Embed Size (px)
Citation preview

ARTICLE IN PRESS
European Journal of Cell Biology 85 (2006) 785–802
0171-9335/$ - se
doi:10.1016/j.ej
Abbreviations
fluorescein thio
STS, staurospo
tetramethylroda�Correspond
Tel.: +49234 3
E-mail addr
www.elsevier.de/ejcb
Dual effects of staurosporine on A431 and NRK cells: Microfilament dis-
assembly and uncoordinated lamellipodial activity followed by cell death
Hans G. Mannherza,b,�, Sabine M. Gonsiorb, Xueqing Wua, Bernhard Polzara,Brian J. Popeb, Lena Wartoschc, Alan G. Weedsb
aDepartment of Anatomy and Embryology, Ruhr-University, Bochum, GermanybMRC Laboratory of Molecular Biology, Cambridge, UKcDepartment of Cell Biology, Institute of Zoology, Technical University of Braunschweig, Germany
Received 26 August 2005; received in revised form 9 February 2006; accepted 13 February 2006
Abstract
The general protein kinase inhibitor staurosporine (STS) has dual effects on human epidermoid cancer cells (A431)and normal rat kidney fibroblasts (NRK). It almost immediately stimulated increased lamellipodial activity of both celllines and after 2 h induced typical signs of apoptosis, including cytoplasmic condensation, nuclear fragmentation,caspase-3 activation and DNA degradation. In the early phase we observed disruption of actin-containing stress fibresand accumulation of monomeric actin in the perinuclear region and cell nucleus. Increased lamellipodial-likeextensions were observed particularly in A431 cells as demonstrated by co-localisation of actin and Arp2/3 complex,whereas NRK cells shrunk and exhibited numerous thin long extensions. These extensions exhibited uncoordinatedcentrifugal motile activity that appeared to tear the cells apart. Both cofilin and ADF were translocated fromperinuclear regions to the cell cortex and, as expected in the presence of a kinase inhibitor, all the cofilin wasdephosphorylated. Myosin II was absent from the extensions, and a reduction of phosphorylated myosin light chainswas observed within the cytoplasm indicating myosin inactivation. Microtubules and intermediate filaments retainedtheir characteristic filamentous organisation after STS exposure even when the cells became rounded and disorganised.Simultaneous treatment of NRK cells with STS and the caspase inhibitor zVAD did not inhibit the morphological andcytoskeletal changes. However, the cells underwent cell death as verified by positive annexin-V-staining. Thus it seemslikely that cell death induced by STS may not only be a consequence of the activation of caspase, instead the disruptionof the many motile processes involving the actin cytoskeleton may by itself suffice to induce caspase-independent celldeath.r 2006 Elsevier GmbH. All rights reserved.
Keywords: Actin; A431 cells; Apoptosis; ADF/cofilin; NRK cells; Tubulin; Vimentin
e front matter r 2006 Elsevier GmbH. All rights reserved.
cb.2006.02.012
: ADF, actin-depolymerising factor; MLCK, myosin light chain kinase; DTT, dithiothreitol; FCS, foetal calf serum; FITC,
isocyanate; Hepes; 4-(hydroxyethyl)-1-piperazineethanesulfonic acid; IF, intermediate filaments; PBS, phosphate-buffered saline;
rine; TBS, Tris-buffered saline; TTBS, TBS with 0.05% Tween-20; Tris, Tris-(hydroxymethyl)-amino methane; TRITC,
mine isothiocynate; P-MLC, phosphorylated myosin light chain; zVAD, z-valine-alanine-aspartate-fluoromethylketone
ing author. Abteilung fur Anatomie und Embryologie, Ruhr-Universitat Bochum, Universitatsstr. 150, D-44780 Bochum, Germany.
22 4553; fax: +49 234 321 4474.
ess: [email protected] (H.G. Mannherz).

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802786
Introduction
The morphology of eukaryotic cells is maintained bythe intracellular cytoskeleton. The microfilament systemplays an important role in the control of cell shape andthe motility of migrating cells (Pollard and Borisy, 2003;DesMarais et al., 2005). Cellular tensile strength isprovided by intermediate filaments, whereas the spatialorganisation of organelles and vesicular trafficking isperformed by microtubules and associated motorproteins. Locomotion of cells has been shown to beindependent of microtubules (Euteneuer and Schliwa,1984). The organisation of these filamentous systems ishighly dynamic allowing rapid adaptation to changingphysiological needs.
A number of drugs, mostly of plant origin, interferewith the organisation of the cytoskeleton. One such drugis staurosporine (STS), a general inhibitor of proteinkinases. STS is commonly used to induce apoptosis ofcells in culture, but the mechanism by which this occursis not well understood. Apoptosis is a special form ofprogrammed cell death characterised by well-defined,dramatic changes in cell shape (Kerr et al., 1972). Thecells first shrink by condensation of both cytoplasm andnucleus leading to rounding. Thereafter motile cytoplas-mic protuberances – apoptotic blebs – are formed whichpinch off as ‘‘apoptotic bodies’’ often including nuclearfragments. A cascade of specific cysteinyl aspartateproteases (caspases) is initiated that leads to theactivation of a number of executor caspases, in particularcaspase-3 (for review see Earnshaw et al., 1999). Thesecaspases cleave cytoplasmic and nuclear proteins that areessential for cell survival. Apoptosis is accompanied bychromatin condensation and further enhanced by itsfragmentation catalysed by endonucleases (Wyllie, 1980;Peitsch et al., 1994; Enari et al., 1998).
The morphological changes occurring during apop-tosis are accompanied by extensive changes of thecytoskeleton. The microfilament system disassembles(for a review see also Mills et al., 1999), although thedetailed mechanisms are not established. It has beenproposed that microfilaments are depolymerised eitherby F-actin-severing proteins like gelsolin or a shift of theG-/F-actin equilibrium due to actin cleavage by caspases(Mashima et al., 1997). Although actin proteolysisduring apoptosis is still controversial (Mashima et al.,1997; Hengstenberg and Mannherz, 1998; Song et al.,1997), a number of actin-binding proteins were shown tobe substrates of caspase-3, including gelsolin (Kothako-ta et al., 1997), a-fodrin (non-erythrocyte spectrin;Janicke et al., 1998) and plectin (Stegh et al., 2000).Recent data have shown that the dissolution of themicrofilament system can be elicited either by thecaspase-3-generated N-terminal fragment of gelsolin(Kothakota et al., 1997) or by activation of gelsolinitself (Paddenberg et al., 2001). Further candidate actin-
binding proteins that modify its state of polymerisationare the members of the cofilin/ADF (actin-depolymeris-ing factor) family, which also sever F-actin (Maciveret al., 1998). The actin-binding activity of cofilin/ADF isinhibited by phosphorylation of serine3 (for a review seeBamburg, 1999) that is catalysed by LIM-kinase 1(Arber et al., 1998; Yang et al., 1998). Dephosphoryla-tion of phospho-cofilin and -ADF by specific phospha-tases (like Slingshot) restores their actin-binding and -depolymerising activity (Niwa et al., 2002).
Here we describe the effects of STS on two cells types:A431 and NRK cells. Both cell types grow toconfluence, but while A431 cells grow in clusters thusmore resembling epithelial sheets, NRK grow as singlecells like fibroblasts and they show a well-developedstress fibre system. After treatment with STS for a fewminutes, changes in their morphology became evidentthat were correlated with changes in the organisation ofthe cytoskeleton. Our results demonstrate a selectivedisassembly of stress fibres, in particular in thefibroblastic NRK cell line, while microtubules andintermediate filaments persisted even during the ensuingapoptotic cell shrinkage. As immediate effect of STS wealso observed an increased and often uncoordinatedlamellipodial activity that appeared to tear the cellsapart. After 2 h of STS exposure classical morphologicalsigns of apoptosis became visible.
Increased lamellipodial activity also depends onADF/cofilin activation. We therefore analysed thedistribution of ADF and cofilin using specific antibodiesbefore and after STS exposure, since both proteins areexpected to be fully active due to LIM kinase inhibitionby STS. Mammalian cofilin shares about 70% sequenceidentity with ADF. Both proteins are co-expressed inepithelia and many cell lines (Hotulainen et al., 2005)and disassemble actin filaments in a pH-dependentfashion, but ADF shows a greater activity than cofilin atpH values greater than 7 (Yeoh et al., 2002). ADF andcofilin were found to be fully dephosporylated andtherefore activated and found redistributed to lamelli-podial areas together with Arp2/3 complex.
The morphological changes typical of classical apop-tosis were largely suppressed by inclusion of the caspaseinhibitor zVAD. Nevertheless a caspase-independentform of cell death still occurred suggesting that celldeath can be induced solely by disruption of the manymotile processes involving the actin cytoskeleton.
Materials and methods
Materials
Foetal calf serum (FCS) and media were obtainedfrom Gibco (Karlsruhe/Germany). FITC-labelled an-

