Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Duration of prepupal summer dormancy regulates synchronization of adultdiapause with winter temperatures in bees of the genus Osmia
Fabio Sgolastra a,⇑, William P. Kemp b, Stefano Maini a, Jordi Bosch c,d
a Dipartimento di Scienze e Tecnologie Agroambientali, Università di Bologna, Italyb Red River Valley Agricultural Research Center, USDA-ARS, Fargo, ND, USAc CREAF, Cerdanyola del Vallès 08193, Spaind Biology Department, Utah State University, Logan, UT, USA
a r t i c l e i n f o
Article history:Received 15 February 2012Received in revised form 17 April 2012Accepted 18 April 2012Available online 27 April 2012
Keywords:PhenologySummer diapauseFluctuating temperatureThermoperiodClimate changeGlobal warming
a b s t r a c t
Osmia (Osmia) bees are strictly univoltine and winter as diapausing adults. In these species, the timing ofadult eclosion with the onset of wintering conditions is critical, because adults exposed to long pre-win-tering periods show increased lipid loss and winter mortality. Populations from warm areas fly in Febru-ary–March and are exposed to longer growth seasons than populations from colder areas, which fly inApril–May. Given their inability to produce an extra generation, early-flying populations should developmore slowly than late-flying populations and thus avoid the negative consequences of long pre-winteringperiods. In this study we compare the development under natural and laboratory conditions of phenolog-ically-distinct populations in two Osmia species. Early-flying populations took �2 months longer todevelop than late-flying populations. Differences between populations in larval and pupal period dura-tion were very small, whereas the prepupal period was much longer in early-flying populations. In con-trast to the larval and pupal stages, the prepupal stage showed a non-linear response to temperature, wasstrongly affected by thermoperiod, and exhibited minimum respiration rates. Coupled with other lines ofevidence, these results suggest that the prepupal period in Osmia corresponds to a summer diapause, andits duration may be under local selection to synchronize adult eclosion with the onset of winter temper-atures. We discuss the implications of our results relative to current expectations of global warming.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Most insect species in temperate regions overwinter in a singlefixed developmental stage (Danks, 1987). For these species, timingof the wintering stage with the onset of wintering conditions maybe critical. Individuals developing too slowly and thus not reachingthe appropriate stage by winter are usually doomed (Taylor, 1986;Bradshaw et al., 1998), whereas individuals developing too quicklymay miss an opportunity for extended feeding and growth (Pullin,1987; Ohtsu et al., 1995). Species with wide distributions are ex-posed to climatic gradients, and thus exhibit substantial variationin seasonality among populations from different latitudes. Onemechanism by which insect populations time their life stages to lo-cal conditions is variation in voltinism. Populations from areaswith longer growing seasons typically have more generations peryear (Danks, 1987), and recent increases in number of generations
per year have been reported, seemingly in response to globalwarming (Kiritani, 2006; Gomi et al., 2007). Obligatorily univoltinespecies, on the other hand, may respond to global warming byreaching the wintering stage in advance of the onset of winter con-ditions, and then enter diapause to minimize energy expenditure(Hahn and Denlinger, 2007, 2011). Alternatively (if wintering in afeeding stage), univoltine populations may feed before enteringdiapause and thus increase their metabolic reserves in preparationfor winter. Thus, the optimal time to enter diapause can be under-stood as an adaptive compromise between building/conserving re-sources and avoiding untimely exposure to winter cold (Bradshawand Holzapfel, 2010).
Solitary bees in the subgenus Osmia (Osmia) are strictly univol-tine and winter as fully-emerged adults within their cocoons (Bos-ch et al., 2001). Because they have no access to food, these adultswinter on a fixed energetic budget. Diapause initiation takes placein late summer, shortly after adult eclosion. However, even at de-pressed metabolic rates, adults consume their fat bodies, experi-ence important lipid losses and rapidly loose weight during pre-wintering (defined as the period from adult eclosion to the onsetof winter temperatures) (Bosch and Kemp, 2004; Bosch et al.,
0022-1910/$ - see front matter � 2012 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jinsphys.2012.04.008
⇑ Corresponding author. Address: Dipartimento di Scienze e Tecnologie Agroam-bientali – Entomologia, Università di Bologna, viale G. Fanin 42, 40127 Bologna,Italy. Tel.: +39 051 2096296; fax: +39 051 2096281.
E-mail address: [email protected] (F. Sgolastra).
Journal of Insect Physiology 58 (2012) 924–933
Contents lists available at SciVerse ScienceDirect
Journal of Insect Physiology
journal homepage: www.elsevier .com/ locate/ j insphys

Author's personal copy
2010; Sgolastra et al., 2011). Individuals kept at pre-wintering(warm) temperatures for periods longer than two months exhibithigh winter mortality and low vigor at emergence and individualswintered as pupae have been found not to be viable (Bosch et al.,2006). Thus, timing of adult eclosion with the onset of winter tem-peratures is particularly critical in Osmia species. Here, we studythe development of phenologically-distinct populations of twounivoltine Osmia (Osmia) species, Osmia cornuta and Osmia lignaria.
O. (Osmia) cornuta (Latreille) is a Palearctic mason bee found incentral and southern Europe, Turkey and parts of North Africa andthe Middle East (Peters, 1977). O. (Osmia) lignaria Say is a Nearcticspecies distributed from southern Canada to northern Mexico,across most of continental USA (Rust, 1974). The two species arethus exposed to a wide range of climatic conditions that result inphenologically distinct local populations. At low latitudes, popula-tions are exposed to longer growing seasons because they emergeand begin flying earlier in the spring, and because wintering tem-peratures arrive later in the autumn. Differences in the timing ofthe flying period are obvious even between populations separatedless than 200 km. For example, O. cornuta populations from LaRibera (41�050N, 0�380E, Tarragona, NE Spain; mean annual tem-perature: 15.6 �C) typically fly in February through mid March,while populations from La Garrotxa (42�110N, 2�300E, Girona, NESpain; mean annual temperature: 12.4 �C) fly from mid Marchthrough April. Given the negative effects of pre-wintering, andtheir univoltine life cycle, populations of warm areas need to de-velop more slowly to synchronize adult eclosion with the onsetof winter temperatures.
In this study, we report on a series of experiments in whichdevelopmental rates of early- and late-flying populations are com-pared. We expect early-flying populations to develop more slowlythan late-flying populations, both under natural conditions andwhen exposed to the same temperature. We also analyze to whatextent different developmental stages respond differently to tem-perature. Then, because the prepupal period appears as the mostvariable developmental stage between populations, and shows aradically distinct response to temperature compared to otherstages, we measure respiration rates and describe associated mor-phological changes during this stage. Our ultimate objective is tounderstand how Osmia species synchronize the timing of adulteclosion to local temperature declines in autumn.
2. Materials and methods
2.1. Life history of O. cornuta and O. lignaria
O. cornuta and O. lignaria are very closely related species, andshare many life history and behavioral traits (Bosch et al., 2001).Both species emerge from the cocoon and fly in late winter andearly spring, and nest in pre-established cavities, in which theybuild series of cells delimited with mud partitions (Torchio,1989; Bosch, 1994). Females provision each cell with a loaf of pol-len mixed with nectar and lay one egg per cell. The nesting periodat a given location lasts approximately 1.5 months. Eggs hatch inabout one week, followed by five larval instars. Upon consumingthe last remnants of the provision, the fifth larval instar spins a co-coon and undergoes a dormant period (prepupal stage) thatapproximately coincides with the summer. Pupation occurs inmid or late summer, and adult eclosion (within the cocoon) in latesummer or early autumn. Upon adulthood, metabolic rates declineand cocooned adult bees enter an obligate diapause (Bosch et al.,2010; Sgolastra et al., 2010). Adults require exposure to cold tem-peratures (wintering) to emerge the following spring as tempera-tures increase (Bosch and Blas, 1994; Bosch and Kemp, 2003,2004; Sgolastra et al., 2010).
