Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7
0006-8993/$ - see frohttp://dx.doi.org/10
Abbreviations: DA
fluorescent protein;
myristate-13-acetate
deoxynucleotidyl tranCorresponding aut
E-mail address:
Research Report
Dopamine induces apoptosis in APPswe-expressingNeuro2A cells following Pepstatin-sensitive proteolysisof APP in acid compartments
Monica Cagnin, Matteo Ozzano, Natascia Bellio, Ilaria Fiorentino, Carlo Follo, Ciro Isidoron
Department of Health Sciences, Laboratory of Molecular Pathology and Nanobioimaging, Universit �a del Piemonte Orientale ‘‘A. Avogadro’’,
Via Solaroli 17, 28100 Novara, Italy
a r t i c l e i n f o
Article history:
Accepted 21 June 2012
A pathological hallmark of Alzheimer’s disease (AD) is the presence within neurons and
the interneuronal space of aggregates of b-amyloid (Ab) peptides that originate from an
Available online 6 July 2012
Keywords:
Alzheimer’s disease
Parkinson’s disease
Pepstatin A
Chloroquine
Lysosome
nt matter & 2012 Elsevie.1016/j.brainres.2012.06.0
PI, 40,6-diamidino-2-ph
IETD-CHO, acetyl-Ile-Gl
; L685,458, [(2R,4R,5S)-2
nsferase-mediated dUTPhor. Fax: þ39 0321 [email protected]
a b s t r a c t
abnormal proteolytic processing of the amyloid precursor protein (APP). The aspartyl
proteases that initiate this processing act in the Golgi and endosomal compartments. Here,
we show that the neurotransmitter dopamine stimulates the rapid endocytosis and
processing of APP and induces apoptosis in neuroblastoma Neuro2A cells over-expressing
transgenic human APP (Swedish mutant). Apoptosis could be prevented by impairing
Pepstatin-sensitive and acid-dependent proteolysis of APP within endosomal–lysosomal
compartments. The g-secretase inhibitor L685,458 and the a-secretase stimulator phorbol
ester elicited protection from dopamine-induced proteolysis of APP and cell toxicity. Our
data shed lights on the mechanistic link between dopamine excitotoxicity, processing of
APP and neuronal cell death. Since AD often associates with parkinsonian symptoms,
which is suggestive of dopaminergic neurodegeneration, the present data provide the
rationale for the therapeutic use of lysosomal activity inhibitors such as chloroquine or
Pepstatin A to alleviate the progression of AD leading to onset of parkinsonism.
& 2012 Elsevier B.V. All rights reserved.
1. Introduction
Alzheimer’s disease (AD) is a late-onset neurological disorder
characterized by progressive loss of memory and cognitive
abilities as a result of excessive neurodegeneration in the
hippocampus and cortex (Sabuncu et al., 2011). A pathological
hallmark of AD is the presence in the interneuronal space of
r B.V. All rights reserved.25
enylindole dihydrochlorid
u-Thr-Asp-aldehyde inhib
-benzyl-5-(Boc-amino)-4-h
-biotin nick end labeling;.(C. Isidoro).
amyloid plaques formed by aggregates of b-amyloid (Ab)
peptides that originate from an abnormal proteolytic proces-
sing of the amyloid precursor protein (APP). APP is a large
transmembrane type 1 (cytosolic C-terminal) glycoprotein
coded by a gene located on chromosome 21 and giving rise
to eight alternative transcripts, of which three are mainly
transcribed into the isoforms containing 695, 751 and 770
e; DA, dopamine; FITC, fluorescein isothiocyanate; GFP, green
itor (Ac-IETD-CHO); PI, propidium iodide; PMA, phorbol 12-
ydroxy-6-phenyl-hexanoyl]-Leu-Phe-NH2; TUNEL, terminal
ZVAD-fmk, benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 103
aminoacids (reviewed in Bekris et al., 2010). APP695 mature
protein differs from the whole length APP770 because it lacks
the 290–364 sequence comprising the Kunitz-Protease Inhibi-
tor peptide.
In the endoplasmic reticulum and during its transport
through the Golgi Complex, nascent APP undergoes co- and
post-translational modifications, including N- and O-glyco-
sylation, phosphorylation and tyrosine sulfation, that lead to
the so-called mature APP (Perdivara et al., 2009). Mature APP
is not permanently resident at the plasma membrane, rather
it is subjected to a continuous retrograde trafficking from the
plasma membrane to intracellular compartments of the
secretory pathway (Vieira et al., 2010), so that at steady state
it is more abundant in the Golgi Complex and in endosomes
(Koo et al., 1996; Xu et al., 1997; Yamazaki et al., 1996).
Moreover, APP is not a stable molecule, as it undergoes
proteolysis through multiple and alternative routes. The
proteolytic pathways involved in the APP processing and
the cellular compartments in which this occurs have been
studied in details (for review see Chow et al., 2010; O’Brien
and Wong, 2011; Thinakaran and Koo, 2008; Zhang et al.,
2011). The order of proteolysis at a, b and g sites determines
whether or not the Ab peptide will be produced: the sequen-
tial action of a- and g-secretases leads to the production of a
soluble APPa fragment (sAPPa), a P3 peptide and an intracel-
lular domain (AICD peptide) at the C-terminus, whereas the
sequential action of b- and g-secretases leads to a soluble
APPb fragment (sAPPb), the Ab peptide (of 40 or 42 aminoa-
cids) and the AICD peptide. Thus, proteolysis at b-site is
alternative to that at a-site and is fundamental for amyloi-
dogenesis. The main protease responsible for such proteoly-
sis is b-APP cleaving enzyme (BACE), a type-1 transmembrane
aspartyl protease mainly localized to endosomes, lysosomes
and the Golgi Complex (Cai et al., 2001; Vassar et al., 1999).
Another protease with potential b-secretase activity is lyso-
somal Cathepsin D, which has been shown able to cleave
in vitro APP and produce Ab (Chevallier et al., 1997; Higaki
et al., 1996), and to be highly expressed in AD brain
(Schechter and Ziv, 2008). However, while BACE-deficient
mice do not produce Ab and show normal phenotype (Luo
et al., 2001; Ohno et al., 2004), Cathepsin D-deficient mice still
produce and accumulate Ab in hippocampal neurons (Saftig
et al., 1996). Amyloidogenic processing of APP has been
proved to occur within the Golgi Complex (Xu et al., 1997)
and the endosomal compartment (Pasternak et al., 2004).
Impairing the internalization of plasma membrane APP
reduces the formation of Ab up to 80% (Koo and Squazzo,
1994), as it does the treatment with drugs that rise the
luminal pH of endosomal–lysosomal compartments
(Schrader-Fischer and Paganetti, 1996).
To what extent the trafficking and processing of APP in vivo
occurs constitutively or is affected by the extracellular sti-
muli, and whether and how neurotransmitters influence the
fate of APP and of cells expressing APP is largely unknown.
Here, we report on the effect of dopamine (DA), a neuro-
transmitter diffused in substantia nigra, striatum and other
brainstem nuclei, in neuroblastoma Neuro2A cells over-
expressing human APP695 (Thinakaran et al., 1996), which
is the isoform mainly expressed in human brain (Kang and
Muller-Hill, 1990). Neuro2A cells express muscarinis receptors
(Edwards et al., 1989) and are prone to cholinergic neuronal
differentiation and neurite development (Kojima et al., 1994).
Under appropriate stimulation, Neuro2A express tyrosine
hydroxylase and produce DA and L-DOPA (Akahoshi et al.,
2009) and respond to DA excitotoxicity (Castino et al., 2005).
Therefore, Neuro2A cells can be assumed bona fide as a
valuable in vitro model to study the effects of dopamine on
APP processing. The data here reported extend the previous
knowledge on the relationship between neuronal cell toxicity
and endocytosis and processing of APP, and also provide new
evidence on the mechanism of DA excitotoxicity in neuronal
cells over-expressing APP. The latter may have clinical rele-
vance, given that Parkinson’s-like dopaminergic neurodegen-
eration has been observed in the postmortem brain of AD
patients with extrapyramidal signs (Burns et al., 2005;
Jellinger, 2003; Schneider et al., 2002).
