Embed Size (px)
Citation preview

Cell Tissue Res (1994) 278:207-215 Cell&Tissue
Research �9 Springer-Verlag 1994
Distribution of p2-1ike adrenergic receptors in the cnidarian Renilla koellikeri as revealed by autoradiography and in situ hybridization Edward W. Awad *, Michel Anctil
D~partement Sciences Biologiques and Centre de Recherche en Sciences Neurologiques, Universit~ de Montr6al, C.P. 6128, Succursale de Centre-Ville, Montreal, Quebec, Canada H3C 3J7
Received: 1 October 1993 / Accepted: 25 March 1994
Abstract. Autoradiography and in situ hybridization were used to examine the histological distribution of the previously characterized fl2-1ike adrenergic receptors in- volved in the bioluminescent activity of the sea pansy RenilIa koellikeri. The use of [3H]-(_+)CGP12177 as radioligand revealed autoradiographic labelling of the re- fringent granule-filled endoderm at the base of autozooid tentacles and autozooid columns, and in the correspond- ing endoderm of siphonozooid polyps, all areas where photocytes are concentrated. The presence of excess (10 pM) unlabelled (_+)CGP12177 or atenolol in the in- cubation mixture substantially reduced total [3H]- (+_)CGP12177 labelling. Under low stringency hy- bridization washing, human /~2-adrenoceptor oligonu- cleotide probe signals were detected in granular cells lo- cated in those areas of polyp endoderm that were labelled by [3H]-(_+)CGP12177. These cells were previously shown to be distinct from, but in close proximity to pho- tocytes. No other cell or tissue type was labelled in polyps or throughout colonial tissues. The results suggest that a conserved form offl2-adrenergic receptors is present and synthesized in a unique type of endodermal cell indirectly involved in sea pansy bioluminescence control.
Key words: fi2-1ike adrenoceptors - Autoradiography - In situ hybridization - Renilla koellikeri (Cnidaria)
Introduction
Coelenterates are considered to be endowed with the most primitive nervous system in the animal kingdom (Anderson and Schwab 1982; Martin and Spencer 1983). The application of intracellular recording techniques has greatly enhanced our understanding of the physiology
* Present address: Department of Biology, Faculty of Sciences, United Arab Emirates University, P.O. Box 17551, A1 Ain, United Arab Emirates
Correspondence to: M. Anctil
of chemical synapses in these animals (see review by Anderson and Spencer /989). However, the identity of their neuroactive substances and pharmacological mech- anisms of neurotransmission are largely unknown (Mar- tin and Spencer 1983; Lesh-Laurie 1988). Yet this knowl- edge is essential for an eventual understanding of the evolutionary origin of neurochemical signalling systems.
Progress in tracking neurochemical mechanisms in coelenterates has been slow. Some evidence for the in- volvement of catecholaminergic-like mechanisms in the control of colonial behavioral responses comes from the bioluminescent sea pansy Renilla koellikeri (Cnidaria, Anthozoa). The colony is composed of two types of indi- viduals, autozooid (feeding) and siphonozooid (water- pumping) polyps. Luminescence is known to originate in photocytes (light-emitting cells) located in two dis- crete regions of autozooid endoderm and in the endoder- mal wall of siphonozooids (Morin 1974). Several lines of evidence suggested that the luminescent behaviour of R. koellikeri is a colonial response mediated by a coordinating nerve net (Parker 1920; Nicol 1955 ; Ander- son and Case 1975). Later, Anctil et al. (/982) showed that epinephrine, but not norepinephrine, induced lu- minescent responses in intact polyps of the sea pansy at low concentrations. In addition, these responses were found to be mimicked by isoproterenol and depressed by the classical /3-adrenergic blocker propranolol, thus indicating that epinephrine-induced luminescence in R. koellikeri could be mediated by a/3-adrenergic receptor. De Waele et al. (1987) and Pani and Anctil (1994) have detected these biogenic amines in the tissues of the sea pansy, using radioenzymatic and HPLC-ED techniques.
Data demonstrating the actual presence of/%adrener- gic receptors in cnidarians were not available until re- cently./32-like adrenergic receptors associated with bio- luminescent activity were characterized by radiobinding studies in polyp tissues of R. koellikeri (Awad and Anctil 1993 a). The pharmacological properties of this receptor class were different from those of the mammalian /32- adrenergic receptors. In addition, adenylate cyclase ac- tivity positively coupled to this cnidarian /3-adrenergic

208
r e c e p t o r was d e t e c t e d in m e m b r a n e p r e p a r a t i o n s ex t r ac t - ed f r o m p o l y p t issues o f this species ( A w a d a n d A n c t i l 1993b) .
A l t h o u g h the p r e s e n c e o f f i- l ike a d r e n o c e p t o r s , the i r p h y s i o l o g i c a l e f fec t a n d the i r c o u p l i n g to s e c o n d mes - sengers w e r e d e m o n s t r a t e d in R. koellikeri, i d e n t i f y i n g a ce l lu la r s u b s t r a t e f o r this r e c e p t o r r e m a i n s a key s tep in u n d e r s t a n d i n g the m o d e o f f l - ad rene rg i c c o n t r o l t h a t is e x e r t e d o n the p h o t o c y t e s o f this c o l o n i a l ~ n t h o z o a n . T h e r e f o r e , this s t udy was p e r f o r m e d to e x a m i n e the dis- t r i b u t i o n o f fi 2- l ike a d r e n e r g i c b i n d i n g sites by a u t o r a d i - o g r a p h y a n d to l oca t e p o t e n t i a l sites o f e x p r e s s i o n o f the fi 2- l ike a d r e n e r g i c r e c e p t o r gene by in s i tu h y b r i d i z a - t i on in the t issues o f this cn ida r i an .