2.2. Experiment 1: effect of temperature and geographic origin ondevelopmental rates (O. cornuta)
We compared developmental rates of an early-flying populationfrom La Ribera (Tarragona, North Eastern Spain: 41�050N, 0�380E),and a late-flying population from El Barcial (Valladolid, North Cen-tral Spain: 41�950N, 5�270W) reared at two different temperatures(22 and 25 �C). In La Ribera, O. cornuta nests in early Februarythrough mid-March. In El Barcial, O. cornuta nests through Aprilinto mid-May. In 1993, before the beginning of the nesting periodin each area, trap-nests were attached to trees and buildings atseveral sites. Each trap-nest consisted of a wood block with 25drilled holes. Each hole was 15 cm long and accommodated an8 mm inside-diameter paper straw. Trap-nests at La Ribera werechecked daily and newly-plugged paper straws (completed nests)were pulled out of the wood block and taken to the laboratory.Nests were then dissected and provisions with eggs were individ-ually placed in clay wells (Torchio and Bosch, 1992). Eggs weresexed based on provision size and cell position within the nest (fe-males are produced deeper in the nest and are assigned larger pro-visions; Bosch, 1994). Eggs were dated assuming a cell productionrate of 0.5 cells/day (Bosch and Vicens, 2005, 2006). Nests from ElBarcial were treated in a similar way, but because trap-nests in thisarea could only be checked once, in the middle of the nesting per-iod, eggs from El Barcial were not dated and thus the duration ofthe larval period could not be measured.
Clay wells with provisions and eggs were placed in clear PVCboxes containing two additional clay wells filled with water to pro-vide adequate humidity, and then transferred to temperature cab-inets at 22 and 25 �C. Clay wells were checked every day and thedates of hatching, completion of cocoon spinning, pupation andadult eclosion were noted. Pupation and adult eclosion werechecked through a slit cut at the apical tip of the cocoon with a ra-zor blade. Sample sizes were �50 males and �20 females per pop-ulation and treatment. Previous studies have already establishedthat males develop faster than females (Bosch and Kemp, 2000).Thus, statistical analyses were conducted separately for malesand females. Similarly, because we were not interested in differ-ences between developmental stages, larval, prepupal and pupalduration were analyzed separately. Differences between tempera-tures in larval stage duration of the early-flying population wereanalyzed with a t-Student. Differences between populations andtemperatures in prepupal duration were analyzed with two-wayfactorial ANOVA. We followed the same procedure to analyze dif-ferences in pupal duration but, because duration of the prepupalstage might condition pupal duration, introduced prepupal dura-tion as a covariate. Within each developmental stage, we usedthe HSD Tukey test for post hoc means separation. All analyseswere conducted with Statistica 7.1 (StatSoft Inc.).
2.3. Experiment 2: effect of temperature regime on developmentalrates (O. cornuta)
In this experiment we further explored potential differences inresponse to temperature between prepupal and pupal stages. Wetested the effect of different temperature regimes on the durationof the prepupal and pupal stages in a late-flying (April) populationfrom Los Apriscos (Valladolid, North-Central Spain: 41�950N,5�270E). Nests were obtained in 1993 from trap-nests attached totrees and buildings. At the end of the nesting period, nests weredissected and male provisions with eggs or young larvae were indi-vidually placed in clay wells and treated as in Experiment 1. Thetemperature regimes tested were: 22, 25, 30 and 17:27 �C on a12:12 h thermoperiod (mean temperature: 22 �C). Sample sizeswere �50 males per treatment. We used one-way ANOVA to ana-lyze differences among temperature regimes in prepupal duration,
F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933 925

Author's personal copy
and one-way ANCOVA to analyze differences in pupal durationwith prepupal duration as a covariate. Within each stage, theHSD Tukey test was used for post hoc multiple comparisons.
2.4. Experiment 3: effect of temperature regime and population originon developmental rates (O. lignaria)
We compared developmental rates and degree-day require-ments of an early-flying (March) O. lignaria population from SanLuis Obispo (Central California: 35�170N, 120�410W), and a late-fly-ing (May) population from Cache Valley (Northern Utah: 40�430N,111�530W) reared at various laboratory temperature regimes andoutdoors. Paper straws containing nests of the early-flying popula-tion were sent to the laboratory on 21 March and 7 April 2005, andprovisions with eggs and first instar larvae (�1-day-old) were indi-vidually placed in clay wells as in Experiments 1 and 2. After co-coon completion, cocoons were placed individually in clear gelcapsules, and X-rayed every 3 days (Stephen and Undurraga,1976). X-ray plates were used to record the dates when bees pu-pated and eclosed as adults within the cocoon. We tested three dif-ferent constant (20, 26 and 29 �C), and one fluctuating (14:27 �C[8:16 h]; mean = 22 �C) temperature regimes. Sample sizes were�35 males and �30 females per treatment. As individual bees be-came adults, they were pre-wintered at treatment temperature(14 days) and subsequently cooled for wintering (7 days at20 �C + 180 days at 4 �C). After wintering, in spring 2006, bees wereincubated at 22 �C and checked daily for emergence from the co-coon. One additional treatment (outdoors) was conducted to mon-itor developmental rates under natural conditions. Somecompleted nests were left within the wood blocks in an open-sidedwooden box held 1.5 m above the ground in an orchard in San LuisObispo. Starting on 6 July 2005, nests were X-rayed every twoweeks to monitor pupation and adult eclosion. Starting in February2006, nests were checked daily to monitor emergence. Develop-mental rates of the late-flying population were obtained from aprevious study (Bosch and Kemp, 2000), in which groups of �60males and �40 females were exposed to 18, 22, 26, 29, 14:27 �C[8:16 h] and outdoors. Throughout the nesting period (May1997), newly plugged nests were taken to the laboratory, and cellswith non-hatched eggs were placed in clay wells as in Experiment1. Pupation and adult eclosion dates were monitored through X-rays every 3 days (Bosch and Kemp, 2000).
As in previous experiments, statistical analyses were conductedseparately for males and females and for each developmentalstage. Data were square root-transformed to improve homoscedas-ticity. Two-way ANOVA was used to analyze differences betweenpopulation and temperatures (only treatments used in both popu-lations: 26, 29 and 14:27 �C) in developmental duration of the lar-val stage. Duration of the prepupal stage was analyzed with two-way ANCOVA with larval duration as a covariate. We used thesame procedure to analyze duration of the pupal stage, with larvaland prepupal duration as covariates. Within each stage, the HSDTukey test was used for post hoc multiple comparisons.
For each individual bee we calculated the degree-days (DD) nec-essary to complete each developmental stage. We used the equa-tion DD = RT�(T � Tmin) (Pruess, 1983; Higley et al., 1986), whereRT is the duration of the developmental stage, T is the developmen-tal temperature, and Tmin is the lower temperature threshold, atwhich developmental rate is zero. We estimated Tmin from theregression line of development rate versus temperature(v = a + bt), where v is rate of development (1/RT), and a and bare constants. Two-way ANOVA was used to compare DD associ-ated to the larval and pupal stages between populations and sexes.It was not possible to calculate DD of the prepupal stage becausedevelopmental rate of this stage did not show a linear responseto temperature.