2. Results
2.1. Dopamine triggers the intrinsic apoptotic deathpathway in Neuro2A cells over-expressing transgenic HumanAPP
To address whether the abnormal expression of APP renders
dopaminergic neuronal cells susceptible to DA toxicity, we
employed an established in vitro model system represented
by neuroblastoma mouse Neuro2A cells sham-transfected or
stably expressing transgenic human APP695 in the Swedish-
mutant form (Thinakaran et al., 1996). The cells were exposed
to DA and observed under the microscope for gross morpho-
logical alterations and cell loss at increasing time of incuba-
tion. Evidence of toxic effects was noted starting at 16 h of
exposure to DA only in the transfected Neuro2A expressing
APP. By this time, nuclei staining with DAPI of cells adherent
on sterile coverslips revealed chromatin condensation and
fragmentation, typical signs of apoptosis, in samples of
Neuro2A-APP exposed to DA (Fig. 1A). TUNEL staining con-
firmed the occurrence of DNA fragmentation in these sam-
ples (Fig. 1B). A quantitative estimation of DA toxicity was
obtained by cytofluorometry of the hypodiploid (so-called
subG1 peak) cell population, which mirrors late apoptotic
cells, in the cultures exposed or not for 16 h to DA. While
sham-transfected Neuro2A cells showed negligible sensitiv-
ity, Neuro2A-APPswe cells showed high sensitivity to DA
toxicity (Fig. 1C). As an additional quantification and proof
of the apoptosis induced by DA, we estimated by cytofluoro-
metry the presence of phosphatidyl-serine on the outer
leaflet of the plasma membrane (an early marker of apopto-
sis) in sham- and APP-transfected Neuro2A cells treated in
the absence or in the presence of the pan-caspase inhibitor
zVAD-fmk. Data showed that as much as 40% of the APP-
over-expressing cells treated with DA for 16 h were positive
for annexinV (indicative of phosphatidyl-serine exterioriza-
tion) and that pre-incubation with zVAD-fmk completely
abrogated this effect (Fig. 1D). Taken together, these data
demonstrate that chronic DA stimulation, while not toxic to
the sham-transfected counterpart, causes apoptotic cell
death in Neuro2A cells over-expressing transgenic human
APP. Because of the pro-oxidative nature of DA excitotoxicity,

Fig. 1 – Dopamine induces apoptosis in Neuro2A cells over-expressing human APP. (A) Sham- and human APP (Swedish
mutant)-transfected Neuro2A cells were plated and let adhere on coverslips and then treated or not for 16 h with 250 lM
dopamine (DA). At the end, the nuclei of the cells were labeled with DAPI to evidence chromatin alterations. DA induced
chromatin condensation and fragmentation (arrows) in Neuro2A-APPswe cells, but not in the sham-transfected counterpart.
(B) The cells treated as above were processed for TUNEL fluorescent staining to evidence nicked DNA as a sign of apoptosis.
Images show the presence of TUNEL-positive nuclei in a larger proportion in APP-expressing cells than in sham-transfected
counterpart. (C) Sham-transfected and APPswe-expressing cells were plated on Petri dishes and treated or not for 16 h with
250 lM DA. At the end, adherent and suspended cells were recovered, fixed in ethanol and labeled with PI, and finally
analyzed by cytofluorometry to estimate the hypodiploid (SubG1) cell population. In DA-treated cultures, the percentage of
cells containing a SubG1 amount of DNA was in APP-expressing clones 2.5–3.0-fold that in the parental sham-transfected
clone. (D) Sham-transfected and APPswe-expressing cells were plated on Petri dishes and treated or not for 16 h with 250 lM
DA in the absence or the presence of the pan-caspase inhibitor zVAD-fmk. At the end, adherent and suspended cells were
recovered, labeled with AnnexinV-FITC, and analyzed by cytofluorometry to estimate the proportion of apoptotic cells. DA
greatly increased the proportion of Annexin-FITC-positive cells in APP-expressing cells and this effect was completely
abolished by zVAD-fmk. The fluorescent images and the cytofluorograms shown in this figure are representative of four
independent experiments in triple.
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7104
we suspected that the apoptotic pathway involved the
lysosome–mitochondrion axis (Castino et al., 2005, 2007).
We checked the integrity of lysosomes with the acidotropic
fluorochrome Acridine Orange, which fluoresces red when
protonated in acidic compartments and green when in
compartments at neutral pH. The images in Fig. 2A show
that endosomes and lysosomes retained their acidity in the
first 8 h of exposure to DA, whereas by 16 h these organelles
lost their integrity in a large number of Neuro2A-APPswe
cells. As quantified by cytofluorometry, at this time 450% of
the cells exposed to DA had lost their staining with red-
emitting Acridine Orange (Fig. 2B). To assess the integrity of
the outer mitochondrial membrane, we employed the fluor-
ochromes Rhodamine-123 and mitotracker, which emit a red
fluorescence when accumulate in the intermembrane space.
These fluorochromes loose their fluorescence when the outer
mitochondrial membrane becomes leaky and the mitochon-
drial membrane potential (DCm) drops. Cytofluorometry data
of Rhodamine-123 staining (upper panel in Fig. 2C) were
compatible with leakage from mitochondria in Neuro2A-
APPswe cells exposed to DA. To further confirm the activation
of the intrinsic apoptotic pathway, the cells were double-
stained with mitotracker and antibodies against the confor-
mational active bax (Castino et al., 2007). In controls, the
mitochondria were red-stained with mitotracker and no bax
oligomerization was evident, whereas in the cells exposed to
DA mitochondria were not labeled with mitotracker and
oligomerization of bax was clearly present (Fig. 2C, lower
panel). We then checked whether the caspase-8 mediated
extrinsic pathway was also activated by DA. To this end, the
cells were pre-incubated or not with IETD-CHO, a specific
inhibitor of caspase-8, and then cell viability was assessed in
cultures exposed for up to 16 h to DA by using CellTracker, a
fluorescent tracer of the mitochondrial metabolic activity.
In DA-treated cultures, early signs of mitochondrial suffer-
ance were apparent at 12 h and cell loss was clearly evident at
16 h, and the caspase-8 inhibitor could not rescue cell
viability (Fig. 2D), indicating that the extrinsic apoptotic
pathway was not involved in DA toxicity. To define the
temporal hierarchy of the events involving lysosomes and
mitochondria, we performed a parallel Acridine Orange and
bax/mitotracker staining in a time-course experiment in cells

Fig. 2 – Dopamine affects lysosome and mitochondrion integrity in APPswe-expressing Neuro2A cells. (A) Neuro2A cells
expressing the Swedish mutant of human APP695 were plated on coverslips and exposed to DA for increasing time of
incubation. Lysosome integrity was assessed by Acridine Orange staining. Upon DA treatment, in a large proportion of the cells the
acidic compartments cluster at one pole of the cell and eventually (at 16 h) loose the metachromatic fluorescent dye as a
consequence of membrane rupture. (B) Neuro2A-APPswe cells were plated on Petri dishes and treated with DA for 16 h. At the end,
the cells were recovered, labeled with Acridine Orange and analyzed by cytofluorometry to assay the shift in emitted fluorescence
associated with lysosomal leakage (upper panel). A parallel set of cultures on coverslips was imaged under the fluorescence
microscope (lower panel). (C) Neuro2A-APPswe cells were plated on Petri dishes and treated with DA for 16 h. At the end, the cells
were recovered, labeled with Rho-123 and analyzed by cytofluorometry to assay the shift in emitted fluorescence associated with
mitochondrial leakage (upper panel). A parallel set of cultures on coverslips was labeled with mitotracker, fixed and processed for
bax immunofluorescence (lower panel). The images show the occurrence of mitochondrial permeability and bax oligomerization in
cells exposed to DA. (D) Neuro2A-APPswe cells were plated on coverslips and treated with DA for the time indicated in the absence
or in the presence of the caspase-8 inhibitor IETD-CHO, and at the end of incubation the cells were processed for CellTracker
staining and imaging. (E) Neuro2A-APPswe cells were plated on coverslips and treated with DA for the time indicated in the
absence or in the presence of the caspase-8 inhibitor IETD-CHO. At each time point, the coverslips were processed for Acridine
Orange or bax/mitotracker/DAPI staining and imaged under the fluorescence microscope. (F) Neuro2A-APPswe cells were plated on
coverslips and treated with DA for 16 h. At the end, the cells were labeled with mitotracker, fixed and processed for cathepsin D
(catD) immunofluorescence. The images show a cytosolic diffuse staining of cathepsin D associated with mitochondrial
permeability in cells exposed to DA. The fluorescent images and the cytofluorograms shown in this figure are representative of
three independent experiments in triple. (For interpretation of the references to color in this figure legend, the reader is referred to
the web version of this article.)
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 105
exposed to DA in the absence or in the presence of the
caspase-8 inhibitor IETD-CHO. This experiment (Fig. 2E)
demonstrated that (1) lysosome leakage occurred between 4
and 8 h, while bax activation and mitochondria leakage
occurred at a time 48 h and (2) inhibition of caspase-8
neither preclude, nor altered the sequence of, such events.

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7106
A direct link between lysosomal leakage and mitochondria
permeabilization was suggested by the concomitant diffuse
cytosolic staining of lysosomal Cathepsin D, a protease
resident in endosomes and lysosomes, and the absence of
mitotracker staining in DA-treated cells at 16 h (Fig. 2E).
Altogether, these data indicate that DA triggers apoptosis in
dopaminergic Neuro2A cells over-expressing APPswe by
sequential destabilization of lysosomes and mitochondria
and subsequent activation of the intrinsic caspase-cascade.
Next, we investigated on the involvement of APP in the
sensitization of neuronal cells toward DA toxicity.
Fig. 3 – Dopamine induces progressive proteolytic
degradation of transgenic human APP in APPswe-
expressing Neuro2A cells. Neuro2A-swe cells adherent on
Petri dishes were exposed to DA for increasing time of
incubation. At the end, cell homogenates were resolved by
electrophoresis and human APP-related peptides were
identified by western blotting using the anti-N46–60
antibody. A specific band with an estimated apparent
molecular weight of 105–110 kDa was detected only in
samples of the Neuro2A-APPswe clone, as expected. The
intensity of this band, as estimated by densitometry
(average of two independent experiments), declined
progressively with time of incubation with DA.