20 nM [3H]-(__)CGP. The slides were then dipped in fresh ice-cold TRIS-HC1 buffer and washed twice for 15 min each in ice-cold buffer, followed by a brief dipping in cold (4 ~ C) H20. Slide- mounted sections were dried on a refrigerated cold plate with a stream of cold dry air and kept overnight at -20 ~ C to ensure com- pletion of the drying process.
Control sections were incubated following the same protocol described above except that the incubation was carried out in the presence of 10 gM unlabelled (_+)CGP, atenolol or trifluoperazine (all from Research Biochemical International). Atenolol, a /~ l-se- lective blocker, was previously found to act as a specific agonist on the/~2-1ike [3H]-(_+)CGP binding sites and to induce lumines- cence like the other/~-adrenergic agonists (Awad and Anctil 1993 a). In contrast, the/~l-like [3H]-dihydroalprenolol binding sites were not sensitive to atenolol, but instead bound trifiuoperazine specifi- cally (Awad and Anctil 1993a).
Materials and methods
Animals
Colonies of R. koellikeri, purchased from Marinus Inc. (Calif., USA), were kept in aerated, filtered and recirculating sea water under standard conditions (1~16 ~ C; pH 7.0-8.0; specific gravity 1.025; 12 h light:12 h dark). A total of 5 animals were processed for autoradiography and 6 animals for in situ hybridization. All experiments were performed between 10 : 00 and 16 : 00.
Autoradiography
The method described by Palacios et al. (1988) was followed with some modifications.
Preparation of autoradiograms. Liquid emulsion autoradiography was used to generate autoradiograms (Descarries and Beaudet 1983). Labelled tissue sections were manually dipped in Ilford K- 5D emulsion (Polysciences Inc.) at 40 ~ C, dried, placed in light- proof boxes containing silica gel and stored at 5 ~ C for 1 4 weeks (optimal time: 3 weeks), then developed for 5 rain in Kodak D-19 (20 ~ C), fixed for 5 min in 30 % sodium thiosulphate, washed in water for at least 10 min, mounted in Aquamount (BDH) and examined with a Zeiss Axiomat photomicroscope equipped for dark-field microscopy and Nomarski optics. Silver grain density was estimated by counting the number of grains within the area delimited by a calibrated microscope ocular grid. All measurements were made on sections subjected to identical lengths of emulsion exposure (3 weeks) and conditions of development. To obtain refer- ence slides of tissue organization (Fig. 1), tissue blocks were fixed in 2 % glutaraldehyde, postfixed in 1% OsO4, embedded in Epon, and i gin-thick sections were stained with methylene blue as de- scribed by Anctil et al. (1984).
Tissue preparation. The animals were anaesthesized by immersion in a mixture of artificial sea water and 0.37 M MgC12 (1:1, v:v) for 1 h, sliced into small pieces and washed in TRIS-HC1 buffer (50 raM, pH 7.4; 20 g/1 NaC1). Tissue pieces were incubated in a solution of 0.04% paraformaldehyde in the TRIS-HCI buffer for 30 rain, frozen in Tissue-Tek embedding medium (Miles Inc.) with dry-ice chilled isopentane (BDH), and stored in small boxes at -80~ until sectioned. Sections, 10-15 ~tm thick, were cut at -21 ~ C with a Hacker-Bright cryotome, thaw-mounted on chrom- alum gelatin-coated slides, and stored in a freezer at - 2 0 ~ for 24 h to facilitate adhesion of sections to the slides.
Receptor labelling. Preliminary experiments were conducted to de- termine optimal incubation and washing conditions. Tissue sections were preincubated in TRIS-HC1 buffer for 10 rain and then incu- bated for 1 h with [3H]-(_+)CGP12177 ([3H]-(_+)CGP, 1920 GBq/ raM, New England Nuclear), a specific //-adrenergic radioligand (Staehelin et al. 1983), at a concentration (20 nM) roughly equiva- lent to twice its KD value (10.23 nM) for the /?2-like adrenergic binding sites (Awad and Anctil 1993a). A curve was generated at different temperatures by varying the wash time for the tissue sections. After washing, the wet tissue was wiped from the slide with a glass-fibre filter (GF/C filter, 24 ram; Fisher Scientific), di- gested with Protosol (DuPont) in scintillation vials and counted. Dissociation was slow, 2 h and 30 rain being required to dissociate 50% of bound [3H]-(_+)CGP at 0~ and 40 ~ C, respectively. Therefore, a wash time of 30 rain at 0 ~ C was chosen as the optimal washing condition and was maintained throughout the experi- ments.
To determine the optimal association rate, slide-mounted sec- tions were incubated with the radioligand (20 nM) at room temper- ature for varying periods of time. A steady state occurred at around 30 min and remained stable for I h. Subsequently, tissue sections were incubated at room temperature for 1 h in the presence of
In situ hybridization
Tissue preparation. Tissue blocks from anaesthesized animals were fixed for 3 h at 4~ in PLPG fixative (Uhl et al. 1985; 1.92 g/1 Na-periodate, 0.1 M lysine, 4 % paraformaldehyde and 8 % glutar- aldehyde in 0.1 M phosphate buffer containing 25 g/1 NaC1, at pH 7.4). Fixed blocks were then washed with 0.1 M phosphate buffer and immersed at 4 ~ C in a graded series of washing buffers containing 5% (1 h), 15% (1 h ) a n d 25% (24h)sucrose. The blocks were frozen in isopentane on dry ice and stored wrapped in aluminum paper at - 8 0 ~ C.
Probe labelling. An oligonucleotide probe mixture complementary to human /~2-adrenergic receptor D N A (New England Nuclear) was used as hybridization probe. The mixture consists of 3 different 48-base long oligonucleotides complementary to 3 different regions of the receptor cDNA coding for amino acid residues 2-17, 220 236 and 257-273. These regions correspond to the amino terminus of the receptor, the amino terminus of the third cytoplasmic loop of the receptor and the carboxyl terminus of the third cytoplasmic loop of the receptor, respectively (Emorine et al. 1987).