2.5. Experiment 4: respiration rates and morphological changes duringthe prepupal stage (O. lignaria)
Bees used in this experiment were obtained from a populationfrom Cache Valley (Utah) released in May 2003 in an apple orchardin North Logan (Cache Valley, Utah). During the nesting period re-cently-plugged nests were brought to the laboratory and dissectedto collect male cells with eggs or first instar larvae (�1-day-old).These cells were placed in clay wells as in previous experimentsand into boxes with screen lids kept in a barn adjacent to the appleorchard. Temperature inside the boxes was recorded hourly with atemperature logger. Clay wells were checked once a week from eggto first silk, and then daily until cocoon completion. Then, cocoonswere placed in gel capsules and X-rayed every three days to mon-itor pupation and adulthood. To measure respiration rates, we se-lected seven individuals in each of the following stages: fifth instar(cocoon spinning) larva (LA); newly-cocooned prepupa (PP);PP + 3 days (PP + 3); PP + 6 days (PP + 6); PP + 9 days (PP + 9);PP + 12 days (PP + 12); PP + 15 days (PP + 15); PP + 18 days(PP + 18); newly-formed pupa (PU); PU + 10 days (PU + 10);PU + 20 days (PU + 20). Specimens of the prepupal stages were ob-served under the microscope and photographed to describe theirappearance.
We used constant volume respirometry to measure O2 con-sumption and CO2 production as indicators of overall metabolicactivity. As in previous studies (Kemp et al., 2004; Bosch et al.,2010; Sgolastra et al., 2010, 2011), we used a Sable Systems FC-1O2 Analyzer and a Li-Cor CO2 Analyzer operating in differentialmode with a 100 ml/min flow rate. This allowed accuracy of mea-surement that exceeded 0.001% in detecting departures from anundepleted air stream that had been scrubbed of CO2 and water va-pour with a Drierite�–Ascarite� column. Measurements lasted 2 hand were conducted in darkness in a 22 �C – cabinet. The first60 min served as a baseline equilibration interval during whichbees were acclimatized and respiration chambers where filled withair scrubbed of CO2 and locked down. Respiration was then al-lowed (O2 consumed and CO2 produced) for 15 min. The last45 min were devoted to measuring the contents (O2/CO2) sequen-tially in each chamber, including the blank. Data were collected viathe Sable Systems data acquisition program DATACAN followingmanufacturer’s protocols. Seven different males were used in eachmeasurement. Upon completing a measurement, we dissected thecocoons and weighed the bees. O2 and CO2 levels were adjusted forthe weight of each individual and expressed as ml/g�h.
3. Results
3.1. Experiment 1: effect of temperature and geographic origin ondevelopmental rates (O. cornuta)
The larval period was only monitored in the early-flying popu-lation and lasted �40 days. This period was slightly shorter inmales reared at the warmer temperature (t(1, 93) = 2.22; P = 0.03),but females showed no differences (t(1, 43) = 0.39; P = 0.7). Temper-ature and population had a significant effect on duration of theprepupal stages in both sexes and the temperature x populationinteraction was significant in males but failed significance in fe-males (Table 1). Overall, the prepupal period was shorter at thewarmer temperature and in the late-flying population, and differ-ences between temperatures were more pronounced in the early-flying population (Fig. 1). As for the duration of the pupal period,we found differences between temperatures (shorter at the war-mer temperature) in both sexes, but no differences between popu-lations (Table 1). The duration of the pupal period was not affectedby prepupal duration in either sex (Table 1). At 22 �C, some individ-
926 F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933

Author's personal copy
uals from both populations remained in the prepupal stagethrough the summer and into the autumn (Table 2). These individ-uals were alive and had a healthy appearance, but showed no signsof pupation. Individuals expressing this prolonged prepupal dor-mancy were not included in development rate calculations. Noprolonged prepupal dormancy occurred at 25 �C (Table 2).
3.2. Experiment 2: effect of temperature regime on prepupal and pupalduration (O. cornuta)
Prepupal duration was significantly affected by temperature(F(3, 211) = 14.50; P < 0.001), and was longest at 22 �C with no differ-ences among the other treatments (Fig. 2). Interestingly, bees ex-posed to 17:27 �C (mean = 22 �C) developed as fast as beesexposed to a 25 �C constant temperture. By contrast, the durationof the pupal period decreased with increasing temperature (F(3,
210) = 51.57, P < 0.001), and was virtually the same at 17:27 �C asat 22 �C (Fig. 2). The duration of pupal period was again not af-fected by prepupal duration (F(1, 210) = 0.84; P = 0.4). At 22 �C, some
individuals exhibited prolonged prepupal dormancy, but not at theother temperature regimes, including 17:27 �C (Table 2).
3.3. Experiment 3: effect of temperature regime and population originon developmental rates (O. lignaria)
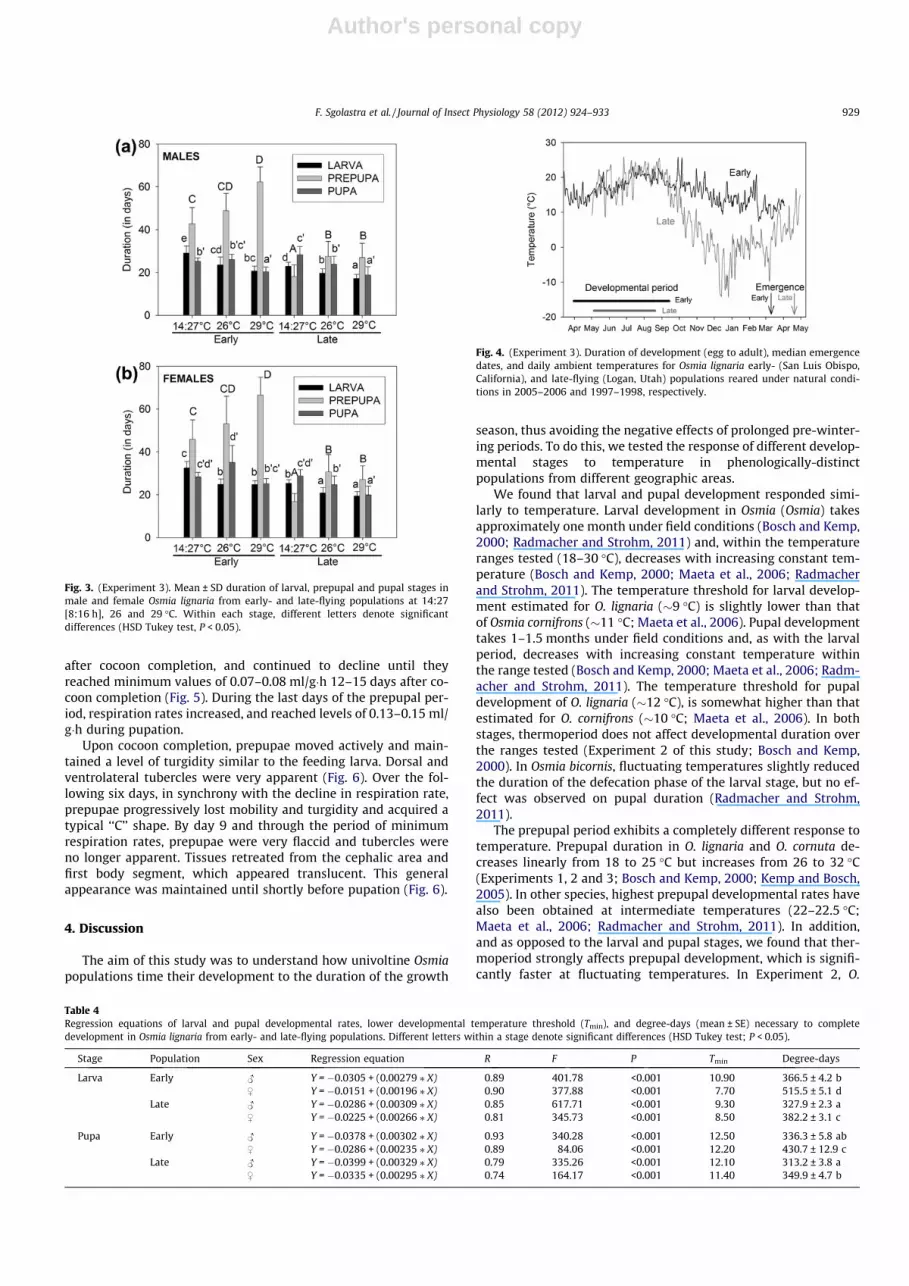
Larval duration decreased with increasing temperature, andwas slightly shorter in the late- than in the early-flying population(Table 3, Fig. 3). The interaction temperature x population was sig-nificant only in males. The duration of the prepupal period was alsoaffected by temperature and population, with a highly significantinteraction between these two factors (Table 3). Prepupal durationwas much shorter in the late-flying population and it increasedwith increasing temperature, especially in the early-flying popula-tion (Fig. 3). Duration of the larval period did not affect duration ofthe prepupal period in either sex. Duration of the pupal period wasinfluenced by temperature (shorter at the warmer temperatures),but an effect of population (shorter in the late-flying population)was only observed in females (Table 3, Fig. 3). The interaction tem-perature x population was highly significant in both sexes becausedifferences between temperatures were more pronounced in thelate-flying population. Pupal duration was not affected by larvalduration, and was only weakly affected by prepupal duration inmales (Table 3).