2.2. Dopamine induces the progressive degradation ofAPPswe
We looked at the fate of APP in transfected Neuro2A cells
exposed to DA for increasing time of incubation. The full
length APP protein was identified with an antibody specific for
human APP (anti-N46–60) directed to the N-terminus (residues
46–60). The APP695-related molecular species potentially
recognized by this antibody are reported in Table 1. Sham-
transfected and APPswe-expressing Neuro2A cells were incu-
bated for up to 8 h with DA (a time at which apoptotic signs
are not yet evident), then the presence of APP-related peptides
were identified in cell homogenates by western blotting. The
antibody revealed the presence of a specific band running at
approximately 110 kDa in APPswe-transfected clones, not in
sham-transfected ones (Fig. 3). A second band, faintly detect-
able in both clones, running at approximately 96 kDa was also
detected. This band could tentatively represent either the
a- or b-APP soluble fragment. However, the fact that it was
present in both clones and that it was not reproduced
in other western blotting (see below) indicates that it is an
occasional contaminant. The amount of cell-associated holo-
APP (as revealed by the anti-N46–60) decayed with time of
exposure to DA. Similar data were obtained in independent
experiments in which APP was identified with antibodies
directed to different epitopes (see below), and therefore
proteolysis of only the N-terminal epitope was excluded.
In a separate study, we found a time-dependent accumulation
of the 4 kDa Ab peptide in the culture medium of DA-treated
APPswe-cells (not shown), suggesting that holo-APP decay
could mirror the amyloidogenic processing of APP.
Table 1 – Antibodies used to detect APP-related molecular spe
Antibody Peptide recognized Approx. M
Anti-N46–60 holo-APP 105–115
sAPPb 94–95
sAPPa 96–97
Anti-AbN1–17 holo-APP 105–115
sAPPa 96–97
C99 (597-695) 12
Ab40–42 4
Anti-C676–695 holo-APP 105–115
C99 (597-695) 12
C83 10
AICD 8
2.3. Dopamine toxicity Is associated with the rapidtranslocation of APP into endosomal–lysosomalcompartments
APP has been shown to undergo amyloidogenic processing
following clathrin-dependent endocytosis driven by the YENPTY
(671–676 in APP695) motif at the C-terminus (Koo and Squazzo,
1994). We therefore looked at the intracellular traffic and
localization of APP as affected by DA treatment. In these
experiments, we followed APP with an antibody (anti-C676–695)
directed to the C-terminus (residues 676–695) that allows to
identify the full length protein and its processed C-fragments
(Table 1). Confocal fluorescence imaging at high magnification
showed the presence of peripheral discrete spots indicative of
the presence of APP on the plasma membrane in control cells
and its rapid (within 30 min) translocation into EEA1-positive
vesicles (Early Endosome Antigen 1 is a marker of early endocytic
vesicles) upon DA treatment (Fig. 4A). Consistent with ongoing
cies.
W (kDa) Notes
Secreted and/or degraded
Secreted and/or degraded
Secreted and/or degraded
C-term after b-secretase cleavage (degraded)
Secreted
C-term after b-secretase cleavage (degraded)
C-term after a-secretase cleavage (degraded)
C-term after g-secretase cleavage (degraded)

Fig. 4 – Dopamine induces the rapid endocytosis of APP. Neuro2A cells expressing the Swedish mutant of human APP were
plated on coverslips and exposed or not to DA for the time indicated. (A) At the end, the cells were processed for
immunofluorescence labeling of EEA1 or APP. Alternatively, the cells were first labeled with Dextran-FITC to trace the
endocytic pathway and then fixed and stained for APP by immunofluorescence. Arrows point to co-staining of APP with
markers of early endosomes and with Dextran-FITC in cells exposed to DA. (B) The cells were processed for
immunofluorescence staining of EEA1 and APP. Images show that after a 4 h treatment with DA the bulk of APP is found in
perinuclear clusters. (C) Neuro2A-APPswe cells were plated on coverslips and exposed to DA for the time indicated and then
processed for immunofluorescence labeling of dynamin and APP. Images show that APP moves toward intracellular
compartments upon exposure to DA, while dynamin consistently remains localized beneath the plasma membrane.
(D) Neuro2A-APPswe cells were transfected with a plasmid coding for the fluorescent chimera Cathepsin D-GFP (CatD-GFP)
and then exposed to DA for 1 or 16 h and, at the end, processed for immunofluorescence labeling of APP. Images show that
colocalization of APP with CD-GFP increases with time of exposure to DA. APP was detected with the anti-C676–695 antibody.
The fluorescent images shown in this figure are representative of three independent experiments. (E) and (F) The cells were
plated on Petri dishes, transfected with a control duplex or a Dynamin I-specific siRNA and then treated or not with DA
for 16 h. At the end, the cells were processed for cytofluorometry analysis of the annexinV-positive (E) and of the subG1
(F) population. Data shown in panels E and F have been reproduced in two independent experiments.
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 107
endocytosis, in DA-treated cells APP colocalized with the endo-
cytosis tracers Dextran-FITC (Fig. 4A) and Lysotracker (not
shown). By 4 h of DA treatment, the bulk of APP appeared
clustered at one pole in the vicinity of the nucleus and showed
reduced colocalization with EEA1 (Fig. 4B). This localization
resembled that of acidic compartments as detected by Acridine
Orange staining (Fig. 2A). DA-induced movement of APP toward
intracellular sites was further assessed using dynamin as a
plasma membrane marker (Fig. 4C). To see whether with time
APP further proceeded downstream the endocytic pathway to
endosomes and lysosomes, we monitored its localization in cells
transiently transfected with a plasmid driving the synthesis of
the endosomal–lysosomal protease Cathepsin D fused with the
green fluorescent protein (CD-GFP). While in control cells no-
colocalization was observed, at 1 h of treatment some organelles
labeled with CD-GFP appeared to also contain APP (Fig. 4D). After

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7108
16 h of DA treatment, a nearly complete colocalization of APP
with lysosomal CD-GFP was observed (Fig. 4D). Note that at this
time as many as 50% of the cells detached and most of the cells
still adherent on plastic showed lysosome leakage (Fig. 2). It is
assumed, however, that the size of chimeric CD-GFP exceeds
that allowed to leak out from permeabilized lysosomes (esti-
mated to be of approximately 40 kDa). We asked about the
possible functional link between endocytosis, APP proteolysis
and apoptosis induced by DA. Dynamin, a GTPase involved in
endocytosis and intracellular membrane trafficking, has been
shown to play a pivotal role in APP endocytosis in Neuro2A cells
(Ehehalt et al., 2003). In a separate experiment, we found that
small-interference RNA-mediated knock-down of dynamin pre-
vented the internalization and degradation of APP in Neruo2A-
APPswe cells exposed to DA (not shown). Of note, cytofluoro-
metric analyses demonstrated the complete absence of the
annexinV-positive and subG1 apoptotic population in dynamin-
silenced cultures exposed for 16 h to DA (Fig. 4E and F).
2.4. Pepstatin A inhibits APP proteolysis and protectsfrom DA toxicity
APP processing in endosomal compartments and leading to
Ab peptide production involves the sequential proteolysis by
Fig. 5 – Pepstatin A inhibits dopamine-induced processing of
100 lM Pepstatin A (Pst) and then exposed or not for the time i
was resolved by SDS-gel electrophoresis and APP molecular sp
incubated for 6 h with DA and APP was detected with the anti-N
and APP was detected with the anti-C676–695 or anti-AbN1–17
and 20 h with DA and APP was detected with the anti-AbN1–17
the end of incubation (20 h). The relative intensity of holo-APP (
in two other independent experiments.
a b-secretase and a g-secretase activity (reviewed in Chow
et al., 2010; O’Brien and Wong, 2011; Zhang et al., 2011). Both
these activities are performed by aspartyl-type proteases that
can be found in endosomes (Schechter and Ziv, 2008;
Fukumori et al., 2006; Kinoshita et al., 2003), and therefore
should be effectively inhibited by large spectrum aspartic
protease inhibitors such as Pepstatin A, able to accumulate
within these organelles (Tian et al., 2002; Wolfe and
Haass, 2001). To confirm that DA toxicity was linked
to endosomal–lysosomal proteolysis of APP, we checked
whether this inhibitor could indeed prevent APPswe proteo-
lysis and at the same time save the cells from DA. Neuro2A-
APPswe cells were pre-incubated 12 h with Pepstatin A and
then exposed to DA for increasing time. Holo-APP was
immunodetected in cell homogenates with antibodies direc-
ted to different epitopes and allowing the detection of the
various molecular species as indicated in Table 1. The result
showed that Pepstatin A could prevent the loss of holo-
APPswe imposed by DA (approximately 40% in 6 h, as
detected by anti-N46–60) (Fig. 5A). We extended the incuba-
tion to 16 h, a time at which almost 50% of the cells exposed
to DA die by apoptosis. At this time, Pepstatin A increased the
amount (about 4-fold) of APP detectable in control (untreated)
cells, whichever the epitope recognized by the antibody,
APP. Neuro2A-APPswe cells were pre-incubated 12 h with
ndicated to dopamine (DA). At the end, the cell homogenate
ecies identified by western blotting. (A) The cells were
46–60 antibody. (B) The cells were incubated for 16 h with DA
antibody, as indicated. (C) The cells were incubated for 1, 6
antibody. Control and Pepstatin A-treated cells were taken at
normalized versus actin) bands is reported. Data reproduced
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 109
suggesting that APP constitutively undergoes a slow proteo-
lytic processing (Fig. 5B and C). As assessed by western
blotting with the anti-C676–695 antibody, by 16 h APP was
reduced in DA-treated cells by some 30% (as compared to
control cells), and this loss was fully prevented by Pepstatin A
(Fig. 5B, left panel). In a parallel independent experiment, APP
was detected by western blotting with an antibody directed to
an epitope placed at the N-terminus (residues 1–17) of the Ab
sequence. It was calculated that, as detected with this anti-
body (anti-AbN1–17), DA imposed a loss of APP of approxi-
mately 80%, and again this loss was practically completely
rescued by Pepstatin A (Fig. 5B, right panel). Finally, we
performed a time-course study of the effects of DA and
Pepstatin A on APP as detected with the anti-AbN1–17. This
experiment confirmed the progressive and extensive loss of
APP detectable with this antibody, suggesting that a large
portion (�80%) of APP was processed to produce the Ab
peptide (Fig. 5C). Again, Pepstain A confirmed its ability to
prevent such proteolysis (Fig. 5C).
