nature of the grafts,with considerable differences in developmental timing. Futurestudies will require detailed in vivo physiological analyses andretrograde tracing from specific muscle groups. However, suchstudies will be technically challenging due to the difficulties infollowing human axons over extended distances from the ven-tral horn to the peripheral muscle target within a single slice.Transplantation of specific motoneuron subtypes into the chickspinal cord will allow us to probe graft host interactions and todefine the required subtypes necessary for restoration of motorfunction for a given spinal cord segment.
Finally, we show that hESC-derived motoneurons surviveand maintain phenotype in the adult rat spinal cord for at least6 weeks after transplantation. To our knowledge, this is thefirst study demonstrating in vivo survival of hESC-derivedmotoneurons, a prerequisite for future applications in pre-clinical models of motoneuron disease. Although our resultsprovide an important first step in demonstrating functionalengraftment, a number of key studies are required for futurepreclinical development. A key issue relates to the questionof whether hESC-derived motoneurons can survive in animalmodels of motoneuron disease such as the SOD1G93A rat[32]. If robust survival can be achieved, it will be critical totest whether hESC-derived motoneurons are sensitive to en-dogenous inhibitors blocking axonal outgrowth in the adult
CNS at levels similar to those observed with mouse ESC-derived motoneurons [33, 34]. Additional steps will includequestions related to the presence of guidance cues in the adultspinal cord, the capacity for long-distance axonal growth, andthe ability to innervate appropriate peripheral targets. Finally,the relationship of target innervation and changes in behav-ioral parameters needs to be explored. Although the road tothe clinical application of hESC-derived motoneurons re-mains extremely challenging, the ability to generate unlim-ited numbers of motoneuron progeny and the capacity for invivo survival and integration in the developing and adultspinal cord are important first steps on this journey.
ACKNOWLEDGMENTS
We thank S. Heller, Q.R. Lu, F. Vaccarino, and T. Jessell forNkx6.1, Olig2, Otx2, and HoxC6 antibodies, H. Wichterle andJ. Timmer for help in setting up chick embryo injections, and S.Noggle and A. Brivanlou for the RUES1-eGFP cell line. Wealso thank T. Bajwa and S. Clairmont for excellent technicalassistance and M. Tomishima and S. Desbordes for criticalreview of the manuscript. This work was supported throughgrants from the ALS Association, Project ALS, the Michael W.McCarthy Foundation, and the Kinetics Foundation.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Cleveland DW, Rothstein JD. From Charcot to Lou Gehrig: Deci-phering selective motor neuron death in ALS. Nat Rev Neurosci 2001;2:806 – 819.
2 Lefebvre S, Burglen L, Reboullet S et al. Identification and characterizationof a spinal muscular atrophy-determining gene. Cell 1995;80:155–165.
3 Shirasaki R, Pfaff SL. Transcriptional codes and the control of neuronalidentity. Annu Rev Neurosci 2002;25:251–281.
4 Wichterle H, Lieberam I, Porter JA et al. Directed differentiation ofembryonic stem cells into motor neurons. Cell 2002;110:385–397.
5 Barberi T, Klivenyi P, Calingasan NY et al. Neural subtype specificationof fertilization and nuclear transfer embryonic stem cells and applicationin parkinsonian mice. Nat Biotechnol 2003;21:1200–1207.
6 Li XJ, Du ZW, Zarnowska ED et al. Specification of motoneurons fromhuman embryonic stem cells. Nat Biotechnol 2005;23:215–221.
7 Singh Roy N, Nakano T, Xuing L et al. Enhancer-specified GFP-basedFACS purification of human spinal motor neurons from embryonic stemcells. Exp Neurol 2005;196:224–234.
8 Zhang SC, Wernig M, Duncan ID et al. In vitro differentiation oftransplantable neural precursors from human embryonic stem cells. NatBiotechnol 2001;19:1129–1133.
9 Cibelli JB, Grant KA, Chapman KB et al. Parthenogenetic stem cells innonhuman primates. Science 2002;295:819.
10 Johe KK, Hazel TG, Muller T et al. Single factors direct the differenti-ation of stem cells from the fetal and adult central nervous system. GenesDev 1996;10:3129–3140.
11 Perrier AL, Tabar V, Barberi T et al. Derivation of midbrain dopamineneurons from human embryonic stem cells. Proc Natl Acad Sci U S A2004;101:12543–12548.
12 Greaney MD, Marshall DL, Bailey BA et al. Improved method for theroutine analysis of acetylcholine release in vivo: Quantitation in thepresence and absence of esterase inhibitor. J Chromatogr 1993;622:125–135.