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 787
nexin-V (Annexin-V-Fluos) was from Boehringer-Mannheim (Germany). Monoclonal antibodies againstmyosin, tubulin, vimentin, TRITC-phalloidin and STSwere purchased from Sigma (Deisenhofen, Germany).The monoclonal anti-b-actin antibody (clone AC74) waspurchased from Sigma (Poole, Dorset, UK) and FITC-labelled anti-rabbit IgG from Amersham (AmershamLife Science/UK). The M30 antibody directed againstthe caspase-3 cleavage product of cytokeratin 18 waspurchased from Boehringer-Mannheim (Germany).Polyclonal anti-Arp2/3 complex antibody directedagainst the ArpC1 subunit was kindly provided byDr. H.N. Higgs (University of Dartmouth, NewHampshire, USA). The antibody against the cytoplas-mic myosin II light chain phosphorylated at Thr 18 andSer19 was purchased from Santa Cruz Biotechnologies(Santa Cruz, California, USA). Monoclonal anti-Tauand -human-phospho-Tau were generous gifts fromProf E.-M. Mandelkow (Hamburg, Germany) and ElanPharmaceuticals (San Francisco, California, USA),respectively. Alexa-488-labelled DNase I and the chro-matin stain TOTO-3 were obtained from MolecularProbes (Eugene, Oregon, USA). The pan-caspaseinhibitor zVAD was obtained form Bachem (Weil amRhein, Germany). Nitrocellulose sheets for Westernblotting (BA 83) were from Schleicher and Schull(Dassel, Germany). All other reagents were of analyticalgrade.
Cell culture and induction of apoptosis
Culture of NRK and human A431 and HeLa cells wasperformed as described previously (Mannherz et al.,2005). The cells were fixed for immunofluorescence orharvested for biochemical analysis at various timepoints.
Polyclonal antibodies specific for cofilin and ADF
Polyclonal antibodies were raised in rabbits againsteither purified recombinant human ADF, cofilin or thecofilin:actin complex. Cofilin:actin complex was pre-pared by mixing F-actin at substoichiometric ratios atpH 8.0. Residual F-actin was pelleted and the super-natant containing cofilin:actin complex used for im-munisation. The use of complex as antigen was tofacilitate a response to these highly conserved proteinsby exposing epitopes that would not exist whenuncomplexed (Polzar et al., 1989). The antibodies wereaffinity purified on Sepharose-cofilin or Sepharose-ADFcolumns using 0.15M glycine-HCl, pH 2.5, to elute theantibodies. Then they were immediately neutralised withTris–HCl, pH 8.0, and dialysed against PBS containing1mM NaN3. The antibody specificity was tested byWestern blotting using purified human recombinant
cofilin and ADF. The results showed that the anti-cofilinwas monospecific, but there was a residual low level ofcross-reactivity for the anti-ADF (Fig. 1A).
Detection of surface phosphatidylserine by FITC-
annexin-V binding
A431 cells were grown for 24 h on sterile coverslips,washed twice with PBS, once with incubation buffer(10mM Hepes-OH, pH 7.4, 140mM NaCl, 5mMCaCl2) then incubated with labelling solution (Annex-in-V-Fluos diluted 1:50 in incubation buffer and 1 mg/mlpropidium iodide) for 15min. Subsequently the cellswere washed in PBS, fixed with 4% formaldehyde and mounted with Mowiol 88 (Hoechst, Frankfurt,Germany).
Fluorescence microscopy
A431 and NRK cells were grown on sterile glasscoverslips in DMEM with added NCS and FCSfor at least 24 h prior to use. Cells were fixed with 4%formaldehyde for 20min, permeabilised with 0.2%Triton X-100 in PBS for 5–10min or ice-cold methanolfor 5min and subsequently washed three times inPBS. Immunohistochemistry was performed as de-scribed previously (Mannherz et al., 2005). Im-munofluorescence microscopy was performed on anMRC-1024 Laser Scanning Confocal Imaging System(BioRAD, Cambridge, UK) or a Zeiss LSM 510 (Jena,Germany). To analyse Hoechst 33342-stained cells, aNikon Diaphot inverted microscope with a PrincetonInstruments Micromax 1300Y 1Mhz CCD cameraor a Zeiss Axiophot microscope equipped with anAxioCam HRc digital camera were used with a DAPI-filter. TOTO-3-stained cells were analysed by confocalmicroscopy.
Transmission electron microscopy
A431 cells were cultured on 75-cm2 culture dishes,washed with PBS, then scraped off with a rubber spatulaand centrifuged at 800g for 10min. The pellet was fixedfor 2 h in Karnovsky buffer (0.1M sodium cacodylate,pH 7.4, 2% paraformaldehyde, 2.5% glutaraldehyde).After a second centrifugation step the pellet was post-fixed in Dalton buffer (5% K2Cr2O7, 3.4% NaCl, 1%OsO4, pH 7.5) and embedded in Epon resin as specified(Fluka, Neu-Ulm/Germany). Ultrathin sections (60 nm)were treated with 1% uranyl acetate and 0.66% leadacetate (10min each). EM images were taken at 80 kVon a Philips EM 420 (Philips, Eindhoven, The Nether-lands).

ARTICLE IN PRESS
Fig. 1. Specificity of the affinity-purified polyclonal anti-ADF and -cofilin antibodies. (A) Immunoblots of purified cofilin and
ADF: Upper gel shows a Coomassie-stained 15% polyacrylamide gel after application of increasing amounts (0.5, 1, 2, and 5 mg of
recombinant human cofilin or ADF. Middle and lower gels show immunoblots with anti-cofilin and -ADF, respectively, under
identical conditions. (B) Immunoblots of A431 or NRK cell homogenates (50 mg protein each) probed with anti-ADF (1st gel) or
anti-cofilin (2nd gel, showing also markers of purified cofilin and ADF). Note the higher expression of ADF in NRK cells. (C–F)
Immunostaining of NRK cells with affinity-purified anti-cofilin (C, red) and anti-ADF (E, red). FITC-phalloidin staining of F-actin
overlayed on cofilin (D) and ADF (F). Bar: 20mm.
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802788
Preparative procedures
Rabbit skeletal muscle actin was purified fromacetone-dried powder as detailed previously (Mannherzet al., 1980). Human cofilin and ADF were expressed inEscherichia coli and purified as described by McGoughet al. (1997). Cell homogenates were prepared from cellsgrown in the presence or absence of FCS. Cells werewashed three times with PBS containing 0.1mM PMSF,scraped off, then broken by three cycles of freezing andthawing and subsequently centrifuged for 10min at15,000 g. Protein concentration of the supernatants wasdetermined as described by Bradford (1976) with bovineserum albumin as standard.
Determination of caspase-3 activity
Cells were collected by centrifugation, washed withPBS, re-suspended in 5mM Tris–HCl, pH 8.0, 5mMDTT and disrupted by three cycles of freezing and
thawing. Homogenates were centrifuged for 20min at17,530 g and the protein concentration of the super-natants (‘‘high speed homogenates’’) was measured(Bradford, 1976). For the caspase-3 fluorogenic enzymeassay, 30 mg of these homogenates were incubated with50 mM DEVD-AMC (Alexis, Grunberg, Germany)dissolved in 600 ml 20mM Hepes-OH, pH 7.5, 10%glycerol, 5mM DTT for 5 h at 37 1C. The AMC releasewas determined by measuring the fluorescence increasewith excitation and emission wavelengths of 380 and460 nm, respectively, using a Shimadzu RF 5001-PCspectrofluorometer as described by Stolzenberg et al.(2000).
Electrophoretic procedures
Cell homogenates were separated on 10–20% acryl-amide gradient gels (Matsudaira and Burgess, 1978;Laemmli, 1970) and transferred to nitrocellulose sheetsusing a semi-dry blotter. The nitrocellulose sheets were