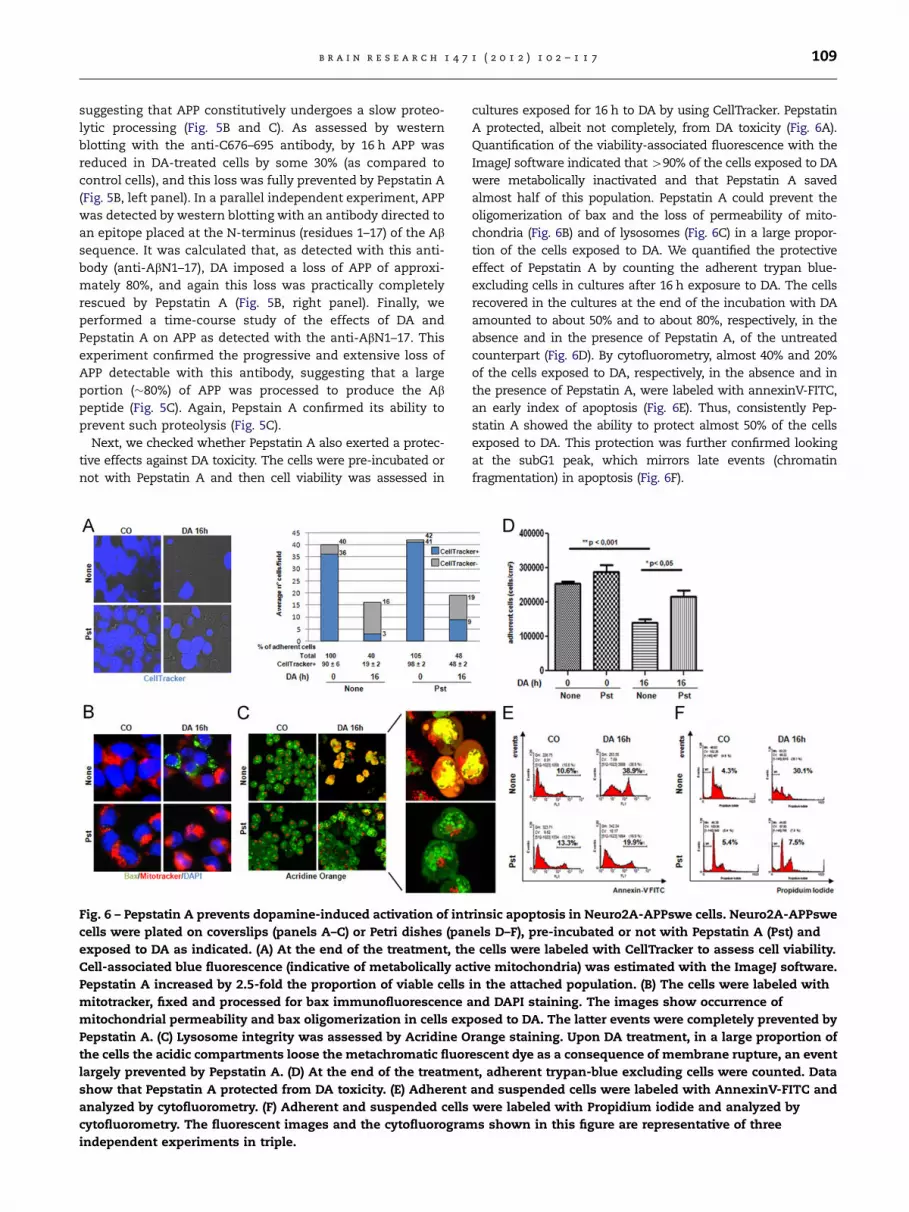
Next, we checked whether Pepstatin A also exerted a protec-
tive effects against DA toxicity. The cells were pre-incubated or
not with Pepstatin A and then cell viability was assessed in
Fig. 6 – Pepstatin A prevents dopamine-induced activation of int
cells were plated on coverslips (panels A–C) or Petri dishes (pan
exposed to DA as indicated. (A) At the end of the treatment, th
Cell-associated blue fluorescence (indicative of metabolically ac
Pepstatin A increased by 2.5-fold the proportion of viable cells
mitotracker, fixed and processed for bax immunofluorescence a
mitochondrial permeability and bax oligomerization in cells exp
Pepstatin A. (C) Lysosome integrity was assessed by Acridine O
the cells the acidic compartments loose the metachromatic fluor
largely prevented by Pepstatin A. (D) At the end of the treatmen
show that Pepstatin A protected from DA toxicity. (E) Adherent
analyzed by cytofluorometry. (F) Adherent and suspended cells
cytofluorometry. The fluorescent images and the cytofluorogram
independent experiments in triple.
cultures exposed for 16 h to DA by using CellTracker. Pepstatin
A protected, albeit not completely, from DA toxicity (Fig. 6A).
Quantification of the viability-associated fluorescence with the
ImageJ software indicated that 490% of the cells exposed to DA
were metabolically inactivated and that Pepstatin A saved
almost half of this population. Pepstatin A could prevent the
oligomerization of bax and the loss of permeability of mito-
chondria (Fig. 6B) and of lysosomes (Fig. 6C) in a large propor-
tion of the cells exposed to DA. We quantified the protective
effect of Pepstatin A by counting the adherent trypan blue-
excluding cells in cultures after 16 h exposure to DA. The cells
recovered in the cultures at the end of the incubation with DA
amounted to about 50% and to about 80%, respectively, in the
absence and in the presence of Pepstatin A, of the untreated
counterpart (Fig. 6D). By cytofluorometry, almost 40% and 20%
of the cells exposed to DA, respectively, in the absence and in
the presence of Pepstatin A, were labeled with annexinV-FITC,
an early index of apoptosis (Fig. 6E). Thus, consistently Pep-
statin A showed the ability to protect almost 50% of the cells
exposed to DA. This protection was further confirmed looking
at the subG1 peak, which mirrors late events (chromatin
fragmentation) in apoptosis (Fig. 6F).
rinsic apoptosis in Neuro2A-APPswe cells. Neuro2A-APPswe
els D–F), pre-incubated or not with Pepstatin A (Pst) and
e cells were labeled with CellTracker to assess cell viability.
tive mitochondria) was estimated with the ImageJ software.
in the attached population. (B) The cells were labeled with
nd DAPI staining. The images show occurrence of
osed to DA. The latter events were completely prevented by
range staining. Upon DA treatment, in a large proportion of
escent dye as a consequence of membrane rupture, an event
t, adherent trypan-blue excluding cells were counted. Data
and suspended cells were labeled with AnnexinV-FITC and
were labeled with Propidium iodide and analyzed by
s shown in this figure are representative of three

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7110
2.5. Chloroquine inhibits APP proteolysis and protectsfrom DA toxicity
The antimalaric drug chloroquine, a lysosomotropic weak
base that impairs endosomal–lysosomal hydrolysis by raising
the luminal pH, has been shown to interfere with the APP
processing (Caporaso et al., 1992; Caporaso et al., 1994) and
Ab peptide-associated toxicity (Liu et al., 2010). We first
checked whether and how chloroquine protected APP from
DA-induced proteolysis at 6 and 16 h of treatment. This
experiment confirmed that APP undergoes basal proteolysis
that could be halted by chloroquine. As assessed by western
blotting with the anti-C676–695 antibody, chloroquine com-
pletely protected APP from DA-induced proteolysis (Fig. 7A).
Of note, in chloroquine-treated samples APP was detected by
the anti-C676–695 as a doublet, the upper band presumably
bearing additional complex-type sugars. An even higher
protection by chloroquine was apparent by detecting APP
with the anti-AbN1–17 antibody. These data are consistent
with our previous finding (Fig. 5) and strongly support the
view that most, if not all, APP is proteolyzed with generation
of the Ab peptide. It is worthy to note that this antibody also
revealed APP as a doublet, but in this case the extra band
migrated faster, showing an apparent molecular weight
diminished of approximately 1.5 kDa (Fig. 7). This second
band was however less protected by chloroquine in the long
incubation with DA (Fig. 7C). Since this band was not detected
with the anti-C676–695, it likely represents a molecular
species that has lost a small peptide at the C-terminus of APP.
Finally, we focused on the link between APP proteolysis and
cell death induced by DA and the protection by chloroquine.