The oligonucleotide probes were labelled with digoxigenin-I 1- dUTP-tagged nucleotide (DIG) at the 3' end by use of the Oligo- nucleotide 3' End Labelling Kit (Boehringer-Mannheim).
In situ hybridization. In situ hybridization of the probes was per- formed as described by Hopman et al. (1988) with some modifica- tions. Fixed tissue blocks were sectioned at - 2 1 ~ and thaw- mounted on chromalum gelatin-coated slides. The sections (10- 15 lain thick) were incubated for 30 min at room temperature in 10 mM TRIS buffer, pH 7.4, containing 2 mM CaC12, 2 gg/ml Pro- teinase K (Sigma Chemical Co.), in 70 and 90% ethanol (v:v) for 5 rain each and air dried. The slides were then transferred to
0
Tentacle
Pinnule
209
Oral di _ongitudinal muscle
_z �9 L)
o o 2
A
scular cavi ty
_ _ E c t o d e r m
Si h~176176176 d i
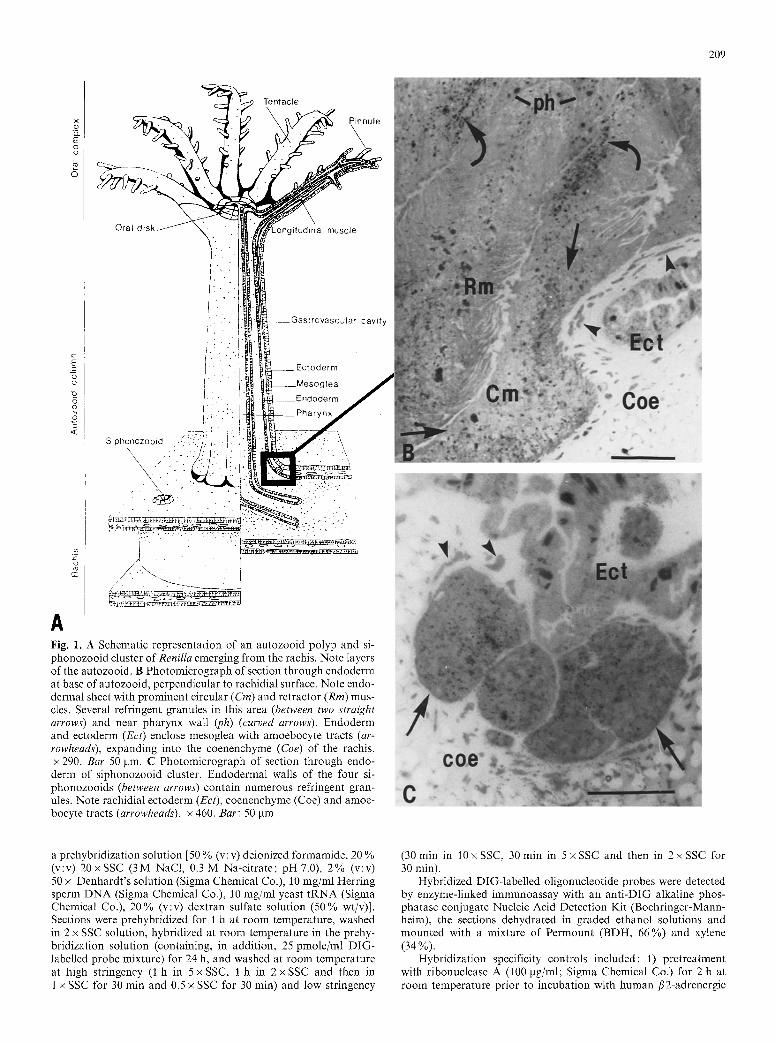
Fig. 1. A Schematic representation of an autozooid polyp and si- phonozooid cluster of Renilla emerging from the rachis. Note layers of the autozooid. B Photomicrograph of section through endoderm at base of autozooid, perpendicular to rachidial surface. Note endo- dermal sheet with prominent circular (Cm) and retractor (Rm) mus- cles. Several refringent granules in this area (between two straight arrows) and near pharynx wall ~h) (curved arrows). Endoderm and ectoderm (Ect) enclose mesoglea with amoebocyte tracts (ar- rowheads), expanding into the coenenchyme (Coe) of the rachis. x 290. Bar 50 pm. C Photomicrograph of section through endo- derm of siphonozooid cluster. Endodermal walls of the four si- phonozooids (between arrows) contain numerous refringent gran- ules. Note rachidial ectoderm (Ect), coenenchyme (Coe) and amoe- bocyte tracts (arrowheads). x 460. Bar: 50 pm
a prehybridization solution [50 % (v:v) deionized formamide, 20 % (v:v) 20xSSC (3M NaC1, 0.3 M Na-citrate; pH 7.0), 2% (v:v) 50 x Denhardt's solution (Sigma Chemical Co.), 10 mg/ml Herring sperm DNA (Sigma Chemical Co.), 10 mg/ml yeast tRNA (Sigma Chemical Co.), 20 % (v:v) dextran sulfate solution (50 % wt/v)]. Sections were prehybridized for 1 h at room temperature, washed in 2 x SSC solution, hybridized at room temperature in the prehy- bridization solution (containing, in addition, 25 pmole/ml DIG- labelled probe mixture) for 24 h, and washed at room temperature at high stringency (1 h in 5 x SSC, 1 h in 2 x SSC and then in I x SSC for 30 rain and 0.5 x SSC for 30 min) and low stringency
(30 min in 10x SSC, 30 min in 5 x SSC and then in 2x SSC for 30 min).
