Embed Size (px)
Citation preview

DTT
RAa
sTb
dT
Atag1d
1
*EAABB1csqqbsteFGGGlsthgsL4dNiPPsStnsSbTTfz
Neuroscience 155 (2008) 1174–1194
0d
ISTINCT MECHANISMS OF 1-METHYL-4-PHENYL-1,2,3,6-ETRAHYDROPYRIMIDINE RESISTANCE REVEALED BY
RANSCRIPTOME MAPPING IN MOUSE STRIATUMamaecnpnrapeAmliaBpnsmredtMtmgir
KB
Pt((mapSar
t(alf
. PATTARINI,a1 Y. RONG,a1 C. QUb
ND J. I. MORGANa*
Department of Developmental Neurobiology, St. Jude Children’s Re-earch Hospital, 262 Danny Thomas Place, Danny Thomas Researchower, Room D2025E, Mail Stop 323, Memphis, TN 38105-3678, USA
Hartwell Center for Bioinformatics and Biotechnology, St. Jude Chil-ren’s Research Hospital, Danny Thomas Research Tower, Memphis,N 38105-3678, USA
bstract—The etiology of idiopathic Parkinson’s disease ishought to involve interplay between environmental factorsnd predisposing genetic traits, although the identification ofenetic risk factors remain elusive. The neurotoxicant,-methyl-4-phenyl-1,2,3,6-tetrahydropyrimidine (MPTP) pro-uces parkinsonian-like symptoms and pathology in mice
Both authors contributed equally to this article.Corresponding author. Tel: �1-901-595-2256; fax: �1-901-595-3143.-mail address: [email protected] (J. I. Morgan).bbreviations: Agxt2l1, alanine-glyoxylate aminotransferase 2-like 1;if1, allograft inflammatory factor 1; Apod, apolipoprotein D; Bach1,TB and CNC homology 1; Bax, BCL2-associated X protein; Btg3,-cell translocation gene 3; Cdkn1a, cyclin-dependent kinase inhibitorA; c-Fos, FBJ osteosarcoma oncogene; c-Jun, Jun oncogene; Comt,atechol-O-methyl transferase; C1qa, complement component 1, qubcomponent, alpha polypeptide; C1qb, complement component 1,subcomponent, beta polypeptide; C1qc, complement component 1,subcomponent, C chain; C4b, complement component 4B (Childo
lood group); DA, dopaminergic; Ddit4, DNA-damage-inducible tran-cript 4; Ddit4l, DNA-damage-inducible transcript 4-like; Edg3, endo-helial differentiation, sphingolipid G-protein-coupled receptor, 3; Egr3,arly growth response 3; Egr4, early growth response 4; Fkbp5,K506 binding protein 5; Fosb, FBJ osteosarcoma oncogene B;add45b, growth arrest and DNA-damage-inducible 45 betaadd45g, growth arrest and DNA-damage-inducible 45 gammaapdh, glyceraldehyde-3-phosphate dehydrogenase; Gfap, glial fibril-
ary acidic protein; Gpr37, G protein-coupled receptor 37; GSEA, geneet enrichment analysis; Hbegf, heparin-binding EGF-like growth fac-or; HC, Hartwell Center for Bioinformatics & Biotechnology; Hmox1,emeoxygenase-1; Hspb6, heat shock protein B6; Junb, Jun-B onco-ene; Klf9, Kruppel-like factor 9; Lgals1, lectin, galactose binding,oluble 1; Lgals3, lectin, galactose binding, soluble 3; Lgals3bp,gals3 binding protein; Ly86, lymphocyte antigen 86; MPP�, 1-methyl--phenylpyridinium; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-ine; Msr2, macrophage scavenger receptor 2; Mt2, metallothionein 2;fkbia, nuclear factor of kappa light chain gene enhancer in B-cells
nhibitor, alpha; Osmr, oncostatin M receptor; Pax8, paired box gene 8;D, Parkinson’s disease; Pdlim4, PDZ and LIM domain 4; Pink1,TEN induced putative kinase 1; qRT-PCR, quantitative reverse tran-cription polymerase chain reaction; Rxrg, retinoid X receptor gamma;gk, serum/glucocorticoid regulated kinase 1; Sgk3, serum/glucocor-
icoid regulated kinase 3; Snca, synuclein, alpha; SNpc, substantiaigra pars compacta; Stat1, signal transducer and activator of tran-cription 1; Stat3, signal transducer and activator of transcription 3;100a6, S100 calcium binding protein A6; S100a10, S100 calciuminding protein A10; S100a13, S100 calcium binding protein A13;nfrsf12a, tumor necrosis factor receptor superfamily, member 12a;sc22d1, TSC22 domain family, member 1; Tsc22d3, TSC22 domain
aamily, member 3; Ucp2, uncoupling protein 2; Vim, vimentin; Zbtb16,inc finger and BTB domain containing 16.
306-4522/08 © 2008 IBRO. Published by Elsevier Ltd. All rights reserved.oi:10.1016/j.neuroscience.2008.06.064
1174
nd humans. As sensitivity to MPTP is genetically deter-ined in mice this provides an opportunity to identify genes
nd biological mechanisms that modify the response to anxogenous agent that produces a Parkinson’s disease–likeondition. MPTP primarily targets dopaminergic nerve termi-als in the striatum and elicits changes in striatal gene ex-ression. Therefore, we used Affymetrix® and qRT-PCR tech-ology to characterize temporal mRNA changes in striatum inesponse to MPTP in genetically MPTP-sensitive, C57BL/6J,nd MPTP-resistant Swiss Webster and BCL2-associated Xrotein (Bax)�/� mice. We identified three phases of mRNAxpression changes composed of largely distinct gene sets.n early response (5 h) occurred in all strains of mice andultiple brain regions. In contrast, intermediate (24 h) and
ate (72 h) phases were striatum specific and much reducedn Swiss Webster, indicating these genes contribute and/orre responsive to MPTP-induced pathology. However,ax�/� mice have robust intermediate responses. We pro-ose a model in which the acute entry of MPP� into dopami-ergic nerve terminals damages them but is insufficient pere to kill the neurons. Rather, we suggest that the compro-ised nerve terminals elicit longer lasting transcriptional
esponses in surrounding cells involving production of mol-cules that feedback on the terminals to cause additionalamage that results in cell death. In Swiss Webster, resis-ance lies upstream in the cascade of events triggered byPTP and uncouples the acute events elicited by MPTP from
he damaging secondary responses. In contrast, in Bax�/�ice resistance lies downstream in the cascade and sug-ests enhanced tolerance to the secondary insult rather than
ts attenuation. © 2008 IBRO. Published by Elsevier Ltd. Allights reserved.
ey words: C57BL/6J mice, Swiss Webster mice, B6.129X1-axtm1Sjk/J mice, Affymetrix array, Parkinson’s disease.
arkinson’s disease (PD) is a progressive neurodegenera-ive disorder characterized by the loss of dopaminergicDA) neurons in the substantia nigra pars compactaSNpc). Although the etiology of idiopathic PD is unclear, itay originate from interplay between environmentalgents (Landrigan et al., 2005; Brown et al., 2006) andredisposing genetic traits (Huang et al., 2004; Benmoyal-egal and Soreq, 2006; Farrer, 2006; Wood-Kaczmar etl., 2006). However, the identification of specific geneticisk factors for idiopathic PD remains elusive.
The most widely studied experimental system of PD ishe murine 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridineMPTP) model (Dauer and Przedborski, 2003; Smeynend Jackson-Lewis, 2005). Although it does not recapitu-
ate PD in its entirety, it reproduces several of its cardinaleatures including loss of DA neurons in the SNpc (Dauer
nd Przedborski, 2003; Przedborski and Vila, 2003;
Ssaag
othTtdmasv1
i(iaAsa1h2H2pKtc
ip
A
Fpsi(mS(wd(sdwibCNtmm
R
T(BafpficeRec
P
Tcwt(ffiaTGtcCkr(uSDCtpmbklalfMhsas(vcb
A
TA3ci(c
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1175
meyne and Jackson-Lewis, 2005). Moreover, as MPTPensitivity in mice is genetically determined (Sundstrom etl., 1987; German et al., 1996; Hamre et al., 1999; Vila etl., 2001), it provides an opportunity to identify potentialenetic risk factors for PD.
Several lines of evidence suggest that the primary sitef injury in both PD and the MPTP model is the SNpc nerve
erminals in the striatum (Bradbury et al., 1986; Herken-am et al., 1991; Nurmi et al., 2001; Rinne et al., 2001).hus the initial event is damage of the synaptic terminals in
he striatum, followed by retrograde degeneration and celleath (Bradbury et al., 1986; Eberling et al., 1997). Thisechanism is particularly relevant for MPTP toxicity as itsctive metabolite, 1-methyl-4-phenylpyridinium (MPP�)electively accumulates in DA nerve terminals in striatumia uptake through the dopamine transporter (Chiba et al.,985; Javitch et al., 1985; Gainetdinov et al., 1997).
Shortly after its administration MPTP elicits the dump-ng of dopamine from SNpc nerve endings in striatumJackson-Lewis et al., 1995). This event coincides with thenduction of several immediate-early genes (Duchemin etl., 1992; Smith et al., 1997; Perez-Otano et al., 1998;gani et al., 2000; Chen et al., 2001). By 24 h, depletion oftriatal tyrosine hydroxylase (Kuhn et al., 2003; Sriram etl., 2004) and damage to DA synapses (Linder et al.,995), coincide with induction of several genes, includingemeoxygenase-1 (Hmox1) (Fernandez-Gonzalez et al.,000) and cytokines/chemokines (Nagatsu et al., 2000;ebert et al., 2003; Sriram et al., 2004, 2006; Shen et al.,005; Pattarini et al., 2007). Subsequent neuronal death isrogressive between 12 h (Jackson-Lewis et al., 1995;uhn et al., 2003) and 7 days (Boyd et al., 2007). Based on
his evidence, these early changes in gene expressionould contribute to neuronal demise.
We hypothesized that sensitivity to MPTP may residen genes expressed in striatum and their identification mayoint to genetic risk factors for PD.
EXPERIMENTAL PROCEDURES
nimals and experiments
emale C57BL/6J and B6.129X1-Baxtm1Sjk/J (BCL2-associated Xrotein (Bax)�/�) mice of both genders were purchased from Jack-on Laboratories (Bar Harbor, ME, USA). Bax�/� mice were bredn-house and intercrossed to obtain homozygous knockout animalsBax�/�) and wild-type littermates (Bax�/�). The genotype for Baxice was performed by Transnetyx (Cordova, TN, USA). FemaleWR mice were purchased from Jackson Laboratories and Harlan
Indianapolis, IN, USA). Animals were housed in micro-isolator unitsith a 12-h light/dark schedule and constant temperature. Furtheretails of the Experimental Procedures are provided in Pattarini et al.2007). MPTP was administered by i.p. injections at the dosage andchedule specified in the Results section. Animals were killed atifferent time points after the first dose of MPTP and brain regionsere dissected and immediately frozen on dry ice to preserve RNA
ntegrity. Samples were stored at �80 °C. All studies were approvedy the St. Jude Children’s Research Hospital Animal Care and Useommittee (ACUC) and were conducted in accordance with theational Institutes of Health Guide for the Care and Use of Labora-
ory Animals (NIH Publication No. 80–23, revised 1996). Efforts wereade to minimize the number of animals involved in each experi-
ent and their suffering. uNA isolation
otal RNA was extracted with TRIzol® Reagent from InvitrogenCarlsbad, CA, USA) according to manufacturer instructions.riefly, 1 ml TRIzol® was added to frozen samples and immedi-tely homogenized. Samples were mixed with 200 �l of chloro-orm and centrifuged for 15 min at 12,000�g at 4 °C. The aqueoushase was transferred to a new vial, mixed with 50 �g of glycogen
rom Roche Applied Science (Penzberg, Germany) and 500 �l ofsopropanol and centrifuged (10 min at 12,000�g, 4 °C) to pre-ipitate total RNA. The pellet was washed with ice cold 70%thanol and air dried for 10 min. Total RNA was resuspended inNase-free water and it was checked for integrity by agarose gellectrophoresis. Samples that appeared degraded were dis-arded.
reparation of samples for microarray analysis
echnical procedures for microarray analysis, including qualityontrol of RNA, labeling, hybridization and scanning of the arraysere performed by the Hartwell Center for Bioinformatics & Bio-
echnology (HC) at St. Jude Children’s Research HospitalSJCRH) according to standard operating procedures for Af-ymetrix protocols (GeneChip® Expression Analysis manual, Af-ymetrix, Santa Clara, CA, USA). Prior to their use, RNA samplentegrity was analyzed with the 2100 Bioanalyzer Laboratory-on--chip system (Agilent Technologies, Santa Clara, CA, USA).otal RNA samples that were not degraded were labeled using theene Chip IVT Labeling Kit (Affymetrix) according to manufac-
urer instructions. Briefly, as a quality control of the labeling pro-ess, samples were first spiked with the GeneChip® Poly-A RNAontrol Kit (Affymetrix) that contains mRNA (not present in eu-aryotic cells) for the following B. subtilis genes: lys (1:100,000atio of copy number), phe (1:50,000), thr (1:25,000) and dap1:7500). Samples were then used to prepare the 1st strand cDNAsing the One-Cycle cDNA Synthesis Kit (Affymetrix) containinguperScript II followed by the 2nd strand cDNA synthesis with T4NA polymerase. cDNA was cleaned using cDNA Cleanup Spinolumn (Affymetrix), and biotin-labeled cRNA was prepared using
he Gene Chip IVT Labeling Kit (Affymetrix). Labeled cRNA wasurified with Cleanup Spin Column (Affymetrix), quantified, frag-ented and spiked with biotin-labeled cRNA for bioB (1.5 pM),ioC (5 pM), bioD (25 pM) and Crex (100 pM) (GeneChip® Eu-aryotic Hybridization Control Kit, Affymetrix). This procedure al-owed us to assess both the linearity of detection and the lowestccurately detectable concentration (1.5 pmol). Samples were
oaded onto the Affymetrix® Mouse Genome 430 2.0 Arrays (Af-ymetrix) previously washed with 1� hybridization buffer (100 mMES, 1 M Na�, 20 mM EDTA, 0.01% Tween-20 pH 6.6) andybridized overnight (16 h) at 37 °C. Arrays were washed andtained with streptavidin conjugated to phycoerthyrin, using theutomated GeneChip® Fluidics Station 400 (Affymetrix) andcanned to produce an image file with the GeneArray™ scannerAffymetrix). Total RNA from each animal was loaded onto indi-idual Affymetrix microarray chips. Experimental reproducibilityan be estimated by comparing columns within a figure as well asetween corresponding columns in Figs. 1, 3, 4, 6, and 8.
nalysis of microarrays
he microarrays used in this study (Mouse Genome 430 2.0rrays, Affymetrix) contain 45,101 probe sets, representing9,000 transcripts and variants, and they are currently the mostomprehensive genechip array available for the mouse. Scannedmages were analyzed with the Gene Chip Operating SoftwareGCOSv1.2, Affymetrix). Assessment of probe set present/absentalls was made using the Single Array Analysis method in GCOS
sing the statistical algorithm with default analysis parameters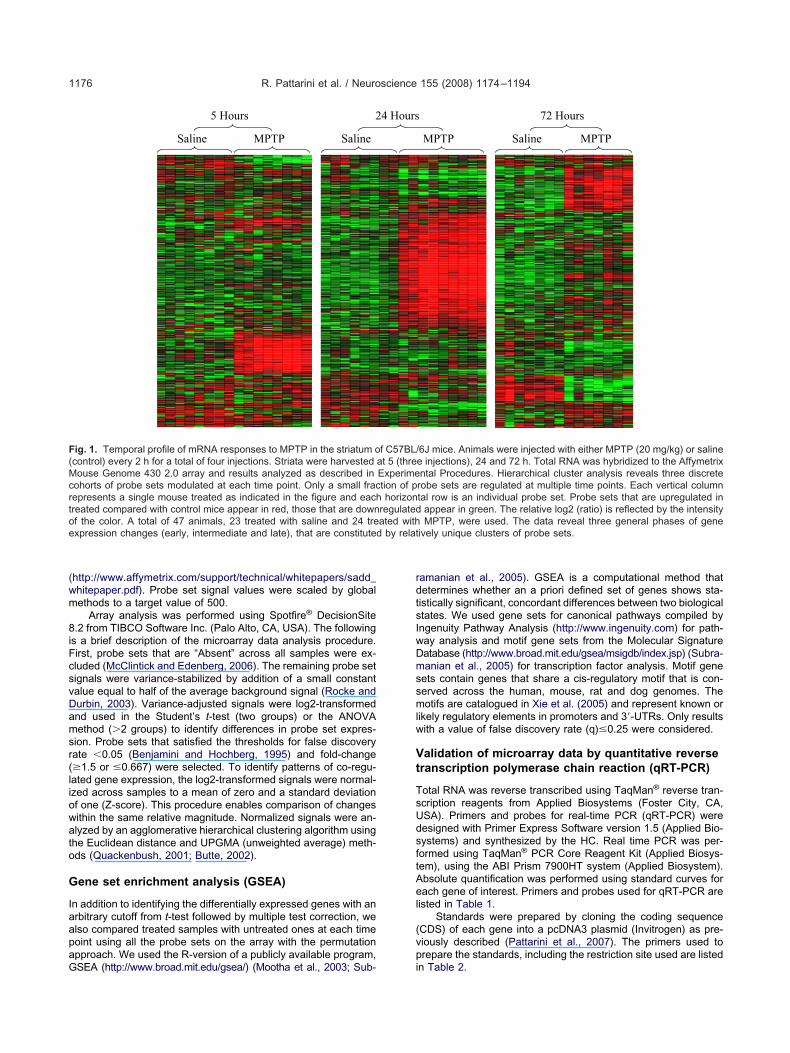
(wm
8iFcsvDamsr(liowato
G
IaapaG
rdtsIwDmssmlw
Vt
TsUdsftAel
(vp
F(Mcrtoe
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941176
http://www.affymetrix.com/support/technical/whitepapers/sadd_hitepaper.pdf). Probe set signal values were scaled by globalethods to a target value of 500.