Based on the CellTracker assay, chloroquine afforded a nearly
complete protection from DA toxicity, unequivocally higher than
that of Pepstatin A (compare Figs. 8A and 6A). Consistently,
chloroquine prevented the DA-induced activation of bax in
those cells in which APP was not processed to produce the Ab
peptide, as suggested by immunofluorescence co-labeling with
anti-conformational active bax and anti-AbN1–17 antibodies
Fig. 7 – Chloroquine inhibits dopamine-induced processing of A
30 lM chloroquine (ClQ) and then exposed or not for the time i
was resolved by SDS-gel electrophoresis and APP molecular sp
incubated for 6 h with DA and APP was detected with the anti-C
cells were incubated for 16 h with DA and APP was detected w
with DA and APP was detected with the anti-AbN1–17 antibody
(Fig. 8B). Chloroquine protection of APP from DA-induced
proteolysis was not ascribable to inhibition of endocytosis, since
APP detected with the anti-AbN1–17 antibody reached the
endosomal–lysosomal compartments as demonstrated by its
colocalization with Cathepsin D (Fig. 8C).
2.6. Inhibition of c-secretase activity or phorbol esterstimulation of a-secretase activity protects from dopamine-induced processing of APP and cell toxicity
Finally, we sought to determine if and to what extent the
amyloidogenic processing of APP was indeed causally linked to
DA toxicity in Neuro2A-APPswe cells. The Ab fragment is formed
following g-secretase hydroysis of the C99 peptide generated by
b-secretase cleavage of holo-APP. To inhibit this processing, we
employed the non-competitive inhibitor of g-secretase L685,458
(Tian et al., 2002). As an additional approach to prevent amyloi-
dogenic processing, we stimulated the a-secretase alternative
pathway (Vincent and Govitrapong, 2011) by using a phorbol
ester (Savage et al., 1998). The cells were or not pre-incubated
with L685,458 or PMA and then exposed (or not) to DA for 16 h,
and at the end APP processing and cell toxicity were assessed.
Both drugs were shown able to prevent holo-APP degradation, as
detected by western blotting with either the anti-C676–695 and
the anti-AbN1–17 antibodies (Fig. 9A). DA-toxicity was largely
prevented in cells incubated with either the g-secretase inhibitor
L685,458 or the a-secretase stimulator PMA, as shown by phase-
contrast imaging of the monolayer, CellTracker staining and
bax staining (Fig. 9B). Of note, both these drugs prevented
DA-induced apoptosis in those cells in which amyloidogenic
processing of APP was abrogated, as suggested by immunofluor-
escence co-labeling of active bax and AbN1–17 APP (Fig. 9B).
3. Discussion
Alpha-synuclein-positive Lewy Bodies, accumulation of
hyperphosphorylated tau and neuron loss have been reported
PP. Neuro2A-APPswe cells were pre-incubated 30 min with
ndicated to dopamine (DA). At the end, the cell homogenate
ecies identified by western blotting. (A) The cells were
676–695 or the anti-AbN1–17 antibody, as indicated. (B) The
ith the anti-C676–695. (C) The cells were incubated for 16 h
. Data representative of two independent experiments.

Fig. 8 – Chloroquine prevents dopamine-induced toxicity in Neuro2A-APPswe cells. Neuro2A-APPswe cells were plated on
coverslips, pre-incubated or not with chloroquine (ClQ) and exposed to DA for 16 h. (A) At the end of the treatment, the cell
viability was assessed by CellTracker staining. Cell-associated blue fluorescence was estimated with the ImageJ software.
Chloroquine increased by 3.5-fold the proportion of viable cells in the attached population. (B) and (C) The cells were fixed
and processed for APP and bax (panel B) or cathepsin D (catD, panel C) immunofluorescence. The images are suggestive of
APP degradation (as revealed by the anti-AbN1–17 antibody) in cells exposed to DA, in concomitance with bax
oligomerization (panel B). Chloroquine inhibits APP processing and loss of cell-associated Ab-reactivity, and prevents bax
oligomerization but not translocation of APP in catD-positive organelles. Data representative of two independent
experiments in triple. (For interpretation of the references to color in this figure legend, the reader is referred to the web
version of this article.)
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 111
in the substantia nigra and extranigral nuclei of AD patients
featuring parkinsonian signs (Burns et al., 2005; Schneider
et al., 2002). Whether the hyper-expression of APP exacer-
bates neuron susceptibility to DA excitotoxicity is not known.
Here, we addressed this issue by studying the molecular and
cellular consequence of DA treatment in Neuro2A cells over-
expressing human APP695swe. We found that DA increased
the recycling of APP determining its translocation and pro-
teolysis within endosomal compartments. These events were
accompanied by rupture of lysosome and mitochondria
integrity and onset of caspase-mediated cell death. We
investigated on the functional relationship between APPswe
processing and neuronal cell death induced by DA. The
Swedish mutation (K595N, M596L in APP695) causes early
onset familial AD and is associated with altered trafficking
and processing of APP (Lo et al., 1994; Lorenzen et al., 2011),
resulting in up to 10-fold higher production of the Ab peptide
(Haass et al., 1995; Sinha and Lieberburg, 1999). Considering
that the Swedish mutation is placed within the b-secretase
consensus sequence, the above findings strengthen the rela-
tionship between cleavage at b-site, excessive generation of
Ab peptide and AD (Thinakaran and Koo, 2008). DA induced
the clustering of endosomes and lysosomes and the rapid
translocation of plasma membrane APPswe into Cathepsin
D-positive acidic compartments. Recently, it has been
shown that, contrary to wild-type APP that can be found in
lysosomes, internalized APPswe localizes to endosomes
(Lorenzen et al., 2011). This would suggest that in our system
DA induced the translocation of APPswe into endosomes. We
asked about the proteases potentially involved in endosomal
processing of APPswe. Cathepsin B was excluded, since this
protease can cleave wild-type APP with production of Ab
peptides, but not APPswe (Hook et al., 2009). Actually, genetic
ablation of Cathepsin B rather increased the accumulation of
Ab peptides in the brain of transgenic mice expressing
APPswe (Mueller-Steiner et al., 2006). The aspartyl-proteases
BACE1 and Cathepsin D were considered as good candidates,
as they have been able to cleave at high rate APPswe, and
with minor efficiency APPwt (Schechter and Ziv, 2008). When
the wide-spectrum aspartyl-protease inhibitor Pepstatin A
was employed, almost 80% of full length APPswe was rescued
from proteolysis induced by DA as detected by the anti-
AbN1–17, indicating that the production of the Ab peptide
was nearly completely abrogated. A similar reduced the
production of Ab peptide was attained when APP internaliza-
tion was abolished by mutation of the YENPTY sequence at

Fig. 9 – L685,458 and PMA prevent APP degradation and cell toxicity induced by dopamine. Neuro2A-APPswe cells were
plated on coverslips or Petri dishes, pre-incubated or not with L685,458 or PMA as indicated, and exposed to DA for 16 h. (A)
At the end, the cell homogenate was resolved by SDS-gel electrophoresis and APP molecular species identified by western
blotting with the anti-C676–695 or the anti-AbN1–17 antibody, as indicated. The relative intensity of holo-APP (normalized
versus actin) bands is reported. Data representative of three independent experiments. (B) At the end of the treatment, the
monolayer in Petri dishes was photographed and the cells grown on coverslips were labeled with CellTracker to assess cell
viability or processed for fluorescence staining with DAPI (nuclei) and anti-bax and anti-AbN1–17 antibodies, as indicated.
The images shown are representative of three independent experiments and demonstrate that both L685,458 and PMA could
protect the cells from DA toxicity.
b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7112
the C-terminus (Koo and Squazzo, 1994). Internalization of
APP relies on dynamin-driven endocytosis (Ehehalt et al.,
2003). Consistently, siRNA-mediated gene knock-down of
dynamin greatly impaired endocytosis and degradation of
APPswe, and also abolished cell toxicity induced by DA. That
proteolysis of APPswe induced by DA likely occurred in acidic
compartments is supported by the observation that chlor-
oquine, a weak base widely used to rise the luminal pH of
endosomes and lysosomes, also blocked this process, in
agreement with a previous report (Caporaso et al., 1994). In
this context, it is to be noted that in chloroquine-treated
samples an additional band of APPswe, showing an apparent
molecular weight reduced of �1.5 kDa, could be detected.
This band was evidenced by the anti-AbN1–17, not by the
anti-C676–695, antibody, suggesting that in the absence of
chloroquine a small peptide is removed at the C-terminus of
APP. The C-terminus of APPswe is indeed very unstable.
Thinakaran et al. (1996) failed to detect the APP C-terminus
fragment generated by the action of g-secretase, and sug-
gested that this fragment undergoes rapid degradation.
Indeed, whichever antibody we used, we never detected
specific bands below the molecular weight of full length
APPswe. In theory, the sAPPb and the transmembrane C99
fragment generated by BACE could be found in cell homo-
genates. We hypothesize that these species either are rapidly
degraded or extruded from the cell under DA stimulation.
Two additional approaches allowed to exclude the possibility
that DA toxicity was associated with the activation of APP
processing pathways other than the amyloidogenic. In fact,
inhibition of the g-secretase activity or the stimulation of
the a-secretase both afforded complete protection from
DA-induced APP degradation and cell toxicity. It has been
shown that the products of APP proteolysis, including the Ab
peptide, transiently accumulate in exosomes of multivesicu-
lar body-endosomes from which are then released extra-
cellularly (Rajendran et al., 2006). Thus, assuming that
proteolysis of APPswe indeed occurred in multivesicular
body-endosomes, it is conceivable that the products were
promptly exocytosed under DA stimulation. The use of
lysosomal activity inhibitors (i.e., Pepstain A and chloroquine)
also evidenced that a portion of APPswe constitutively under-
goes processing, though in the presence of DA this process
was accelerated and associated with lysosomes and mito-
chondria dysfunction and cell death. Recently, it has been
shown that lysosome leakage may be due to the insertion in
the lysosomal membrane of toxic Ab-42 peptide taken up

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 113
from the extracellular mileau (Liu et al., 2010). In this context,
it is to note that chloroquine could protect Neuro2A-APPswe
cells from Ab42 toxicity by inhibiting the lysosomal mem-
brane insertion of the Ab peptide and thus preventing lyso-
some leakage (Liu et al., 2010). However, the effective toxicity
of the soluble Ab peptide is debatable. Therefore, we consider
the possibility that DA-induced degradation of APP leads to
cell toxicity through the production of pro-oxidative frag-
ments that generate reactive oxygen species.