Hybridized DIG-labelled oligonucleotide probes were detected by enzyme-linked immunoassay with an anti-DIG alkaline phos- phatase conjugate Nucleic Acid Detection Kit (Boehringer-Mann- heim), the sections dehydrated in graded ethanol solutions and mounted with a mixture of Permount (BDH, 66%) and xylene (34%).
Hybridization specificity controls included: 1) pretreatment with ribonuclease A (100 pg/ml; Sigma Chemical Co.) for 2 h at room temperature prior to incubation with human fl2-adrenergic
receptor oligonucleotide probe labelled with DIG, 2) pretreatment of sections with excess unlabelled hybridization probe (100 pmole/ ml) of the same sense as the DIG-labelled probe and 3) exposure of sections to high temperatures (40 and 60 ~ C) after hybridization. An additional control of detection specificity was conducted by omitting the labelled probe from the hybridization solution.
211
Results
Histological organization of the colony
The histology of the colonial tissues of R. koellikeri, as described by Lyke (1965), is depicted in Fig. 1. The colony consists of two types of polyps: the autozooids which are columnar retractile polyps involved in feeding and reproduction, and the siphonozooids which are small clusters of water pumping polyps. The hydra-like autozooids bear eight pinnate tentacles at the oral end of their column and emerge from a flattened discoidal colonial mass, the rachis. The colony is anchored to the substratum by a muscular cylindrical mass called the peduncle (now shown).
The polyps show an outer cell layer (ectoderm) sepa- rated from an inner cell layer (endoderm) by an extracel- lular matrix (mesoglea) (Fig. 1). These layers constitute the wall of the autozooid column, of its tentacles, and of the invaginated pharynx. Photocytes (light-emitting cells 5-10 gm in diameter) are endodermal clusters of modified circular myoepithelial cells (Germain and Anc- til 1988) located at the sites where cells filled with re- fringent granules are concentrated (Fig. 1 B, C; Morin 1974).
Autoradiographic visualization
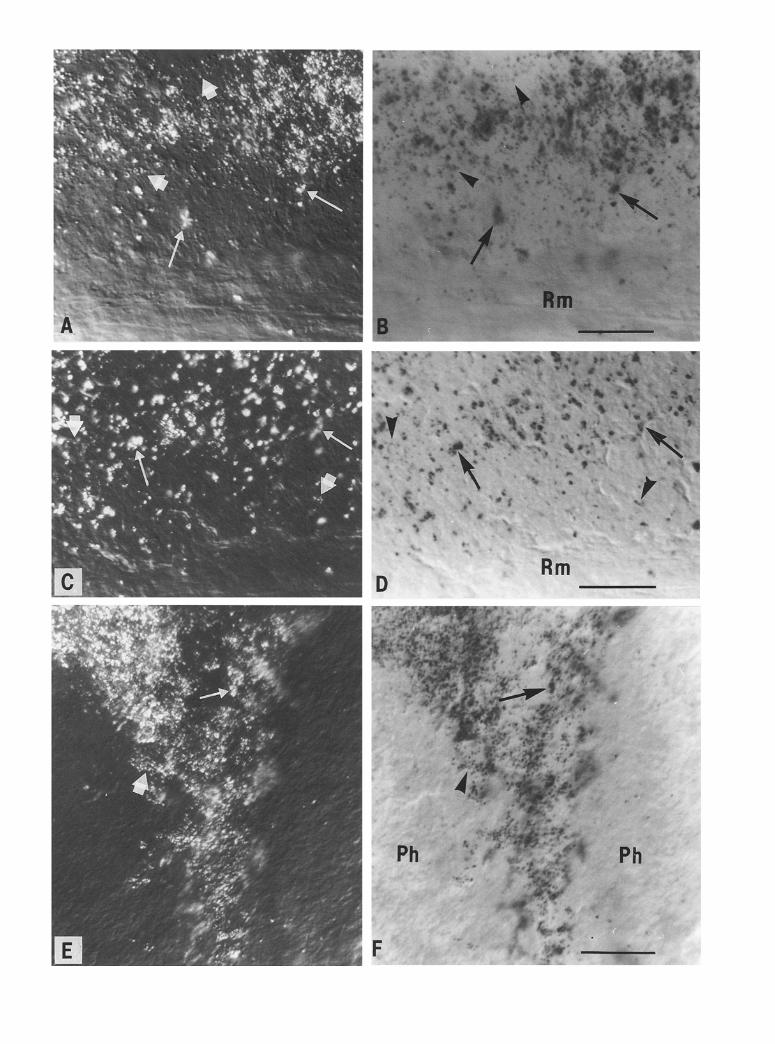
The autozooid and siphonozooid bioluminescent regions contain a large number of refringent granules in the en- dodermal layers (Fig. 2A, C, E). These endodermal granules are also scattered throughout the entire length of the autozooid column. However, their density is high- er in and around bioluminescent areas (Fig. 1 B). The high density of these endogenous granules made the vi- sualization of silver grains difficult. Silver grains were differentiated from the refringent granules by their smaller size and spherical appearance compared with the larger and more irregular shape of the granules (Fig. 2).