13 Kakinohana O, Cizkova D, Tomori Z et al. Region-specific cell graftinginto cervical and lumbar spinal cord in rat: A qualitative and quantitativestereological study. Exp Neurol 2004;190:122–132.
14 Tabar V, Panagiotakos G, Greenberg ED et al. Migration and differen-tiation of neural precursors derived from human embryonic stem cells inthe rat brain. Nat Biotechnol 2005;23:601–606.
15 Kawasaki H, Mizuseki K, Nishikawa S et al. Induction of midbraindopaminergic neurons from ES cells by stromal cell-derived inducingactivity. Neuron 2000;28:31–40.
16 Kawasaki H, Suemori H, Mizuseki K et al. Generation of dopaminergicneurons and pigmented epithelia from primate ES cells by stromal cell-derived inducing activity. Proc Natl Acad Sci U S A 2002;99:1580–1585.
17 Walther C, Gruss P. Pax-6, a murine paired box gene, is expressed in thedeveloping CNS. Development 1991;113:1435–1449.
18 Li HS, Yang JM, Jacobson RD et al. Pax-6 is first expressed in a regionof ectoderm anterior to the early neural plate: Implications for stepwisedetermination of the lens. Dev Biol 1994;162:181–194.
19 Pevny LH, Sockanathan S, Placzek M et al. A role for SOX1 in neuraldetermination. Development 1998;125:1967–1978.
20 Tao W, Lai E. Telencephalon-restricted expression of BF-1, a newmember of the HNF-3/fork head gene family, in the developing rat brain.Neuron 1992;8:957–966.
21 Soundararajan P, Miles GB, Rubin LL et al. Motoneurons derived fromembryonic stem cells express transcription factors and develop pheno-types characteristic of medial motor column neurons. J Neurosci 2006;26:3256–3268.
22 James D, Noggle SA, Swigut T et al. Contribution of human embryonicstem cells to mouse blastocysts. Dev Biol 2006;295:90–102.
23 Vrana KE, Hipp JD, Goss AM et al. Nonhuman primate parthenogeneticstem cells. Proc Natl Acad Sci U S A 2003;100(suppl 1):11911–11916.
24 Dasen JS, Liu JP, Jessell TM. Motor neuron columnar fate imposed bysequential phases of Hox-c activity. Nature 2003;425:926–933.
25 Dasen JS, Tice BC, Brenner-Morton S et al. A Hox regulatory networkestablishes motor neuron pool identity and target-muscle connectivity.Cell 2005;123:477–491.
26 Nieuwkoop PD. The neural induction process; its morphogenetic aspects.Int J Dev Biol 1999;43:615–623.
27 Briscoe J, Pierani A, Jessell TM et al. A homeodomain protein codespecifies progenitor cell identity and neuronal fate in the ventral neuraltube. Cell 2000;101:435–445.
28 Ericson J, Briscoe J, Rashbass P et al. Graded sonic hedgehog signalingand the specification of cell fate in the ventral neural tube. Cold SpringHarb Symp Quant Biol 1997;62:451–466.
1938 Human Embryonic Stem Cell-Derived Motoneurons

29 Diez del Corral, Olivera-Martinez I, Goriely A et al. Opposing FGF andretinoid pathways control ventral neural pattern, neuronal differentiation,and segmentation during body axis extension. Neuron 2003;40:65–79.
30 Diez del Corral R, Storey KG. Opposing FGF and retinoid pathways: Asignalling switch that controls differentiation and patterning onset in theextending vertebrate body axis. Bioessays 2004;26:857–869.
31 Nordstrom U, Maier E, Jessell TM et al. An early role for Wnt signalingin specifying neural patterns of Cdx and Hox gene expression and motorneuron subtype identity. PLoS Biol 2006;4:e252.
32 Nagai M, Aoki M, Miyoshi I et al. Rats expressing human cytosoliccopper-zinc superoxide dismutase transgenes with amyotrophic lateralsclerosis: Associated mutations develop motor neuron disease. J Neuro-sci 2001;21:9246–9254.
33 Deshpande DM, Kim YS, Martinez T et al. Recovery from paralysis inadult rats using embryonic stem cells. Ann Neurol 2006;60:32–44.
34 Harper JM, Krishnan C, Darman JS et al. Axonal growth of embryonicstem cell-derived motoneurons in vitro and in motoneuron-injured adultrats. Proc Natl Acad Sci U S A 2004;101:7123–7128.
See www.StemCells.com for supplemental material available online.
1939Lee, Shamy, Elkabetz et al.
www.StemCells.com





