Parkinsonian-like motor signs (including rigidity, tremor,
bradykinesia) have been reported in 13–36% of AD patients
Fig. 10 – Schematic representation of the results. The upper par
and the peptide generated by a, b and c secretase activity. The
potential sites of cleavage by proteases, the sequence specifica
indicated. The lower part of the scheme illustrates the principa
work. DA stimulates the recycling of endocytic vesicles and indu
Here, APP is subject to amyloidogenic processing and the prod
degradation is likely associated with the production of reactive
undergo permeabilization and leakage of cathepsin D. Addition
within lysosomes, and lead to lysosomes leakage. The followin
mitochondrial membranes, the permeabilization of mitochondr
(Scarmeas et al., 2004; Wilson et al., 2000) and appear to be
related to morbidity and mortality. Such parkinsonian symp-
toms are associated with more rapid cognitive decline and
deterioration of physical conditions (Chui et al., 1994;
Mortimer et al., 1992).
The present findings (schematically reproduced in Fig. 10)
strongly support the view that inhibiting aspartyl-protease-
mediated proteolysis of APP within endosomes is a good
strategy to protect neurons in AD patients. Also chloroquine,
an FDA approved drug that can freely pass the blood–brain
barrier, would be a good candidate for this purpose. Indeed, a
t of the scheme shows the transgenic holo-APPswe protein
position of relevant post-translational modifications, the
lly recognized by the antibodies used in this work, are
l findings, and their interpretation, reported in the present
ces the translocation of APP into endosomal compartments.
ucts, including the Ab peptide, are rapidly secreted. APP
oxygen species (ROS) within lysosomes, which eventually
ally, the Ab peptide could be endocytosed and accumulate
g events include sequentially the oligomerization of bax on
ia, the activation of caspases and cell death.

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7114
clinical trial with hydroxychloroquine revealed no benefits in
terms of progression of dementia in AD patients treated for
18 months (Van Gool et al., 2001). Based on the data reported
here, we propose the prolonged use of this drug in AD
patients to prevent the progression toward parkinsonism, of
course keeping in mind the potential side effects (Block, 1998;
Good and Shader, 1982).
4. Experimental procedures
4.1. Cells and treatments
Mouse neuroblastoma Neuro2A cells (American Type Culture
Collection, Rockville, MD) and Neuro2A cells stably expres-
sing the transgenic Swedish-mutant APP695 (Thinakaran
et al., 1996) were cultivated under standard culture condi-
tions (37 1C; 95 v/v% air: 5 v/v% CO2) in Dulbecco Modified
Eagle’s Medium (cod. D5671, Sigma-Aldrich, St. Louis, USA)
supplemented with 10% heat-inactivated fetal bovine serum
(cod. DE14-801F, Lonza Group Ltd., Basel, Switzerland), 2 mM
L-glutamine (cod. 35050, Life Technologies Ltd., Paisley, UK),
1 mM sodium pyruvate (cod. S8636, Sigma-Aldrich), 1 w/v% of
non-essential aminoacids (cod. M7145, Sigma-Aldrich) and
1 w/v% of a penicillin–streptomycin solution (cod. P0781,
Sigma-Aldrich). Experiments were carried out during the log
phase of cell growth. Cells (25,000/cm2) were seeded on sterile
plastic dishes or coverslip and allowed to adhere for 24 h
prior to start any treatment. Treatments included 250 mM
dopamine (DA, cod. H8502, Sigma-Aldrich) and, prior to
exposure to DA, 30 mM chloroquine (ClQ, cod. C6628, Sigma-
Aldrich; 30 min in advance), 100 mM Pepstatin A (cod. P5318,
Sigma-Aldrich; 12 h in advance), 30 mM ZVAD (OMe)-fmk
(ZVAD, cod. 260-020-M005, Alexis Laboratories, San Diego,
CA; 1 h in advance), 20 mM IETD-CHO (IETD, cod. A1216,
Sigma-Aldrich; 1 h in advance), 1.5 mM L685,458 (cod.
H-5106, Bachem; 6 h in advance), 5 mM PMA (cod. 8139,
Sigma-Aldrich; 1 h in advance).
4.2. Small-interference RNA silencing of dynamin
Post-transcriptional silencing of dynamin expression was
achieved by the small interference RNA (siRNA) technology.
Duplexes of 27-nucleotide siRNA including two 30-overhan-
ging TT were synthesized by MWG Biotech AG (Washington,
DC). The sequence and use of the siRNA for sham transfec-
tion have been described previously (Trincheri et al., 2007).
The sense strand of siRNA targeting dynamin-1 mRNA was
50–CAG AAC ACA CUG AUG GAA GAA UCG GCC-30. Adherent
cells (plated at 15,000/cm2 in Petri dish) were incubated for
4 h with 400 pmol RNA-duplexes in the presence of 10 ml
Lipofectamine 2000 in 1 ml of Optimem. The cells were then
washed and treated 36 h post-transfection to allow maximal
effect on protein down-regulation.
4.3. Assessment of cell toxicity
At the end of incubation, adherent and suspended cells were
collected, diluted in a solution containing trypan blue and
counted to determine cell loss and occurrence of necrosis
(trypan blue positive cells). For cytofluorometry assessment
of cell death, adherent and suspended cells were collected,
washed in PBS, fixed in ice-cold 70 v/v% ethanol and labeled
with 0.18 mg/ml propidium iodide (PI, cod. P4170, Sigma-
Aldrich) in the presence of RNase A (0.4 mg/ml). Hypodiploid
(SubG1) labeled cells were assumed as apoptotic. In addition,
the presence of phosphatidyl-serine on the plasma mem-
brane, an index of apoptosis, was assessed by cytofluorome-
try in the whole cell population by Annexin-FITC labeling
(cod. ALX-209–256, Alexis Laboratories; 10 min at room tem-
perature) of non-fixed cells. At least 10,000 cells were ana-
lyzed using a FacScan flow cytometer (Becton Dickinson,
Mountain View, CA, USA) equipped with a 488 nm argon
laser. Data were elaborated with the winMDI software.
Apoptosis-associated chromatin alterations were detected
by staining the cells adherent on coverslips with the DNA-
labeling fluorescent dye 40,6-diamidino-2-phenylindole dihy-
drochloride (DAPI, cod. 32670, Sigma-Aldrich). In situ Term-
inal deoxynucleotidyl transferase-mediated dUTP-biotin nick
end labeling (TUNEL) for detection of apoptotic cells was
performed with the ‘‘In situ Cell Death Detection’’ fluorescent
Kit (cod. 1684817, Roche Diagnostics Corporation Indianapo-
lis, IN, USA) (Trincheri et al., 2008). To test cell viability, the
cells adherent on coverslips were labeled with CellTrackerTM
(CellTrackerTMBlue-CMAC 7-amino-4-chloromethylcoumarin)
(cod. C2110, Life Technologies Ltd.), a fluorescent dye that
emits blue fluorescence of intensity proportional to the
mitochondrial respiratory activity. At the end of the treat-
ment, the cells were loaded with CellTracker (5 mM for
20 min), then the cells were washed and incubated for
30 min and imaged under the fluorescence microscope
(Ekkapongpisit et al., 2012).
4.4. Primary antibodies used for immunofluorescence andwestern blotting
The following primary antibodies were used: a rabbit poly-
clonal antiserum specific for human Cathepsin D (CD) (Follo
et al., 2007), a polyclonal antibody specific for conformational
active bax (cod. 2772, Cell Signaling Technology, MA, USA), a
mouse monoclonal antibody specific for EEA1 (cod. 610456,
Becton, Dickinson and Company, Franklin Lakes, NJ, USA), a
mouse monoclonal antibody specific for dynamin (cod. 05319,
Millipore, Billerica, MA, USA), a mouse monoclonal antibody
specific for b-actin (cod. A5441, Sigma-Aldrich) and mouse
monoclonal antibody specific for b-tubulin (cod. T5293,
Sigma-Aldrich). APP-related peptides were detected with the
following antibodies (see also Table 1): rabbit polyclonal
anti-C-terminus (cod. 171610, Calbiochem, Merck KGaA,
Darmstadt, Germany); rabbit polyclonal anti-N-terminus
(cod. A8967, Sigma-Aldrich); mouse monoclonal anti-Ab-N-
terminus sequence (cod. 12266, Abcam, Cambridge, UK).
4.5. Immunofluorescence staining
Cells on coverslip were fixed in cold methanol and processed
for immunofluorescence as previously reported (Castino et al.,
2010). Immunocomplexes were revealed with secondary anti-
bodies, either IRIS-2 (green fluorescence)- or IRIS-3 (red fluor-
escence)-conjugated goat-anti-rabbit IgG or goat-anti-mouse

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 115
IgG (cod. 2W5-08, 2W5-07, 3W5-08, 3W5-07, Cyanine Technology
SpA, Turin, I), as appropriate.