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 789
stained with 0.2% Ponceau-S in 10% acetic acid tomonitor the success of transfer, incubated with 3% fishgelatine (Serva, Heidelberg, Germany) in TTBS for 1 hat room temperature and then blocked with 3%skimmed milk powder at 4 1C in TTBS overnight. Theblots were incubated with polyclonal anti-cofilin or anti-ADF in TTBS containing 0.5% skimmed milk powderovernight at 4 1C, washed 3 times in TTBS, thenincubated with horseradish peroxidase-conjugated goatanti-rabbit or goat-anti-mouse antibodies (1:3000). Theenhanced chemiluminescence (ECL) system (Amersham,Braunschweig, Germany) was used with exposure timesbetween 20 and 120 s.
Two-dimensional gel electrophoresis was performedon 106 cells washed with PBS and homogenised in 500 mlsample buffer containing 9M urea, 2% (w/v) ampholine(pH 3.5–10), 2% (v/v) Triton X-100 and 20mM DTTand centrifuged for 10min at 16,000g. The supernatantswere stored at �20 1C before processing on pH 3.5–10immobiline gradient strips (IPGs, Pharmacia, Freiburg,Germany) according to Wu et al. (2000). The gels werefixed and transferred to nitrocellulose for immunoblot-ting as described above.
Results
Differential localisation of ADF and cofilin in NRK
cells
Fig. 1B gives the levels of expression of cofilin andADF in NRK and A431 cells. Cofilin expression isroughly similar in both cell types, but there isconsiderably more ADF present in NRK cells than inA431 cells (Fig. 1B). We therefore looked to see whetherthere was any differential localisation of the twoproteins in NRK cells. Both cofilin and ADF werepreferentially located around the nucleus, but cofilinoften showed a nuclear distribution in addition (Fig. 1Cand D vs E and F). Staining of F-actin with FITC-phalloidin highlighted the differences in the nuclearcontent between cofilin and ADF (Fig. 1D and E), but itshould be noted that phalloidin does not bind tofilaments decorated with cofilin or ADF (Moon et al.,1993), so this does not mean that actin is absent fromthese nuclei. Indeed nuclear actin has been demon-strated by others using anti-actin antibodies (Gonsioret al., 1999).
STS promotes stress fibre disassembly, shape
changes and redistribution of actin in NRK cells
NRK cells showed the presence of stress fibres whenstained with TRITC-phalloidin (Fig. 2A), while mono-meric actin was concentrated in the perinuclear region as
visualised by Alexa 488-labelled-DNase I (Fig. 2B).Within 10min of exposure to 2 mM STS, the stress fibreshad disappeared (Fig. 2D). Disassembly of the stressfibres was accompanied by accumulation of the remain-ing F-actin (phalloidin stainable) at the cell periphery(Fig. 2D) and of monomeric actin around and within thecell nucleus (Fig. 2E). After 30min, the cells wereshrunken and adopted a star-like appearance with manyremaining thin and often branched extensions thatstained with TRITC-phalloidin (Fig. 2G). Monomericactin was restricted to cell nuclei (Fig. 2H) as shown alsoby counterstaining with Hoechst 33342 (Fig. 2I).
STS induces increased but uncoordinated
lamellipodial activity in A431 cells
A431 cells grow in clusters. When treated with 0.5 mMSTS we observed an increased lamellipodial activity ofthe cells at the periphery of the clusters (arrows inFig. 3B–D). Time-lapse analysis demonstrated that thediameter of the cell clusters increased with time(Fig. 3A–D). Particularly, the peripheral cells exhibiteda slow outward movement with the formation of verybroad lamellipodial structures that after 2–3 h adopted afan-like structure connected to the cell body by onlysmall stalks (clearly visible in Fig. 3J). A431 in contrastto NRK cells did not exhibit a strong cytoplasmicshrinkage. In addition, a decrease in intercellularcontacts was observed leading to broader intercellularspaces. These morphological changes also occurred atconsiderably lower STS concentrations. Even at 1 nMSTS increased lamellipodial activity was observedthough it then lasted at least 5 h till they became evident(see Fig. 8A).
To verify the lamellipodial nature of the describedextensions we stained A431 cells with a polyclonal anti-Arp2/3 complex antibody as previously described(Mannherz et al., 2005). In control cells, Arp2/3complex was mainly concentrated in the perinuclearregion of both cell lines and only weakly at the cellperiphery (shown for A431 cells in Fig. 3E), although atthe peripheral region presumably cortical actin wasstrongly stained by TRITC-phalloidin (Fig. 3F). AfterSTS treatment, Arp2/3 complex was found concentratedat the distal end of (arrow in Fig. 3H) and within thebroad lamellipodial structures (arrow in Fig. 3K),although a large proportion remained in the perinuclearregion. Staining with TRITC-phalloidin resulted in analmost homogenous staining marking the broad STS-induced lamellipodial structures (Fig. 3I and L), co-localising with the distal and more punctate Arp2/3complex staining within these lamellipodia (Fig. 3J andM). Similar changes in Arp2/3 distribution wereobtained for NRK cells (not shown). These datasuggested that the cytoskeletal organisation of the

ARTICLE IN PRESS
Fig. 2. STS-induced stress fibre degradation, morphological changes and cofilin redistribution in NRK cells. Micrographs of NRK
cells before (A–C) and after treatment with 2mM STS for 10min (D–F) and 30min (G–I). Staining for F-actin with TRITC-
phalloidin (A, D, G); G-actin with Alexa 488-DNase I (B, E, H); and merged, showing also Hoechst 33342 nuclear stain (C, F, I).
Same magnification for all micrographs. Bar: 20mm.
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802790
STS-induced lamellipodial structures resembled that ofnormal lamellipodia in agreement with previous dataobtained for fish keratocytes after STS exposure(Verkhovsky et al., 1999).
STS induces redistribution of cofilin/ADF into the
extending lamellipodial structures
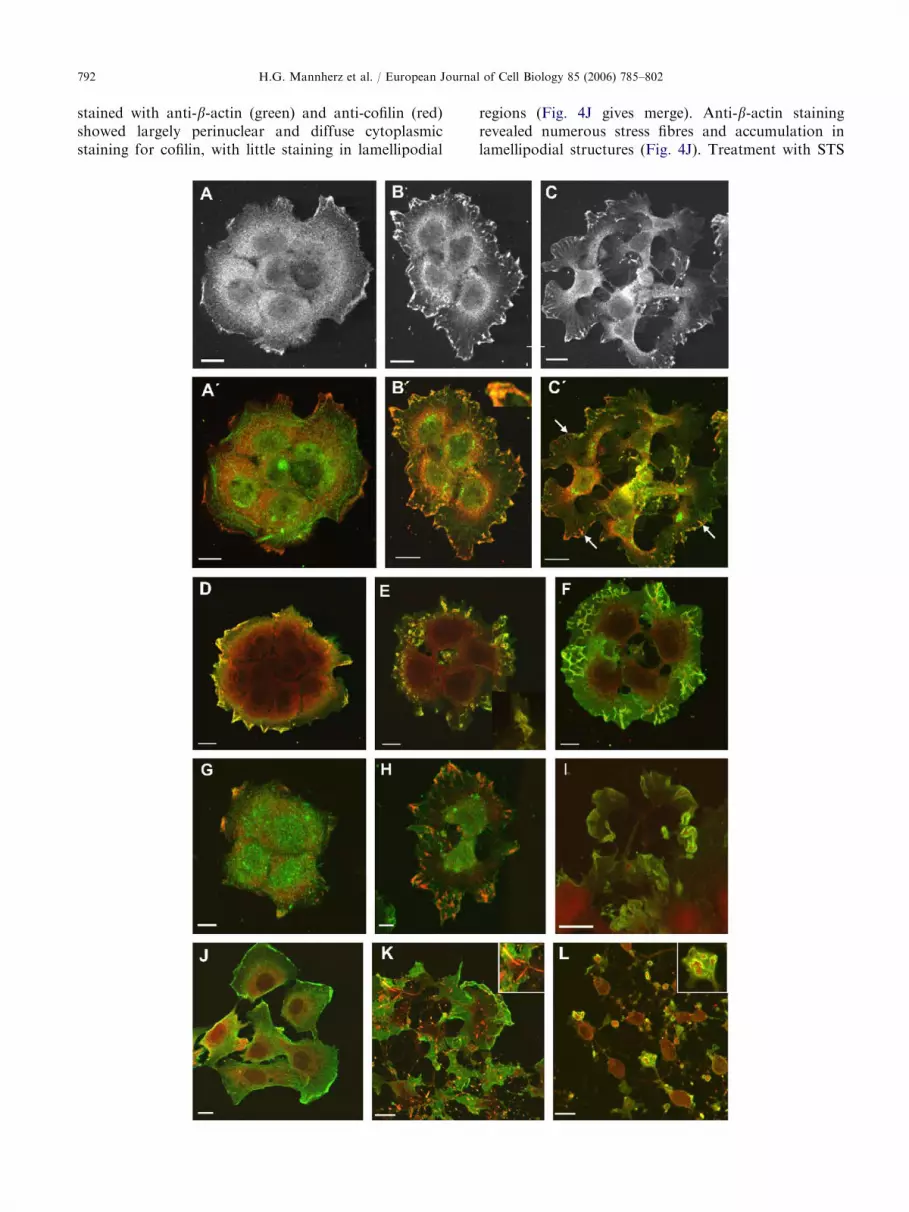
Disruption of the stress fibres would be expected afteractivation of severing proteins like cofilin and ADF.Most of the cofilin staining in resting A431 cells islocalised to the perinuclear region, with very littleconcentrated at the cell cortex (Fig. 4A and A0 red).Following exposure to 2 mM STS for up to 1 h, there wasgreatly increased concentration of cofilin in the periph-ery of the lamellipodia-like structures, while the peri-nuclear zone appeared less diffuse. Concomitantly weobserved increased protrusive activity of the peripheral
cells of the clusters (Fig. 4B and C) that was also evidentfrom their increased diameter as detailed above. Thestaining showed co-localisation of actin and cofilinwithin the surface projections (see inset Fig. 4B0). Afterlonger periods of STS exposure (shown for 4 h inFig. 4C), the cells adopted a star-like appearancecharacterised by nuclei surrounded by a thin rim ofcytoplasm that elongated into broad lamellipodia. Theperiphery of these lamellipodia stained for cofilin andactin with stronger cofilin staining in ‘‘streaks’’ perpen-dicular to the membrane (Fig. 4C0 arrows).
After fixation we also permeabilised A431 cells withmethanol (Fig. 3D–F), because it was reported thatdifferent permeabilisation conditions lead to differentmorphologies and the exposure of different epitopes,especially of actin (Gonsior et al., 1999). Control cellslooked very similar under both extraction conditions (cf.Fig. 4A with D). However, after 1 h STS treatment weobserved better preservation of small blebs and dorsal