The degree-days necessary to complete larval developmentwere significantly greater in the early- than in the late-flying pop-ulation (F(1, 616) = 585.12; P < 0.001), and in females than in males(F(1, 616) = 818.52; P < 0.001) (Table 4). We also found a significantpopulation by sex interaction (F(1, 616) = 177.50; P < 0.001), as dif-ferences between sexes were higher in the early- than in thelate-flying population. Similarly, the degree-days needed to com-plete the pupal stage were significantly greater in the early-flyingpopulation (F(1, 424) = 51.46; P < 0.001) and in females (F(1,
424) = 81.77; P < 0.001), and again, there was a population by sex ef-fect (F(1, 424) = 15.86; P < 0.001), with differences between sexesbeing more pronounced in the early-flying population (Table 4).The response of larval and pupal stage to temperature was linearin both sexes (Table 4). Minimum threshold temperatures for lar-val development ranged from 8 to 11 �C, and were �12 �C for pupaldevelopment. We did not calculate degree-days or Tmin for theprepupal stage because its duration did not show a linear responseto temperature. However, the fact that some individuals at thelowest temperatures (20 �C for the early-flying population, 18 �Cfor the late-flying population; see below) expressed prolongedprepupal dormancy provides some indication of threshold temper-atures for development of this stage.
By 1 October, 28 individuals from the early-flying populationreared at 20 �C had not yet pupated (Table 2). These bees werewintered as prepupae at 4 �C for 180 days. In the following spring,they were transferred to 14:27 �C [8:16 h] (the treatment yieldingfastest prepupal development). Some of these bees died in the pu-pal stage, but 23 (82%) became adults by 13 July and were wintered
Table 1(Experiment 1). Results of two-way factorial ANOVA of prepupal and pupal duration in relation to population (Early vs Late) and temperature (22� vs 25 �C) in Osmia cornuta.Prepupal duration was used as a covariate in pupal stage analyses.
Stage Factors Males Females
F df P F df P
Prepupa Temperature 72.95 1, 176 <0.001 25.74 1, 90 <0.001Population 40.13 1, 176 <0.001 6.62 1, 90 0.01Temperature x Population 4.40 1, 176 0.04 3.57 1, 90 0.06
Pupa Temperature 45.91 1, 175 <0.001 6.29 1, 89 0.01Population 3.73 1, 175 0.06 1.34 1, 89 0.3Temperature x Population 1.45 1, 175 0.2 0.06 1, 89 0.8Prepupal duration 1.97 1, 175 0.2 0.85 1, 89 0.4
Fig. 1. (Experiment 1). Mean ± SD duration of prepupal and pupal stages in malesand females of Osmia cornuta from early- and late-flying populations at 22 and25 �C. Within each stage, different letters denote significant differences (HSD Tukeytest, P < 0.05).
F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933 927

Author's personal copy
at 4 �C for 180 days. In spring 2007, these individuals were incu-bated at 22 �C and checked daily for emergence. Five of them diedwithin the cocoon, but 18 (64% of the original 28) were able toemerge, and lived for 4–5 days. In the late-flying population 17bees reared at 18 �C were still in the prepupal stage by 23 October,and their fate is described in Bosch and Kemp (2000). Nine of thesebees were wintered as prepupae and incubated the followingspring. Most of them never pupated, but 3 reached adulthood. Of
these, 2 survived their second winter, and 1 (11% of the 9 individ-uals wintered as prepupae) emerged following incubation andlived for 3 days.
By 3 June, all bees from the early-flying population reared out-doors were in prepupal stage. By 19 August (median date: 5 Au-gust), all individuals had pupated, and by 4 October all of themwere adults (median date: 21 September). Therefore, bees of theearly-flying population took �170 days to complete development(Fig. 4). In the following spring, emergence took place in Febru-ary–March. All bees of the late-flying population reached theprepupal stage by 20 June, that is, slightly later than bees of theearly-flying population. Nonetheless, bees of the late-flying popu-lation pupated earlier (by 31 July; median date: 13 July), and by30 August all of them were adults (median date: 15 August). Totaldevelopment time in this population was �110 days, and springemergence occurred in April (Fig. 4). Differences in egg-adultdevelopmental duration between populations (longer in theearly-flying population) were confirmed at all artificial tempera-ture regimes tested (Appendix A).
3.4. Experiment 4: respiration rates and morphological changes duringthe prepupal stage (O. lignaria)
Respiration rates of O. lignaria males from a late-flying popula-tion reared outdoors was 0.7–0.9 ml/g�h in the cocoon-spinninglarva. Then, respiration rates dropped to �0.15 ml/g�h immediately
Table 2(Experiments 1, 2, 3). Percent individuals remaining in the prepupal stage by autumn (prolonged prepupal dormancy) according to population, temperature and sex.
Experiment Species Population Temperature (�C) % Prolonged prepupal dormancy
# $
1 O. cornuta Early 22 15.3 14.3Early 25 0 0Late 22 11.7 32.5Late 25 0 0
2 O. cornuta Late 22 15.0 –Late 25 0 –Late 30 0 –Late 17:27 0 –
3 O. lignaria Early 20 46.7 63.6Early 26 0 0Early 29 0 0Early 14:27 0 0Late 18 15.7 37.5Late 22 0 0Late 26 0 0Late 29 0 0Late 14:27 0 0
Fig. 2. (Experiment 2). Mean ± SD duration of prepupal and pupal stages in Osmiacornuta males reared at three constant temperatures (22, 25 and 30 �C) and onefluctuating temperature regime (17:27 [12:12 h]; mean = 22 �C). Within each stage,different letters denote significant differences (HSD Tukey test, P < 0.05).