4.6. Endocytosis and lysosomal and mitochondrialmembranes integrity
The endocytosis process was monitored using Dextran-FITC
(Life Technologies Ltd., Paisley, UK) as a fluorescent tracer
(Dragonetti et al., 2000). Lysosomal membrane integrity was
assessed with the metachromatic fluorescent dye Acridine
Orange (cod. A6529, Sigma-Aldrich), which emits a red-
orange fluorescence when reside within acidic compartments
(endosomes and lysosomes), and a yellow-green fluorescence
when resides in neutral compartments (cytoplasm). The cells
were incubated with Acridine Orange (15 mg/ml, 15 min), then
washed and rapidly imaged under the fluorescence micro-
scope or analyzed by cytofluorometry (Dragonetti et al., 2000).
Mitochondrial membrane integrity was tested by using Rho-
damine-123 hydrochloride (Rho-123, cod. 610-018-M005,
Alexis Laboratories) or Mitotracker Red (cod. M22425, Life
Technologies Ltd.). The cells were incubated for 10 min at
37 1C with 50 nM Rho-123, then washed and rapidly imaged
under the fluorescence microscope or analyzed by cytofluoro-
metry. Alternatively, the cells on coverslips were incubated
for 15 min at 37 1C with 0.2 ml/ml of Mitotracker solution,
fixed in 3.7% paraformaldehyde for 30 min and permeabilized
with 0.2% Triton X-100 for 20 min, and further processed for
fluorescence staining with anti-bax antibody (Castino et al.,
2007).
4.7. Cathepsin D fluorescent chimera and plasmidtransfection
The cDNA coding for human lysosomal Cathepsin D (Isidoro
et al., 1991) devoid of the stop codon was subcloned in the
multiple cloning site of the plasmids peGFP-N1 (cod. 6085-1,
Clontech Lab., Takara Bio Inc., Shiga, Japan) in order to drive
the synthesis of the fluorescent chimeras CD-GFP
(Ekkapongpisit et al., 2012). The cells were transfected with
the plasmid using the Lipofectamine 2000 Reagent (cod.
11668-019, Life Technologies Ltd.) method as suggested by
the purchaser. Briefly, cells were plated in P35 Petri dish at
15,000/cm2 and let adhere 24 h before to proceed with the
transfection. The DNA–Lipofectamine complexes were pre-
pared in 500 ml of Opti-MEM I Reduced Serum Medium (cod.
11058021, Life Technologies Ltd.) with 5 mg of plasmid and
10 ml of Lipofectamine. After 6 h of incubation, medium of
transfection was removed and replaced with a serum-con-
taining culture medium (10% FBS-DMEM) and the cells were
cultivated for 36 h to allow for maximal protein expression
prior to any treatment.
4.8. Fluorescence microscope imaging
Fluorescently labeled cells were observed under the fluores-
cence microscopes Leica DMI6000 or the confocal Leica
DMIRE2 (Leica Microsystems AG, Wetzlad, Germany). For each
experimental condition, three coverslips were prepared. Five
to ten fields (for a minimum of 100 cells) in each coverslip
were examined independently by two investigators. Selected
images of representative fields are shown. The ImageJ
software freely available at http://rsbweb.nih.gov/ij/ was
employed for quantification of the fluorescent signal.
4.9. Protein expression analysis
Protein expression was evaluated by standard immunoblot-
ting procedure as previously reported (Castino et al., 2007).
Cell homogenates were prepared by freeze–thawing and
ultrasonication in a buffer containing detergents and pro-
tease inhibitors. About 50 mg of cell proteins were denatured
with Laemmli sample buffer, separated by electrophoresis on
a 10% SDS-containing polyacrylamide gel and then electro-
blotted onto nitrocellulose membrane (Bio-Rad, Hercules, CA,
USA). Protein of interest was detected with the specific
primary antibody as detailed above. As an index of homo-
genate protein loading in the lanes was used b-Actin and
b-tubulin. Immunocomplexes were revealed by using a per-
oxidase-conjugated secondary antibody (cod. 170515, 1706516,
Bio-Rad), as appropriate, and subsequent peroxidase-induced
chemiluminescence reaction (cod. NEL103E001EA, PerkinEl-
mer, Waltham, MA, USA). Western blotting data were repro-
duced at least three times independently, unless otherwise
specified. Intensity of the bands was estimated by densito-
metry (Quantity One Software, Bio-Rad; ImageJ software).
4.10. Statistical analysis
All the experiments were performed in triple and data shown
have been reproduced at least three times (unless otherwise
specified). Densitometric data are reported for western blot-
ting shown (difference between replicates was less than 20%
of the absolute value). Quantification data from ImageJ and
cytofluorometry analyses and cell counting data were given
as average7SD. The Student’s t-test (with po0.05 for statis-
tical significance) was employed to compare the results from
different treatments. The Microsoft Excel XLStats software
was used.
Acknowledgments
Research supported by grants from San Paolo (Project Neu-
roscienze 2008.2395), Regione Piemonte (Ricerca Sanitaria
Finalizzata, Torino), Consorzio InterUniversitario per le Bio-
tecnologie (CIB, Trieste). The bio-imaging facility was donated
by Comoli, Ferrari & SpA (Novara, Italy). Thanks are due
to Dr. D.L. Feinstein (University of Illinois, Chicago, USA)
for advices for Ab western blotting, to Dr. V. Bruno
(Mahidol University, Bangkok, Thailand) for discussion, and
to Dr. C. Peracchio for excellent artwork and editorial
assistance.
r e f e r e n c e s
Akahoshi, E., Yoshimura, S., Uruno, S. Effect of dioxins on regulationof tyrosine hydroxylase gene expression by aryl hydrocarbonreceptor: a neurotoxicology study. Environ. Health 8, 24.

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7116
Bekris, L.M., Yu, C.E., Bird, T.D. Genetics of Alzheimer disease. J.Geriatr. Psychiatry Neurol. 23, 213–227.
Block, J.A., 1998. Hydroxychloroquine and retinal safety. Lancet351, 771.
Burns, J.M., Galvin, J.E., Roe, C.M. The pathology of the substantianigra in Alzheimer disease with extrapyramidal signs. Neu-rology 64, 1397–1403.
Cai, H., Wang, Y., McCarthy, D. BACE1 is the major beta-secretasefor generation of Abeta peptides by neurons. Nat. Neurosci. 4,233–234.
Caporaso, G.L., Gandy, S.E., Buxbaum, J.D. Chloroquine inhibitsintracellular degradation but not secretion of Alzheimer beta/A4 amyloid precursor protein. Proc. Natl. Acad. Sci. U.S.A. 89,2252–2256.
Caporaso, G.L., Takei, K., Gandy, S.E. Morphologic and biochem-ical analysis of the intracellular trafficking of the Alzheimerbeta/A4 amyloid precursor protein. J. Neurosci. 14, 3122–3138.
Castino, R., Bellio, N., Follo, C. Inhibition of PI3k class III-dependentautophagy prevents apoptosis and necrosis by oxidative stress indopaminergic neuroblastoma cells. Toxicol. Sci. 117, 152–162.
Castino, R., Bellio, N., Nicotra, G. Cathepsin D-Bax death pathwayin oxidative stressed neuroblastoma cells. Free Radic. Biol.Med. 42, 1305–1316.
Castino, R., Isidoro, C., Murphy, D., 2005. Autophagy-dependentcell survival and cell death in an autosomal dominant familialneurohypophyseal diabetes insipidus in vitro model. FASEB J.19, 1024–1026.
Chevallier, N., Vizzavona, J., Marambaud, P. Cathepsin D displaysin vitro b-secretase-like specificity. Brain Res. 750, 11–19.
Chow, V.W., Mattson, M.P., Wong, P.C. An overview of APPprocessing enzymes and products. Neuromol. Med. 12, 1–12.
Chui, H.C., Lyness, S.A., Sobel, E. Extrapyramidal signs andpsychiatric symptoms predict faster cognitive decline inAlzheimer’s disease. Arch. Neurol. 51, 676–681.
Dragonetti, A., Baldassarre, M., Castino, R. The lysosomal pro-tease cathepsin D is efficiently sorted to and secreted fromregulated secretory compartments in the rat basophilic/mastcell line RBL. J. Cell Sci. 113, 3289–3298.
Edwards, A., Gillard, M., Merler, E., 1989. Characterization ofmuscarinic receptors: type M2 (subtype B) on neuro-2A neu-roblastoma cells. J. Recept. Res. 9, 259–270.
Ehehalt, R., Keller, P., Haass, C. Amyloidogenic processing of theAlzheimer b-amyloid precursor protein depends on lipid rafts.J. Cell Biol. 160, 113–123.
Ekkapongpisit, M., Giovia, A., Nicotra, G. Labeling and exocytosisof secretory compartments in RBL mastocytes by polystyreneand mesoporous silica nanoparticles. Int. J. Nanomed. 7,1829–1840 2012.
Follo, C., Castino, R., Nicotra, G. Folding, activity and targeting ofmutated human cathepsin D that cannot be processed intothe double-chain form. Int. J. Biochem. Cell Biol. 39, 638–649.
Fukumori, A., Okochi, M., Tagami, S. Presenilin-dependentgamma-secretase on plasma membrane and endosomes isfunctionally distinct. Biochemistry 45, 4907–4914.
Good, M.I., Shader, R.I., 1982. Lethality and behavioral side effectsof chloroquine. J. Clin. Psychopharmacol. 2, 40–47.