ARTICLE IN PRESS
Fig. 3. STS-induced increased lamellipodial activity in A431 cells. (A–D) Time-lapse recording of effect of STS on the lamellipodial
activity of the peripheral cells of an A431 cluster during the exposure to 0.5 mM STS. Regions with increased lamellipodial activity
are marked by arrows. (A) t ¼ 0; (B) t ¼ 30 min; (C) T ¼ 120 min, and (D) t ¼ 180 min. (E–M) Staining of A431 cells with anti-
Arp2/3 complex (E, H, K) and TRITC-phalloidin (F, I, L) before (E–G) and after treatment with 1 mM STS for 2 h (H–J) and 4 h
(K–M). (G, J, M) merge. Bars: 50mm (D) and 20mm (G, J, M).
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 791
ruffles, where actin appeared more distal and cofilinmore proximal (Fig. 4E and inset, see also Fig. 4L). Thisdistribution was also maintained 3 h after STS treatment(Fig. 4F). We assume that the dorsal ruffles in Fig. 4Eand F correspond to the ‘‘streaks’’ seen in Fig. 4C(arrows).
If cofilin plays a role in this process, we might expectsimilar behaviour for ADF. Immunofluorescencedemonstrates that like cofilin, ADF is diffusely cyto-plasmic in A431 cells showing very little co-localisationwith actin, (cf. Fig. 4G and A). Two hours after drug
exposure, its distribution had changed in a manneridentical to cofilin, i.e. co-localised with actin at the cellperiphery (Fig. 4H after Triton X-100 permeabilisation)and was detected in blebs and ruffles after methanolpermeabilisation with a more proximal distribution ofADF (red) in comparison to actin (green) (Fig. 4I;cf. with Fig. 4B0 and F, respectively, for cofilin).
In NRK cells, STS-exposure led to similar morpho-logical changes. However, the cytoplasmic extensionswere smaller and their connections to the cell body weremuch thinner (Fig. 4K and L). Control NRK cells
ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802792
stained with anti-b-actin (green) and anti-cofilin (red)showed largely perinuclear and diffuse cytoplasmicstaining for cofilin, with little staining in lamellipodial
regions (Fig. 4J gives merge). Anti-b-actin stainingrevealed numerous stress fibres and accumulation inlamellipodial structures (Fig. 4J). Treatment with STS

ARTICLE IN PRESS
Fig. 5. Immunoblots of two-dimensional gels of NRK cell
homogenates. Homogenates of NRK cells before (A) and after
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 793
for 2 h and permeabilisation with Triton X-100 demon-strated dramatic shape changes, disassembly of thestress fibres and the presence of numerous anti-actin-and anti-cofilin-positive cytoplasmic rods (see also insetin Fig. 4K). Methanol extraction revealed the formationof small membrane blebs and ruffles predominantly atthe ends of the star-like cytoplasmic extensions (Fig.4L). Cofilin was always localised more proximally andactin more distally within these membrane extensions(inset in Fig. 4L). The nuclei of these shrunken, star-likecells were surrounded by thin cytoplasmic rims, whichwere also immunopositive for cofilin (Fig. 4L). Thus inboth cells lines STS induces dramatic changes inmorphology characterised by cytoplasmic extensionsthat terminate in lamellipodial structures. NRK cellswere often surrounded by these lamellipodial extensionsthat appeared to migrate in opposing directions and totear the cells apart (Fig. 4L).
exposure (B) to 1 mM STS for 2 h. Only the anti-cofilin-reactive
region of the 2D-gel is shown (see also Mannherz et al., 2005).
Arrows in (A) mark (1) phospho-cofilin (or cofilin-1),
(2) phopho-m-cofilin (muscle type cofilin or cofilin-2), and
(3) unphosphorylated cofilin. After STS treatment only
unphosphorylated cofilin was detectable (B).
Cofilin dephosphorylation after STS treatment
The changes observed in the distribution of cofilinand ADF after STS exposure suggested their involve-ment in the observed uncoordinated lamellipodialmotility and therefore their activation by dephosphor-ylation of the regulatory serine3 residue. We suspectedinhibition of LIM kinase-1 by STS that should lead tocomplete dephosphorylation and hence activation ofADF/cofilin. Western blots of 2D gels of homogenatesof control NRK cells showed three anti-cofilin reactivebands (Fig. 5A): phospho-cofilin (arrow 1), phospho-m-cofilin (arrow 2) and dephosphorylated cofilin (seealso Mannherz et al., 2005). About 20% of the cofilinappeared to be phosphorylated. After 2 h exposure toSTS, all the cofilin was dephosphorylated (Fig. 5B).Similar observations were reported for A431 cells inresponse to STS: these cells showed a considerablyhigher percentage of phosphorylated cofilin prior totreatment (Mannherz et al., 2005).
Fig. 4. STS-induced redistribution of cofilin, ADF and actin in A43
with formaldehyde and permeabilised with Triton X-100 before (A;
C0), stained with anti-cofilin (A, B,C) and merged images (A0,B0,C0)
increased cofilin staining at the cell periphery and lamellipodia in
Peripheral streaks immunopositive for cofilin and actin became visibl
double stained for cofilin (red) and b-actin (green) before (D), 1 h aft
4% paraformaldehyde and permeabilised by a short treatment with
blebs and ruffles in (E) and (F). Inset to (E) shows an enlargement of
and more distal actin (green) distribution. (G–I) Similar cell cluster
actin (green). Control cells (G), 2 h after treatment with 2mM STS (H
(J–L) NRK cells stained with anti-cofilin; Control cells (J); note cytop
hours after exposure to 1mM STS. In (K) the cells were permeabilis
and the presence of numerous actin- and cofilin-containing rods
permeabilisation; note the cone-like structures at the ends of the cy
distal actin and a proximal cofilin immunoreactivity; see also inset.
STS leads to myosin II redistribution
We used an antibody against cytoplasmic myosin II toimmunostain NRK cells, which because of their well-developed stress fibres should give a clear correlationwith the microfilament system. Indeed, in resting NRKcells we observed punctate anti-myosin II staining alongstress fibres and around the nucleus (Fig. 6A–C). InA431 cells we observed myosin II staining additionally inproximal parts of lamellipodia (not shown). Afterexposing NRK cells for 1 h to 1mM STS, myosin IIwas localised entirely in the perinuclear region and,unexpectedly, absent from the large cytoplasmic exten-sions, which stained strongly for F-actin (Fig. 6D and F).
1 and NRK cells. Confocal images of A431 cell clusters, fixed
A0) and after addition of 2 mM STS for 1 h (B, B0) and 4 h (C,
of anti-cofilin (red) stained also with anti-b-actin (green). Note
(B) and (C), where it colocalised with actin (see inset in B0).
e after STS (B and marked by arrows in C0). (D–F) Cell clusters
er (E) and 3 h after (F) STS treatment. These cells were fixed in
ice-cold methanol. Note the preservation of numerous surface
one of the ruffles demonstrating the more proximal cofilin (red)
s double stained with anti-ADF (red) and monoclonal anti-b-,I) permeabilised with Triton X-100 (H) and with methanol (I).
lasmic and peri- and nuclear distribution of cofilin. (K, L) Two
ed with Triton X100; note elongated and star-like morphology
within the cytoplasm, see also inset in (K). (L) Methanol
toplasmic extensions containing numerous blebs with a more
Bars: 10 mm.