Array analysis was performed using Spotfire® DecisionSite.2 from TIBCO Software Inc. (Palo Alto, CA, USA). The following
s a brief description of the microarray data analysis procedure.irst, probe sets that are “Absent” across all samples were ex-luded (McClintick and Edenberg, 2006). The remaining probe setignals were variance-stabilized by addition of a small constantalue equal to half of the average background signal (Rocke andurbin, 2003). Variance-adjusted signals were log2-transformednd used in the Student’s t-test (two groups) or the ANOVAethod (�2 groups) to identify differences in probe set expres-
ion. Probe sets that satisfied the thresholds for false discoveryate �0.05 (Benjamini and Hochberg, 1995) and fold-change�1.5 or �0.667) were selected. To identify patterns of co-regu-ated gene expression, the log2-transformed signals were normal-zed across samples to a mean of zero and a standard deviationf one (Z-score). This procedure enables comparison of changesithin the same relative magnitude. Normalized signals were an-lyzed by an agglomerative hierarchical clustering algorithm using
he Euclidean distance and UPGMA (unweighted average) meth-ds (Quackenbush, 2001; Butte, 2002).
ene set enrichment analysis (GSEA)
n addition to identifying the differentially expressed genes with anrbitrary cutoff from t-test followed by multiple test correction, welso compared treated samples with untreated ones at each timeoint using all the probe sets on the array with the permutationpproach. We used the R-version of a publicly available program,
5 Hours 2
SalineSaline MPTP
ig. 1. Temporal profile of mRNA responses to MPTP in the striatum ocontrol) every 2 h for a total of four injections. Striata were harvestedouse Genome 430 2.0 array and results analyzed as described in
ohorts of probe sets modulated at each time point. Only a small fracepresents a single mouse treated as indicated in the figure and eachreated compared with control mice appear in red, those that are downf the color. A total of 47 animals, 23 treated with saline and 24 trexpression changes (early, intermediate and late), that are constituted
SEA (http://www.broad.mit.edu/gsea/) (Mootha et al., 2003; Sub- i
amanian et al., 2005). GSEA is a computational method thatetermines whether an a priori defined set of genes shows sta-istically significant, concordant differences between two biologicaltates. We used gene sets for canonical pathways compiled byngenuity Pathway Analysis (http://www.ingenuity.com) for path-ay analysis and motif gene sets from the Molecular Signatureatabase (http://www.broad.mit.edu/gsea/msigdb/index.jsp) (Subra-anian et al., 2005) for transcription factor analysis. Motif gene
ets contain genes that share a cis-regulatory motif that is con-erved across the human, mouse, rat and dog genomes. Theotifs are catalogued in Xie et al. (2005) and represent known or
ikely regulatory elements in promoters and 3=-UTRs. Only resultsith a value of false discovery rate (q)�0.25 were considered.
alidation of microarray data by quantitative reverseranscription polymerase chain reaction (qRT-PCR)
otal RNA was reverse transcribed using TaqMan® reverse tran-cription reagents from Applied Biosystems (Foster City, CA,SA). Primers and probes for real-time PCR (qRT-PCR) wereesigned with Primer Express Software version 1.5 (Applied Bio-ystems) and synthesized by the HC. Real time PCR was per-ormed using TaqMan® PCR Core Reagent Kit (Applied Biosys-em), using the ABI Prism 7900HT system (Applied Biosystem).bsolute quantification was performed using standard curves forach gene of interest. Primers and probes used for qRT-PCR are
isted in Table 1.Standards were prepared by cloning the coding sequence
CDS) of each gene into a pcDNA3 plasmid (Invitrogen) as pre-iously described (Pattarini et al., 2007). The primers used torepare the standards, including the restriction site used are listed
72 Hours
MPTP Saline MPTP
6J mice. Animals were injected with either MPTP (20 mg/kg) or salinee injections), 24 and 72 h. Total RNA was hybridized to the Affymetrixntal Procedures. Hierarchical cluster analysis reveals three discreterobe sets are regulated at multiple time points. Each vertical columntal row is an individual probe set. Probe sets that are upregulated in
appear in green. The relative log2 (ratio) is reflected by the intensityMPTP, were used. The data reveal three general phases of gene
ively unique clusters of probe sets.
4 Hours
f C57BL/at 5 (threExperimetion of phorizon
regulatedated with
by relat
n Table 2.
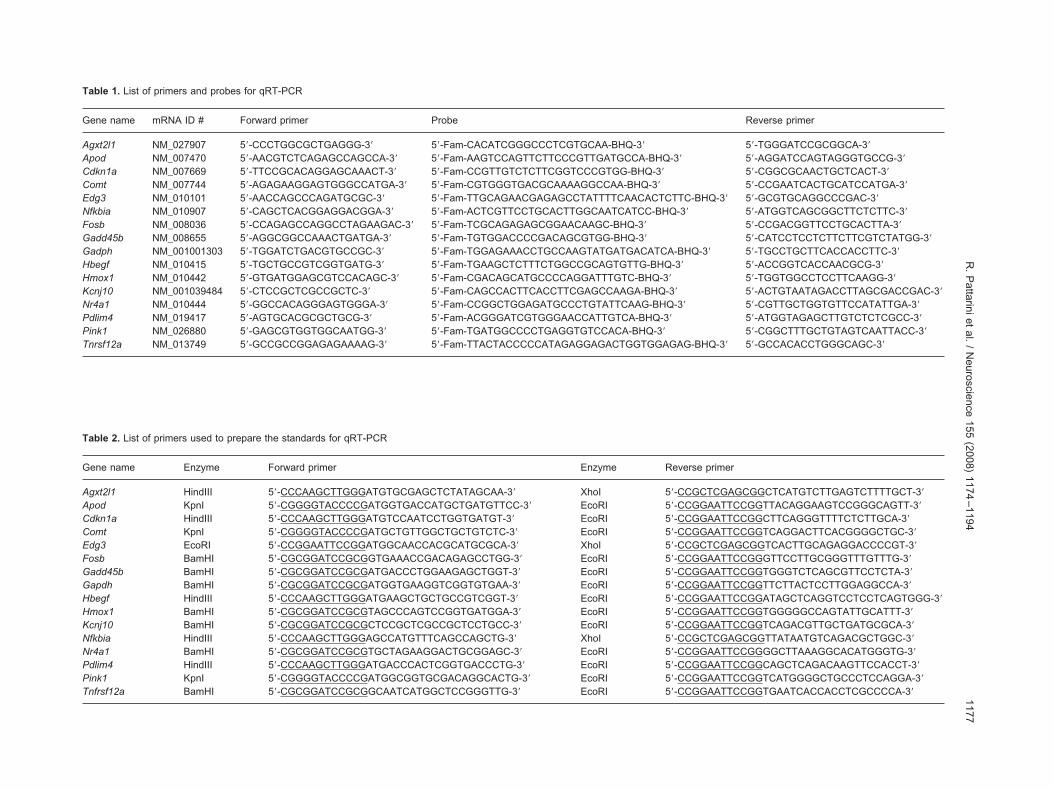
Table 1. List of primers and probes for qRT-PCR
Gene name mRNA ID # Forward primer Probe Reverse primer
Agxt2l1 NM_027907 5=-CCCTGGCGCTGAGGG-3= 5=-Fam-CACATCGGGCCCTCGTGCAA-BHQ-3= 5=-TGGGATCCGCGGCA-3=Apod NM_007470 5=-AACGTCTCAGAGCCAGCCA-3= 5=-Fam-AAGTCCAGTTCTTCCCGTTGATGCCA-BHQ-3= 5=-AGGATCCAGTAGGGTGCCG-3=Cdkn1a NM_007669 5=-TTCCGCACAGGAGCAAACT-3= 5=-Fam-CCGTTGTCTCTTCGGTCCCGTGG-BHQ-3= 5=-CGGCGCAACTGCTCACT-3=Comt NM_007744 5=-AGAGAAGGAGTGGGCCATGA-3= 5=-Fam-CGTGGGTGACGCAAAAGGCCAA-BHQ-3= 5=-CCGAATCACTGCATCCATGA-3=Edg3 NM_010101 5=-AACCAGCCCAGATGCGC-3= 5=-Fam-TTGCAGAACGAGAGCCTATTTTCAACACTCTTC-BHQ-3= 5=-GCGTGCAGGCCCGAC-3=Nfkbia NM_010907 5=-CAGCTCACGGAGGACGGA-3= 5=-Fam-ACTCGTTCCTGCACTTGGCAATCATCC-BHQ-3= 5=-ATGGTCAGCGGCTTCTCTTC-3=Fosb NM_008036 5=-CCAGAGCCAGGCCTAGAAGAC-3= 5=-Fam-TCGCAGAGAGCGGAACAAGC-BHQ-3= 5=-CCGACGGTTCCTGCACTTA-3=Gadd45b NM_008655 5=-AGGCGGCCAAACTGATGA-3= 5=-Fam-TGTGGACCCCGACAGCGTGG-BHQ-3= 5=-CATCCTCCTCTTCTTCGTCTATGG-3=Gadph NM_001001303 5=-TGGATCTGACGTGCCGC-3= 5=-Fam-TGGAGAAACCTGCCAAGTATGATGACATCA-BHQ-3= 5=-TGCCTGCTTCACCACCTTC-3=Hbegf NM_010415 5=-TGCTGCCGTCGGTGATG-3= 5=-Fam-TGAAGCTCTTTCTGGCCGCAGTGTTG-BHQ-3= 5=-ACCGGTCACCAACGCG-3=Hmox1 NM_010442 5=-GTGATGGAGCGTCCACAGC-3= 5=-Fam-CGACAGCATGCCCCAGGATTTGTC-BHQ-3= 5=-TGGTGGCCTCCTTCAAGG-3=Kcnj10 NM_001039484 5=-CTCCGCTCGCCGCTC-3= 5=-Fam-CAGCCACTTCACCTTCGAGCCAAGA-BHQ-3= 5=-ACTGTAATAGACCTTAGCGACCGAC-3=Nr4a1 NM_010444 5=-GGCCACAGGGAGTGGGA-3= 5=-Fam-CCGGCTGGAGATGCCCTGTATTCAAG-BHQ-3= 5=-CGTTGCTGGTGTTCCATATTGA-3=Pdlim4 NM_019417 5=-AGTGCACGCGCTGCG-3= 5=-Fam-ACGGGATCGTGGGAACCATTGTCA-BHQ-3= 5=-ATGGTAGAGCTTGTCTCTCGCC-3=Pink1 NM_026880 5=-GAGCGTGGTGGCAATGG-3= 5=-Fam-TGATGGCCCCTGAGGTGTCCACA-BHQ-3= 5=-CGGCTTTGCTGTAGTCAATTACC-3=Tnrsf12a NM_013749 5=-GCCGCCGGAGAGAAAAG-3= 5=-Fam-TTACTACCCCCATAGAGGAGACTGGTGGAGAG-BHQ-3= 5=-GCCACACCTGGGCAGC-3=
Table 2. List of primers used to prepare the standards for qRT-PCR
Gene name Enzyme Forward primer Enzyme Reverse primer
Agxt2l1 HindIII 5=-CCCAAGCTTGGGATGTGCGAGCTCTATAGCAA-3= XhoI 5=-CCGCTCGAGCGGCTCATGTCTTGAGTCTTTTGCT-3=Apod KpnI 5=-CGGGGTACCCCGATGGTGACCATGCTGATGTTCC-3= EcoRI 5=-CCGGAATTCCGGTTACAGGAAGTCCGGGCAGTT-3=Cdkn1a HindIII 5=-CCCAAGCTTGGGATGTCCAATCCTGGTGATGT-3= EcoRI 5=-CCGGAATTCCGGCTTCAGGGTTTTCTCTTGCA-3=Comt KpnI 5=-CGGGGTACCCCGATGCTGTTGGCTGCTGTCTC-3= EcoRI 5=-CCGGAATTCCGGTCAGGACTTCACGGGGCTGC-3=Edg3 EcoRI 5=-CCGGAATTCCGGATGGCAACCACGCATGCGCA-3= XhoI 5=-CCGCTCGAGCGGTCACTTGCAGAGGACCCCGT-3=Fosb BamHI 5=-CGCGGATCCGCGGTGAAACCGACAGAGCCTGG-3= EcoRI 5=-CCGGAATTCCGGGTTCCTTGCGGGTTTGTTTG-3=Gadd45b BamHI 5=-CGCGGATCCGCGATGACCCTGGAAGAGCTGGT-3= EcoRI 5=-CCGGAATTCCGGTGGGTCTCAGCGTTCCTCTA-3=Gapdh BamHI 5=-CGCGGATCCGCGATGGTGAAGGTCGGTGTGAA-3= EcoRI 5=-CCGGAATTCCGGTTCTTACTCCTTGGAGGCCA-3=Hbegf HindIII 5=-CCCAAGCTTGGGATGAAGCTGCTGCCGTCGGT-3= EcoRI 5=-CCGGAATTCCGGATAGCTCAGGTCCTCCTCAGTGGG-3=Hmox1 BamHI 5=-CGCGGATCCGCGTAGCCCAGTCCGGTGATGGA-3= EcoRI 5=-CCGGAATTCCGGTGGGGGCCAGTATTGCATTT-3=Kcnj10 BamHI 5=-CGCGGATCCGCGCTCCGCTCGCCGCTCCTGCC-3= EcoRI 5=-CCGGAATTCCGGTCAGACGTTGCTGATGCGCA-3=Nfkbia HindIII 5=-CCCAAGCTTGGGAGCCATGTTTCAGCCAGCTG-3= XhoI 5=-CCGCTCGAGCGGTTATAATGTCAGACGCTGGC-3=Nr4a1 BamHI 5=-CGCGGATCCGCGTGCTAGAAGGACTGCGGAGC-3= EcoRI 5=-CCGGAATTCCGGGGCTTAAAGGCACATGGGTG-3=Pdlim4 HindIII 5=-CCCAAGCTTGGGATGACCCACTCGGTGACCCTG-3= EcoRI 5=-CCGGAATTCCGGCAGCTCAGACAAGTTCCACCT-3=Pink1 KpnI 5=-CGGGGTACCCCGATGGCGGTGCGACAGGCACTG-3= EcoRI 5=-CCGGAATTCCGGTCATGGGGCTGCCCTCCAGGA-3=Tnfrsf12a BamHI 5=-CGCGGATCCGCGGCAATCATGGCTCCGGGTTG-3= EcoRI 5=-CCGGAATTCCGGTGAATCACCACCTCGCCCCA-3=
R.
Pattariniet
al./
Neuroscience
155(2008)
1174–1194
1177

S
Spq4Uapawtrpm(
T
Tti(smaldewimmdftiufmtb
da(uIrlmtg2alomtc
wet
mePeMetsJGt((Sefst(hlmra
vcatggnc(Psa22Setvna(
2isbstL
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941178
tatistical analysis
tatistical methods used to analyze microarray results are ex-lained in the microarray analysis section. Statistical analysis forRT-PCR results was performed with GraphPad Prism® version.03 for Windows® (GraphPad Software Inc., San Diego, CA,SA). Results are expressed as the ratio of number of copies ofspecific gene over the number of copies of glyceraldehyde-3-
hosphate dehydrogenase (Gapdh). Each time point is the aver-ge of at least three animals. The temporal profile of each geneas analyzed by one-way ANOVA followed by Bonferroni’s mul-
iple comparisons test to assess statistical significance versusespective control (time zero). Comparison between strains waserformed either by two-way ANOVA followed by Bonferroni’sultiple comparisons test (�two groups) or by Student’s t-test
two groups).
RESULTS
he MPTP striatal transcriptome in C57BL/6J mice
o investigate the temporal transcriptional responses inhe striatum in MPTP-sensitive strain, C57BL/6J mice werenjected every 2 h with either saline (as control) or MPTP20 mg/kg) for a total of four injections. This injectionchedule, sometimes referred to as the acute MPTPodel, is used widely to simulate PD in mice and leads totemporally predictable sequence of molecular and cellu-
ar events that culminate in the relatively synchronouseath of SNpc neurons (Przedborski and Vila, 2003; Millert al., 2005; Smeyne and Jackson-Lewis, 2005). Animalsere killed at 5 (three injections), 24 and 72 h (both four
njections) after the first dose of MPTP, the striatum re-oved and total RNA isolated and used for Affymetrixicroarray analysis as described in Experimental Proce-ures. These time points were chosen to span the periodrom the acute consequences of MPTP intoxicationhrough times when formal perturbation of DA nerve end-ngs and compromised SNpc neuron function are evidentp to the point when DA neurons begin to die. Total RNArom each animal was loaded onto individual Affymetrixicroarray chips. Experimental reproducibility can be es-
imated by comparing columns within a figure as well asetween corresponding columns in Fig. 1.
Using criteria described in the Experimental Proce-ures section, we identified 107, 287 and 191 probe setss differentially expressed at 5, 24 and 72 h, respectivelySupplemental Table 1). This target list of probe sets wassed to perform hierarchical cluster, Gene Ontology and
ngenuity Pathway analyses. Hierarchical cluster analysiseveals three largely discrete sets of genes whose mRNAevels change sequentially over time following MPTP ad-
inistration (Fig. 1 and Supplemental Table 1). At earlyime points (Fig. 1), the levels of mRNA for a number ofenes increase and then largely decline to basal values by4 h (Fig. 1 and Supplemental Table 1). By 24 h a distinctnd larger set of mRNAs is increased (Fig. 1) and then
argely declines to baseline by 72 h at which time a new setf gene expression changes is evident (Fig. 1 and Supple-ental Table 1). Although less in number, there were also
ransient decreases in some mRNAs over the same time
ourse (Supplemental Table 1). Additional microarray data 2ere obtained at 2 and 96 h post-MPTP treatment. Genexpression changes seen at 2 and 96 h were subsets ofhose seen at 5 and 72 h, respectively (data not shown).
A variety of bioinformatic tools was used to analyzeRNA changes. As expected from prior studies (Duch-min et al., 1992; Doucet et al., 1996; Smith et al., 1997;erez-Otano et al., 1998; Hunot et al., 2004), immediatearly genes are prominent in the early phase followingPTP treatment and include the transcriptional regulatorsarly growth response 3 and 4 (Egr3 and Egr4, respec-
ively), FBJ osteosarcoma oncogene (c-Fos), FBJ osteo-arcoma oncogene B (Fosb), Jun oncogene (c-Jun) andun oncogene B (Junb) (Supplemental Table 1). Moreover,ene Ontology analysis revealed that transcription fac-
ors/regulators were over-represented in the early phase20.9%) versus intermediate (5.3%) and late phases6.3%). Similarly, Ingenuity Pathway Analysis (Ingenuityystem Inc., Redwood City, CA, USA) indicated that genexpression was statistically the most over-representedunction (2.28e-08�P�8.72e-03) in the early phase re-ponse. Examples include changes in mRNA levels for theranscriptional regulators BTB and CNC homology 1Bach1), B-cell translocation gene 3 (Btg3), CCAAT/en-ancer binding protein (C/EBP), beta (Cebpb), Kruppel-
ike factor 9 (Klf9), nuclear receptor subfamily 4, group A,ember 3 (Nr4a3), paired box gene 8 (Pax8), retinoid X
eceptor gamma (Rxrg), superoxide dismutase two (Sox2)nd zinc finger and BTB domain containing 16 (Zbtb16).