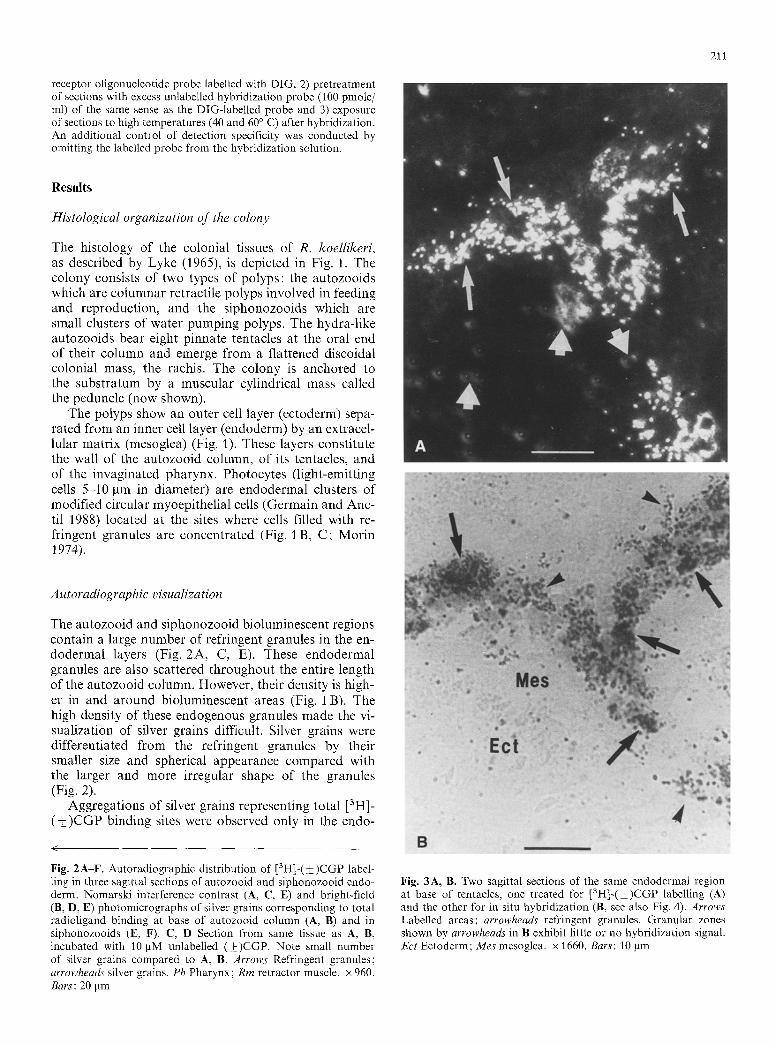
Aggregations of silver grains representing total [3H]- ( • binding sites were observed only in the endo-
<
Fig. 2A-F. Autoradiographic distribution of [3H]-(_+)CGP label- ling in three sagittal sections of autozooid and siphonozooid endo- derm. Nomarski interference contrast (A, C, E) and bright-field (B, D, E) photomicrographs of silver grains corresponding to total radioligand binding at base of autozooid column (A, B) and in siphonozooids (E, F). C, D Section from same tissue as A, B, incubated with 10 BM unlabelled (+)CGP. Note small number of silver grains compared to A, B. Arrows Refringent granules; arrowheads silver grains. Ph Pharynx; Rm retractor muscle. • 960. Bars: 20 pm
Fig. 3A, B. Two sagittal sections of the same endodermal region at base of tentacles, one treated for [3H]-(_+)CGP labelling (A) and the other for in situ hybridization (B, see also Fig. 4). Arrows Labelled areas; arrowheads refringent granules. Granular zones shown by arrowheads in B exhibit little or no hybridization signal. Ect Ectoderm; Mes mesoglea. • 1660. Bars: 10 gm
212

213
derm of autozooids, in two discrete regions, one of which is located in the lower portion of the column (Fig. 2A, B) and the other in the upper portion of the column at the base of the tentacles (Fig. 3 A). This labelling was associated with endodermal areas of the column wall joining with radial walls (septa). These areas contain cells with endogenous refringent granules and are known to include photocytes, such as the photocyte-rich areas shown in Fig. lB. Silver grain density invariably dropped to less than 20% of controls when sections were incubated with the radioligand in the presence of 10 ~tM unlabelled (+_CGP (Fig. 2C, D) or 10 ~tM ateno- lol (not shown). For example, 144.2+9.1 grains/ 1000 g m 2 w e r e counted on sections incubated with the radioligand alone and 26.8 _+ 2.4 in the presence of unla- belled (+)CGP (~_+ sem, N = 10 sections of autozooid endoderm). Addition of trifluoperazine (10 gM) to the incubation medium did not cause any noticeable change in [3H]-(-t-)CGP labelling. No specific labelling was de- tected in rachidial and peduncle tissues.
Similar aggregations of silver grains also overlaid the endoderm surrounding the pharynx of siphonozooids (Fig. 2E, F). As in autozooids, these areas contain nu- merous cells with refringent granules (Fig. 2 E) and pho- tocytes are known to be interspersed among them.