ARTICLE IN PRESS
Fig. 6. STS-induced changes in the distribution of myosin II. (A) Single NRK cell stained with anti-myosin II antibody; (B) with
TRITC-phalloidin, and (C) merge including nuclear staining with Hoechst 33342. (D–E) Two NRK cells after 1 h of 1 mM STS
exposure stained with anti-myosin II (D), note concentration of anti-myosin II immunoreactivity to the perinuclear region; TRITC-
phalloidin (E); and (F) merge including nuclear stain with Hoechst 33342. Staining of A431 cells with anti-P-MLC before (G–I) and
after exposure to 0.5mM STS for 4 h (J–L). Anti-P-MLC (green: G, J), TRITC-phalloidin (H, K) and merge (I, L). Bars: 10mm (F),
20 (I, L).
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802794
Thus the outward migration of these lamellipodialstructures does not appear to depend on myosin IImotors. Instead it could be due either solely to thepropulsive force generated by increased microfilamentreorganisation stimulated by activation of ADF/cofilin(Carlier et al., 1997; Pope et al., 2000) or to theinvolvement of other myosins.
We assumed that STS also inhibited myosin lightchain kinase (MLCK) leading to myosin II inactivation.To test this hypothesis, we immunostained NRK andA431 cells before and after treatment with 0.5 mM STSfor 4 h using an antibody specific for the phosphorylatedregulatory light chain (P-MLC). We obtained a clearreduction in the peripheral immunostain after STS

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 795
exposure of both cell lines (Fig. 6G–L). In resting NRK(not shown) and A431 cells the anti-P-MLC stainingwas punctate along microfilamentous bundles andconcentrated in the periphery of the A431 cells (Fig.6G and I). After STS exposure, the peripheral cyto-plasmic stain was decreased and almost absent in thelarge fan-like lamellipodial extensions, but still localisedaround and within the nucleus (Fig. 6J and L).
STS induces apoptosis in A431 and NRK cells
After longer periods of STS exposure, both A431 andNRK cells showed signs of apoptosis: control A431 cellsexhibited normal nuclear and chromatin morphology(Fig. 7B). Following STS exposure for 2 h many cellsexhibited punctate chromatin condensation (Fig. 7C).Using electron microscopy for higher resolution mor-phological analysis, semi-lunar chromatin deposits wereseen along the inner nuclear membrane typical ofapoptosis (Fig. 7D). Provided the STS exposure wasno longer than 2 h, removal of STS followed by arecovery period of 24 h resulted in complete disappear-ance of the apoptotic morphology (data not shown).Induction of apoptosis was verified by staining for thecaspase-3-specific cleavage product of cytokeratin 18(M30) (Fig. 7F–H), whereas there was no equivalentstaining in untreated cells (Fig. 7E). We often observedM30-positive apoptotic cells being engulfed by neigh-
Fig. 7. STS-induced apoptotic morphology in A431 cells. A431 cells
C) Staining with Hoechst 33342. Electron micrographs before (B) and
cells before (E) and after treatment with 2mM STS for 2 h (F), 3 h (
nuclear chromatin and with anti-M30 antibody (green) directed ag
absence of M30 staining in control cells (E), but positive M30 st
filamentous organisation of truncated cytokeratin 18. Nuclear fragm
10mm (A, C, E–H), 2mm (B, D).
bouring cells after 3 h (Fig. 7G and H). Further evidencefor apoptosis was obtained by a time-dependent increasein FITC-annexin-V staining of phosphatidylserineshifted to the outer leaflet of the plasma membraneduring apoptosis (see Fig. 11B). However, no nuclearstaining by propidium iodide (PI) was observed up to3 h, indicating intactness of the plasma membrane for atleast this period (not shown).
These data indicated that 1–2 mM STS inducedapoptosis of both cell lines after about 2 h exposure.However, the apoptosis-inducing effect of STS mighthave been due to its usage at these high concentrations.We therefore exposed both cell lines to 1, 5, 10 and50 nM STS for various time periods. Even at 1 nM STS,we observed morphological changes in many cells ofboth lines similar as described for 1 mM STS. After 5 h, aconsiderable number of A431 cells possessed condensednuclei of typical apoptotic morphology as verified byHoechst 33342 staining that were apparently phagocy-tosed by neighbouring cells (Fig. 8B). Similarly, after 5 hof exposure to 5 nM STS the majority of the A431 cellexhibited apoptotic chromatin condensation and frag-mentation (Fig. 8C).
Biochemical evidence for apoptosis
The caspase-3 activation seen in immunofluorescenceexperiments was confirmed in supernatants of cell
before (A, B) and 2 h after treatment with 2mM STS (C, D). (A,
2 h after (D) addition of STS. (E–H) Confocal images of A431
G), and 4 h (H). Cells were stained with Toto-3 (blue) to stain
ainst the caspase-3 cleavage product of cytokeratin 18. Note
aining (F–H) after addition of STS. Note too persistence of
entation and phagocytotic events are depicted in (G, H). Bars:

ARTICLE IN PRESS
Fig. 8. Treatment of A431 cells with nanomolar concentrations of STS. (A) Phase-contrast image of a A431 cluster after treatment
with 1 nM STS for 5 h. Note lamellipodial activity of the cells at the periphery of the cluster. (B, C) Apoptotic cells after 5 h
treatment with 1 nM STS (B, B0) and with 5 nM STS (C, C0). (B, C) staining with Hoechst 33342 and (B0, C0) phase-contrast images.
Note in (B) condensed nuclei surrounded by crescent nuclei of phagocytosing cells and (C) fragmented nucleus. Bars: 20 mm.
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802796
homogenates obtained from adherent A431 or NRKcells using acetyl–Asp–Glu–Val–Asp (DEVD) linked to7-amino-4-methylcoumarin as substrate. Control cellsshowed no or very little caspase-3 activity, but both celllines generated a significant increase in coumarinfluorescence after 3–4 h of STS exposure. Thus inA431 cells we observed a continuous increase incaspase-3 activity after initiation treatment with 2 mMSTS with a maximal activation of about 35-fold reachedafter 3 h (Fig. 9A). We also examined apoptosis-specificprotein degradation. Proteolysis of lamin B, which isspecifically cleaved by caspase-6 (Lazebnik et al., 1995),started after 1 h and was complete after 4 h STStreatment (Fig. 9D), but in contrast no degradation ofeither actin or cofilin was detected (Fig. 9B and C). Thusthe combined morphological and biochemical evidenceindicates that prolonged STS treatment results in theapoptotic demise of both A431 and NRK cells.
Fig. 9. Analysis of biochemical markers of apoptosis. (A)
Time-dependent activation of caspase-3 in A431 cells after
addition of 2mM STS as determined by the increasing
fluorescence signal of liberated AMC with time. Ordinate
gives fluorescence intensity (FI) in arbitrary units. (B–D)
Western blots of A431 cell homogenates to test for proteolysis
of actin (B), cofilin (C), and lamin B (D). (AC: purified rabbit
skeletal muscle actin, COF: purified recombinant human
cofilin as markers).
The STS-induced morphological changes are not
accompanied by disassembly of intermediate
filaments or microtubules
The M30 staining demonstrated the typical distribu-tion of intermediate filaments (IF) even after prolongedSTS treatment in A431 cells (see Fig. 7F–H). Thisassumption was verified by staining also NRK cells withanti-vimentin antibodies; again there was no indicationof IF depolymerisation after STS treatment even inrounded cells (Fig. 10A–F). Similarly staining NRKcells before and after exposure to 2 mM STS with anti-tubulin antibodies demonstrated the persistence offilamentous tubulin-positive structures (Fig. 10G–L)after 1 h even in rounded cells.
This apparent stability of the microtubular organisa-tion after STS exposure might be due to inhibition ofmicrotubular affinity regulating kinase (MARK). Un-phosphorylated Tau-protein remains firmly bound toMTs, strongly increasing their stability. MARK phos-phorylates Tau inducing its dissociation from MTs,thereby rendering them dynamically unstable. Using an
anti-Tau antibody that recognises Tau irrespective of itsstate of phosphorylation, we showed that some of theTau co-localised with the microtubular network incontrol cells but there was also considerable diffusecytoplasmic staining (Fig. 10M–O). There was nochange in Tau localisation after STS exposure for 2 h(Fig. 10P–R). We examined the state of Tau phosphor-ylation by using an anti-human phospho-Tau antibody.