Another major component of the early response in-olves genes implicated in oxidative stress and includesyclin-dependent kinase inhibitor 1A (Cdkn1a), DNA-dam-ge-inducible transcript 4 (Ddit4), DNA-damage-inducibleranscript 4-like (Ddit4l), FK506 binding protein 5 (Fkbp5),rowth arrest and DNA-damage-inducible 45 beta andamma (Gadd45b and Gadd45g), metallothionein 2 (Mt2),uclear factor of kappa light chain gene enhancer in B-ells inhibitor, alpha (NFkBia) and uncoupling protein 2Ucp2). These changes are also consistent with studies inD and models of the disorder where evidence of oxidativetress have been reported (Kindt et al., 1987; Krueger etl., 1990; Bates et al., 1994; Hasegawa et al., 1995; Beal,003; Przedborski and Vila, 2003; Hald and Lotharius,005; Jackson-Lewis and Smeyne, 2005; Hald et al., 2007;chapira, 2008). Other gene expression changes in thearly phase represent inflammatory responses (Tnf recep-or-associated factor 5 (Traf5) and interferon-related de-elopmental regulator one (Ifrd1)) and steroid/stress sig-aling (serum/glucocorticoid regulated kinase 1 and 3 (Sgknd Sgk3), TSC22 domain family, members 1 and 3Tsc22d1 and Tsc22d3)).
GSEA (Mootha et al., 2003; Subramanian et al., 2005,007) revealed that the intermediate phase is character-
zed by enrichment for transcripts implicated in cytokineignaling and inflammatory responses (Supplemental Ta-les 1 and 6). This result is consistent with many studieshowing the presence of inflammatory responses in stria-um in both PD and animal models thereof (Hald andotharius, 2005; Nagatsu and Sawada, 2005; Hald et al.,
007; Hunter et al., 2007; McGeer and McGeer, in press;
Wf(ewa(ccinCpfiaabt16tec
ct(daGgAStrRapcsapiioladab(ptecBpf
ccmdeL(Si9ss
ciafgscaspmoam4asapaoicta(e((rtls
Ab
TuoBgt(p
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1179
ilms et al., 2007). Expression of genes involved in TNFamily signaling such as the receptors for TNF-alphaTnfrsf1a) and Tweak (Tnfsf12a) is increased. Likewise,xpression of genes involved in interleukin signaling path-ays such as suppressor of cytokine signaling 3 (Socs3)nd signal transducer and activator of transcription 1 and 3Stat1 and Stat3) is elevated. Besides genes involved inytokine and chemokine signaling, many effector mole-ules of the inflammatory response are increased in thentermediate phase, including the complement compo-ents 1, q subcomponent, alpha and beta polypeptide andchain (C1qa, C1qb and C1qc, respectively), and com-
lement component 4B (C4b); Fc receptor, IgG, high af-nity I (Fcgr1); cathepsin B, C, D and Z (Ctsb, Ctsc, Ctsdnd Ctsz, respectively); lectin, galactose binding, soluble 1nd 3 (Lgals1 and Lgals3, respectively) and the Lgals3inding protein (Lgals3bp). Similarly, markers of inflamma-ory and immune cells such as allograft inflammatory factor
(Aif1 a.k.a. Iba1, a microglia marker), CD antigens 44,8, 151 and 180 (Cd44, Cd68, Cd151 and Cd180, respec-
ively), lymphocyte antigen 86 (Ly86), macrophage scav-nger receptor 2 (Msr2) and oncostatin M receptor (Osmr)hange in the intermediate phase.
Also prominent in the intermediate phase are in-reased transcript levels for genes associated with activa-ion of astrocytes, including glial fibrillary acidic proteinGfap) and vimentin (Vim). We also, confirm our earlieremonstration of elevated Hmox1 expression in striatalstrocytes following MPTP administration (Fernandez-onzalez et al., 2000). Although not a specific marker forliosis, the levels of S100 calcium binding proteins A6,10, A11, A13 and A16 (S100a6, S100a10, S100a11,100a13 and S100a16, respectively) as well as their in-
eracting proteins, annexin A2 and A3 (Anxa2 and Anxa3,espectively) (Bianchi et al., 1992; Tokumitsu et al., 1992;ety et al., 1999; Santamaria-Kisiel et al., 2006; Reschernd Gerke, 2008) are also increased in the intermediatehase. In addition, a number of other gene products asso-iated with protein folding, modification and elimination,uch as heat shock protein 1, B6 and 8 (Hspb1, Hspb6nd Hspb8, respectively), transglutaminase 1, K and Colypeptides (Tgm1 and Tgm2, respectively) and tissue
nhibitor of metalloproteinase 1 (Timp1) are elevated. Alsondicative of ongoing responses to cellular damage andxidative stress are elevation in levels of mRNAs for apo-
ipoprotein D (Apod), fatty acid binding protein 7 (Fabp7)nd Mt2. In addition mRNA levels of genes linked with celleath such as myeloid cell leukemia sequence 1 (Mcl1)nd transmembrane BAX inhibitor motif containing 1 (Tm-im1) and macroautophagy BclII-associated athanogene 3Bag3) (Carra et al., 2008) change in the intermediatehase. Besides gene products overtly linked to inflamma-ion, gliosis, and cellular damage and stress responses,xpression of genes involved in other signaling pathwayshanges, including bone morphogenetic protein 1 (Bmp1),MP2 inducible kinase (Bmp2k), CD9 antigen (Cd9), he-arin-binding EGF-like growth factor (Hbegf) and trans-
orming growth factor, beta receptor II (Tgfbr2). 4By 72 h post-treatment the majority of the mRNAhanges seen at 24 h return to basal levels and a newohort of transcripts are altered. The persistently alteredRNAs are those linked to gliosis, inflammation and oxi-ative stress and include, Gfap, Vim, C1qc and C4b, Ly86,ndothelin receptor type B (Ednrb), Hspb6, Lgals1 andgals3bp, lysosomal-associated membrane protein 2Lamp2), legumain (Lgmn), metallothionein 1 (Mt1),100a6 and S100a13, and transferrin (Trf). The same
nflammation/gliosis-related mRNAs are also elevated at6 h post treatment indicating persistent inflammatory re-ponses and ongoing astrogliosis in striatum (data nothown).
In the late phase, a new cluster of gene expressionhanges is evident. Several immediate-early genes includ-
ng Egr3 and Fos-like antigen 2 (Fosl2) are down-regulatedt 72 and 96 h. The mRNA levels for the transcriptionactor ets variant gene 5 (Etv5) and for brain-specific an-iogenesis inhibitor 1-associated protein 2 (Baiap2), a pre-umptive immediate-early gene are also persistently de-reased whereas levels of the transcriptional regulatorsctivating transcription factor 6 (Atf6), nuclear receptorubfamily 2, group F, member 2 (Nr2f2) and zinc fingerrotein of the cerebellum 1 (Zic1) are increased. TheRNAs levels for many membrane and secreted proteinsr proteins that modify the extracellular matrix also changet 72 h and include aquaporin 4 (Aqp4), gap junctionembrane channel protein alpha 1 (Gja1 a.k.a. connexin3), myelin oligodendrocyte glycoprotein (Mog), neural celldhesion molecule 1 (Ncam1), proteolipid protein 1 (Plp1),olute carrier family 44, member 1 (Slc44a1), secretedcidic cysteine rich glycoprotein (Sparc), secreted phos-hoprotein 1 (Spp1) and tissue inhibitor of metalloprotein-se 4 (Timp4). Also prominent are changes in expressionf genes associated with specific neuronal subtypes and
nclude, parvalbumin (Pvalb), potassium voltage-gatedhannel, subfamily Q, member 5 (Kcnq5), and the GABAransporter solute carrier family 6, member 11, (Slc6a11),s well as general neuronal proteins such as bassoonBsn) and homer homolog 1 (Homer1). Finally, the mRNAsncoding two proteins implicated in PD, alpha-synucleinSnca) (decrease) and G protein-coupled receptor 37Gpr37 a.k.a. Pael-R) (increase) are altered in the lateesponse phase. Moreover, the same changes in thesewo transcripts are also evident at 96 h suggesting that theatter two are more long-lasting alterations in gene expres-ion (data not shown).
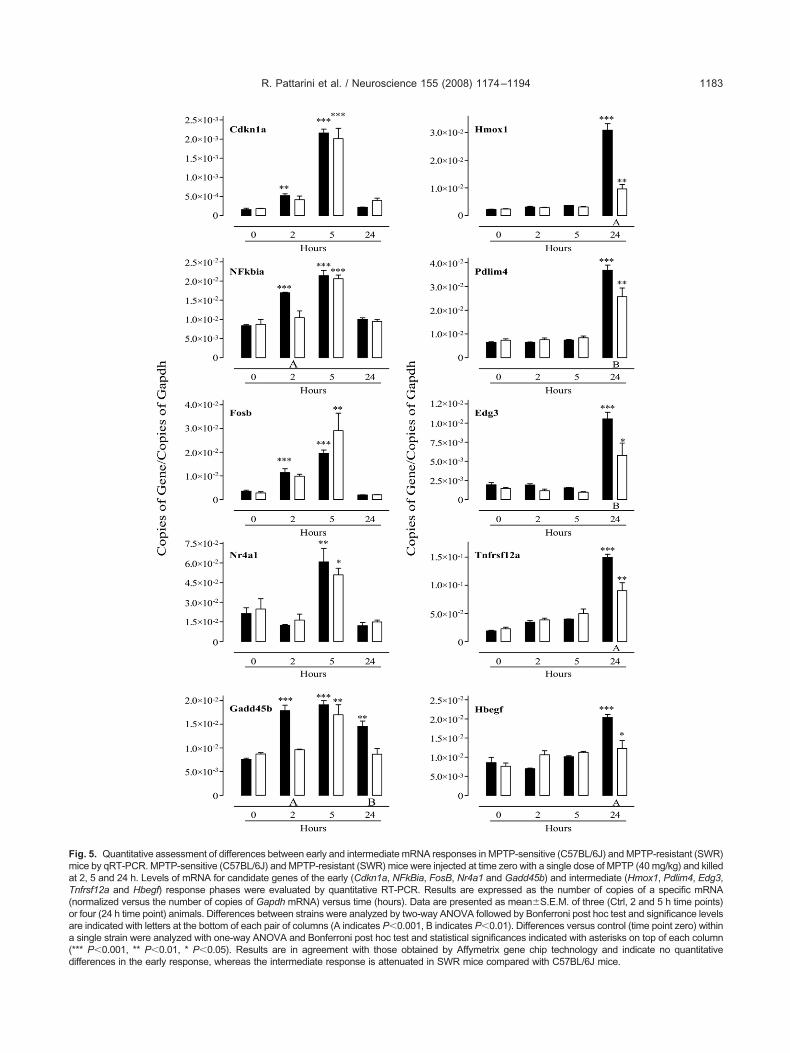
ssessment of temporal mRNA changesy qRT-PCR
o confirm and extend the microarray data, qRT-PCR wassed to assess the temporal profiles of mRNA expressionf selected genes representative of early (Cdkn1a, NFk-ia, Gadd45b, Fosb and nuclear receptor subfamily 4,roup A, member 1 (Nr4a1)) and intermediate (Hmox1,umor necrosis factor receptor superfamily, member 12Tnfrsf12a), endothelial differentiation, sphingolipid G-rotein-coupled receptor 3 (Edg3), PDZ and LIM domain
(Pdlim4) and Hbegf) phase transcripts (Fig. 2). Early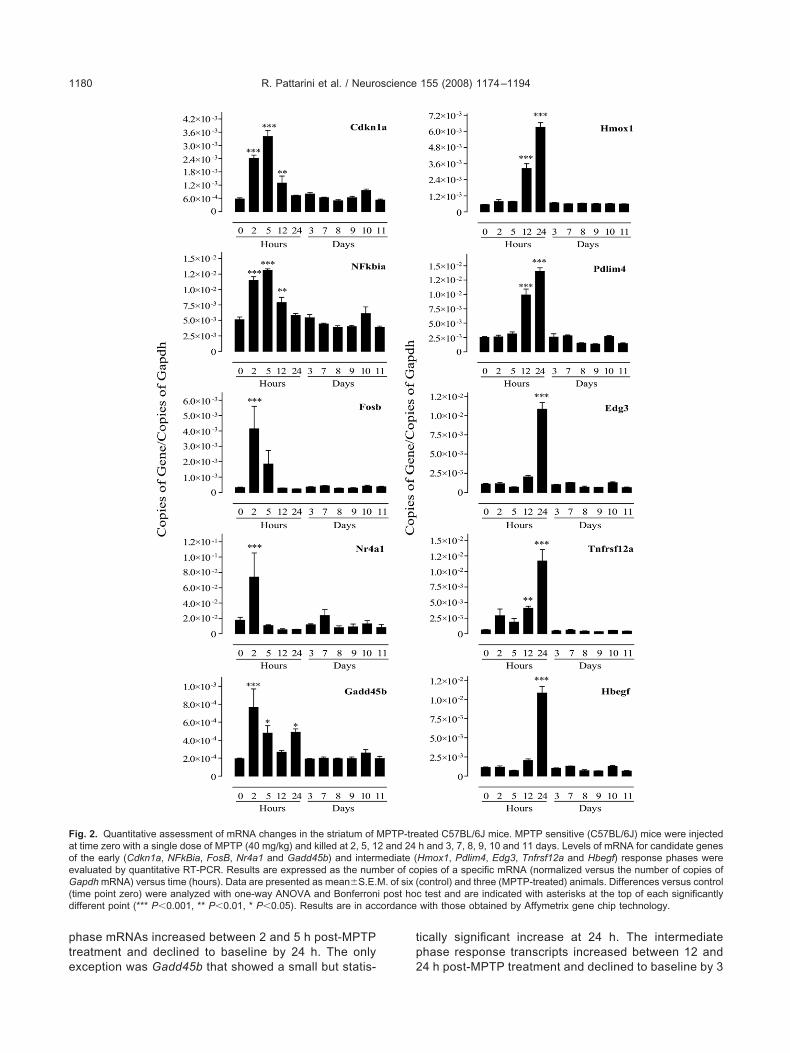
pte
tp
FaoeG(d ordance
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941180
hase mRNAs increased between 2 and 5 h post-MPTPreatment and declined to baseline by 24 h. The only
ig. 2. Quantitative assessment of mRNA changes in the striatum oft time zero with a single dose of MPTP (40 mg/kg) and killed at 2, 5, 1f the early (Cdkn1a, NFkBia, FosB, Nr4a1 and Gadd45b) and intermvaluated by quantitative RT-PCR. Results are expressed as the numapdh mRNA) versus time (hours). Data are presented as mean�S.E.
time point zero) were analyzed with one-way ANOVA and Bonferronifferent point (*** P�0.001, ** P�0.01, * P�0.05). Results are in acc
xception was Gadd45b that showed a small but statis- 2
ically significant increase at 24 h. The intermediatehase response transcripts increased between 12 and
ated C57BL/6J mice. MPTP sensitive (C57BL/6J) mice were injectedh and 3, 7, 8, 9, 10 and 11 days. Levels of mRNA for candidate genesHmox1, Pdlim4, Edg3, Tnfrsf12a and Hbegf) response phases werepies of a specific mRNA (normalized versus the number of copies ofcontrol) and three (MPTP-treated) animals. Differences versus controlc test and are indicated with asterisks at the top of each significantlywith those obtained by Affymetrix gene chip technology.
MPTP-tre2 and 24ediate (
ber of coM. of six (i post ho
4 h post-MPTP treatment and declined to baseline by 3
dc
Bm
WtGeswficruiatertc
tud
efttstepaag
Ts
Tow6s22sacccr
Fasatc1dfii
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1181
ays. These data serve to confirm and extend the mi-roarray analysis.
rain region specificity of MPTP-inducedRNA changes
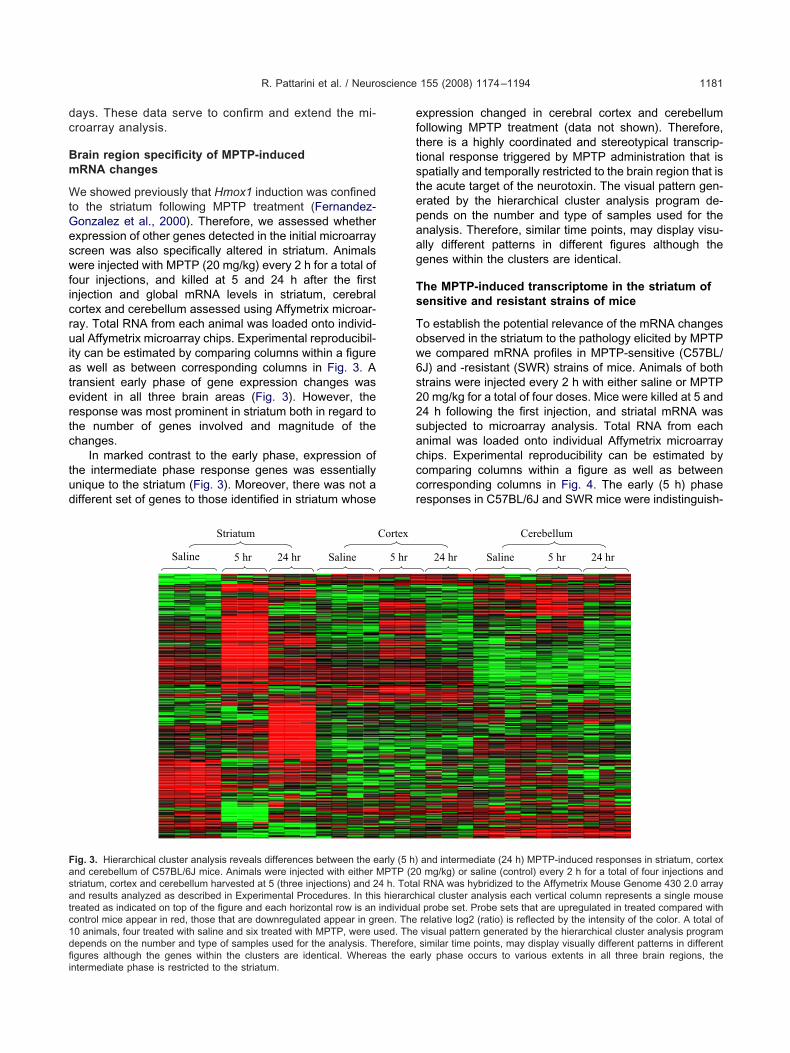
e showed previously that Hmox1 induction was confinedo the striatum following MPTP treatment (Fernandez-onzalez et al., 2000). Therefore, we assessed whetherxpression of other genes detected in the initial microarraycreen was also specifically altered in striatum. Animalsere injected with MPTP (20 mg/kg) every 2 h for a total of
our injections, and killed at 5 and 24 h after the firstnjection and global mRNA levels in striatum, cerebralortex and cerebellum assessed using Affymetrix microar-ay. Total RNA from each animal was loaded onto individ-al Affymetrix microarray chips. Experimental reproducibil-
ty can be estimated by comparing columns within a figures well as between corresponding columns in Fig. 3. A
ransient early phase of gene expression changes wasvident in all three brain areas (Fig. 3). However, theesponse was most prominent in striatum both in regard tohe number of genes involved and magnitude of thehanges.