Table 3(Experiment 3). Results of two-way ANOVA of larval, prepupal and pupal duration in relation to population (Early vs Late) and temperature (14:27 [mean = 22 �C], 26 and 29 �C) inOsmia lignaria. Larval duration was used as a covariate in the prepupal stage analyses, and larval and prepupal duration as covariates in the pupal stage analyses.
Stage Factors Males Females
F df P F df P
Larva Temperature 188.44 2, 234 <0.001 123.95 2, 163 <0.001Population 128.87 1, 234 <0.001 131.96 1, 163 <0.001Temp. x Pop. 3.38 2, 234 0.04 2.08 2, 163 0.1
Prepupa Temperature 30.10 2, 233 <0.001 24.25 2, 162 <0.001Population 307.73 1, 233 <0.001 208.38 1, 162 <0.001Temp. x Pop. 6.78 2, 233 0.001 6.03 2, 162 0.003Larval duration 0.09 1, 233 0.8 0.02 1, 162 0.9
Pupa Temperature 32.95 2, 232 <0.001 19.65 2, 161 <0.001Population 2.41 1, 232 0.1 13.65 1, 161 <0.001Temp. x Pop. 9.44 2, 232 <0.001 15.81 2, 161 <0.001Larval duration 0.71 1, 232 0.4 1.135 1, 161 0.3Prepupal duration 4.40 1, 232 0.04 2.02 1, 161 0.2
928 F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933
Author's personal copy
after cocoon completion, and continued to decline until theyreached minimum values of 0.07–0.08 ml/g�h 12–15 days after co-coon completion (Fig. 5). During the last days of the prepupal per-iod, respiration rates increased, and reached levels of 0.13–0.15 ml/g�h during pupation.
Upon cocoon completion, prepupae moved actively and main-tained a level of turgidity similar to the feeding larva. Dorsal andventrolateral tubercles were very apparent (Fig. 6). Over the fol-lowing six days, in synchrony with the decline in respiration rate,prepupae progressively lost mobility and turgidity and acquired atypical ‘‘C’’ shape. By day 9 and through the period of minimumrespiration rates, prepupae were very flaccid and tubercles wereno longer apparent. Tissues retreated from the cephalic area andfirst body segment, which appeared translucent. This generalappearance was maintained until shortly before pupation (Fig. 6).
4. Discussion
The aim of this study was to understand how univoltine Osmiapopulations time their development to the duration of the growth
season, thus avoiding the negative effects of prolonged pre-winter-ing periods. To do this, we tested the response of different develop-mental stages to temperature in phenologically-distinctpopulations from different geographic areas.
We found that larval and pupal development responded simi-larly to temperature. Larval development in Osmia (Osmia) takesapproximately one month under field conditions (Bosch and Kemp,2000; Radmacher and Strohm, 2011) and, within the temperatureranges tested (18–30 �C), decreases with increasing constant tem-perature (Bosch and Kemp, 2000; Maeta et al., 2006; Radmacherand Strohm, 2011). The temperature threshold for larval develop-ment estimated for O. lignaria (�9 �C) is slightly lower than thatof Osmia cornifrons (�11 �C; Maeta et al., 2006). Pupal developmenttakes 1–1.5 months under field conditions and, as with the larvalperiod, decreases with increasing constant temperature withinthe range tested (Bosch and Kemp, 2000; Maeta et al., 2006; Radm-acher and Strohm, 2011). The temperature threshold for pupaldevelopment of O. lignaria (�12 �C), is somewhat higher than thatestimated for O. cornifrons (�10 �C; Maeta et al., 2006). In bothstages, thermoperiod does not affect developmental duration overthe ranges tested (Experiment 2 of this study; Bosch and Kemp,2000). In Osmia bicornis, fluctuating temperatures slightly reducedthe duration of the defecation phase of the larval stage, but no ef-fect was observed on pupal duration (Radmacher and Strohm,2011).
The prepupal period exhibits a completely different response totemperature. Prepupal duration in O. lignaria and O. cornuta de-creases linearly from 18 to 25 �C but increases from 26 to 32 �C(Experiments 1, 2 and 3; Bosch and Kemp, 2000; Kemp and Bosch,2005). In other species, highest prepupal developmental rates havealso been obtained at intermediate temperatures (22–22.5 �C;Maeta et al., 2006; Radmacher and Strohm, 2011). In addition,and as opposed to the larval and pupal stages, we found that ther-moperiod strongly affects prepupal development, which is signifi-cantly faster at fluctuating temperatures. In Experiment 2, O.
Fig. 3. (Experiment 3). Mean ± SD duration of larval, prepupal and pupal stages inmale and female Osmia lignaria from early- and late-flying populations at 14:27[8:16 h], 26 and 29 �C. Within each stage, different letters denote significantdifferences (HSD Tukey test, P < 0.05).
Table 4Regression equations of larval and pupal developmental rates, lower developmental temperature threshold (Tmin), and degree-days (mean ± SE) necessary to completedevelopment in Osmia lignaria from early- and late-flying populations. Different letters within a stage denote significant differences (HSD Tukey test; P < 0.05).
Stage Population Sex Regression equation R F P Tmin Degree-days
Larva Early # Y = �0.0305 + (0.00279 ⁄ X) 0.89 401.78 <0.001 10.90 366.5 ± 4.2 b$ Y = �0.0151 + (0.00196 ⁄ X) 0.90 377.88 <0.001 7.70 515.5 ± 5.1 d
Late # Y = �0.0286 + (0.00309 ⁄ X) 0.85 617.71 <0.001 9.30 327.9 ± 2.3 a$ Y = �0.0225 + (0.00266 ⁄ X) 0.81 345.73 <0.001 8.50 382.2 ± 3.1 c
Pupa Early # Y = �0.0378 + (0.00302 ⁄ X) 0.93 340.28 <0.001 12.50 336.3 ± 5.8 ab$ Y = �0.0286 + (0.00235 ⁄ X) 0.89 84.06 <0.001 12.20 430.7 ± 12.9 c
Late # Y = �0.0399 + (0.00329 ⁄ X) 0.79 335.26 <0.001 12.10 313.2 ± 3.8 a$ Y = �0.0335 + (0.00295 ⁄ X) 0.74 164.17 <0.001 11.40 349.9 ± 4.7 b
Fig. 4. (Experiment 3). Duration of development (egg to adult), median emergencedates, and daily ambient temperatures for Osmia lignaria early- (San Luis Obispo,California), and late-flying (Logan, Utah) populations reared under natural condi-tions in 2005–2006 and 1997–1998, respectively.
F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933 929

Author's personal copy
cornuta prepupal duration dropped from 73 days at constant 22 �Cto 57 days at 17:27 �C (mean = 22 �C). In O. lignaria, it went from29 days at 22 �C to 17 days at 14:27 �C (mean = 22 �C) (Bosch andKemp, 2000). In Osmia bicornis, Radmacher and Strohm (2011)found a similar response when they compared three constant tem-peratures (17.5, 22.5, 27.5 �C) with their equivalent fluctuatingregimes.