In situ hybridization
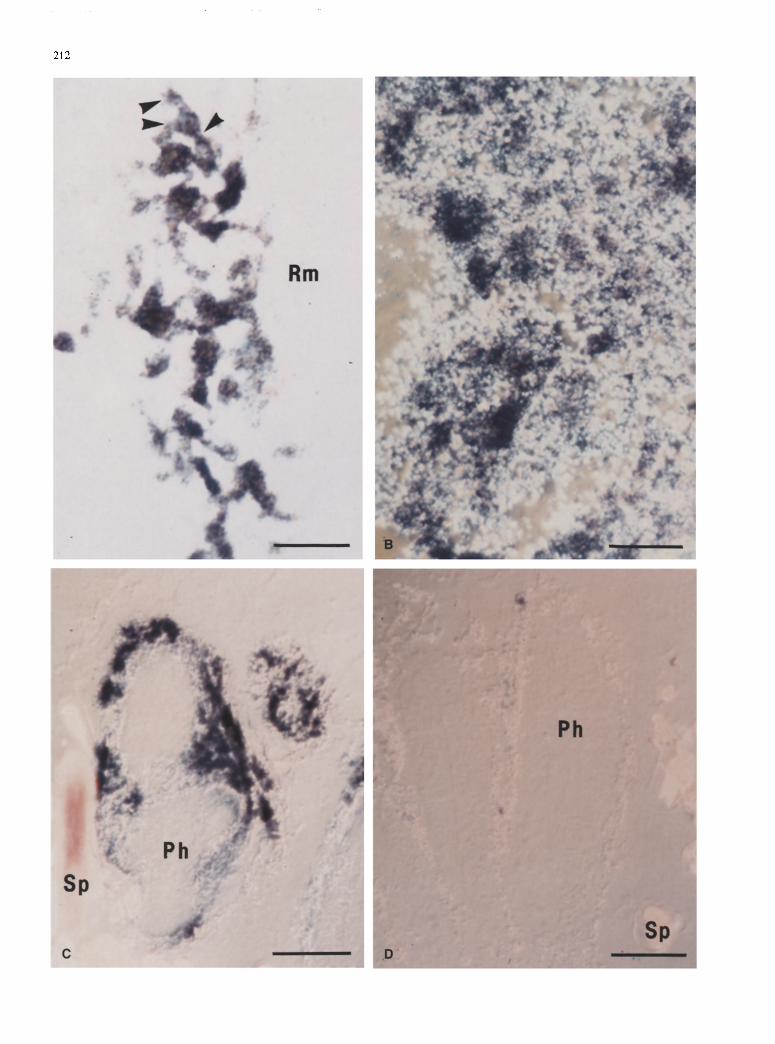
When a high stringency washing strategy was used after hybridization, very weak hybridization signals were ob- served with human /~2-adrenergic receptor probe mix- ture in the endoderm of polyps (not shown). Under low stringency, however, endodermal areas of autozooids that were labelled by [3H]-(_+)CGP also exhibited hy- bridization signals (Fig. 3). Strong signals were con- sistently observed in endodermal cells located at the base of the tentacles (Fig. 3 B), and at the base of the columns (Fig. 4A, B). The signal was mostly confined in the peri- karyon of the refringent granule-containing endodermal cells, but weaker signals were also observed in their pro- cesses (Fig. 4A). The granules vary in shape and diame- ter (1-4 gm), and they appear to be randomly distributed inside the cells (Fig. 4 B). The granular cells are multipo- lar, their perikaryon measuring between 15-20 gm in di- ameter, with stubby processes that appear to end bluntly (Fig. 4A). Similar hybridization signals were observed
<
Fig. 4A-D. Sagittal sections of autozooid and siphonozooid endo- derm treated for in situ hybridization with human/~2-adrenergic receptor oligonucleotide probe mixture. A Strong hybridization sig- nal (blue color) in meshwork of granular endodermal cells at base of autozooid column. Granules, pink color. Note processes extend- ing from granular cell (arrowheads). Rm Position of retractor mus- cle. B Dense meshwork of endodermal cells at base of autozooid column viewed by Nomarski optics to show hybridization signal over numerous refringent (white) granules. C Labelling of granular cells in endoderm of two siphonozooids (Nomarski optics). Parts of two other siphonozooids at right. D Ribonuclease A pretreat- ment eliminated hybridization signal of siphonozooid endoderm. Ph Pharynx; Sp calcareous spicules, x 1000. Bars: 20 gm
in the granular endodermal cell wall of siphonozooids (Fig. 4 C). No other cell or tissue type exhibited any spe- cific signal, even in populations of endodermal granular cells outside the photocyte-rich areas.
Sections digested with ribonuclease A showed no sig- nificant hybridization signal under low stringency wash- ing (Fig. 4D). Pretreatment of sections with excess unla- belled hybridization probe (100 pmole/ml) of the same sense as the DIG-labelled probe eliminated the hybrid- ization signals, as did omission of labelled probes from the hybridization solution. Testing for thermal stability of hybridized sections exposed to temperature values of 40 and 60 ~ C showed a complete loss of DIG labelling (not shown).
Discussion
As demonstrated by control experiments where the ra- dioligand was displaced by excess (+)CGP and atenolol but not trifiuoperazine, the localized autoradiographic labelling detected in R. koellikeri represents specifically bound [3H]-(__ )CGP. As shown earlier (Awad and Anc- til 1993a), this particular binding is characterized by a/~2-1ike adrenergic pharmacological profile. Thus the distribution of specific [3H]-(_+)CGP binding in the en- doderm of polyps which matches that of photocyte-rich areas is considered to represent labelling of/~-adrenergic receptors, here observed for the first time in the tissues of a cnidarian.
Low stringency hybridization strategy has been used to clone a variety of membrane-bound G protein-cou- pled receptors (Albert 1992). Under conditions which tolerate a certain degree of mismatch, hybridization of a cloned cDNA is used to search for related protein cDNAs encoding domains of high homology. Since se- quences homologous to human /?2-adrenergic receptor DNA are present in lower vertebrates, many inverte- brates and unicellular eukaryotes (Palacios et al. 1989), human/~2-adrenergic oligonucleotide probes may be a useful tool for the isolation and study of gene structures of homologous receptors in the sea pansy. The human /~2-adrenergic oligonucleotide probe mixture used in this study is complementary to three different mRNA se- quences coding for amino acids in three different seg- ments of the receptor, thus increasing the probability of matching the probes with Renilla mRNA encoding domains of high homology. In addition, the use of ap- propriate control experiments minimizes the possibility of non-specific hybridization.