In marked contrast to the early phase, expression ofhe intermediate phase response genes was essentiallynique to the striatum (Fig. 3). Moreover, there was not aifferent set of genes to those identified in striatum whose
Saline 5 hr 24 hr Saline
Striatum
ig. 3. Hierarchical cluster analysis reveals differences between the end cerebellum of C57BL/6J mice. Animals were injected with either Mtriatum, cortex and cerebellum harvested at 5 (three injections) and 2nd results analyzed as described in Experimental Procedures. In thisreated as indicated on top of the figure and each horizontal row is anontrol mice appear in red, those that are downregulated appear in gr0 animals, four treated with saline and six treated with MPTP, were uepends on the number and type of samples used for the analysis. T
gures although the genes within the clusters are identical. Whereas the eantermediate phase is restricted to the striatum.xpression changed in cerebral cortex and cerebellumollowing MPTP treatment (data not shown). Therefore,here is a highly coordinated and stereotypical transcrip-ional response triggered by MPTP administration that ispatially and temporally restricted to the brain region that ishe acute target of the neurotoxin. The visual pattern gen-rated by the hierarchical cluster analysis program de-ends on the number and type of samples used for thenalysis. Therefore, similar time points, may display visu-lly different patterns in different figures although theenes within the clusters are identical.
he MPTP-induced transcriptome in the striatum ofensitive and resistant strains of mice
o establish the potential relevance of the mRNA changesbserved in the striatum to the pathology elicited by MPTPe compared mRNA profiles in MPTP-sensitive (C57BL/J) and -resistant (SWR) strains of mice. Animals of bothtrains were injected every 2 h with either saline or MPTP0 mg/kg for a total of four doses. Mice were killed at 5 and4 h following the first injection, and striatal mRNA wasubjected to microarray analysis. Total RNA from eachnimal was loaded onto individual Affymetrix microarrayhips. Experimental reproducibility can be estimated byomparing columns within a figure as well as betweenorresponding columns in Fig. 4. The early (5 h) phaseesponses in C57BL/6J and SWR mice were indistinguish-
Saline 5 hr 24 hr24 hr
Cerebellum
and intermediate (24 h) MPTP-induced responses in striatum, cortexmg/kg) or saline (control) every 2 h for a total of four injections and
l RNA was hybridized to the Affymetrix Mouse Genome 430 2.0 arrayical cluster analysis each vertical column represents a single mousel probe set. Probe sets that are upregulated in treated compared withrelative log2 (ratio) is reflected by the intensity of the color. A total ofvisual pattern generated by the hierarchical cluster analysis programsimilar time points, may display visually different patterns in different
5 hr
Cortex
arly (5 h)PTP (20
4 h. Totahierarch
individuaeen. Thesed. Theherefore,
rly phase occurs to various extents in all three brain regions, the
adtiIamtmtmwstMm(w(metamS
rswt
lpCisetlatbHmaSws
t
FoheMvabtpdM nuated in
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941182
ble (Fig. 4). This suggests there is unlikely to be a strain-ependent difference in entry of MPTP into brain, consis-ent with a recent chemical determination of MPP� levelsn brains of C57BL/6J and SWR mice (Boyd et al., 2007).n contrast, the intermediate (24 h) response was attenu-ted in SWR mice (Fig. 4). Whereas the magnitude ofRNA changes observed in C57BL/6J mice was consis-
ent from animal to animal the degree of change in SWRice varied greatly between animals and with respect
o individual genes. Thus, some MPTP-treated SWRice were indistinguishable from saline-treated animalshereas others showed more robust responses that forome probe sets approximated levels observed in sensi-ive C57BL/6J mice. Although SWR mice are consideredPTP resistant, this is a relative term. In the acute MPTPodel, SWR mice exhibit an approximate 14% loss of DA
tyrosine hydroxylase-positive) SNpc neurons comparedith 58% loss in C57BL/6J mice under identical conditions
Hamre et al., 1999). Thus, it is expected that if the inter-ediate response is linked to neuronal loss, it should bevident to some extent even in SWR mice. Furthermore,he neuronal loss in SWR mice is variable, with somenimals having no apparent loss whereas others haveore substantial losses. Therefore, it is possible that the
C57BL/6J
Saline Saline 5 hr
SWR C
Saline5 hr
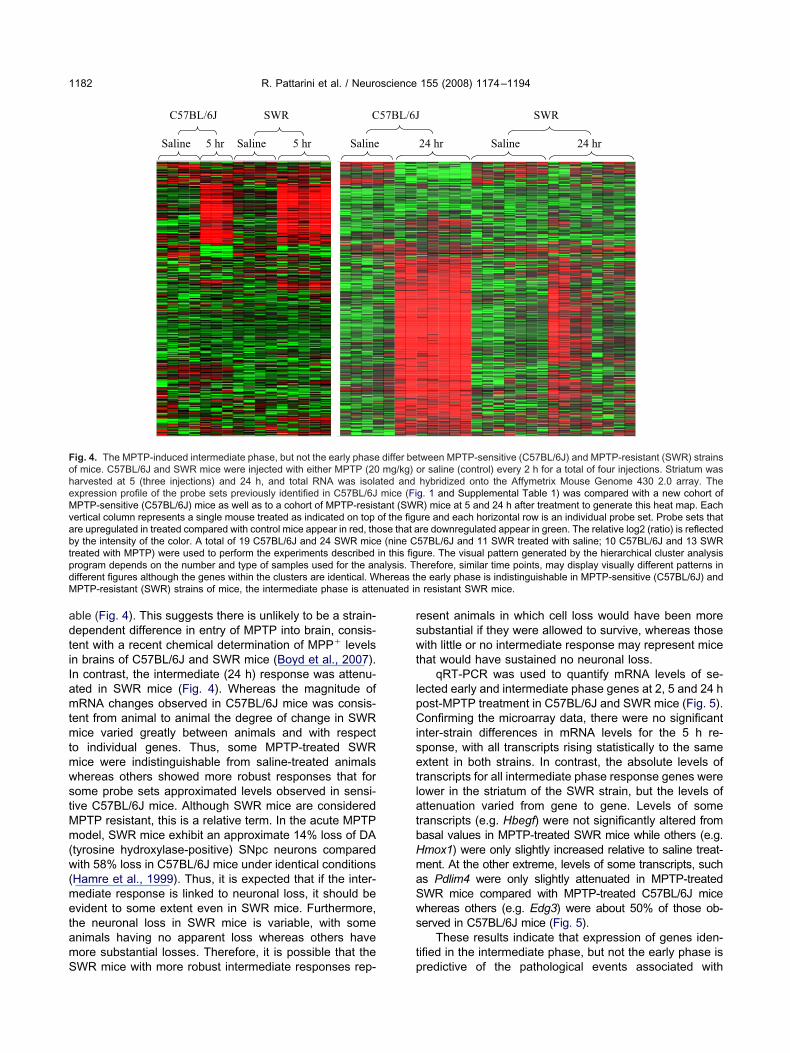
ig. 4. The MPTP-induced intermediate phase, but not the early phasef mice. C57BL/6J and SWR mice were injected with either MPTP (20arvested at 5 (three injections) and 24 h, and total RNA was isolaxpression profile of the probe sets previously identified in C57BL/6JPTP-sensitive (C57BL/6J) mice as well as to a cohort of MPTP-resis
ertical column represents a single mouse treated as indicated on topre upregulated in treated compared with control mice appear in red, thy the intensity of the color. A total of 19 C57BL/6J and 24 SWR micereated with MPTP) were used to perform the experiments describedrogram depends on the number and type of samples used for the anifferent figures although the genes within the clusters are identical. WPTP-resistant (SWR) strains of mice, the intermediate phase is atte
WR mice with more robust intermediate responses rep- p
esent animals in which cell loss would have been moreubstantial if they were allowed to survive, whereas thoseith little or no intermediate response may represent mice
hat would have sustained no neuronal loss.qRT-PCR was used to quantify mRNA levels of se-
ected early and intermediate phase genes at 2, 5 and 24 host-MPTP treatment in C57BL/6J and SWR mice (Fig. 5).onfirming the microarray data, there were no significant
nter-strain differences in mRNA levels for the 5 h re-ponse, with all transcripts rising statistically to the samextent in both strains. In contrast, the absolute levels ofranscripts for all intermediate phase response genes wereower in the striatum of the SWR strain, but the levels ofttenuation varied from gene to gene. Levels of someranscripts (e.g. Hbegf) were not significantly altered fromasal values in MPTP-treated SWR mice while others (e.g.mox1) were only slightly increased relative to saline treat-ent. At the other extreme, levels of some transcripts, suchs Pdlim4 were only slightly attenuated in MPTP-treatedWR mice compared with MPTP-treated C57BL/6J micehereas others (e.g. Edg3) were about 50% of those ob-erved in C57BL/6J mice (Fig. 5).
These results indicate that expression of genes iden-ified in the intermediate phase, but not the early phase is
24 hr
SWR
Saline 24 hr
tween MPTP-sensitive (C57BL/6J) and MPTP-resistant (SWR) strainsr saline (control) every 2 h for a total of four injections. Striatum washybridized onto the Affymetrix Mouse Genome 430 2.0 array. Theg. 1 and Supplemental Table 1) was compared with a new cohort ofR) mice at 5 and 24 h after treatment to generate this heat map. Eachre and each horizontal row is an individual probe set. Probe sets thatre downregulated appear in green. The relative log2 (ratio) is reflected57BL/6J and 11 SWR treated with saline; 10 C57BL/6J and 13 SWRure. The visual pattern generated by the hierarchical cluster analysiserefore, similar time points, may display visually different patterns ine early phase is indistinguishable in MPTP-sensitive (C57BL/6J) andresistant SWR mice.
57BL/6J
differ bemg/kg) oted andmice (Fi
tant (SWof the figuose that a
(nine Cin this figalysis. Thhereas th
redictive of the pathological events associated with
FmaT(oaa(d
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1183
ig. 5. Quantitative assessment of differences between early and intermediate mRNA responses in MPTP-sensitive (C57BL/6J) and MPTP-resistant (SWR)ice by qRT-PCR. MPTP-sensitive (C57BL/6J) and MPTP-resistant (SWR) mice were injected at time zero with a single dose of MPTP (40 mg/kg) and killedt 2, 5 and 24 h. Levels of mRNA for candidate genes of the early (Cdkn1a, NFkBia, FosB, Nr4a1 and Gadd45b) and intermediate (Hmox1, Pdlim4, Edg3,nfrsf12a and Hbegf) response phases were evaluated by quantitative RT-PCR. Results are expressed as the number of copies of a specific mRNAnormalized versus the number of copies of Gapdh mRNA) versus time (hours). Data are presented as mean�S.E.M. of three (Ctrl, 2 and 5 h time points)r four (24 h time point) animals. Differences between strains were analyzed by two-way ANOVA followed by Bonferroni post hoc test and significance levelsre indicated with letters at the bottom of each pair of columns (A indicates P�0.001, B indicates P�0.01). Differences versus control (time point zero) withinsingle strain were analyzed with one-way ANOVA and Bonferroni post hoc test and statistical significances indicated with asterisks on top of each column
*** P�0.001, ** P�0.01, * P�0.05). Results are in agreement with those obtained by Affymetrix gene chip technology and indicate no quantitative
ifferences in the early response, whereas the intermediate response is attenuated in SWR mice compared with C57BL/6J mice.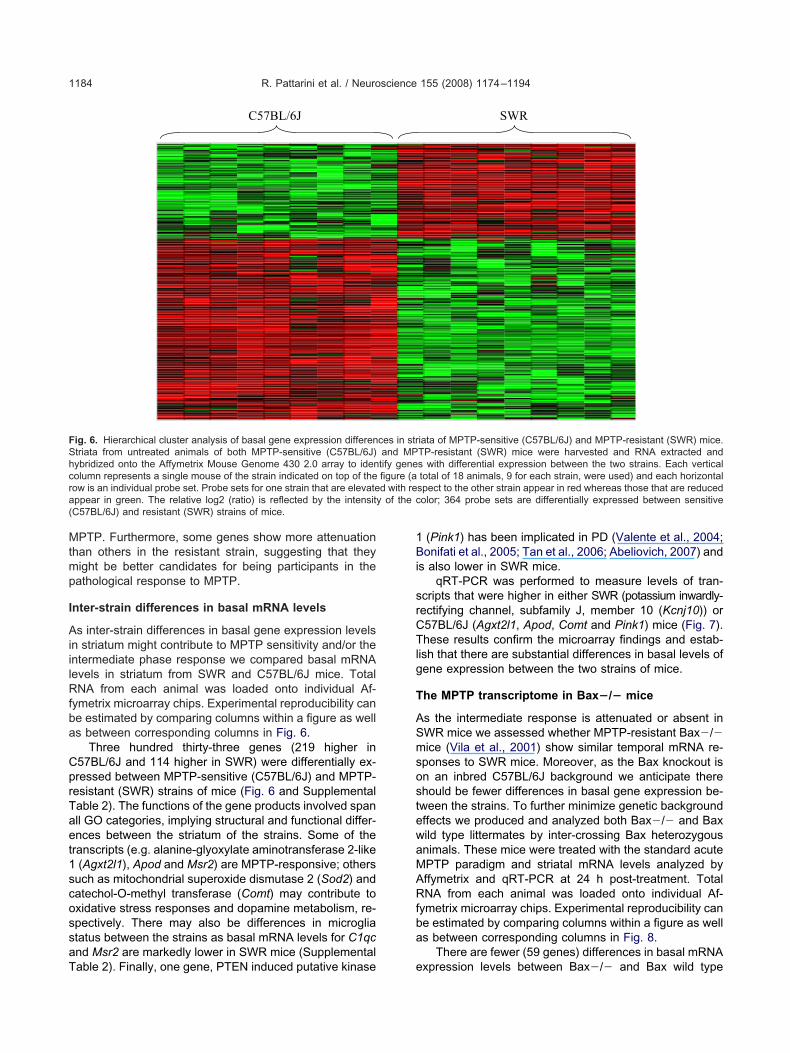
Mtmp
I
AiilRfba
CprTaet1scossaT
1Bi
srCTlg
T
ASmsostewaMARfba
FShcra(
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941184
PTP. Furthermore, some genes show more attenuationhan others in the resistant strain, suggesting that theyight be better candidates for being participants in theathological response to MPTP.
nter-strain differences in basal mRNA levels
s inter-strain differences in basal gene expression levelsn striatum might contribute to MPTP sensitivity and/or thentermediate phase response we compared basal mRNAevels in striatum from SWR and C57BL/6J mice. TotalNA from each animal was loaded onto individual Af-
ymetrix microarray chips. Experimental reproducibility cane estimated by comparing columns within a figure as wells between corresponding columns in Fig. 6.