Effects of thermoperiod on developmental rates have been ob-served in some insects, but not in others, and in a few cases, fluc-tuating temperatures have resulted in lower developmental rates(Beck, 1983; Ratte, 1984; Bryant et al., 1999). In species with facul-tative diapause, thermoperiod, alone or in concert with photope-riod, may enhance or diminish the incidence of diapausedepending on the upper and lower temperatures, the amplitudeof the cycle, and the duration of the cryophase and the thermo-phase (Beck, 1983; Tauber et al., 1986; Saulich and Musolin,
1996; Yamashiro et al., 1998; Kalushkov et al., 2001; Fontinouet al., 2002; Mironidis and Savopoulou-Soultani, 2012). In general,thermoperiods that result in a high incidence of diapause also yieldlow rates of diapause development (Danks, 1987). The biochemicalmechanisms underlying the effects of fluctuating temperature ondevelopment duration are not completely understood, but byencompassing a wider range of temperatures, fluctuating regimesare expected to provide favorable conditions for a wider varietyof enzyme systems with different thermokinetic characteristics(Morden and Waldbauer, 1980). Particularly relevant is the factthat some O. cornuta individuals could not complete prepupaldevelopment at 22 �C constant, but all individuals were able todo so when exposed to the same mean temperature under a fluc-tuating regime (Table 2).
Several lines of physiological and phylogenetic evidence sug-gest that prepupal dormancy in Osmia is diapause-mediated.First, respiration rates reach minimum values of �0.07 ml/g hduring this stage. These values are similar to rates obtained inpre-wintering and wintering prepupae of another Megachilidbee, the alfalfa leafcutting bee, Megachile rotundata, whichspends the winter in prepupal diapause (Kemp et al., 2004).These values are also similar to rates of adult diapausing Osmia(Kemp et al., 2004; Bosch et al., 2010; Sgolastra et al., 2011).Second, weight loss during the prepupal period in Osmia is min-imal (�0.08 mg/day) compared with 0.3–0.7 during the pupalperiod at the same temperatures (�22 �C) (Bosch and Vicens,2002; Kemp et al., 2004). Under natural conditions, the prepupalperiod normally coincides with the warmest summer tempera-tures (Kemp and Bosch, 2005), and therefore likely representsa very important reduction in metabolic expenditure during thisstage. Third, individuals reared at 18 and 20 �C that did not com-plete prepupal development by the end of autumn remained inthis stage for several months without dying or changing theirappearance. In O. cornifrons, which is also univoltine, prepupaewere kept at 10 �C for as long as 14 months and then resumedtheir development when exposed to 22 �C (Maeta et al., 2006).Fourth, parsivoltinism (the coexistence of univoltine and semi-voltine individuals within a population) has been reported in
Fig. 5. (Experiment 4). Mean ± SE O2 consumption and CO2 production measured at22 �C throughout the late larval, prepupal and pupal stages in Osmia lignaria malesfrom a late-flying population reared under natural conditions (n = 7 per treatment).LA: cocoon spinning larva; PP: prepupa upon cocoon completion; PP + 3 to PP + 18:3- to 18-day-old prepupa; PU: newly formed pupa; PU + 10 to PU + 20: 10- to 20-day-old pupa.
Fig. 6. (Experiment 4). Appearance of Osmia lignaria males from a late-flying population throughout the prepupal period. PP: prepupa upon cocoon completion; PP + 3:prepupa + 3 days; PP + 6: prepupa + 6 days; PP + 9: prepupa + 9 days; PP + 12: prepupa + 12 days; PP + 15: prepupa + 15 days; PP + 18: prepupa + 18 days.
930 F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933

Author's personal copy
many Osmia species in other subgenera (Torchio and Tepedino,1982; Bosch et al., 2001). Semivoltine individuals of these spe-cies spend their first winter as prepupae and their second winteras adults. Wintering in the adult stage is a derived trait withinthe Megachilidae, most of which winter as prepupae (Boschet al., 2001). In these species, the prepupa enters diapause insummer and resumes development in late spring or early sum-mer (Kemp et al., 2004). Wintering in the adult stage allows Os-mia species to emerge early in the spring, at a time when fewother bees are active (Bosch et al., 2001). Interestingly, someof the O. lignaria individuals expressing prolonged prepupal dor-mancy that were overwintered in this stage were able to com-plete development in the second year and eventually emergefrom the cocoon. Their longevity in the laboratory (3–5 days)was similar to that of univoltine individuals (Bosch and Kemp,2000). Taken together, these results suggest that prepupal sum-mer dormancy in Osmia may be an abbreviated version of thelong (summer–winter) diapause found in other Megachilidae,including the ancestral Lithurgini, and that some Osmia (Osmia)individuals have retained the capacity to overwinter as prepu-pae. Ongoing studies are analyzing expression of diapause-re-lated genes (Denlinger, 2002; Kostal, 2006; Yocum et al., 2005,2006) during the various stages of the life cycle in O. lignaria.Life cycles with two distinct diapause episodes occurring in dif-ferent developmental stages have been documented in a few in-sect species, including moths, sawflies, chrysomelid beetles anddamselflies (Masaki, 1980). All these species reproduce in theautumn and winter as eggs. Some mosquito species in the gen-era Wyeomyia and Aedes go through a summer diapause in theegg stage and then spend the winter as diapausing larvae (Tau-ber et al., 1986). The pentatomid bug Carbula humerigera goesthrough two facultative diapauses, one in the adult stage inthe summer and the other in the nymphal stage in the winter.The adult diapause has have been interpreted as a mechanismto maintain a univoltine life cycle under conditions otherwisefavorable for the development of multiple generations (Saulichand Musolin, 1996).
O. cornuta and O. lignaria populations from different geographicareas show important phenological differences. Under natural con-ditions, development from egg to adult takes about 5.5 months inearly-flying populations, compared to 3.5 months in late-flyingpopulations. These differences in development could be due to dif-ferences between geographic areas in ambient temperatures and/or to differences between populations in response to temperature(Nechols et al., 1987; Tauber et al., 1987; Ayres and Scriber,1994; Papanatasiou et al., 2011). The fact that differences in ambi-ent temperatures between Logan, Utah, and San Luis Obispo, Cali-fornia, are small during the developmental period of O. lignaria(Fig. 4), suggests that the two populations studied respond differ-ently to temperature. This was confirmed in our laboratory exper-iments, showing longer egg to adult development time in the earlypopulation at all temperatures tested (Appendix A). The early-fly-ing population required more degree-days to complete both stages,but differences between populations in larval and pupal develop-mental time were small in O. lignaria (and non-significant in O.cornuta). In contrast, populations showed large differences inprepupal stage duration. Under natural conditions, the early-flyingO. lignaria population spent two and half months as prepupae,compared to approximately one month in the late-flying popula-tion, and longer prepupal duration in the early-flying populationswas evident at all artificial temperatures tested. We conclude thatthe duration of the prepupal summer dormancy is the main mech-anism through which Osmia populations synchronize adult eclo-sion with the onset of wintering temperatures, thus avoiding thenegative effects of prolonged pre-wintering periods (Bosch et al.,2010; Sgolastra et al., 2011). Available evidence suggests that
prepupal dormancy duration is at least in part genetically con-trolled. When bees from a late-flying O. lignaria population fromnorthern Utah were released in a February-blooming almond orch-ard in California and reared from egg to adult under central Califor-nia natural conditions, they expressed the short prepupal diapausetypical of their area of origin, became adults early in the summer,and were thus exposed to an extended pre-wintering period thatresulted in high winter mortality (Bosch et al., 2000). Interestingly,except in males of Experiment 3, in which we found a weak nega-tive correlation between prepupal and pupal duration, bees did notadjust the duration of the pupal period to the duration of theprepupal period, further reinforcing our conclusion that prepupalduration is the main variable regulating synchronization with win-ter temperatures.