ARTICLE IN PRESS
Fig. 10. STS induced no disassembly of intermediate filaments and microtubules. (A–C) Micrographs of control NRK cells stained
with TRITC-phalloidin (A), anti-vimentin (B), and (C) merge plus Hoechst 33342 staining. (D–F) Identical sequence of NRK cells
2 h after 2mM staurosporine exposure. Confocal micrographs of NRK cells before (G–I) and after treatment with 2 mM STS for
60min (J–L) stained with TRITC-phalloidin (G, J) and polyclonal antibody against tubulin (H, K), and merge (I, L) including
nuclear stain by TOTO-3. Micrographs of NRK cells before (M–O) and after exposure to 1mM staurosporine for 2 h (P–R). Stained
with anti-tubulin (M, P), anti-pan-Tau-protein (N, Q), and merge (O, R). Note perinuclear concentration of Tau-protein and co-
localisation with MTs. Staining of HeLa cells with anti-phospho-Tau-protein before (S) and after (U) exposure to 1mMstaurosporine for 5 h. Note staining of all control cells in (S) and almost complete loss of staining after staurosporine treatment (U).
(T, V) Corresponding phase-contrast images. Bars: 20mm.
H.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 797
Since this antibody does not stain rat NRK cells, weused HeLa cells, which possess a well-developedmicrotubular network, and observed a significant
decrease in phospho-Tau fluorescence after STSexposure (Fig. 10U and V) as compared to control cells(Fig. 10S and T).

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802798
The STS-induced morphological changes and cell
death are not prevented by caspase inhibition
In order to evaluate the direct effects of STS on thecytoskeleton, NRK cells exposed to STS were treatedsimultaneously with 40 mM zVAD, a general caspaseinhibitor for up to 4 h. Already after 2 h of treatment thecells showed the typical star-like appearance irrespectiveof the inclusion of zVAD (not shown). After 4 h, thetreated cells maintained the star-like appearance underboth conditions (Fig. 11B00 and C00), and in the absenceof zVAD many of the nuclei showed morphologicalchanges typical of apoptosis (Fig. 11B0). Many of thesecells were also annexin-V positive (Fig. 11B) in contrastto control cells (Fig. 11A). Typical apoptotic nuclearchanges were not observed in cells exposed simulta-neously to 40 mM zVAD, although their nuclei appeareddeformed and more condensed than those of controlcells (Fig. 11C0). Staining these cells with FITC-annexin-V revealed also positively stained cells though lessnumerous (Fig. 11C) suggesting caspase-independentcell death in agreement with Deas et al. (1998). Thepossibility of necrosis occurring was excluded by the factthat nuclei of the treated cells were not stained by themembrane impermeable propidium iodide (data notshown). These results confirm that the morphological
Fig. 11. Effects of the caspase inhibitor zVAD on cell and nuclear m
(B); and cells treated with 1 mM STS and 40 mM zVAD for 4 h (C). (A
and (A00–C00) phase contrast. Note positive annexin-V staining in (B)
did not inhibit changes in cell morphology (C00) and annexin-V stain
(C0). Bars: 20 mm.
and the underlying cytoskeletal alterations are notdependent on caspases.
Discussion
Differential localisation of ADF and cofilin in
control NRK cells
In this study we analysed the changes of thedistribution of ADF and cofilin and a number of othercytoskeletal proteins after STS treatment mainly usingthe anti-cofilin antibody, because the Western blotanalysis of our affinity-purified polyclonal anti-cofilinand anti-ADF showed absolute specificity for the anti-cofilin, but some residual cross-reactivity of the anti-ADF antibody with cofilin. Immunostaining NRK cellswith either antibody demonstrated certain differences inthe distribution of cofilin and ADF. While both ADFand cofilin showed predominantly perinuclear andcytoplasmic localisation in resting cells, cofilin was alsofound within the nuclei of NRK cells. Previous reportshave indicated that unphosphorylated cofilin in complexwith monomeric actin was localised within cell nucleiunder stress conditions (Ohta et al., 1989) or afterdephosphorylation (Moon and Drubin, 1995). The
orphology. NRK control cells (A); cells treated with 1 mM STS
–C) stained with FITC-annexin-V, (A0-C0) with Hoechst 33342
and apoptotic nuclear morphology in (B0) Addition of zVAD
ing (C), but the appearance of apoptotic nuclear morphology

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 799
reason for the preferential nuclear accumulation ofcofilin but not of ADF is presently unclear.
The morphological effects of STS
Treatment of both NRK and A431 cells with STSinduced marked morphological changes characterisedby an increased lamellipodial activity particularly of theA431 cells. The elevated lamellipodial activity did notresult in an increased directional migration instead itappeared to be uncoordinated in nature and led toextremely broad fan-like lamellipodia in A431 and astar-like appearance in NRK cells (Figs. 2–4). Theseeffects were observed over a wide range of STSconcentrations (1 nM–1 mM) and reversible even at1 mM STS provided the drug was removed within 2 h.However, longer exposure resulted in apoptosis. Apop-tosis was also induced by 1–50 nM STS albeit with aconsiderably longer delay. The STS-induced shapechanges were accompanied by disruption of the stressfibres and an accumulation of G-actin in the nucleus ofNRK cells as detected by fluorescent DNase I binding.Nuclear accumulation of actin has been reported inresponse to stress conditions and suggested that actin incomplex with unphosphorylated cofilin is preferentiallytranslocated to the nucleus (Ohta et al., 1989).
After STS exposure we observed a translocation ofcofilin and ADF to the cell cortex, where they co-localised with F-actin in lamellipodial regions as alsodemonstrated for activated neutrophils (Heyworth et al.,1997). A separation into a more proximal ADF/cofilinlocalisation was only observed for membrane ruffles orblebs that appeared after 2 h of drug exposure, asreported previously (Mannherz et al., 2005). A similarmore distal distribution of actin was reported forXenopus keratocytes with ADF/cofilin excluded fromthe leading edge of lamellipodia, but not in fibroblasts(Svitkina and Borisy, 1999). This spatial difference wasnot attributed to the rate of locomotion of the two celltypes, because in keratocytes, it did not change whenlocomotion was slowed by low concentrations of STS(Svitkina and Borisy, 1999). The exclusion of ADF/cofilin from the leading edge of blebs or ruffles mayreflect continuous addition of actin:ATP subunits(which do not bind ADF/cofilin) to the distal plus-endsof F-actin (Maciver and Weeds, 1994; Carlier et al.,1997) under conditions of constant treadmilling result-ing in a continuous protrusion (DesMarais et al., 2005).
We suspected that STS as a general inhibitor ofprotein kinases inhibited LIM kinases and therebyblocked phosphorylation (promote inhibition) of ADFand cofilin (Arber et al., 1998; Yang et al., 1998). Thecomplete dephosphorylation of cofilin observed afterSTS exposure (Fig. 4) provides indirect evidence forLIM kinase inhibition. Therefore we assume that both
cofilin and ADF were maximally active, although itshould be noted that phosphorylation is not the solemechanism for the regulation of activity of theseproteins, since they are also inhibited by PIP2 orbinding to other proteins (DesMarais et al., 2005).Elevated F-actin-severing activity by activated ADF/cofilin is associated with lamellipodial extension in ratadenocarcinoma cells (DesMarais et al., 2004; Chanet al., 2000). It is therefore likely that a considerablefraction of the actin subunits liberated during stress fibredisassembly was reused for the increased lamellipodialactivity, because the DNase I staining did not indicate adramatic relative increase in monomeric actin. A similaractivation has been reported for isolated lamellipodiafrom fish keratocytes after STS exposure (Verkhovskyet al., 1999). Thus the disorganised motility after STSexposure appeared to be at least partially due to thepersistent over-activation of ADF/cofilin.
It is noteworthy that after STS treatment myosin IIwas absent from the cytoplasmic extensions andrestricted to the perinuclear region, indicating that themotile activity of these lamellipodial structures does notrequire this form of myosin, although other myosinscannot be excluded (Krendel and Mooseker, 2005). STSwould be expected to inactivate MLCK, which woulduncouple the normal relationship between actin assem-bly and myosin II-based cell body retraction at the rearof lamellipodia that normally occurs during celllocomotion. Using an anti-P-MLC antibody we indeedobserved a decrease in specific cytoplasmic staining butnot its complete disappearance suggesting residualactive myosin II possibly due to the activity of death-associated protein (DAP) kinase which was shown to beactivated during apoptosis and to phosphorylate themyosin II light chain (Kuo et al., 2004) provided it is notinhibited by STS. Svitkina and Borisy (1999) alsoinferred incomplete myosin II inactivation by STS whenusing this drug to slow down fish keratocyte migration.Reduced MLC phosphorylation, stress fibre disassemblyand reduced attachment of the actin cytoskeleton tofocal adhesions (Kuo et al., 2004) may account for thedisorganised and uncoordinated motility and cytoplas-mic fragmentation observed after STS exposure. Sur-prisingly the anti-P-MLC antibody also gave a strongnuclear immunostaining that was not altered in intensityafter STS treatment (see Fig. 6G). The fact that anti-cytoplasmic myosin II did not give a nuclear stainpossibly suggests that this light chain might attach to adifferent myosin unless the nuclear staining was due tocross-reactivity with an unknown phospho-protein. Thepresence of myosins within the nucleus has repeatedlybeen demonstrated (Philimonenko et al., 2004), there-fore it will be interesting to identify this nuclear antigenin future experiments.
Active cytoplasmic myosin II is also necessary for theformation of large apoptotic blebs (Mills et al., 1998).