Three hundred thirty-three genes (219 higher in57BL/6J and 114 higher in SWR) were differentially ex-ressed between MPTP-sensitive (C57BL/6J) and MPTP-esistant (SWR) strains of mice (Fig. 6 and Supplementalable 2). The functions of the gene products involved spanll GO categories, implying structural and functional differ-nces between the striatum of the strains. Some of theranscripts (e.g. alanine-glyoxylate aminotransferase 2-like(Agxt2l1), Apod and Msr2) are MPTP-responsive; others
uch as mitochondrial superoxide dismutase 2 (Sod2) andatechol-O-methyl transferase (Comt) may contribute toxidative stress responses and dopamine metabolism, re-pectively. There may also be differences in microgliatatus between the strains as basal mRNA levels for C1qcnd Msr2 are markedly lower in SWR mice (Supplemental
C57BL/6J
ig. 6. Hierarchical cluster analysis of basal gene expression differentriata from untreated animals of both MPTP-sensitive (C57BL/6J)ybridized onto the Affymetrix Mouse Genome 430 2.0 array to idenolumn represents a single mouse of the strain indicated on top of theow is an individual probe set. Probe sets for one strain that are elevateppear in green. The relative log2 (ratio) is reflected by the intensityC57BL/6J) and resistant (SWR) strains of mice.
able 2). Finally, one gene, PTEN induced putative kinase e
(Pink1) has been implicated in PD (Valente et al., 2004;onifati et al., 2005; Tan et al., 2006; Abeliovich, 2007) and
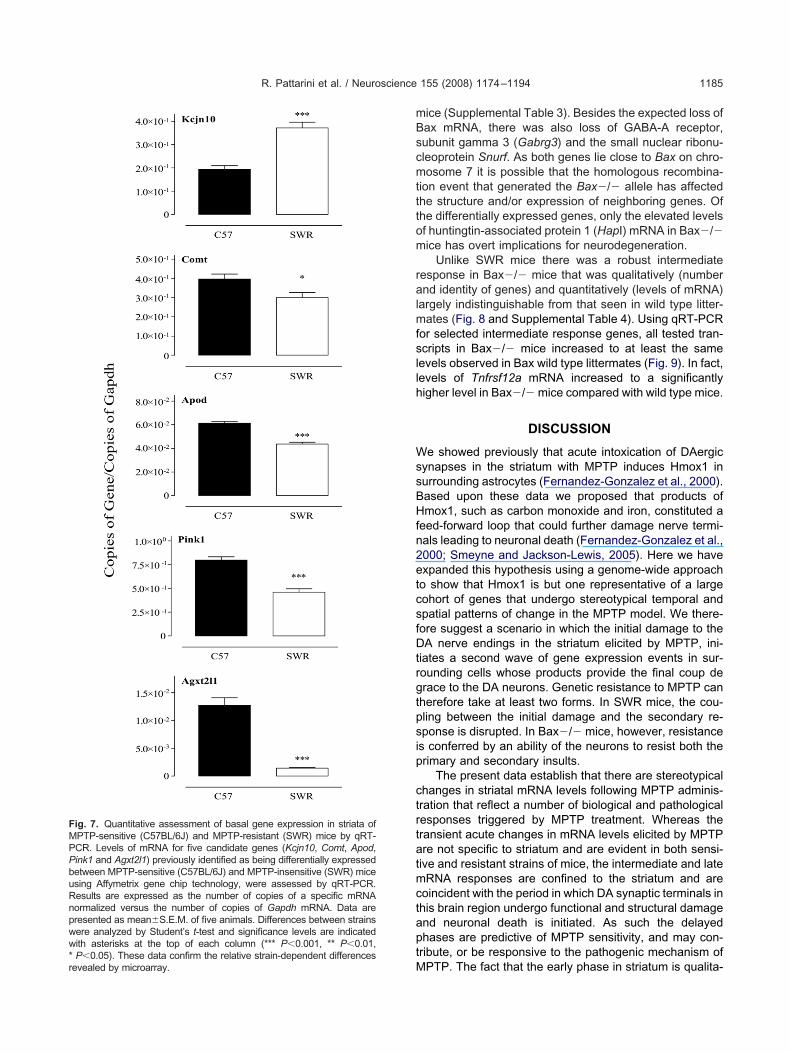
s also lower in SWR mice.qRT-PCR was performed to measure levels of tran-
cripts that were higher in either SWR (potassium inwardly-ectifying channel, subfamily J, member 10 (Kcnj10)) or57BL/6J (Agxt2l1, Apod, Comt and Pink1) mice (Fig. 7).hese results confirm the microarray findings and estab-
ish that there are substantial differences in basal levels ofene expression between the two strains of mice.
he MPTP transcriptome in Bax�/� mice
s the intermediate response is attenuated or absent inWR mice we assessed whether MPTP-resistant Bax�/�ice (Vila et al., 2001) show similar temporal mRNA re-
ponses to SWR mice. Moreover, as the Bax knockout isn an inbred C57BL/6J background we anticipate therehould be fewer differences in basal gene expression be-ween the strains. To further minimize genetic backgroundffects we produced and analyzed both Bax�/� and Baxild type littermates by inter-crossing Bax heterozygousnimals. These mice were treated with the standard acutePTP paradigm and striatal mRNA levels analyzed byffymetrix and qRT-PCR at 24 h post-treatment. TotalNA from each animal was loaded onto individual Af-
ymetrix microarray chips. Experimental reproducibility cane estimated by comparing columns within a figure as wells between corresponding columns in Fig. 8.
There are fewer (59 genes) differences in basal mRNA
SWR
iata of MPTP-sensitive (C57BL/6J) and MPTP-resistant (SWR) mice.TP-resistant (SWR) mice were harvested and RNA extracted ands with differential expression between the two strains. Each verticaltotal of 18 animals, 9 for each strain, were used) and each horizontalspect to the other strain appear in red whereas those that are reducedolor; 364 probe sets are differentially expressed between sensitive
ces in strand MPtify genefigure (ad with reof the c
xpression levels between Bax�/� and Bax wild type
mBscmtttom
ralmfsllh
WssBHfn2etcsfDtrgtpsip
ctrtatmctaptM
FMPPbuRnpww* P�0.05). These data confirm the relative strain-dependent differencesrevealed by microarray.
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1185
ice (Supplemental Table 3). Besides the expected loss ofax mRNA, there was also loss of GABA-A receptor,ubunit gamma 3 (Gabrg3) and the small nuclear ribonu-leoprotein Snurf. As both genes lie close to Bax on chro-osome 7 it is possible that the homologous recombina-
ion event that generated the Bax�/� allele has affectedhe structure and/or expression of neighboring genes. Ofhe differentially expressed genes, only the elevated levelsf huntingtin-associated protein 1 (HapI) mRNA in Bax�/�ice has overt implications for neurodegeneration.
Unlike SWR mice there was a robust intermediateesponse in Bax�/� mice that was qualitatively (numbernd identity of genes) and quantitatively (levels of mRNA)
argely indistinguishable from that seen in wild type litter-ates (Fig. 8 and Supplemental Table 4). Using qRT-PCR
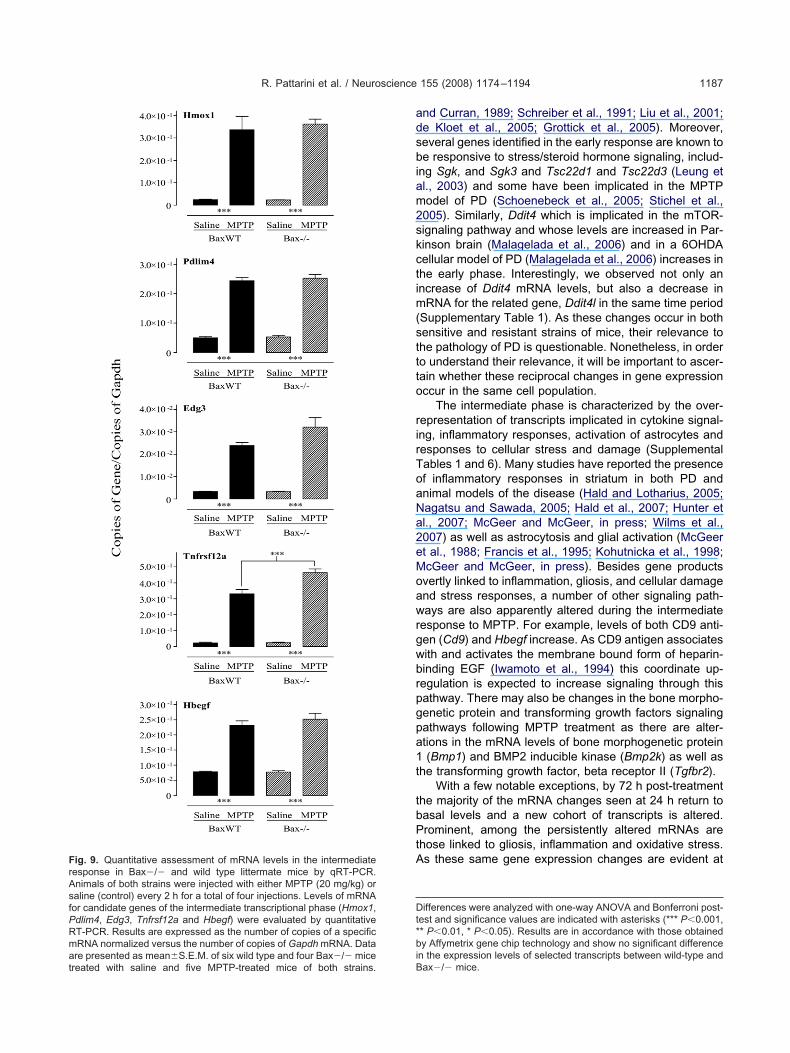
or selected intermediate response genes, all tested tran-cripts in Bax�/� mice increased to at least the sameevels observed in Bax wild type littermates (Fig. 9). In fact,evels of Tnfrsf12a mRNA increased to a significantlyigher level in Bax�/� mice compared with wild type mice.
DISCUSSION
e showed previously that acute intoxication of DAergicynapses in the striatum with MPTP induces Hmox1 inurrounding astrocytes (Fernandez-Gonzalez et al., 2000).ased upon these data we proposed that products ofmox1, such as carbon monoxide and iron, constituted a
eed-forward loop that could further damage nerve termi-als leading to neuronal death (Fernandez-Gonzalez et al.,000; Smeyne and Jackson-Lewis, 2005). Here we havexpanded this hypothesis using a genome-wide approacho show that Hmox1 is but one representative of a largeohort of genes that undergo stereotypical temporal andpatial patterns of change in the MPTP model. We there-ore suggest a scenario in which the initial damage to theA nerve endings in the striatum elicited by MPTP, ini-
iates a second wave of gene expression events in sur-ounding cells whose products provide the final coup derace to the DA neurons. Genetic resistance to MPTP canherefore take at least two forms. In SWR mice, the cou-ling between the initial damage and the secondary re-ponse is disrupted. In Bax�/� mice, however, resistances conferred by an ability of the neurons to resist both therimary and secondary insults.
The present data establish that there are stereotypicalhanges in striatal mRNA levels following MPTP adminis-ration that reflect a number of biological and pathologicalesponses triggered by MPTP treatment. Whereas theransient acute changes in mRNA levels elicited by MPTPre not specific to striatum and are evident in both sensi-ive and resistant strains of mice, the intermediate and lateRNA responses are confined to the striatum and are
oincident with the period in which DA synaptic terminals inhis brain region undergo functional and structural damagend neuronal death is initiated. As such the delayedhases are predictive of MPTP sensitivity, and may con-ribute, or be responsive to the pathogenic mechanism of
ig. 7. Quantitative assessment of basal gene expression in striata ofPTP-sensitive (C57BL/6J) and MPTP-resistant (SWR) mice by qRT-CR. Levels of mRNA for five candidate genes (Kcjn10, Comt, Apod,ink1 and Agxt2l1) previously identified as being differentially expressedetween MPTP-sensitive (C57BL/6J) and MPTP-insensitive (SWR) micesing Affymetrix gene chip technology, were assessed by qRT-PCR.esults are expressed as the number of copies of a specific mRNAormalized versus the number of copies of Gapdh mRNA. Data areresented as mean�S.E.M. of five animals. Differences between strainsere analyzed by Student’s t-test and significance levels are indicatedith asterisks at the top of each column (*** P�0.001, ** P�0.01,
PTP. The fact that the early phase in striatum is qualita-

tesMrttiBiasn
src(awa(HakPJ
Spgsm(caagmedpsmcs1mn(a
ge
FaaiMiiae two geno
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941186
ively (genes represented) and quantitatively (mRNA lev-ls) indistinguishable between C57BL/6J and SWR miceuggests that peripheral metabolism and/or penetration ofPTP into the brain is not the underlying mechanism of
esistance in the latter strain. Rather the data are consis-ent with a resistance mechanism in SWR involving limita-ion of nerve terminal damage and/or attenuation of thentermediate transcriptional response. In contrast, inax�/� mice there is a robust intermediate response
mplying a different mode of MPTP resistance where dam-ge to DA synaptic terminals, inflammation and astroglio-is in the striatum are tolerated by the DAergic SNpceurons.
The early response has two general features. First, aubset of genes is induced to similar levels in all brainegions examined. Most of these genes have been impli-ated in responses to oxidative stress and include Cdkn1aHershenson, 2004; O’Reilly, 2005), Gadd45g (Edwards etl., 2004) and Fkbp5 (Grigoryev et al., 2006) implyingidespread metabolic compromise, presumptively attribut-ble to inhibition of mitochondrial respiration by MPP�
Kindt et al., 1987; Krueger et al., 1990; Bates et al., 1994;asegawa et al., 1995). These findings are consistent withn extensive literature showing oxidative stress and cyto-ine signaling responses in the MPTP model (Beal, 2003;rzedborski and Vila, 2003; Hald and Lotharius, 2005;
Saline MPTP
Bax +/+
ig. 8. The intermediate response in Bax knockout mice does not diffnd wild type littermates were injected with either MPTP (20 mg/kg) ot 24 h, and total RNA isolated and hybridized onto the Affymetrix Mou
dentified in C57BL/6J mice (Fig. 1 and Supplemental Table 1) was comPTP to generate this heat map. Each vertical column represents a s
s an individual probe set. Probe sets that are upregulated in treated con green. The relative log2 (ratio) is reflected by the intensity of the colond two male Bax�/� mice were treated with saline and five malexperiments. The transcriptional response did not differ between the
ackson-Lewis and Smeyne, 2005; Hald et al., 2007; p
chapira, 2008). Second there is a striatum-enriched com-onent of the early response involving a larger number ofenes whose levels of expression change in the ordertriatum�cerebral cortex��cerebellum. For example,any immediate-early genes are induced to lesser extents
e.g. Fosb, Junb) or not at all (e.g. c-Jun) in cortex anderebellum when compared with striatum. A number ofdditional genes linked to oxidative stress responses suchs Mt2, Ucp2, Gadd45b and NFkBia are also increased toreater extents in striatum, suggesting that this structureay experience additional metabolic stress. MPP� prefer-ntially accumulates in DA nerve terminals and elicitsumping of DA (Chiba et al., 1985; Rollema et al., 1988), aotent oxidant as well as a receptor ligand. Therefore, thetriatum selective responses may be the result of dopa-ine receptor activation and local oxidative damage
aused by dopamine. The finding of elevated Fosb intriatal neurons in the MPTP model (Perez-Otano et al.,998) is consistent with dopamine receptor interactionsediating some of the early responses. Furthermore, theon-receptor-mediated induction of Hmox1 by dopaminebut not other catecholamines) in glia (Schmidt et al., 1999)lso supports a local oxidative stress model.
Some of the early mRNA changes may also be trig-ered as part of the central stress response to the periph-ral toxicity of MPTP. Indeed many stressors trigger ex-
Saline MPTP
Bax -/-
hat in wild type littermates. Bax null mice on a C57BL/6J backgroundontrol) every 2 h for a total of four injections. Striatum was harvestedme 430 2.0 array. The expression profile of the probe sets previouslya cohort of Bax�/� and wild type littermates 24 h after treatment withuse treated as indicated on top of the figure and each horizontal rowwith control mice appear in red, those that are downregulated appearl of 20 animals (three female and three male wild type and two femaleh genotype were treated with MPTP) were used to perform thesetypes, although Bax�/� mice are resistant to MPTP.
er from tr saline (cse Genopared iningle momparedr. A totas of eac
ression of immediate-early genes in the brain (Morgan
adsbiam2skctim(sttto
rirToaNa2eMoawrgwbrpgpa1t
tbPtAF
rAsfPRmare presented as mean�S.E.M. of six wild type and four Bax�/� micetreated with saline and five MPTP-treated mice of both strains.
Dt*biB
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1187
nd Curran, 1989; Schreiber et al., 1991; Liu et al., 2001;e Kloet et al., 2005; Grottick et al., 2005). Moreover,everal genes identified in the early response are known toe responsive to stress/steroid hormone signaling, includ-
ng Sgk, and Sgk3 and Tsc22d1 and Tsc22d3 (Leung etl., 2003) and some have been implicated in the MPTPodel of PD (Schoenebeck et al., 2005; Stichel et al.,005). Similarly, Ddit4 which is implicated in the mTOR-ignaling pathway and whose levels are increased in Par-inson brain (Malagelada et al., 2006) and in a 6OHDAellular model of PD (Malagelada et al., 2006) increases inhe early phase. Interestingly, we observed not only anncrease of Ddit4 mRNA levels, but also a decrease in
RNA for the related gene, Ddit4l in the same time periodSupplementary Table 1). As these changes occur in bothensitive and resistant strains of mice, their relevance tohe pathology of PD is questionable. Nonetheless, in ordero understand their relevance, it will be important to ascer-ain whether these reciprocal changes in gene expressionccur in the same cell population.
The intermediate phase is characterized by the over-epresentation of transcripts implicated in cytokine signal-ng, inflammatory responses, activation of astrocytes andesponses to cellular stress and damage (Supplementalables 1 and 6). Many studies have reported the presencef inflammatory responses in striatum in both PD andnimal models of the disease (Hald and Lotharius, 2005;agatsu and Sawada, 2005; Hald et al., 2007; Hunter etl., 2007; McGeer and McGeer, in press; Wilms et al.,007) as well as astrocytosis and glial activation (McGeert al., 1988; Francis et al., 1995; Kohutnicka et al., 1998;cGeer and McGeer, in press). Besides gene productsvertly linked to inflammation, gliosis, and cellular damagend stress responses, a number of other signaling path-ays are also apparently altered during the intermediate
esponse to MPTP. For example, levels of both CD9 anti-en (Cd9) and Hbegf increase. As CD9 antigen associatesith and activates the membrane bound form of heparin-inding EGF (Iwamoto et al., 1994) this coordinate up-egulation is expected to increase signaling through thisathway. There may also be changes in the bone morpho-enetic protein and transforming growth factors signalingathways following MPTP treatment as there are alter-tions in the mRNA levels of bone morphogenetic protein(Bmp1) and BMP2 inducible kinase (Bmp2k) as well as
he transforming growth factor, beta receptor II (Tgfbr2).With a few notable exceptions, by 72 h post-treatment
he majority of the mRNA changes seen at 24 h return toasal levels and a new cohort of transcripts is altered.rominent, among the persistently altered mRNAs are
hose linked to gliosis, inflammation and oxidative stress.s these same gene expression changes are evident at
ifferences were analyzed with one-way ANOVA and Bonferroni post-est and significance values are indicated with asterisks (*** P�0.001,* P�0.01, * P�0.05). Results are in accordance with those obtainedy Affymetrix gene chip technology and show no significant difference
ig. 9. Quantitative assessment of mRNA levels in the intermediateesponse in Bax�/� and wild type littermate mice by qRT-PCR.nimals of both strains were injected with either MPTP (20 mg/kg) oraline (control) every 2 h for a total of four injections. Levels of mRNAor candidate genes of the intermediate transcriptional phase (Hmox1,dlim4, Edg3, Tnfrsf12a and Hbegf) were evaluated by quantitativeT-PCR. Results are expressed as the number of copies of a specificRNA normalized versus the number of copies of Gapdh mRNA. Data
n the expression levels of selected transcripts between wild-type andax�/� mice.

9iowl7
ttai(aao3(aZecsjc(hmas
tMdacdapeiaddirsdmlwtfyt(7tmK
cn(gvoBctmg(ka2hmasersA(fectdgtgtmlt
ipaspit1a2PggWbstnept
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941188
6 h, this provides evidence for an ongoing and persistentnflammatory response in striatum that initiates within 24 hf MPTP treatment. Nevertheless, the majority of geneshose expression is altered at 24 h have returned to basal
evels and another cohort of mRNA changes is evident at2 and 96 h.