Summer temperatures are expected to increase under thecurrent scenario of global warming (Bale and Hayward, 2010).Other studies have emphasized phenological mismatches be-tween bee activity and spring flower resources (Hegland et al.,2009; Forrest and Thomson, 2011; Bartomeus et al., 2011). Ourresults suggest another potentially important mismatch betweenadult eclosion and the arrival of winter temperatures. As shownin this and other studies (Kemp and Bosch, 2005; Maeta et al.,2006; Radmacher and Strohm, 2011) prepupal duration de-creases with increasing temperature up to a threshold of 22–26 �C, beyond which it increases. If temperature increases ex-pected under global warming were moderate, we would expectfaster prepupal development, leading to earlier adult eclosion,which, together with an extended warm season, would exacer-bate the negative consequences of prolonged pre-wintering peri-ods (Bosch et al., 2010; Sgolastra et al., 2011). On the otherhand, if temperature increases were more severe, Osmia popula-tions could exhibit protracted prepupal periods thereby re-syn-chronizing adult eclosion with the delayed onset of wintertemperatures. It is important to note, however, that this physio-logical response to increased temperatures is unlikely to comewithout a metabolic cost. In the green stink bug, Nezara viridula,individuals exposed to a 2.5 �C temperature increase over naturalconditions in late-summer expressed slower developmentalrates, but suffered higher mortality and showed lower post-win-ter performance compared to individuals exposed to natural con-ditions (Musolin et al., 2010). For this reason, we expect thatincreased summer temperatures will more likely result is the lo-cal selection of Osmia phenotypes with extended prepupal dia-pause and/or the northward migration of southerly populations(Kemp and Bosch, 2005; Sgolastra et al., 2011).
Acknowledgements
Thanks are due to J. Calzadilla, M.A. Escolano, M. López andN. Vicens (Barcelona), G. Trostle, S. Kalaskar and P. Rieger (Lo-gan) and S. Pryor (San Luis Obispo) for their help with obtainingpopulations and monitoring development. The manuscript wasimproved through comments from J. Bowsher (North DakotaState University), J.P. Rinehart (USDA-ARS, Fargo, North Dakota),and two anonymous reviewers. This study was partially sup-ported by DGICYT project AGR. 91-0988 and CONSOLIDER pro-ject CSD2008-00040 (Spanish Ministry of Science andInnovation) to J.B., and a Ph.D. scholarship from the Universityof Bologna to F.S.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.jinsphys.2012.04.008.
F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933 931

Author's personal copy
References
Ayres, M.P., Scriber, J.M., 1994. Local adaptation to regional climates in Papiliocanadensis (Lepidoptera: Papilionidae). Ecological Monographs 64, 465–482.
Bale, J.S., Hayward, S.A.L., 2010. Insect overwintering in a changing climate. TheJournal of Experimental Biology 213, 980–994.
Bartomeus, I., Ascher, J.S., Wagner, D., Danforth, B.N., Colla, S., Kornbluth, S.,Winfree, R., 2011. Climate-associated phenological advances in bee pollinatorsand bee-pollinated plants. Proceeding of National Academy of Sciences of theUnited States of America 108, 20645–20649.
Beck, S.D., 1983. Insect thermoperiodism. Annual Review of Entomology 28,91–108.
Bosch, J., 1994. The nesting behaviour of the mason bee Osmia cornuta (Latr) withspecial reference to its pollinating potential (Hymenoptera, Megachilidae).Apidologie 25, 84–93.
Bosch, J., Blas, M., 1994. Effect of over-wintering and incubation temperatures onadult emergence in Osmia cornuta Latr (Hymenoptera, Megachilidae).Apidologie 25, 265–277.
Bosch, J., Kemp, W.P., 2000. Development and emergence of the orchard pollinator,Osmia lignaria (Hymenoptera, Megachilidae). Environmental Entomology 29, 8–13.
Bosch, J., Kemp, W.P., 2003. Effect of wintering duration and temperature onsurvival and emergence time in males of the orchard pollinator Osmialignaria (Hymenoptera: Megachilidae). Environmental Entomology 32, 711–716.
Bosch, J., Kemp, W.P., 2004. Effect of pre-wintering and wintering temperatureregimes on weight loss, survival, and emergence time in the mason bee Osmiacornuta (Hymenptera: Megachilidae). Apidologie 35, 469–479.
Bosch, J., Vicens, N., 2002. Body size as an estimator of production costs in a solitarybee. Ecological Entomology 27, 129–137.
Bosch, J., Vicens, N., 2005. Sex allocation in the solitary bee Osmia cornuta: dofemales behave in agreement with Fisher’s theory? Behavioral Ecology andSociobiology 59, 124–132.
Bosch, J., Vicens, N., 2006. Relationship between body size, provisioning rate,longevity and reproductive success in females of the solitary bee Osmia cornuta.Behavioral Ecology and Sociobiology 60, 26–33.
Bosch, J., Kemp, W.P., Peterson, S.S., 2000. Management of Osmia lignaria(Hymenoptera: Megachilidae) populations for almond pollination: methods toadvance bee emergence. Environmental Entomology 29, 874–883.
Bosch, J., Maeta, Y., Rust, R.W., 2001. A phylogenetic analysis of nest nestingbehaviour in the genus Osmia (Hymenoptera: Megachilidae). Annals of theEntomological Society of America 94, 617–627.
Bosch, J., Kemp, W.P., Trostle, G.E., 2006. Bee population returns and cherry yields inan orchard pollinated with Osmia lignaria (Hymenoptera: Megachilidae).Journal of Economic Entomology 99, 408–413.
Bosch, J., Sgolastra, F., Kemp, W.P., 2010. Timing of eclosion affects diapausedevelopment, fat body consumption and longevity in Osmia lignaria, aunivoltine, adult-wintering solitary bee. Journal of Insect Physiology 56,1949–1957.
Bradshaw, W.E., Holzapfel, C.M., 2010. Insects at not so low temperature: climatechange in the temperate zone and its biotic consequences. In: Denlinger, D.L.,Lee, R.E. (Eds.), Low Temperature Biology of Insects. Cambridge University Press,pp. 242–275.
Bradshaw, W.E., Armbruster, P.A., Holzapfel, C.M., 1998. Fitness consequences ofhibernal diapause in the pitcher-plant mosquito, Wyeomyia smithii. Ecology 79,1458–1462.
Bryant, S.R., Bale, J.S., Thomas, C.D., 1999. Comparison of development andgrowth of nettle-feeding larvae Nymplaidae (Lepidoptera) under constant andalternating temperature regimes. European Journal of Entomology 96,143–148.
Danks, H.V., 1987. Insect Dormancy: An Ecological Perspective. Biological Survey ofCanada Monopraph, Ottawa.
Denlinger, D.L., 2002. Regulation of diapause. Annual Review of Entomology 47, 93–122.
Fontinou, A.A., Chatzoglou, C.S., Kagkov, E.A., 2002. Thermoperiodic effects ondiapause of Sesamia nonagrioides (Lepidoptera: Noctuidae). European Journal ofEntomology 99, 421–425.
Forrest, J.R.K., Thomson, J.D., 2011. An examination of synchrony between insectemergence and flowering in Rocky Mountain meadows. Ecological Monographs81, 469–491.
Gomi, T., Nagasaka, M., Fukuda, T., Hagihara, H., 2007. Shifting of the life cycle andlife-traits of the fall webworm in relation to climate change. EntomologiaExperimentalis et Applicata 125, 179–184.
Hahn, D.A., Denlinger, D.L., 2007. Meeting the energetic demands of insectdiapause: nutrient storage and utilization. Journal of Insect Physiology 53,760–773.