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802800
Indeed, a peculiarity of STS-induced apoptosis is theformation of only small apoptotic blebs (see Fig. 3E andF; Mills et al., 1998). In this respect, the morphologicalchanges induced by STS differ from other apoptosis-inducing drugs like etoposide, which was shown toinduce a typical apoptotic morphology characterised bynumerous large apoptotic blebs probably because it doesnot affect the equilibrium between de- and phosphory-lated proteins as demonstrated for cofilin (Mannherzet al., 2005).
Similarly, STS inhibited the phosphorylation of themicrotubule-stabilising Tau-protein. Dissociation ofTau-protein from MTs occurs only after its phosphor-ylation, which is a prerequisite of MT disassembly.Likewise lack of phosphorylation of IF proteinsstabilises IFs. Thus inhibition of the relevant proteinkinases probably accounts for the observed lack of aglobal disassembly of both MTs and IFs after STStreatment.
Longer exposure of cells to STS induces cell death
STS has been shown to induce apoptosis by themitochondrial pathway in which cytochrome c isliberated, which together with Apaf-1 (apoptosis pro-tease activating factor) and inactive pro-caspase-9 formsthe apoptosome, thereby initiating activation of caspase-9 and subsequently of the executioner caspases-3 and -6(Deshmukh and Johnson, 2000; Stepczynska et al.,2001) leading to classical apoptosis as also shown here.Simultaneous treatment of NRK cells with STS and thecaspase inhibitor zVAD prevented apoptosis as verifiedby the lack of apoptotic nuclear condensation andchromatin fragmentation (not shown). However, thistreatment did not inhibit the STS-typical shape changesincluding microfilament disassembly and increasedlamellipodial activity. This shows that the changes incell morphology and behaviour induced by STS areindependent of its apoptosis-inducing effect. Our dataindicate that the simultaneous exposure of NRK cells toSTS and zVAD induced a caspase-independent form ofcell death as verified by positive annexin-V-stainingalbeit at delayed rate. Similar results were previouslyreported by Deas et al. (1998) for peripheralT-lymphocytes. Since STS leads to disassembly of onlythe microfilament system, these results suggest that thewell-balanced equilibrium between monomeric andfilamentous actin is essential for cell survival. Ourresults on these two eukaryotic cell lines are inagreement with recent reports demonstrating that afinely balanced dynamic behaviour of actin is essentialfor survival of yeast cells (Gourlay et al., 2004).However, these authors observed a correlation of yeastcell aging and apoptosis with decreased actin dynamics.It will be interesting to analyse in future experiments in
greater detail the role of this equilibrium for survival ordemise of eukaryotic cells.
Apart from its apoptosis-inducing effect by cyto-chrome c release from mitochondria, our data highlightthe plethora of effects elicited by STS. They also indicatethat STS is not as suitable a purely apoptosis-inducingagent as would appear from its widespread usage.
Acknowledgements
It is a pleasure for us to thank Mrs. Eva Konieczny,Marion Otto and Swantje Wulf for expert technicalassistance and the Deutsche Forschungsgemeinschaftfor financial support. Sabine Gonsior thanks theEuropean Commission for a TMR Marie-Curie-Fellow-ship and H.G. Mannherz Trinity College, Cambridge,UK, for a Visiting Fellowship.
References
Arber, S., Barbayannis, F.A., Hanser, H., Schneider, C.,
Stanyon, C.A., Bernard, O., Caroni, P., 1998. Regulation of
actin dynamics through phosphorylation of cofilin by LIM-
kinase. Nature 393, 805–809.
Bamburg, J.R., 1999. Proteins of the ADF/cofilin family:
essential regulators of actin dynamics. Annu. Rev. Cell
Dev. Biol. 15, 185–230.
Bradford, M.M., 1976. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal. Biochem. 72,
248–254.
Carlier, M.-F., Laurent, V., Santolini, J., Melki, R., Didry, D.,
Xia, G.-A., Hong, Y., Chua, N.-H., Pantaloni, D., 1997.
Actin depolymerizing factor (ADF/cofilin) enhances the
rate of filament turnover: implication in actin-based
motility. J. Cell Biol. 136, 1307–1323.
Chan, A.Y., Bailly, M., Zebda, N., Segall, J.E., Condeelis, J.S.,
2000. Role of cofilin in epidermal growth factor-stimulated
actin polymerization and lamellipod protrusion. J. Cell
Biol. 148, 531–542.
Deas, O., Dumont, C., MacFarlane, M., Rouleau, M., Hebib,
C., Harper, F., Hirsch, F., Charpentier, B., Cohen, G.M.,
Senik, A., 1998. Caspase-independent cell death induced by
anti-CD2 or staurosporine in activated human peripheral T
lymphocytes. J. Immunol. 161, 3375–3383.
Deshmukh, M., Johnson Jr., E.M., 2000. Staurosporine-
induced neuronal death: multiple mechanisms and method-
ological implications. Cell Death Differ. 7, 250–261.
DesMarais, V., Macaluso, F., Condeelis, J., Bailly, M., 2004.
Synergistic interaction between the Arp2/3 complex and
cofilin drives stimulated lamellipod extension. J. Cell Sci.
117, 3499–3510.
DesMarais, V., Gosh, M., Eddy, R., Condeelis, J., 2005.
Cofilin takes the lead. J. Cell Sci. 118, 19–29.
Earnshaw, W.C., Martins, L.M., Kaufmann, S.H., 1999.
Mammalian caspases: structure, activation, substrates and