The biological functions of genes that alter uniquely inhe late phase are diverse and potentially represent adap-ive responses occurring in neurons and oligodendrocytess well as astrocytes and microglia. For example, changes
n parvalbumin and solute carrier family 6, member 11Slc6a11, a GABA transporter) expression imply alter-tions in striatal GABAergic interneurons (Kawaguchi etl., 1995; Marin et al., 2000) whereas changes in myelinligodendrocyte glycoprotein (Mog), 2=,3=-cyclic nucleotide= phosphodiesterase (Cnp) and proteolipid protein 1Plp1) indicate responses in oligodendrocytes (Scolding etl., 1989; Hardy and Friedrich, 1996; Jakovcevski andecevic, 2005). A number of mRNAs in the late phasencode membrane or secreted proteins involved in inter-ellular communication and extracellular matrix functionuch as neural cell adhesion molecule 1 (Ncam1), gap
unction membrane channel protein alpha 1 (Gja1 a.k.a.onnexin 43), secreted acidic cysteine rich glycoproteinSparc), secreted phosphoprotein 1 (Spp1) and tissue in-ibitor of metalloproteinase 4 (Timp4). These responsesay reflect the process of synaptic terminal eliminationnd remodeling as might changes in mRNA levels for theynaptic protein, bassoon (Bsn).
The mRNA levels of a number of genes identified inhis analysis have been reported to change in variousPTP models. However, as the models diverge in terms ofosing regimens, brain regions studied, time courses ex-mined as well as microarray/analytical platforms, statisti-al criteria and sample size used, we cannot readily makeirect comparisons, although we can highlight similaritiesmong the present analysis and previously published re-orts. Here we identify 443 genes of which 88 belong to thearly response (68 increased and 20 reduced), 226 to the
ntermediate response (199 increased and 27 reduced)nd 170 to the late response (108 increased and 62 re-uced). Moreover, we have chosen time points when celleath in the SNpc has not yet started (early response), is
n its infancy (intermediate response) or is advanced (lateesponse). When we compared previously published re-ults obtained in striatum of MPTP-treated mice with ourataset we observed relatively little overlap. Using cDNAicroarray techniques, Grünblatt and colleagues (Grunb-
att et al., 2001; Mandel et al., 2002) identified 51 geneshose levels were modulated 8 days after the first MPTP
reatment in the striatum, only one of which, solute carrieramily 6, member 11 (Slc6a11), was detected in our anal-sis (elevated at 3 days post treatment). The analysis ofhe striatal response to MPTP performed by Miller et al.2005) using Affymetrix arrays (U74A v2) revealed 178 and16 genes modulated at 8 and 15 days, respectively, afterhe first MPTP injection. Of these genes, only 25 wereodulated in a similar fashion in our study (early response
it ligand (Kitl), Mt2 and serine/arginine-rich protein spe- eific kinase 1 (Srpk1); intermediate response AMP deami-ase 3 (Ampd3), cytochrome b-245, alpha polypeptideCyba), C1qb, Mt2 and Osmr; late response angiotensino-en (Agt), cathepsin S (Ctss), human immunodeficiencyirus type I enhancer binding protein 2 (Hivep2), myelinligodendrocyte glycoprotein (Mog), ribonuclease T2A and
(Rnaset2a and Rnaset2b), Rho-associated coiled-coilontaining protein kinase 2 (Rock2), secreted phosphopro-ein 1 (Spp1) and tropomyosin 1, alpha (Tpm1) and inter-ediate/late Apod, aquaporin 4 (Aqp4), C1qc, Gfap, lectin,alactose binding, soluble 1 (Lgals1), metallothionein 1Mt1), S100a6 and S100a13 and TYRO protein tyrosineinase binding protein (Tyrobp)). In a recent study (Chin etl., 2008) using a similar dosing paradigm (15 mg/kg everyh, four injections) and the same Affymetrix chip used
ere, Chin and coworkers identified 181 genes whoseRNA is changed in the striatum of C57BL/6J mice 7 daysfter MPTP treatment. Ten genes identified in the lattertudy were also detected in our analysis: eight genes werelevated 7 days post-MPTP treatment and were similarlyegulated in the intermediate and late responses in ourtudy (Osmr and Serpina3n, both elevated at 24 h; andTP-binding cassette, sub-family A (ABC1), member 1
Abca1), aquaporin 4 (Aqp4), Gfap, Ly86, transcriptionactor 7-like 2, T-cell specific, HMG-box (Tcf7l2) and Vimlevated at both 24 and 72 h). The remaining two genes inommon were downregulated (protein tyrosine phospha-ase 4a3 (Ptp4a3) and ryanodine receptor 1 (Ryr1), bothecreased at 72 h). Despite the relatively low overlap, theenes consistently identified by all studies suggests long-erm changes in processes such as inflammation, astro-liosis and protein trafficking. Our analysis indicates thathese processes are initiated within the first 24 h of treat-ent. Therefore, strategies aimed at ameliorating damage
ikely need to target early events that couple the insult tohe pathological responses.
The early response to MPTP treatment in striatumnvolves changes in expression levels of many genes im-licated in transcriptional regulation (approximately 21%)nd is replete in immediate-early gene transcription factorsuch as Egr3 and Egr4, Fos, Fosb, Jun and Junb (Sup-lemental Table 1). In addition to transient increases in
mmediate-early gene expression there are increases inranscriptional repressors such as Bach1 (Oyake et al.,996), Btg3 (Ou et al., 2007) and Zbtb16 (Costoya, 2007)s well as putative activators such as Klf9 (Zhang et al.,003) and transient decreases in others such as Rxrg andax8. The implication is that these alterations in turn trig-er subsequent changes in expression of other targetenes, such as those in the intermediate response.hereas the acute response occurs to the same extent in
oth sensitive and resistant strains, the intermediate re-ponse is much attenuated in SWR mice, suggesting thathe first is not causative of the second. However, we can-ot exclude the possibility that the early responses aressential but not sufficient to trigger the intermediatehase response. In this scenario the mechanism of resis-ance in the SWR strain could involve the uncoupling of the
arly transcriptional response from the intermediate re-
sctaepiccs1vwtgcMaasaptet3gdsFesampottts
goitsmwlclg9ssBLmk
LseabiTiaittat(traoi(tiwsa
CcMaamM(atlm(liew(a(ammi
fise(n(
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1189
ponse. The late response is also characterized byhanges in expression of several transcription factors. No-ably, the immediate-early gene transcription factors, Egr3nd Fos-like antigen 2 (Fosl2) that are up-regulated in thearly response are actually down-regulated in the latehase. This is reminiscent of the behavior of c-Fos follow-
ng seizures, where its levels first increase and then de-line to below basal values, at which point the gene be-omes un-responsive or refractory to re-induction by sub-equent challenges with chemoconvulsants (Morgan et al.,987). Levels of mRNA for the transcription factor, etsariant gene 5 (Etv5) are also decreased in the late phasehereas levels of the transcriptional regulators activating
ranscription factor 6 (Atf6), nuclear receptor subfamily 2,roup F, member 2 (Nr2f2) and zinc finger protein of theerebellum 1 (Zic1) are increased. This again implies thatPTP elicits coordinated transcriptional cascades in stri-tum that are correlated with pathology. Like MPTP, meth-mphetamine also causes damage to DAergic synapses intriatum (O’Callaghan and Miller, 1994). Using a cDNArray platform Cadet et al. (2001) showed that metham-hetamine treatment elicited a rapid (2–4 h) increase inhe levels of many mRNAs. As in the MPTP model, thisarly component was enriched in transcripts encodingranscription factors/DNA binding proteins (33.6% and5.4%, respectively) many of which were immediate-earlyenes. Although a direct comparison is limited by theifferences in platform and strains of mice (CD1) used,everal genes including c-Jun, c-Fos, Pax8, JunB andosB are in common with our dataset. This suggests thearly component may be part of a common striatal re-ponse to synaptic impairment/damage. The same studylso reported gene expression changes at 16 h post-treat-ent, a time intermediate between the 5 and 24-h timeoints investigated here. Comparing their 16 h dataset withur 24 h dataset revealed only three gene products, Ca-hepsin D, GADD45 and Stat3 to be in common. However,he time differences between the studies do not enable uso conclude whether or not methamphetamine elicits theame intermediate response as MPTP.
Although we determined the temporal relationships ofene expression changes in striatum in response to MPTPur methods do not have cellular resolution thereby limiting
nterpretation of signaling cascades, i.e. we cannot provehat any two changes in gene expression occur in theame cell. Nevertheless, valuable information can beined from the data regarding potential signaling path-ays activated by MPTP. To identify transcriptional regu-
ators in the early response that potentially contribute tohanges in the intermediate response, we used the Mo-
ecular Signature Database (MSigDB, www.broad.mit.edu/sea/msigdb/msigdb_index.html) (Supplemental Tables 8,and 10). The most significant transcription factor binding
ite associations to genes during the intermediate re-ponse include JunD, Nrf2, Stat1 and Stat3, Bach1 andach2, and members of the NFkB, AP-1 and E2F families.evels of mRNAs for both STATs increased in the inter-ediate response and presumptively contribute to cyto-
ine signaling associated with the inflammatory response. b
evels of Bach1 mRNA increase in the early phase. Bachignaling has been implicated in regulation of Cdkn1axpression (Shim et al., 2006) that is ubiquitously andcutely up regulated by MPTP. In addition, putative Bach1inding sites are present in several genes identified in the
ntermediate response, including Gfap, S100a10 andnfrsf12a. Moreover, induction of Hmox1 requires NRF2 to
nactivate the transcriptional repressor Bach1 (Reichard etl., in press). Putative Elf1 binding sites are also enriched
n the intermediate phase. Elf1 is an Ets-related transcrip-ion factor that can associate with other transcription fac-ors, such as AP-1 complexes (Bassuk and Leiden, 1995)nd has been implicated in gene regulation in many con-exts, most notably immune and inflammatory responsesSerdobova et al., 1997; Gallant and Gilkeson, 2006). Pu-ative Elf-1 sites are present in genes encoding C1qa, Fceceptor, IgE, high affinity I, gamma polypeptide (Fcer1 g)nd Vim that increase in the intermediate response. More-ver, Vim also harbors an AP-1 site, members of which
ncrease in both the early (e.g. Fos, Junb) and intermediatec-Jun) responses opening the possibility for co-regulationhrough Elf1-AP-1 associations. An Elf1 site is also presentn Aif1 (a.k.a. Iba1) a marker for microglia (Ito et al., 1998)hose expression also increases in the intermediate re-ponse. Thus, Elf1 may contribute to both microgliosis andstrocytosis in the MPTP model.
The relative resistance to MPTP in SWR versus57BL/6J mice is a polygenic trait (Cook et al., 2003) thatould be attributable to differences in both the basal andPTP modulated levels of gene expression as well asmino acid polymorphisms. In striatum alone 333 genesre differentially expressed between the strains (Supple-ental Table 2) and several could potentially contribute toPTP resistance. For example, superoxide dismutase 2
Sod2) has been implicated in oxidative stress responsesnd Comt contributes to dopamine metabolism. Of the 333ranscripts, 12 (e.g. Agxt2l1, ApoD, Msr2) are also regu-ated by MPTP and belong almost exclusively to the inter-
ediate and late phases that are attenuated in SWR miceSupplemental Table 11). Notably, a number of the regu-ated genes, such as C1qc and Msr2 are likely expressedn microglia and are reduced in abundance in SWR miceven under basal conditions. Furthermore, another geneith reduced expression in SWR mice, CD34 antigen
Cd34) has been associated with microgliosis (Ladeby etl., 2005) whereas the complement antagonist Cd59aQian et al., 2000) that attenuates damage in experimentalllergic encephalitis (Mead et al., 2004) is elevated in SWRice. This could imply intrinsic functional differences inicroglia between the strains that warrant further analysis
n the MPTP model.A previous quantitative trait loci (QTL) analysis identi-
ed a region of chromosome 1 (mptp1) that showed aignificant association with the strain dependent differ-nces in MPTP sensitivity in SWR and C57BL/6J miceCook et al., 2003). We identified three genes (Kcnj10,uclear VCP-like (Nvl) and signal recognition particle 9Srp9)) in the mptp1 locus that are differentially expressed
etween the strains (Supplemental Table 12 and Fig. 9). In
aamd
maueamdwtaist1Memsir
pwstAPS7hm
(s2s2pSaorsG12e(tnNite
CftiPaise
ttTpapeausPbK2betDpgom
AfCrspCag
A
A
A
A
A
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941190
ddition, mRNA levels for another six genes in the locuslter following MPTP treatment in C57BL/6J mice (Supple-ental Table 13). These genes therefore become candi-ates for more detailed analysis.
The results in SWR mice indicate that both the inflam-atory response and gliosis seen in C57BL/6J mice isttenuated in the resistant strain. However, this is not aniversal mechanism of MPTP resistance as Bax�/� micexhibit a robust intermediate response that is qualitativelynd quantitatively indistinguishable from wild type litter-ates. In addition, Bax�/� mice have very few intrinsicifferences in their basal striatal mRNA profiles comparedith wild type littermates. Of the differentially expressed
ranscripts, only the elevated levels of huntingtin-associ-ted protein 1 (HapI) mRNA in Bax�/� mice has any overt
mplications for neurodegeneration. HapI can bind andequester polyglutamine-expanded proteins such as Hun-ingtin thereby antagonizing aggregate formation (Li et al.,995). Therefore, it is conceivable that this leads to greaterPTP resistance in Bax�/� mice. An alternative hypoth-sis is that Bax resistance lies downstream of the inflam-atory response, possibly in the SNpc DA neurons them-
elves, making them tolerant of the insult. These possibil-ties could be tested using a floxed Bax allele and a Creecombinase targeted to DA neurons.
The present data provide insight into the biological andathological processes triggered by MPTP treatment asell as the genes and mechanisms that might contribute toensitivity to this neurotoxin. However, the ultimate goal ofhe study is to identify genetic factors that contribute to PD.nalysis of our datasets for genes that have been linked toD, identified three candidates: Gpr37 (a.k.a. PaelR) andnca that are increased and decreased, respectively, at2 h post-MPTP treatment, and Pink1 that is expressed atigher levels in the MPTP-sensitive C57BL/6J strain ofice.
Polymorphism of Snca have been linked with familialPolymeropoulos et al., 1997; Kruger et al., 1998) andporadic forms of PD (Chiba-Falek et al., 2006; Ahn et al.,008). Although the role of Snca in PD has been exten-ively reviewed (Dauer and Przedborski, 2003; Cookson,005; Schapira, 2006; Wood-Kaczmar et al., 2006), itshysiological and pathophysiological roles remain elusive.everal functions have been ascribed to this protein suchs: activation of microglia (Su et al., in press), modulationf glutamatergic (Gureviciene et al., 2007) and DAergicelease (Abeliovich et al., 2000) and regulation of NFkBignaling pathway (Yuan et al., in press). The orphan-protein-coupled receptor, Gpr37 (Marazziti et al., 1997,998) is a known substrate for Parkin (Imai et al., 2003,007), another gene linked to familial forms of PD (Kitadat al., 1998). Gpr37 has been implicated in DA metabolismImai et al., 2007) as well as in the death of DA neurons inhe SNpc (Kitao et al., 2007). Pink1 (PTEN-inducible ki-ase 1) is a serine/threonine protein kinase (Unoki andakamura, 2001) localized in the cytoplasm (Haque et al.,
n press) and mitochondria (Valente et al., 2004). Muta-ions in Pink1 segregate with familial forms of PD (Valente
t al., 2004; Albanese et al., 2005; Bonifati et al., 2005;ookson, 2005). Pink1 has been reported to protect cellsrom apoptosis (Petit et al., 2005) and neurons from MPTPoxicity (Haque et al., in press). Although its genetic elim-nation does not cause DA neuron loss (Zhou et al., 2007),ink1 knockout mice display altered striatal DA releasend synaptic plasticity (Kitada et al., 2007). Its lower levels
n SWR mice seems at odds with the suggested pro-urvival effect of Pink1 in PD (Abeliovich, 2007; Pridgeont al., 2007).
A major question raised by this study is to what extenthe risk for, and/or progression of PD may be influenced byhe transcriptome of the striatum versus that of the SNpc.his issue is especially relevant as a number of generoducts linked to PD are broadly expressed in the brainnd it is sometimes unclear how they result in the specificattern of neurodegeneration seen in this disease. Forxample, Pink1 is widely expressed in brain (Taymans etl., 2006) and how disruption of its function leads to PD isnclear. Hence changed Pink1 expression or function intriatum might be critical in PD. Alpha-synuclein and thearkin substrate, Gpr37 are also broadly expressed inrain (Jakes et al., 1994; Zeng et al., 1997; Li et al., 2002;ingsbury et al., 2004) as is Parkin itself (Stichel et al.,000). However, Gpr37 and Snca are components of Lewyodies (Arima et al., 1999; Ishizawa et al., 2003; Murakamit al., 2004; Wakabayashi et al., 2007) and it is most likelyhat their pathophysiological roles are exerted in SNpcAergic neurons. To resolve the question of the patho-hysiologically relevant sites of expression of susceptibilityenes for PD it will be necessary to selectively alter levelsf their transcripts in either SNpc or striatum by geneticeans.
cknowledgments—The authors would like to thank Dr. D. Soleckior critical reading of this manuscript. We also thank the Hartwellenter for Bioinformatics and Biotechnology for the synthesis of
eal-time PCR primers and probes, sequencing and for processingamples for Affymetrix GeneChip technology. This work was sup-orted in part by the National Institutes of Health Cancer CenterORE grant CA 21765, the American Lebanese Syrian Associ-ted Charities (ALSAC) and the National Institutes of Healthrants R01-NS042828 and R01-ES010772 to J.I.M.