The match between the consistently strong hybridiza- tion signals in granular endodermal cells of polyps and the distribution of silver grains corresponding to specific [3H]-(_+)CGP binding strongly suggests that these cells both contain and synthesize/?2-like adrenergic receptors associated with bioluminescent activity in this cnidarian. This match makes it unlikely that the human/~2-adren- ergic oligonucleotide probe mixture is labelling other membrane-bound G protein-coupled receptors that might be existing in the tissues of the sea pansy. This possibility exists because two of the oligonucleotide

214
probes used in the mixture for hybridization code for segments belonging to the third cytoplasmic loop of the human fi 2 adrenoceptor which is believed to be responsi- ble for the interaction of the receptor with G protein (Strosberg 1991). In addition, those regions of the third cytoplasmic loop in closest apposition to the plasma membrane are considered to be the most highly con- served cytoplasmic regions of the receptor among the different adenylyl cyclase coupled receptors that have thus far been cloned (O'Dowd et al. 1988). This possibili- ty is countered by evidence showing that activation of other G protein-related receptor types (ill adrenergic- like, serotoninergic-like) stimulates adenylate cyclase ac- tivity in rachidial and peduncle tissues (Awad and Anctil 1993b), yet neither fi2-1ike hybridization signal (in this study) nor fi2-1ike activation of adenylate cyclase (Awad and Anctil 1993 b) was detected in those tissues.
Our description of the granular endodermal cells largely corresponds with their histological description by Lyke (1965). In his study, he described a type of "epithelio-muscular" cells located in the bioluminescent regions of the sea pansy that are characterized by heavy concentrations of highly refringent granules within their cytoplasm. According to Lyke (1965), the granules (0.5- 3.0 gm in diameter) are guanine-related compounds that occur in a wide variety of shapes and may act as micro- reflectors for the emitted light. Based on his observa- tions, Lyke suggested a role in the bioluminescent reac- tion for these granular cells. However, Anctil et al. (1984) have demonstrated that the cells containing these refringent granules are distinct from, but in close prox- imity to polyp photocytes identifiable by their autofluor- escence. In addition, an ultrastructural study of disso- ciated photocytes failed to reveal the presence of inclu- sions corresponding in size or shape to these refringent granules (Germain and Anctil 1988). Therefore, the co- localization of hybridization signals described in this study and refringent granules located in cells from pho- tocyte-rich endoderm (Anctil et al. 1984) suggests that these granular cells, but not the photocytes, contain the fi2-1ike adrenergic receptors.
The existence of fi2-1ike adrenergic receptors in gran- ular endodermal cells proximal to photocytes substan- tiates previous reports on the role of fi-adrenergic mech- anisms in bioluminescence in R. koellikeri. Physiological concentrations of fl-adrenergic agonists failed to induce luminescence in isolated photocytes (Germain and Anc- til 1988). In addition, bioluminescence activation by fi- adrenergic agonists can be blocked by isotonic MgC12 in intact polyps suggesting the presence of an intermedi- ate step between the release of the adrenergic substance and the activation of luminescence (Awad and Anctil 1993 a). Together, these findings support a mediator role for granular endodermal cells in the activation of photo- cytes by the fl2-adrenergic system. Upon activation by fi-adrenergic agonists, the granular endodermal cells might release a hitherto unknown substance that acts on photocytes to induce luminescence. This indirect path for the fl-adrenergic effect on bioluminescence is analo- gous to the mediation of many vertebrate functions by fi-adrenergic receptors, where stress conditions lead to
the activation of prejunctional fi-adrenergic receptors by epinephrine released from the adrenal gland (Hieble and Ruffolo Jr. 1991). When subjected to stressful conditions (e.g., contact with a predator), the fl-adrenergic system in the sea pansy might be activated leading to the release of epinephrine in the vicinity of the granular endodermal cells. Although neurites were reported in the vicinity of the contractile tube-feet of photocytes and other myo- epithelial cells (Germain and Anctil 1988), neither evi- dence of the presence of adrenergic neurites in photo- cyte-rich endodermal regions nor ultrastructural obser- vations of the relationship between photocytes and gran- ular cells, is yet available. Once activated, the granular endodermal cells may release a "paracrine hormone" or a "final transmitter" which acts on photocytes to enhance or trigger luminescence, respectively.
The indirect circuitry of bioluminescence activation in R. koellikeri presents a striking parallel with that of photocyte activation in the hydrozoan Obelia geniculata. In the latter species, non-luminescent neighbor cells of the endoderm are also involved in activation of photo- cyte luminescence, but in this case Ca 2 + is the signalling transmitter (Dunlap et al. 1987; Brehm et al. 1989). Acti- vation of the neighbor cells opens voltage-dependent Ca 2+ channels, thus causing an influx of Ca 2+ which then enters the photocytes through gap junctions linking the two cell types. Light emission occurs when the newly interiorized Ca 2 + binds to the photoprotein obelin pres- ent in Obelia photocytes (Morin and Hastings 1971; Campbell 1974; Morin and Reynolds 1974). Thus both Renilla and Obelia depend on the mediation of neighbor- ing, non-nervous cells for luminescence control even though the mechanisms involved differ between the two cnidarian species.
In this study, we identified a unique type of endoder- real cells in a cnidarian that appears to synthesize, con- tain and use fl2-1ike adrenergic receptors to possibly con- trol bioluminescent activity. Future studies should be aimed at investigating a putative secretory role for these cells, at cloning the gene for these receptors and using them as models to understand the evolution of fl-adren- ergic receptors.
Acknowledgements. This work was funded by the Natural Sciences and Engineering Research Council of Canada (grant no. A6447 to M.A.). Other support was provided by fellowships to E.W.A. from the Facult6 des 6tudes sup6rieures, Universit~ de Montr6al and Groupe de recherche FCAR sur le syst6me nerveux central.