Hahn, D.A., Denlinger, D.L., 2011. Energetic of insect diapause. Annual Review ofEntomology 56, 103–121.
Hegland, S.J., Nielsen, A., Lázaro, A., Bjerknes, A.-L., Totland, Ø., 2009. How doesclimate warming affect plant-pollinator interactions? Ecology Letters 12, 184–195.
Higley, L.G., Pedigo, L.P., Ostlie, K.R., 1986. DEGDAY: a program for calculatingdegree-days, and assumptions behind the degree-day approach. EnvironmentalEntomology 15, 999–1016.
Kalushkov, P., Hodková, M., Nedved, O., Hodek, I., 2001. Effect of thermoperiod ondiapause intensity in Pyrrhocoris apterus (Heteroptera Pyrrhocoridae). Journal ofInsect Physiology 47, 55–61.
Kemp, W.P., Bosch, J., 2005. Effect of temperature on Osmia lignaria (Hymenoptera:Megachilidae) prepupa-adult development, survival, and emergence. Journal ofEconomic Entomology 98, 1917–1923.
Kemp, W.P., Bosch, J., Dennis, B., 2004. Oxygen consumption during the life cycle ofthe prepupa-wintering bee Megachile rotundata (F.) and the adult-wintering beeOsmia lignaria Say (Hymenoptera: Megachilidae). Annals of the EntomologicalSociety of America 97, 161–170.
Kiritani, K., 2006. Predicting impacts of global warming on population dynamicsand distribution of arthropods in Japan. Population Ecology 48, 5–12.
Kostal, V., 2006. Eco-physiological phases of insect diapause. Journal of InsectPhysiology 52, 113–127.
Maeta, Y., Nakanishi, K., Fujii, K., Kitamura, K., 2006. Exploitation of systems to use aunivoltine Japanese mason bee, Osmia cornifrons (Radoszkowski), throughoutthe year for pollination of greenhouse crops (Hymenoptera: Megachilidae).Chugoku Kontyu 20, 1–17.
Masaki, S., 1980. Summer diapause. Annual Review of Entomology 25, 1–25.Mironidis, G.K., Savopoulou-Soultani, M., 2012. Effects of constant and changing
temperature conditions on diapause induction in Helicoverpa armigera(Lepidoptera: Noctuidae). Bulletin of Entomological Research 102, 139–147.
Morden, R.D., Waldbauer, G.P., 1980. Diapause and its termination in the psychidmoth, Thyridopteryx ephemeraeformis. Entomologia Experimentalis et Applicata28, 322–333.
Musolin, D.L., Tough, D., Fujisaki, K., 2010. Too hot to handle? Phenological and life-history responses to simulated climate change of the southern green stink bugNezara viridula (Heteroptera: Pentatomidae). Global Change Biology 16, 73–87.
Nechols, J.R., Tauber, M.J., Tauber, C.A., 1987. Geographical variability inecophysiological traits controlling dormancy in Chrysopa oculata (Neuroptera:Chrysopidae). Journal of Insect Physiology 33, 627–633.
Ohtsu, T., Kimura, M.T., Hori, S.H., 1995. The influence of eclosion timing on wintersurvival and triacylglycerol accumulation in four temperate species ofDrosophila. Physiological Entomology 20, 248–252.
Papanatasiou, S.A., Nestel, D., Diamantidis, A.D., Nakas, L.T., Papadopoulos, N.T.,2011. Physiological and biological patterns of highland and a coastal populationof the European cherry fruit fly during diapause. Journal of Insect Physiology 57,83–93.
Peters, D.S., 1977. Systematik und Zoogeographie der westpaleärktischen Arten vonOsmia’s str., Monosmia, und Orientosmia. Senckenbergiana Biologica 58, 287–346.
Pruess, K.P., 1983. Day-degree methods for pest management. EnvironmentalEntomology 12, 613–619.
Pullin, A.S., 1987. Adult feeding time, lipid accumulation, and overwintering inAglais urticae and Inachis io (Lepidoptera: Nymphalidae). Journal of Zoology211, 631–641.
Radmacher, S., Strohm, E., 2011. Effects of constant and fluctuating temperatures onthe development of the solitary bee Osmia bicornis (Hymenoptera:Megachilidae). Apidologie 42, 711–720.
Ratte, H.T., 1984. Temperature and insect development. In: Hoffamann, K.H. (Ed.),Environmental Physiology and Biochemestry of Insects. Springer, Berlin, pp. 33–66.
Rust, R.W., 1974. The systematics and biology of the genus Osmia, subgenera Osmia,Chalcosmia and Cephalosmia (Hymenoptera: Megachilidae). The WasmannJournal of Biology 32, 1–93.
Saulich, A.K., Musolin, D.L., 1996. Univoltinism and its regulation in some temperatetrue bugs (Heteroptera). European Journal of Entomology 93, 507–518.
Sgolastra, F., Bosch, J., Molowny-Horas, R., Maini, S., Kemp, W.P., 2010. Effect oftemperature regime on diapause intensity in an adult-wintering Hymenopteranwith obligate diapause. Journal of Insect Physiology 56, 185–194.
Sgolastra, F., Kemp, W.P., Buckner, J.S., Pitts-Singer, T.L., Maini, S., Bosch, J., 2011. Thelong summer: pre-wintering temperatures affect metabolic expenditure andwinter survival in a solitary bee. Journal of Insect Physiology 57, 1651–1659.
Stephen, W.P., Undurraga, J.M., 1976. X-radiography, an analytical tool inpopulation studies of the leafcutter bee, Megachile pacifica. Journal ofApicultural Research 15, 81–87.
Tauber, C.A., Tauber, M.J., Nechols, J.R., 1987. Thermal requirements fordevelopment in Chrysopa oculata: a geographically stable trait. Ecology 68,1479–1487.
Tauber, M.J., Tauber, C.A., Masaki, S., 1986. Seasonal Adaptations of Insects. OxfordUniversity Press, Oxford, UK.
Taylor, F., 1986. The fitness functions associated with diapuse induction inarthropods I. The effects of age structure. Theoretical Population Biology 30,76–92.
Torchio, P.F., 1989. In-nest biologies and immature development of three Osmiaspecies (Hymenoptera: Megachilidae). Annals of the Entomological Society ofAmerica 82, 599–615.
Torchio, P.F., Bosch, J., 1992. Biology of Tricrania stansburyi, a meloid beetlecleptoparasite of the bee Osmia lignaria propinqua (Hymenoptera:Megachilidae). Annals of the Entomological Society of America 85, 713–721.
Torchio, P.F., Tepedino, V.J., 1982. Parsivoltinism in three species of Osmia bees.Psyche 89, 221–238.
Yamashiro, C., Ando, Y., Masaki, S., 1998. Thermoperiod reduces the thermalconstant required for oviposition in the leaf beetle Atrachya menetriesi.Entomological Science 1, 299–307.
932 F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933

Author's personal copy
Yocum, G.D., Kemp, W.P., Bosch, J., Knoblett, J.N., 2005. Temporal variation inoverwintering gene expression and respiration in the solitary bee Megachilerotundata. Journal of Insect Physiology 51, 621–629.
Yocum, G.D., Kemp, W.P., Bosch, J., Knoblett, J.N., 2006. Thermal history influencesdiapause development in the solitary bee Megachile rotundata. Journal of InsectPhysiology 52, 1113–1120.
F. Sgolastra et al. / Journal of Insect Physiology 58 (2012) 924–933 933





