Haass, C., Lemere, C.A., Capell, A. The Swedish mutation causesearly-onset Alzheimer’s disease by beta-secretase cleavagewithin the secretory pathway. Nat. Med. 1, 1291–1296.
Higaki, J., Catalano, R., Guzzetta, A.W. Processing of beta-amyloidprecursor protein by cathepsin D. J. Biol. Chem. 271, 31885–31893.
Hook, V.Y., Kindy, M., Reinheckel, T. Genetic cathepsin B defi-ciency reduces beta-amyloid in transgenic mice expressinghuman wild-type amyloid precursor protein. Biochem. Bio-phys. Res. Commun. 386, 284–288.
Isidoro, C., Horst, M., Baccino, F.M. Differential segregation ofhuman and hamster cathepsin D in transfected baby-hamsterkidney cells. Biochem. J. 273, 363–367.
Jellinger, K.A., 2003. Alpha-synuclein pathology in Parkinson’sand Alzheimer’s disease brain: incidence and topographicdistribution—a pilot study. Acta Neuropathol. 106, 191–201.
Kang, J., Muller-Hill, B., 1990. Differential splicing of Alzheimer’sdisease amyloid A4 precursor RNA in rat tissues: PreA4(695)mRNA is predominantly produced in rat and human brain.Biochem. Biophys. Res. Commun. 166, 1192–1200.
Kinoshita, A., Fukumoto, H., Shah, T. Demonstration by FRET ofBACE interaction with the amyloid precursor protein at thecell surface and in early endosomes. J. Cell Sci. 116, 3339–3346.
Kojima, N., Kurosawa, N., Nishi, T. Induction of cholinergicdifferentiation with neurite sprouting by de novo biosynthesisand expression of GD3 and b-series gangliosides in Neuro2acells. J. Biol. Chem. 269, 30451–30456.
Koo, E.H., Squazzo, S.L., 1994. Evidence that production andrelease of amyloid beta-protein involves the endocytic path-way. J. Biol. Chem. 269, 17386–17389.
Koo, E.H., Squazzo, S.L., Selkoe, D.J. Trafficking of cell-surfaceamyloid beta-protein precursor. I. Secretion, endocytosis andrecycling as detected by labeled monoclonal antibody. J. CellSci. 109, 991–998.
Liu, R.Q., Zhou, Q.H., Ji, S.R. Membrane localization of beta-amyloid 1–42 in lysosomes: a possible mechanism for lyso-some labilization. J. Biol. Chem. 285, 19986–19996.
Lo, A.C., Haass, C., Wagner, S.L. Metabolism of the ‘‘Swedish’’amyloid precursor protein variant in Madin–Darby caninekidney cells. J. Biol. Chem. 269, 30966–30973.
Lorenzen, A., Samosh, J., Vandewark, K. Rapid and direct trans-port of cell surface APP to the lysosome defines a novelselective pathway. Mol. Brain 3, 11.
Luo, Y., Bolon, B., Kahn, S. Mice deficient in BACE1, the Alzheimer’sbeta-secretase, have normal phenotype and abolished beta-amyloid generation. Nat. Neurosci. 4, 231–232.
Mortimer, J.A., Ebbitt, B., Jun, S.P. Predictors of cognitive andfunctional progression in patients with probable Alzheimer’sdisease. Neurology 42, 1689–1696.
Mueller-Steiner, S., Zhou, Y., Arai, H. Antiamyloidogenic andneuroprotective functions of cathepsin B: implications forAlzheimer’s disease. Neuron 51, 703–714.
O’Brien, R.J., Wong, P.C., 2011. Amyloid precursor protein processingand Alzheimer’s disease. Annu. Rev. Neurosci. 34, 185–204.
Ohno, M., Sametsky, E.A., Younkin, L.H. BACE1 deficiency rescuesmemory deficits and cholinergic dysfunction in a mousemodel of Alzheimer’s disease. Neuron 41, 27–33.
Pasternak, S.H., Callahan, J.W., Mahuran, D.J., 2004. The role of theendosomal/lysosomal system in amyloid-beta production andthe pathophysiology of Alzheimer’s disease: reexamining thespatial paradox from a lysosomal perspective. J. AlzheimersDis. 6, 53–65.
Perdivara, I., Petrovich, R., Allinquant, B. Elucidation of O-glyco-sylation structures of the beta-amyloid precursor protein byliquid chromatography-mass spectrometry using electrontransfer dissociation and collision induced dissociation. J.Proteome Res. 8, 631–642.
Rajendran, L., Honsho, M., Zahn, T.R. Alzheimer’s disease beta-amyloid peptides are released in association with exosomes.Proc. Natl. Acad. Sci. U.S.A. 103, 11172–11177.
Sabuncu, M.R., Desikan, R.S., Sepulcre, J. Alzheimer’s disease neuroi-maging initiative. The dynamics of cortical and hippocampalatrophy in Alzheimer disease. Arch. Neurol. 68, 1040–1048.
Saftig, P., Peters, C., von Figura, K. Amyloidogenic processing ofhuman amyloid precursor protein in hippocampal neuronsdevoid of cathepsin D. J. Biol. Chem. 271, 27241–27244.
Savage, M.J., Trusko, S.P., Howland, D.S. Turnover of amyloid beta-protein in mouse brain and acute reduction of its level byphorbol ester. J. Neurosci. 18, 1743–1752.
Scarmeas, N., Hadjigeorgiou, G.M., Papadimitriou, A. Motor signsduring the course of Alzheimer disease. Neurology 63, 975–982.

b r a i n r e s e a r c h 1 4 7 1 ( 2 0 1 2 ) 1 0 2 – 1 1 7 117
Schechter, I., Ziv, E., 2008. Kinetic properties of cathepsin D andBACE 1 indicate the need to search for additional beta-secretase candidate(s). Biol. Chem. 389, 313–320.
Schneider, J.A., Bienias, J.L., Gilley, D.W. Improved detection ofsubstantia nigra pathology in Alzheimer’s disease. J. Histo-chem. Cytochem. 50, 99–106.
Schrader-Fischer, G., Paganetti, P.A., 1996. Effect of alkalizingagents on the processing of the beta-amyloid precursorprotein. Brain Res. 716, 91–100.
Sinha, S., Lieberburg, I., 1999. Cellular mechanisms of beta-amyloid production and secretion. Proc. Natl. Acad. Sci.U.S.A. 96, 11049–11053.
Thinakaran, G., Koo, E.H., 2008. Amyloid precursor protein trafficking,processing, and function. J. Biol. Chem. 283, 29615–29619.
Thinakaran, G., Teplow, D.B., Siman, R. Metabolism of the ‘‘Swedish’’amyloid precursor protein variant in neuro2a (N2a) cells.Evidence that cleavage at the ‘‘beta-secretase’’ site occurs inthe golgi apparatus. J. Biol. Chem. 271, 9390–9397.
Tian, G., Sobotka-Briner, C.D., Zysk, J. Linear non-competitiveinhibition of solubilized human gamma-secretase by pepstatinA methylester, L685,458, sulfonamides, and benzodiazepines.J. Biol. Chem. 277, 31499–31505.
Trincheri, N.F., Follo, C., Nicotra, G. Resveratrol-induced apoptosisdepends on the lipid kinase activity of Vps34 and on theformation of autophagolysosomes. Carcinogenesis 29, 381–389.
Trincheri, N.F., Nicotra, G., Follo, C. Resveratrol induces cell deathin colorectal cancer cells by a novel pathway involvinglysosomal cathepsin D. Carcinogenesis 28, 922–931.
Van Gool, W.A., Weinstein, H.C., Scheltens, P. Effect of hydroxy-chloroquine on progression of dementia in early Alzheimer’sdisease: an 18-month randomised, double-blind, placebo-controlled study. Lancet 358, 455–460.
Vassar, R., Bennett, B.D., Babu-Khan, S. Beta-secretase cleavage ofAlzheimer’s amyloid precursor protein by the transmembraneaspartic protease BACE. Science 286, 735–741.
Vieira, S.I., Rebelo, S., Esselmann, H. Retrieval of the Alzheimer’samyloid precursor protein from the endosome to the TGN isS655 phosphorylation state-dependent and retromer-mediated. Mol. Neurodegener. 5, 40.
Vincent, B., Govitrapong, P., 2011. Activation of the a-secretaseprocessing of AbPP as a therapeutic approach in Alzheimer’sdisease. J. Alzheimers Dis. 24 (2), 75–94.
Wilson, R.S., Bennett, D.A., Gilley, D.W. Progression of parkinso-nian signs in Alzheimer’s disease. Neurology 54, 1284–1289.
Wolfe, M.S., Haass, C., 2001. The role of presenilins in gamma-secretase activity. J. Biol. Chem. 276, 5413–5416.
Xu, H., Sweeney, D., Wang, R. Generation of Alzheimer beta-amyloid protein in the trans-Golgi network in the apparentabsence of vesicle formation. Proc. Natl. Acad. Sci. U.S.A. 94,3748–3752.
Yamazaki, T., Koo, E.H., Selkoe, D.J., 1996. Trafficking of cell-surfaceamyloid beta-protein precursor. II. Endocytosis, recycling andlysosomal targeting detected by immunolocalization. J. Cell Sci.109, 999–1008.
Zhang, Y.W., Thompson, R., Zhang, H. APP processing in Alzheimer’sdisease. Mol. Brain 4, 3.




