References
Albert PR (1992) Molecular biology of the 5HT1A receptors: low- stringency cloning and eukaryotic expression. J Chem Neuroan- atomy 5:283-288
Anctil M, Boulay D, LaRivi6re L (1982) Monoaminergic mecha- nisms associated with control of luminescence and contractile activities in the coelenterate, Renilla k6llikeri. J Exp Zool 223:11 24
Anctil M, Germain G, LaRivi6re L (1984) Catecholamines in the coelenterate Renilla k61likeri. Uptake and radioautographic lo- calization. Cell Tissue Res 238:69-80

215
Anderson PAV, Case JF (1975) Electrical activity associated with luminescence and other colonial behavior in the pennatulid Renilla k6llikeri. Biol Bull (Woods Hole) 149:80-95
Anderson PAV, Schwab WE (1982) Recent advances and model systems in coelenterate neurobiology. Progr Neurobiol 19:213- 236
Anderson PAV, Spencer AN (1989) The importance of cnidarian synapses for neurobiology. J Neurobiol 20:435-457
Awad EW, Anctil M (1993 a) Identification of fl-like adrenoceptors associated with bioluminescence in the sea pansy Renilla koel- likeri. J Exp Biol 177 : 181-200
Awad EW, Anctil M (1993 b) Positive coupling of fi-like adrenergic receptors with adenylate cyclase in the cnidarian Renilla koellik- eri. J Exp Biol 182:131-146
Brehm P, Leichleiter J, Smith S, Dunlap K (1989) Intercellular signaling as visualized by endogenous calcium-dependent biolu- minescence. Neuron 3:191-198
Campbell AK (1974) Extraction, partial purification and properties of obelin, the calcium-activated photoprotein from the hydroid Obelia geniculata. Biochem J 143:411-418
De Waele J-P, Anctil M, Carlberg M (1987) Biogenic catechol- amines in the cnidarian Renilla kgllikeri: radioenzymatic and chromatographic detection. Can J Zool 65:2458-2465
Descarries L, Beaudet A (1983) The use of radioautography for investigating transmitter-specific neurons. In: H6kfelt T, Bj6rklund J (eds) Handbook of Chemical Neuroanatomy, vol 1. Elsevier, Amsterdam, pp 286-364
Dunlap K, Takeda K, Brehm P (1987) Activation of a calcium- dependent photoprotein by chemical signalling through gap junctions. Nature 325 : 60-62
Emorine L J, Marullo S, Delavier-Klutchko C, Kaveri SV, Durieu- Trantman O, Strosberg AD (1987) Structure of the gene for the human fl2-adrenergic receptor: expression and promoter characterization. Proc Natl Acad Sci USA 84:6995-6999
Germain G, Anctil M (1988) Luminescent activity and ultrastruc- tural characterization of photocytes dissociated from the coe- lenterate Renilla kdllikeri. Tissue Cell 20:701-720
Hieble JP, Rufolo RP Jr (1991) Functions mediated by fl-adreno- ceptor activation. Prog Basic Clin Pharmacol 7 : 173-209
Hopman AHN, Raap AK, Landegent JE, Wiegant J, Boerman RH, Van der Ploeg M (1988) Non-radioactive hybridization. In: Van Leeuwen FW, Buijs RM, Pool CW, Pach O (eds) Mo- lecular neurobiology. Elsevier, Amsterdam, pp 43-68
Lesh-Laurie GE (1988) Coelenterate endocrinology. In: Laufer H, Downer RGH (eds) Endocrinology of selected invertebrates. Liss, New York, pp 3-29
Lyke EB (1965) The histology of the sea pansies, Renilla reniformis (Pallas) and Renilla k6llikeri (Pfeffer) with a note on the fine structure of the latter species. Ph. D. thesis, University of Wisconsin, Madison
Martin SM, Spencer AN (1983) Neurotransmitters in coelenterates. Comp Biochem Physiol [C] 74:1-14
Morin JG (1974) Coelenterate bioluminescence. In: Muscatine L, Lenhoff HM (eds) Coelenterate biology. Reviews and new per- spectives. Academic Press, New York, pp 397-438
Morin JG, Hastings JW (1971) Biochemistry of the biolumines- cence of colonial hydroids and other coelenterates. J Cell Phys- iol 77:305-311
Morin JG, Reynolds G (1974) The ceIlular origin of biolumines- cence in the colonial hydroid, Obelia. BioI Bull (Woods Hole) 147:397-410
Nicol JAC (1955) Nervous regulation of luminescence in the sea pansy Renilla k6llikeri. J Exp Biol 32:619-635
O'Dowd BF, Hnatowich M, Regan JW, Leader WM, Caron MG, Lefkowitz RJ (1988) Site-directed mutagenesis of the cytoplas- mic domains of the human beta2-adrenergic receptor. J Biol Chem 263:15985-15992
Palacios JM, Cartes R, Diet1 MM (1988) A laboratory guide for the in vitro labelling of receptors in tissue sections for autoradi- ography. In: Van Leeuwen FW, Buijs RM, Pool CW, Pach O (eds) Molecular neuroanatomy. Elsevier, Amsterdam, pp 95 109
Palacios JM, O'Dowd BF, Cotecchia S, Hnatowich M, Caron MG, Lefkowitz RJ (1989) Adrenergic receptor homologies in verte- brate and invertebrate species examined by DNA hybridization. Life Sci 44:2057-2065
Pani AK, Anctil M (1994) Quantitative survey of biogenic mono- amines, their precursors and metabolites in the coelenterate Renilla koellikeri. Biogen Amines 10 : 161-180
Parker GH (1920) Activities of colonial animals. II: Neuromuscu- lar movements and phosphorescence in Renilla. J Exp Zool 31:475-515
Staehelin M, Simons P, Jaeggi K, Wigger N (1983) CGP-12177: a hydrophilic beta-adrenergic receptor radioligand reveals high affinity binding of agonists to intact cells. J Biol Chem 258 : 3496-3502
Strosberg AD (1991) Structure/function relationship of proteins belonging to the family of receptors coupled to GTP-binding proteins. Eur J Biochem 196:1-10
Uhl GR, Zingg HH, Habener JF (1985) Vasopressin mRNA in situ hybridization: localization and regulation studied with oli- gonucleotide cDNA probes in normal and Brattleboro rat hy- pothalamus. Proc Natl Acad Sci USA 82 : 5555-5559