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802 801
functions during apoptosis. Annu. Rev. Biochem. 68,
383–424.
Enari, M., Sakahira, H., Yokoyama, H., Okawa, K.,
Iwamatsu, A., Nagata, S., 1998. A caspase-activated
DNase that degrades DNA during apoptosis, and its
inhibitor ICAD. Nature 39, 43–50.
Euteneuer, U., Schliwa, M., 1984. Persistent, directional
motility of cells and fragments in the absence of micro-
tubules. Nature 310, 58–61.
Gonsior, S.M., Platz, S., Buchmeier, S., Scheer, U., Jockusch,
B.M., Hinssen, H., 1999. Conformational difference
between nuclear and cytoplasmic actin as detected by a
monoclonal antibody. J. Cell Sci. 112, 797–809.
Gourlay, C.W., Carpp, L.N., Timpson, P., Winder, S.J.,
Ayscough, K.R., 2004. A role for the actin cytoskeleton in
cell death and aging in yeast. J. Cell Biol. 164, 803–809.
Hengstenberg, K., Mannherz, H.G., 1998. Actin cleavage by
caspases during apoptosis. Eur. J. Cell Biol. 75 (Suppl. 48), 39.
Heyworth, P.G., Robinson, J.M., Ding, J., Ellis, B.A.,
Badwey, J.A., 1997. Cofilin undergoes rapid dephosphor-
ylation in stimulated neutrophils and translocates to
ruffled membranes enriched in products of the NADPH
oxidase complex. Evidence for a novel cycle of phosphor-
ylation and dephosphorylation. Histochem. Cell Biol. 108,
221–233.
Hotulainen, P., Paunola, E., Vartiainen, M.K., Lappalainen,
P., 2005. Actin-depolymerizing factor and cofilin-1 play
overlapping roles in promoting rapid F-actin depolymer-
ization in mammalian non-muscle cells. Mol. Biol. Cell 16,
649–664.
Janicke, R.U., Ng, P., Sprengart, M.L., Porter, A.G., 1998.
Caspase-3 is required for a-fodrin cleavage but dispensable
for cleavage of other death substrates in apoptosis. J. Biol.
Chem. 273, 15540–15545.
Kerr, J.F.R., Wyllie, A.H., Currie, A.R., 1972. Apoptosis: a
basic biological phenomenon with wide-ranging implica-
tions in tissue kinetics. Br. J. Cancer 26, 239–257.
Kothakota, S., Azuma, T., Reinhard, C., Klippel, A., Tang, J.,
Chu, K., McGarry, T.J., Kirschner, M.-W., Koths, K.,
Kwiatkowski, D.J., Williams, L.T., 1997. Caspase-3-gener-
ated fragment of gelsolin: effector of morphological change
in apoptosis. Science 278, 294–298.
Krendel, M., Mooseker, M.S., 2005. Myosin: tails (and heads)
of functional diversity. Physiology 20, 239–251.
Kuo, J.-C., Lin, J.-R., Staddon, J.M., Hosoya, H., Chen, R.-
H., 2004. Uncoordinated regulation of stress fibres
and focal adhesions by DAP kinase. J. Cell Sci. 116,
4777–4790.
Laemmli, U.K., 1970. Cleavage of structural proteins during
the assembly of the head of bacteriophage T4. Nature 227,
680–685.
Lazebnik, Y.A., Takahashi, A., Moir, R., Goldman, R.,
Poirier, G.G., Kaufmann, S.H., Earnshaw, W.C., 1995.
Studies of the lamin proteinase reveal multiple parallel
biochemical pathways during apoptotic execution. Proc.
Natl. Acad. Sci. USA 92, 9042–9046.
Maciver, S.K., Weeds, A.G., 1994. Actophorin preferentially
binds monomeric ADP-actin over ATP-bound actin:
consequences for cell locomotion. FEBS Lett. 347,
251–256.
Maciver, S.K., Pope, B.J., Whytock, S., Weeds, A.G., 1998.
The effect of two actin depolymerizing factors (ADF/
cofilin) on actin filament turnover: pH sensitivity of F-actin
binding by human ADF, but not of Acanthamoeba
actophorin. Eur. J. Biochem. 256, 388–397.
Mannherz, H.G., Goody, R.S., Konrad, M., Nowak, E.,
1980. The interaction of bovine pancreatic deoxyribonu-
clease I and skeletal muscle actin. Eur. J. Biochem. 104,
367–379.
Mannherz, H.G., Gonsior, S.M., Gremm, D., Wu, X., Pope,
B.J., Weeds, A.G., 2005. Activated cofilin colocalizes with
Arp2/3 complex in apoptotic blebs during programmed cell
death. Eur. J. Cell Biol. 84, 503–515.
Mashima, T., Naito, M., Noguchi, K., Miller, D.K., Nichol-
son, D.W., Tsuruo, T., 1997. Actin cleavage by CPP-32/
apopain during the development of apoptosis. Oncogene
14, 1007–1012.
Matsudaira, P.T., Burgess, D.R., 1978. SDS microslab linear
gradient polyacrylamide gel electrophoresis. Anal. Bio-
chem. 87, 386–396.
McGough, A., Pope, B., Chiu, W., Weeds, A.G., 1997. Cofilin
changes the twist of F-actin: implications for actin
filament dynamics and cellular function. J. Cell Biol. 138,
771–781.
Mills, J.C., Stone, N.L., Erhardt, J., Pittman, R.N., 1998.
Apoptotic membrane blebbing is regulated by myosin light
chain phosphorylation. J. Cell Biol. 140, 627–636.
Mills, J.C., Sone, N.L., Pittman, R.N., 1999. Extranuclear
apoptosis: the role of the cytoplasm in the execution phase.
J. Cell Biol. 146, 703–706.
Moon, A., Drubin, D.G., 1995. The ADF/cofilin proteins:
stimulus-responsive modulators of actin dynamics. Mol.
Biol. Cell 6, 1423–1431.
Moon, A.L., Janmey, P.A., Louie, K.A., Drubin, D.G., 1993.
Cofilin is an essential component of the yeast cortical
cytoskeleton. J. Cell Biol. 120, 421–435.
Niwa, R., Nagata-Ohashi, K., Takeichi, M., Mizuno, K.,
Uemura, T., 2002. Control of actin reorganization by
Slingshot, a family of phosphatases that dephosphorylate
ADF/cofilin. Cell 108, 233–246.
Ohta, Y., Nishida, E., Sakai, H., Miyamoto, E., 1989.
Dephosphorylation of cofilin accompanies heat shock-
induced nuclear accumulation of cofilin. J. Biol. Chem.
264, 16143–16148.
Paddenberg, R., Loos, S., Schoneberger, H.-J., Wulf, S.,
Muller, A., Iwig, M., Mannherz, H.G., 2001. Serum
withdrawal induces a redistribution of intracellular gelsolin
towards F-actin in NIH 3T3 fibroblasts preceding apopto-
tic cell death. Eur. J. Cell Biol. 80, 366–378.
Peitsch, M.C., Mannherz, H.G., Tschopp, J., 1994. The
apoptotis endonucleases: cleaning up after cell death.
Trends Cell Biol. 4, 37–41.
Philimonenko, V.V., Zhao, J., Iben, S., Dingova, H., Kysela,
K., Kahle, M., Zentgraf, H., Hofmann, W.A., de Lanerolle,
P., Hozak, P., Grummt, I., 2004. Nuclaer actin and myosin
I are required for RNA polymerase I transcription. Nat.
Cell Biol. 6, 1165–1172.
Pollard, T.D., Borisy, G.G., 2003. Cellular motility driven by
assembly and disassembly of actin filaments. Cell 112,
453–465.

ARTICLE IN PRESSH.G. Mannherz et al. / European Journal of Cell Biology 85 (2006) 785–802802
Polzar, B., Rosch, A., Mannherz, H.G., 1989. A simple
procedure to produce monospecific polyclonal antibodies
of high affinity against actin. Eur. J. Cell Biol. 50,
220–229.
Pope, B., Gonsior, S.M., Yeoh, S., McGough, A., Weeds,
A.G., 2000. Uncoupling filament fragmentation by cofilin
from increased subunit turnover. J. Mol. Biol. 298,
649–661.
Stegh, A.H., Herrmann, H., Lampel, S., Weisenberger, D.,
Andra, K., Seper, M., Wiche, G., Krammer, P.H., Peter,
M.E., 2000. Identification of the cytolinker plectin as a
major early in vivo substrate for caspase 8 during CD95-
and tumor necrosis factor receptor-mediated apoptosis.
Mol. Cell. Biol. 20, 5665–5679.
Stepczynska, A., Lauber, K., Engels, I.H., Janssen, O.,
Kabelitz, D., Wesselborg, S., Schulze-Osthoff, K., 2001.
Staurosporine and conventional anticancer drugs induce
overlapping, yet distinct pathways of apoptosis and caspase
activation. Oncogene 20, 1193–1202.
Song, Q., Wie, T., Lees-Miller, S., Alnemri, E., Watters, D.,
Lavin, M.F., 1997. Resistance of actin to cleavage during
apoptosis. Proc. Natl. Acad. Sci. USA 94, 157–162.
Stolzenberg, I., Wulf, S., Mannherz, H.G., Paddenberg, R.,
2000. Different sublines of Jurkat cells respond with
varying susceptibility of internucleosomal DNA degrada-
tion to different mediators of apoptosis. Cell Tissue Res.
301, 273–282.
Svitkina, T.M., Borisy, G.G., 1999. Arp2/3 complex and actin
depolymerizing factor/cofilin in dendritic organization and
treadmilling of actin filament array in lamellipodia. J. Cell
Biol. 145, 1009–1026.
Verkhovsky, A.B., Svitkina, T.M., Borisy, G.G., 1999. Self-
polarization and directional motility of cytoplasm. Curr.
Biol. 9, 11–20.
Wu, X., Ritter, B., Schlattjan, J.H., Lessmann, V., Heumann,
R., Dietzel, I.D., 2000. Protein expression patterns of
identified neurons and of sprouting cells from leech central
nervous system. J. Neurobiol. 44, 320–332.
Wyllie, A.H., 1980. Glucocorticoid-induced thymocyte apop-
tosis is associated with endogenous endonuclease activa-
tion. Nature 284, 555–556.
Yang, N., Higuchi, O., Ohashi, K., Nagata, K., Wada, A.,
Kangawa, K., Nihishi, E., Mizuno, K., 1998. Cofilin
phosphorylation by LIM-kinase 1 and its role in Rac-
mediated actin organisation. Nature 393, 809–812.
Yeoh, S., Pope, B., Mannherz, H.G., Weeds, A., 2002.
Determining the differences in actin binding by human
ADF and cofilin. J. Mol. Biol. 315, 911–925.







![New pyrazolo[3,4-d]pyrimidines endowed with A431 antiproliferative activity and inhibitory properties of Src phosphorylation](https://img.pdfslide.net/doc/110x75/63334435ce61be0ae50e8fdc/new-pyrazolo34-dpyrimidines-endowed-with-a431-antiproliferative-activity-and.jpg)