REFERENCES
beliovich A (2007) Parkinson’s disease: pro-survival effects ofPINK1. Nature 448:759–760.
beliovich A, Schmitz Y, Farinas I, Choi-Lundberg D, Ho WH, CastilloPE, Shinsky N, Verdugo JM, Armanini M, Ryan A, Hynes M,Phillips H, Sulzer D, Rosenthal A (2000) Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopaminesystem. Neuron 25:239–252.
gani FH, Pichiule P, Chavez JC, LaManna JC (2000) The role ofmitochondria in the regulation of hypoxia-inducible factor 1 expres-sion during hypoxia. J Biol Chem 275:35863–35867.
hn TB, Kim SY, Kim JY, Park SS, Lee DS, Min HJ, Kim YK, Kim SE,Kim JM, Kim HJ, Cho J, Jeon BS (2008) alpha-Synuclein geneduplication is present in sporadic Parkinson disease. Neurology70:43–49.
lbanese A, Valente EM, Romito LM, Bellacchio E, Elia AE, Dallapi-ccola B (2005) The PINK1 phenotype can be indistinguishable
from idiopathic Parkinson disease. Neurology 64:1958–1960.
A
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
D
d
D
D
E
E
F
F
F
G
G
G
G
G
G
G
H
H
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1191
rima K, Hirai S, Sunohara N, Aoto K, Izumiyama Y, Ueda K, Ikeda K,Kawai M (1999) Cellular co-localization of phosphorylated tau- andNACP/alpha-synuclein-epitopes in lewy bodies in sporadic Parkin-son’s disease and in dementia with Lewy bodies. Brain Res843:53–61.
assuk AG, Leiden JM (1995) A direct physical association betweenETS and AP-1 transcription factors in normal human T cells. Im-munity 3:223–237.
ates TE, Heales SJ, Davies SE, Boakye P, Clark JB (1994) Effects of1-methyl-4-phenylpyridinium on isolated rat brain mitochondria:evidence for a primary involvement of energy depletion. J Neuro-chem 63:640–648.
eal MF (2003) Mitochondria, oxidative damage, and inflammation inParkinson’s disease. Ann N Y Acad Sci 991:120–131.
enjamini Y, Hochberg Y (1995) Controlling the false discovery rate: apractical and powerful approach to multiple testing. J R Stat Soc57:289–300.
enmoyal-Segal L, Soreq H (2006) Gene-environment interactions insporadic Parkinson’s disease. J Neurochem 97:1740–1755.
ianchi R, Pula G, Ceccarelli P, Giambanco I, Donato R (1992) S-100protein binds to annexin II and p11, the heavy and light chains ofcalpactin I. Biochim Biophys Acta 1160:67–75.
onifati V, Rohe CF, Breedveld GJ, Fabrizio E, De Mari M, TassorelliC, Tavella A, Marconi R, Nicholl DJ, Chien HF, Fincati E, Abbruzz-ese G, Marini P, De Gaetano A, Horstink MW, Maat-Kievit JA,Sampaio C, Antonini A, Stocchi F, Montagna P, Toni V, Guidi M,Dalla Libera A, Tinazzi M, De Pandis F, Fabbrini G, Goldwurm S,de Klein A, Barbosa E, Lopiano L, Martignoni E, Lamberti P,Vanacore N, Meco G, Oostra BA (2005) Early-onset parkinsonismassociated with PINK1 mutations: frequency, genotypes, and phe-notypes. Neurology 65:87–95.
oyd JD, Jang H, Shepherd KR, Faherty C, Slack S, Jiao Y, SmeyneRJ (2007) Response to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dine (MPTP) differs in mouse strains and reveals a divergence inJNK signaling and COX-2 induction prior to loss of neurons in thesubstantia nigra pars compacta. Brain Res 1175:107–116.
radbury AJ, Costall B, Jenner PG, Kelly ME, Marsden CD, Naylor RJ(1986) MPP� can disrupt the nigrostriatal dopamine system byacting in the terminal area. Neuropharmacology 25:939–941.
rown TP, Rumsby PC, Capleton AC, Rushton L, Levy LS (2006)Pesticides and Parkinson’s disease: is there a link? Environ HealthPerspect 114:156–164.
utte A (2002) The use and analysis of microarray data. Nat Rev DrugDiscov 1:951–960.
adet JL, Jayanthi S, McCoy MT, Vawter M, Ladenheim B (2001)Temporal profiling of methamphetamine-induced changes in geneexpression in the mouse brain: evidence from cDNA array. Syn-apse 41:40–48.
arra S, Seguin SJ, Lambert H, Landry J (2008) HSPB8 chaperoneactivity towards poly-Q containing proteins depends on its associ-ation with BAG3, a stimulator of macroautophagy. J Biol Chem283:1437–1444.
hen JY, Hsu PC, Hsu IL, Yeh GC (2001) Sequential up-regulation ofthe c-fos, c-jun and bax genes in the cortex, striatum and cerebel-lum induced by a single injection of a low dose of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in C57BL/6 mice. Neu-rosci Lett 314:49–52.
hiba-Falek O, Lopez GJ, Nussbaum RL (2006) Levels of alpha-synuclein mRNA in sporadic Parkinson disease patients. Mov Dis-ord 21:1703–1708.
hiba K, Trevor AJ, Castagnoli N Jr (1985) Active uptake of MPP�, ametabolite of MPTP, by brain synaptosomes. Biochem BiophysRes Commun 128:1228–1232.
hin MH, Qian WJ, Wang H, Petyuk VA, Bloom JS, Sforza DM, LacanG, Liu D, Khan AH, Cantor RM, Bigelow DJ, Melega WP, Camp DG2nd, Smith RD, Smith DJ (2008) Mitochondrial dysfunction, oxida-
tive stress, and apoptosis revealed by proteomic and transcrip-tomic analyses of the striata in two mouse models of Parkinson’sdisease. J Proteome Res 7:666–677.
ook R, Lu L, Gu J, Williams RW, Smeyne RJ (2003) Identification ofa single QTL, Mptp1, for susceptibility to MPTP-induced substantianigra pars compacta neuron loss in mice. Brain Res Brain Res Rev110:279–288.
ookson MR (2005) The biochemistry of Parkinson’s disease. AnnuRev Biochem 74:29–52.
ostoya JA (2007) Functional analysis of the role of POK transcrip-tional repressors. Brief Funct Genomic Proteomic 6:8–18.
auer W, Przedborski S (2003) Parkinson’s disease: mechanisms andmodels. Neuron 39:889–909.
e Kloet ER, Sibug RM, Helmerhorst FM, Schmidt MV (2005) Stress,genes and the mechanism of programming the brain for later life.Neurosci Biobehav Rev 29:271–281.
oucet JP, Nakabeppu Y, Bedard PJ, Hope BT, Nestler EJ, JasminBJ, Chen JS, Iadarola MJ, St-Jean M, Wigle N, Blanchet P,Grondin R, Robertson GS (1996) Chronic alterations in dopami-nergic neurotransmission produce a persistent elevation ofdeltaFosB-like protein (s) in both the rodent and primate striatum.Eur J Neurosci 8:365–381.
uchemin AM, Gudehithlu KP, Neff NH, Hadjiconstantinou M (1992)c-fos mRNA in mouse brain after MPTP treatment. Neurochem Int20:281–287.
berling JL, Bankiewicz KS, Jordan S, VanBrocklin HF, Jagust WJ(1997) PET studies of functional compensation in a primate modelof Parkinson’s disease. Neuroreport 8:2727–2733.
dwards MG, Sarkar D, Klopp R, Morrow JD, Weindruch R, Prolla TA(2004) Impairment of the transcriptional responses to oxidativestress in the heart of aged C57BL/6 mice. Ann N Y Acad Sci1019:85–95.
arrer MJ (2006) Genetics of Parkinson disease: paradigm shifts andfuture prospects. Nat Rev 7:306–318.
ernandez-Gonzalez A, Perez-Otano I, Morgan JI (2000) MPTP se-lectively induces haem oxygenase-1 expression in striatal astro-cytes. Eur J Neurosci 12:1573–1583.
rancis JW, Von Visger J, Markelonis GJ, Oh TH (1995) Neuroglialresponses to the dopaminergic neurotoxicant 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in mouse striatum. Neurotoxicol Teratol17:7–12.
ainetdinov RR, Fumagalli F, Jones SR, Caron MG (1997) Dopaminetransporter is required for in vivo MPTP neurotoxicity: evidencefrom mice lacking the transporter. J Neurochem 69:1322–1325.
allant S, Gilkeson G (2006) ETS transcription factors and regulationof immunity. Arch Immunol Ther Exp 54:149–163.
erman DC, Nelson EL, Liang CL, Speciale SG, Sinton CM, SonsallaPK (1996) The neurotoxin MPTP causes degeneration of specificnucleus A8, A9 and A10 dopaminergic neurons in the mouse.Neurodegeneration 5:299–312.
rigoryev DN, Liu M, Cheadle C, Barnes KC, Rabb H (2006) Genomicprofiling of kidney ischemia-reperfusion reveals expression of spe-cific alloimmunity-associated genes: Linking “immune” and “non-immune” injury events. Transplant Proc 38:3333–3336.
rottick AJ, Bagnol D, Phillips S, McDonald J, Behan DP, ChalmersDT, Hakak Y (2005) Neurotransmission- and cellular stress-relatedgene expression associated with prepulse inhibition in mice. BrainRes Brain Res Rev 139:153–162.
runblatt E, Mandel S, Maor G, Youdim MB (2001)Gene expressionanalysis in N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine micemodel of Parkinson’s disease using cDNA microarray: effect ofR-apomorphine. J Neurochem 78:1–12.
ureviciene I, Gurevicius K, Tanila H (2007) Role of alpha-synuclein insynaptic glutamate release. Neurobiol Dis 28:83–89.
ald A, Lotharius J (2005) Oxidative stress and inflammation in Par-kinson’s disease: is there a causal link? Exp Neurol 193:279–290.
ald A, Van Beek J, Lotharius J (2007) Inflammation in Parkinson’sdisease: causative or epiphenomenal? Subcell Biochem 42:
249–279.
H
H
H
H
H
H
H
H
H
H
I
I
I
I
I
J
J
J
J
J
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941192
amre K, Tharp R, Poon K, Xiong X, Smeyne RJ (1999) Differentialstrain susceptibility following 1-methyl-4-phenyl-1,2,3,6-tetrahy-dropyridine (MPTP) administration acts in an autosomal dominantfashion: quantitative analysis in seven strains of Mus musculus.Brain Res Brain Res Rev 828:91–103.
aque ME, Thomas KJ, D’Souza C, Callaghan S, Kitada T, Slack RS,Fraser P, Cookson MR, Tandon A, Park DS (2008) CytoplasmicPink1 activity protects neurons from dopaminergic neurotoxinMPTP. Proc Natl Acad Sci U S A, in press.
ardy RJ, Friedrich VL Jr (1996) Progressive remodeling of the oligo-dendrocyte process arbor during myelinogenesis. Dev Neurosci18:243–254.
asegawa E, Asagami H, Kang D, Minakami S, Takeshige K (1995)1-Methyl-4-phenylpyridinium (MPP�) inhibits mitochondrial oxy-gen consumption mediated by succinate as well as malate in ratpheochromocytoma PC12 cells. Biochem Mol Biol Int 35:409–413.
ebert G, Arsaut J, Dantzer R, Demotes-Mainard J (2003) Time-course of the expression of inflammatory cytokines and matrixmetalloproteinases in the striatum and mesencephalon of miceinjected with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, a do-paminergic neurotoxin. Neurosci Lett 349:191–195.
erkenham M, Little MD, Bankiewicz K, Yang SC, Markey SP, Johan-nessen JN (1991) Selective retention of MPP� within the mono-aminergic systems of the primate brain following MPTP adminis-tration: an in vivo autoradiographic study. Neuroscience 40:133–158.
ershenson MB (2004) p21Waf1/Cip1 and the prevention of oxidativestress. Am J Physiol 286:L502–L505.
uang Y, Cheung L, Rowe D, Halliday G (2004) Genetic contributionsto Parkinson’s disease. Brain Res 46:44–70.
unot S, Vila M, Teismann P, Davis RJ, Hirsch EC, Przedborski S,Rakic P, Flavell RA (2004) JNK-mediated induction of cyclooxy-genase 2 is required for neurodegeneration in a mouse model ofParkinson’s disease. Proc Natl Acad Sci U S A 101:665–670.
unter RL, Dragicevic N, Seifert K, Choi DY, Liu M, Kim HC, Cass WA,Sullivan PG, Bing G (2007) Inflammation induces mitochondrialdysfunction and dopaminergic neurodegeneration in the nigrostri-atal system. J Neurochem 100:1375–1386.
mai Y, Inoue H, Kataoka A, Hua-Qin W, Masuda M, Ikeda T, TsukitaK, Soda M, Kodama T, Fuwa T, Honda Y, Kaneko S, Matsumoto S,Wakamatsu K, Ito S, Miura M, Aosaki T, Itohara S, Takahashi R(2007) Pael receptor is involved in dopamine metabolism in thenigrostriatal system. Neurosci Res 59:413–425.
mai Y, Soda M, Murakami T, Shoji M, Abe K, Takahashi R (2003) Aproduct of the human gene adjacent to parkin is a component ofLewy bodies and suppresses Pael receptor-induced cell death.J Biol Chem 278:51901–51910.
shizawa T, Mattila P, Davies P, Wang D, Dickson DW (2003) Colo-calization of tau and alpha-synuclein epitopes in Lewy bodies.J Neuropathol Exp Neurol 62:389–397.
to D, Imai Y, Ohsawa K, Nakajima K, Fukuuchi Y, Kohsaka S (1998)Microglia-specific localisation of a novel calcium binding protein,Iba1. Brain Res Brain Res Rev 57:1–9.
wamoto R, Higashiyama S, Mitamura T, Taniguchi N, Klagsbrun M,Mekada E (1994) Heparin-binding EGF-like growth factor, whichacts as the diphtheria toxin receptor, forms a complex with mem-brane protein DRAP27/CD9, which up-regulates functional recep-tors and diphtheria toxin sensitivity. EMBO J 13:2322–2330.
ackson-Lewis V, Jakowec M, Burke RE, Przedborski S (1995) Timecourse and morphology of dopaminergic neuronal death caused bythe neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Neu-rodegeneration 4:257–269.
ackson-Lewis V, Smeyne RJ (2005) MPTP and SNpc DA neuronalvulnerability: role of dopamine, superoxide and nitric oxide in neu-rotoxicity. Minireview. Neurotox Res 7:193–202.
akes R, Spillantini MG, Goedert M (1994) Identification of two distinct
synucleins from human brain. FEBS Lett 345:27–32.akovcevski I, Zecevic N (2005) Sequence of oligodendrocyte devel-opment in the human fetal telencephalon. Glia 49:480–491.
avitch JA, D’Amato RJ, Strittmatter SM, Snyder SH (1985) Parkin-sonism-inducing neurotoxin, N-methyl-4-phenyl-1,2,3,6-tetrahy-dropyridine: uptake of the metabolite N-methyl-4-phenylpyridine bydopamine neurons explains selective toxicity. Proc Natl Acad SciU S A 82:2173–2177.
awaguchi Y, Wilson CJ, Augood SJ, Emson PC (1995) Striatalinterneurones: chemical, physiological and morphological charac-terization. Trends Neurosci 18:527–535.
indt MV, Heikkila RE, Nicklas WJ (1987) Mitochondrial and metabolictoxicity of 1-methyl-4-(2=-methylphenyl)-1,2,3,6-tetrahydropyridine.J Pharmacol Exp Ther 242:858–863.
ingsbury AE, Daniel SE, Sangha H, Eisen S, Lees AJ, Foster OJ(2004) Alteration in alpha-synuclein mRNA expression in Parkin-son’s disease. Mov Disord 19:162–170.
itada T, Asakawa S, Hattori N, Matsumine H, Yamamura Y,Minoshima S, Yokochi M, Mizuno Y, Shimizu N (1998) Mutations inthe parkin gene cause autosomal recessive juvenile parkinsonism.Nature 392:605–608.
itada T, Pisani A, Porter DR, Yamaguchi H, Tscherter A, Martella G,Bonsi P, Zhang C, Pothos EN, Shen J (2007) Impaired dopaminerelease and synaptic plasticity in the striatum of PINK1-deficientmice. Proc Natl Acad Sci U S A 104:11441–11446.
itao Y, Imai Y, Ozawa K, Kataoka A, Ikeda T, Soda M, Nakimawa K,Kiyama H, Stern DM, Hori O, Wakamatsu K, Ito S, Itohara S,Takahashi R, Ogawa S (2007) Pael receptor induces death ofdopaminergic neurons in the substantia nigra via endoplasmicreticulum stress and dopamine toxicity, which is enhanced undercondition of parkin inactivation. Hum Mol Genet 16:50–60.
ohutnicka M, Lewandowska E, Kurkowska-Jastrzebska I, CzlonkowskiA, Czlonkowska A (1998) Microglial and astrocytic involvement in amurine model of Parkinson’s disease induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Immunopharmacology 39:167–180.
rueger MJ, Singer TP, Casida JE, Ramsay RR (1990) Evidence thatthe blockade of mitochondrial respiration by the neurotoxin 1-methyl-4-phenylpyridinium (MPP�) involves binding at the same site asthe respiratory inhibitor, rotenone. Biochem Biophys Res Commun169:123–128.
ruger R, Kuhn W, Muller T, Woitalla D, Graeber M, Kosel S, PrzuntekH, Epplen JT, Schols L, Riess O (1998) Ala30Pro mutation in thegene encoding alpha-synuclein in Parkinson’s disease. Nat Genet18:106–108.
uhn K, Wellen J, Link N, Maskri L, Lubbert H, Stichel CC (2003) Themouse MPTP model: gene expression changes in dopaminergicneurons. Eur J Neurosci 17:1–12.
adeby R, Wirenfeldt M, Dalmau I, Gregersen R, Garcia-Ovejero D,Babcock A, Owens T, Finsen B (2005) Proliferating resident mi-croglia express the stem cell antigen CD34 in response to acuteneural injury. Glia 50:121–131.
andrigan PJ, Sonawane B, Butler RN, Trasande L, Callan R, DrollerD (2005) Early environmental origins of neurodegenerative dis-ease in later life. Environ Health Perspect 113:1230–1233.
eung YF, Tam PO, Lee WS, Lam DS, Yam HF, Fan BJ, Tham CC,Chua JK, Pang CP (2003) The dual role of dexamethasone onanti-inflammation and outflow resistance demonstrated in culturedhuman trabecular meshwork cells. Mol Vision 9:425–439.
i JY, Henning Jensen P, Dahlstrom A (2002) Differential localizationof alpha-, beta- and gamma-synucleins in the rat CNS. Neuro-science 113:463–478.
i XJ, Li SH, Sharp AH, Nucifora FC Jr, Schilling G, Lanahan A, WorleyP, Snyder SH, Ross CA (1995) A huntingtin-associated proteinenriched in brain with implications for pathology. Nature 378:398–402.
inder JC, Young SJ, Groves PM (1995) Electron microscopic evi-dence for neurotoxicity in the basal ganglia. Neurochem Int
26:195–202.
L
M
M
M
M
M
M
M
M
M
M
M
M
M
M
N
N
N
O
O
O
O
P
P
P
P
P
P
Q
Q
R
R
R
R
R
R
S
S
S
R. Pattarini et al. / Neuroscience 155 (2008) 1174–1194 1193
iu PK, Grossman RG, Hsu CY, Robertson CS (2001) Ischemic injuryand faulty gene transcripts in the brain. Trends Neurosci 24:581–588.
alagelada C, Ryu EJ, Biswas SC, Jackson-Lewis V, Greene LA(2006) RTP801 is elevated in Parkinson brain substantia nigralneurons and mediates death in cellular models of Parkinson’sdisease by a mechanism involving mammalian target of rapamycininactivation. J Neurosci 26:9996–10005.
andel S, Grunblatt E, Maor G, Youdim MB (2002) Early and lategene changes in MPTP mice model of Parkinson’s disease em-ploying cDNA microarray. Neurochem Res 27:1231–1243.
arazziti D, Gallo A, Golini E, Matteoni R, Tocchini-Valentini GP(1998) Molecular cloning and chromosomal localization of themouse Gpr37 gene encoding an orphan G-protein-coupled peptidereceptor expressed in brain and testis. Genomics 53:315–324.
arazziti D, Golini E, Gallo A, Lombardi MS, Matteoni R, Tocchini-Valentini GP (1997) Cloning of GPR37, a gene located on chro-mosome 7 encoding a putative G-protein-coupled peptide recep-tor, from a human frontal brain EST library. Genomics 45:68–77.
arin O, Anderson SA, Rubenstein JL (2000) Origin and molecularspecification of striatal interneurons. J Neurosci 20:6063–6076.
cClintick JN, Edenberg HJ (2006) Effects of filtering by Present callon analysis of microarray experiments. BMC Bioinformat 7:49.
cGeer PL, Itagaki S, Boyes BE, McGeer EG (1988) Reactive micro-glia are positive for HLA-DR in the substantia nigra of Parkinson’sand Alzheimer’s disease brains. Neurology 38:1285–1291.
cGeer PL, McGeer EG (2007) Glial reactions in Parkinson’s disease.Mov Disord, in press.
ead RJ, Neal JW, Griffiths MR, Linington C, Botto M, Lassmann H,Morgan BP (2004) Deficiency of the complement regulator CD59aenhances disease severity, demyelination and axonal injury inmurine acute experimental allergic encephalomyelitis. Lab Invest84:21–28.
iller RM, Chen LL, Kiser GL, Giesler TL, Kaysser-Kranich TM,Palaniappan C, Federoff HJ (2005) Temporal evolution of mousestriatal gene expression following MPTP injury. Neurobiol Aging26:765–775.
ootha VK, Lindgren CM, Eriksson KF, Subramanian A, Sihag S,Lehar J, Puigserver P, Carlsson E, Ridderstrale M, Laurila E,Houstis N, Daly MJ, Patterson N, Mesirov JP, Golub TR, TamayoP, Spiegelman B, Lander ES, Hirschhorn JN, Altshuler D, GroopLC (2003) PGC-1alpha-responsive genes involved in oxidativephosphorylation are coordinately downregulated in human diabe-tes. Nat Genet 34:267–273.
organ JI, Cohen DR, Hempstead JL, Curran T (1987) Mappingpatterns of c-fos expression in the central nervous system afterseizure. Science 237:192–197.
organ JI, Curran T (1989) Stimulus-transcription coupling in neurons:role of cellular immediate-early genes. Trends Neurosci 12:459–462.
urakami T, Shoji M, Imai Y, Inoue H, Kawarabayashi T, Matsubara E,Harigaya Y, Sasaki A, Takahashi R, Abe K (2004) Pael-R isaccumulated in Lewy bodies of Parkinson’s disease. Ann Neurol55:439–442.
agatsu T, Mogi M, Ichinose H, Togari A (2000) Cytokines in Parkin-son’s disease. J Neural Transm Suppl 143–151.
agatsu T, Sawada M (2005) Inflammatory process in Parkinson’sdisease: role for cytokines. Curr Pharm Des 11:999–1016.
urmi E, Ruottinen HM, Bergman J, Haaparanta M, Solin O, SonninenP, Rinne JO (2001) Rate of progression in Parkinson’s disease: a6-[18F]fluoro-L-dopa PET study. Mov Disord 16:608–615.
’Callaghan JP, Miller DB (1994) Neurotoxicity profiles of substitutedamphetamines in the C57BL/6J mouse. J Pharmacol Exp Ther270:741–751.
’Reilly MA (2005) Redox activation of p21Cip1/WAF1/Sdi1: a multi-functional regulator of cell survival and death. Antioxid Redox
Signal 7:108–118.u YH, Chung PH, Hsu FF, Sun TP, Chang WY, Shieh SY (2007) Thecandidate tumor suppressor BTG3 is a transcriptional target of p53that inhibits E2F1. EMBO J 26:3968–3980.
yake T, Itoh K, Motohashi H, Hayashi N, Hoshino H, Nishizawa M,Yamamoto M, Igarashi K (1996) Bach proteins belong to a novelfamily of BTB-basic leucine zipper transcription factors that interactwith MafK and regulate transcription through the NF-E2 site. MolCell Biol 16:6083–6095.
attarini R, Smeyne RJ, Morgan JI (2007) Temporal mRNA profiles ofinflammatory mediators in the murine 1-methyl-4-phenyl-1,2,3,6-tetrahydropyrimidine model of Parkinson’s disease. Neuroscience145:654–668.
erez-Otano I, Mandelzys A, Morgan JI (1998) MPTP-Parkinsonism isaccompanied by persistent expression of a delta-FosB-like proteinin dopaminergic pathways. Brain Res 53:41–52.
etit A, Kawarai T, Paitel E, Sanjo N, Maj M, Scheid M, Chen F, Gu Y,Hasegawa H, Salehi-Rad S, Wang L, Rogaeva E, Fraser P, Rob-inson B, St George-Hyslop P, Tandon A (2005) Wild-type PINK1prevents basal and induced neuronal apoptosis, a protective effectabrogated by Parkinson disease-related mutations. J Biol Chem280:34025–34032.
olymeropoulos MH, Lavedan C, Leroy E, Ide SE, Dehejia A, Dutra A,Pike B, Root H, Rubenstein J, Boyer R, Stenroos ES, Chan-drasekharappa S, Athanassiadou A, Papapetropoulos T, JohnsonWG, Lazzarini AM, Duvoisin RC, Di Iorio G, Golbe LI, NussbaumRL (1997) Mutation in the alpha-synuclein gene identified in fam-ilies with Parkinson’s disease. Science 276:2045–2047.
ridgeon JW, Olzmann JA, Chin LS, Li L (2007) PINK1 protectsagainst oxidative stress by phosphorylating mitochondrial chaper-one TRAP1. PLoS Biol 5:e172.
rzedborski S, Vila M (2003) The 1-methyl-4-phenyl-1,2,3,6-tetrahy-dropyridine mouse model: a tool to explore the pathogenesis ofParkinson’s disease. Ann N Y Acad Sci 991:189–198.
ian YM, Qin X, Miwa T, Sun X, Halperin JA, Song WC (2000)Identification and functional characterization of a new gene encod-ing the mouse terminal complement inhibitor CD59. J Immunol165:2528–2534.
uackenbush J (2001) Computational analysis of microarray data. NatRev 2:418–427.
eichard JF, Motz GT, Puga A (2007) Heme oxygenase-1 induction byNRF2 requires inactivation of the transcriptional repressor BACH1.Nucleic Acids Res, in press.
escher U, Gerke V (2008) S100A10/p11: family, friends and func-tions. Pflugers Arch 455:575–582.
ety S, Sopkova J, Renouard M, Osterloh D, Gerke V, Tabaries S,Russo-Marie F, Lewit-Bentley A (1999) The crystal structure of acomplex of p11 with the annexin II N-terminal peptide. Nat StructBiol 6:89–95.
inne OJ, Nurmi E, Ruottinen HM, Bergman J, Eskola O, Solin O(2001) [(18)F]FDOPA and [(18)F]CFT are both sensitive PETmarkers to detect presynaptic dopaminergic hypofunction in earlyParkinson’s disease. Synapse 40:193–200.
ocke DM, Durbin B (2003) Approximate variance-stabilizing transfor-mations for gene-expression microarray data. Bioinformatics19:966–972.
ollema H, Kuhr WG, Kranenborg G, De Vries J, Van den Berg C(1988) MPP�-induced efflux of dopamine and lactate from ratstriatum have similar time courses as shown by in vivo braindialysis. J Pharmacol Exp Ther 245:858–866.
antamaria-Kisiel L, Rintala-Dempsey AC, Shaw GS (2006) Calcium-dependent and -independent interactions of the S100 protein fam-ily. Biochem J 396:201–214.
chapira AH (2006) Etiology of Parkinson’s disease. Neurology66:S10–S23.
chapira AH (2008) Mitochondria in the aetiology and pathogenesis of
Parkinson’s disease. Lancet Neurol 7:97–109.
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
T
T
T
U
V
V
W
W
W
X
Y
Z
Z
Z
S
S
R. Pattarini et al. / Neuroscience 155 (2008) 1174–11941194
chmidt J, Mertz K, Morgan JI (1999) Regulation of heme oxygen-ase-1 expression by dopamine in cultured C6 glioma and primaryastrocytes. Brain Res Brain Res Rev 73:50–59.
choenebeck B, Bader V, Zhu XR, Schmitz B, Lubbert H, Stichel CC(2005) Sgk1, a cell survival response in neurodegenerative dis-eases. Mol Cell Neurosci 30:249–264.
chreiber SS, Tocco G, Shors TJ, Thompson RF (1991) Activation ofimmediate early genes after acute stress. Neuroreport 2:17–20.
colding NJ, Frith S, Linington C, Morgan BP, Campbell AK, Comp-ston DA (1989) Myelin-oligodendrocyte glycoprotein (MOG) is asurface marker of oligodendrocyte maturation. J Neuroimmunol22:169–176.
erdobova I, Pla M, Reichenbach P, Sperisen P, Ghysdael J, WilsonA, Freeman J, Nabholz M (1997) Elf-1 contributes to the function ofthe complex interleukin (IL)-2-responsive enhancer in the mouseIL-2 receptor alpha gene. J Exp Med 185:1211–1221.
hen YQ, Hebert G, Lin LY, Luo YL, Moze E, Li KS, Neveu PJ (2005)Interleukine-1beta and interleukine-6 levels in striatum and otherbrain structures after MPTP treatment: influence of behaviorallateralization. J Neuroimmunol 158:14–25.
him KS, Rosner M, Freilinger A, Lubec G, Hengstschlager M (2006)Bach2 is involved in neuronal differentiation of N1E-115 neuroblas-toma cells. Exp Cell Res 312:2264–2278.
meyne RJ, Jackson-Lewis V (2005) The MPTP model of Parkinson’sdisease. Brain Res 134:57–66.
mith TS, Trimmer PA, Khan SM, Tinklepaugh DL, Bennett JP Jr(1997) Mitochondrial toxins in models of neurodegenerative dis-eases. II: Elevated zif268 transcription and independent temporalregulation of striatal D1 and D2 receptor mRNAs and D1 and D2receptor-binding sites in C57BL/6 mice during MPTP treatment.Brain Res 765:189–197.
riram K, Benkovic SA, Hebert MA, Miller DB, O’Callaghan JP (2004)Induction of gp130-related cytokines and activation of JAK2/STAT3 pathway in astrocytes precedes up-regulation of glial fibril-lary acidic protein in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dine model of neurodegeneration: key signaling pathway forastrogliosis in vivo? J Biol Chem 279:19936–19947.
riram K, Matheson JM, Benkovic SA, Miller DB, Luster MI,O’Callaghan JP (2006) Deficiency of TNF receptors suppressesmicroglial activation and alters the susceptibility of brain regions toMPTP-induced neurotoxicity: role of TNF-alpha. FASEB J 20:670–682.
tichel CC, Augustin M, Kuhn K, Zhu XR, Engels P, Ullmer C, LubbertH (2000) Parkin expression in the adult mouse brain. Eur J Neu-rosci 12:4181–4194.
tichel CC, Schoenebeck B, Foguet M, Siebertz B, Bader V, Zhu XR,Lubbert H (2005) sgk1, A member of an RNA cluster associatedwith cell death in a model of Parkinson’s disease. Eur J Neurosci21:301–316.
u X, Maguire-Zeiss KA, Giuliano R, Prifti L, Venkatesh K, Federoff HJ(2007) Synuclein activates microglia in a model of Parkinson’sdisease. Neurobiol Aging, in press.
ubramanian A, Kuehn H, Gould J, Tamayo P, Mesirov JP (2007)GSEA-P: a desktop application for gene set enrichment analysis.Bioinformatics 23:3251–3253.
ubramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL,Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES,Mesirov JP (2005) Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles.Proc Natl Acad Sci U S A 102:15545–15550.
undstrom E, Stromberg I, Tsutsumi T, Olson L, Jonsson G (1987)
Studies on the effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri- tdine (MPTP) on central catecholamine neurons in C57BL/6 mice.Comparison with three other strains of mice. Brain Res 405:26–38.
an EK, Yew K, Chua E, Puvan K, Shen H, Lee E, Puong KY, Zhao Y,Pavanni R, Wong MC, Jamora D, de Silva D, Moe KT, Woon FP,Yuen Y, Tan L (2006) PINK1 mutations in sporadic early-onsetParkinson’s disease. Mov Disord 21:789–793.
aymans JM, Van den Haute C, Baekelandt V (2006) Distribution ofPINK1 and LRRK2 in rat and mouse brain. J Neurochem 98:951–961.
okumitsu H, Mizutani A, Minami H, Kobayashi R, Hidaka H (1992) Acalcyclin-associated protein is a newly identified member of theCa2�/phospholipid-binding proteins, annexin family. J Biol Chem267:8919–8924.
noki M, Nakamura Y (2001) Growth-suppressive effects of BPOZand EGR2, two genes involved in the PTEN signaling pathway.Oncogene 20:4457–4465.
alente EM, Abou-Sleiman PM, Caputo V, Muqit MM, Harvey K,Gispert S, Ali Z, Del Turco D, Bentivoglio AR, Healy DG, AlbaneseA, Nussbaum R, Gonzalez-Maldonado R, Deller T, Salvi S, CortelliP, Gilks WP, Latchman DS, Harvey RJ, Dallapiccola B, AuburgerG, Wood NW (2004) Hereditary early-onset Parkinson’s diseasecaused by mutations in PINK1. Science 304:1158–1160.
ila M, Jackson-Lewis V, Vukosavic S, Djaldetti R, Liberatore G, OffenD, Korsmeyer SJ, Przedborski S (2001) Bax ablation preventsdopaminergic neurodegeneration in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. Proc NatlAcad Sci U S A 98:2837–2842.
akabayashi K, Tanji K, Mori F, Takahashi H (2007) The Lewy bodyin Parkinson’s disease: molecules implicated in the formation anddegradation of alpha-synuclein aggregates. Neuropathology 27:494–506.
ilms H, Zecca L, Rosenstiel P, Sievers J, Deuschl G, Lucius R (2007)Inflammation in Parkinson’s diseases and other neurodegenera-tive diseases: cause and therapeutic implications. Curr Pharm Des13:1925–1928.
ood-Kaczmar A, Gandhi S, Wood NW (2006) Understanding themolecular causes of Parkinson’s disease. Trends Mol Med 12:521–528.
ie X, Lu J, Kulbokas EJ, Golub TR, Mootha V, Lindblad-Toh K,Lander ES, Kellis M (2005) Systematic discovery of regulatorymotifs in human promoters and 3= UTRs by comparison of severalmammals. Nature 434:338–345.
uan Y, Jin J, Yang B, Zhang W, Hu J, Zhang Y, Chen NH (2007)Overexpressed Alpha-Synuclein Regulated the Nuclear Factor-kappaB Signal Pathway. Cell Mol Neurobiol, in press.
eng Z, Su K, Kyaw H, Li Y (1997) A novel endothelin receptortype-B-like gene enriched in the brain. Biochem Biophys Res Com-mun 233:559–567.
hang XL, Zhang D, Michel FJ, Blum JL, Simmen FA, Simmen RC(2003) Selective interactions of Klf9/basic transcription element-binding protein with progesterone receptor isoforms A and B de-termine transcriptional activity of progesterone-responsive genesin endometrial epithelial cells. J Biol Chem 278:21474–21482.
hou H, Falkenburger BH, Schulz JB, Tieu K, Xu Z, Xia XG (2007)Silencing of the Pink1 gene expression by conditional RNAi doesnot induce dopaminergic neuron death in mice. Int J Biol Sci3:242–250.
APPENDIX
upplementary data
upplementary data associated with this article can be found, in
he online version, at doi:10.1016/j.neuroscience.2008.06.064.(Accepted 20 June 2008)(Available online 8 July 2008)


![Synthesis of (2E)-2-methyl-3-(4-{[4-(quinolin-2-ylmethoxy)phenyl]sulfanyl}phenyl)prop-2-enoic acid (VUFB 20609) and 2-methyl-3-(4-{[4-(quinolin-2-ylmethoxy)phenyl]sulfanyl}phenyl)propanoic](https://img.pdfslide.net/doc/110x75/6348f97bb88fb0854f02bc13/synthesis-of-2iei-2-methyl-3-4-4-quinolin-2-ylmethoxyphenylsulfanylphenylprop-2-enoic.jpg)











![1-Phenyl-3-{4-[4-(4-undecyloxybenzoyloxy)phenyloxycarbonyl]phenyl}triazene 1-oxide](https://img.pdfslide.net/doc/110x75/6347f12809e11fdd740a83cc/1-phenyl-3-4-4-4-undecyloxybenzoyloxyphenyloxycarbonylphenyltriazene-1-oxide.jpg)


methanone](https://img.pdfslide.net/doc/110x75/634485ed6cfb3d4064093fa6/3-5-nitro-2-furyl-1-phenyl-1-h-pyrazol-4-ylphenylmethanone.jpg)











