Embed Size (px)
Citation preview

Journal of Plankton Research VoL18no.8 pp.1407-1412, 1996
Summer diapause in Daphnia as a reaction to the presence offish
Joanna Pijanowska1 and Gisela Stolpe2
Max-Planck Institutfur Limnologie, Postfach 165, D-24302 Plon, FRG1 Permanent address: Department of Hydrobiology, University of Warsaw,Banacha 2, 00 097 Warsaw, PolandPermanent address: INA, Insel ofVilm, D-18581 Lauterbach, FRG
Abstract A chemical signal, released by a fish predator under summer-like high water temperatureand long-day photoperiod, caused the formation of resting eggs in a clone of Daphnia magna. Noephippial females were recorded and no ephippia were released in the control treatment during 45days of the experiment. When exposed to fish water, the fraction of ephippial females reached amaximum of 3.7%, a value comparable to that registered in summer in the GroBer Binnensee (North-ern Germany), a hypertrophic lake inhabited by fish, which was the source lake for our experimentalclone. The number of ephippia released within 45 days was on average 34 ± 22. Ephippia formationcould not result from the between-treatment differences in population density, and related patterns offood depletion, since no substantial difference between control and fish treatment was observed.Instead, specific information on the presence of a predator provided a cue which induced the formationof resting eggs in Daphnia. Under heavy predation and very low survival probability of parthenogeneticfemales, ephippia formation in summer can be adaptive, i.e. higher fitness can be achieved through sur-vival in the diapausing state than through the immediate reproductive gain via parthenogenesis.
Introduction
In planktonic animals, diapause is generally considered to be adaptive: once livingconditions deteriorate, animals can withdraw from the active phase of the life cyclein open water and bide their time in the form of resting eggs, which are often sexual.Although there is a general consensus which considers diapause in cyclicallyparthenogenetic animals as an escape from temporarily unfavorable conditions,there is still no agreement on what are the ultimate causes inducing the formationof inactive stages. While the majority of published data provide evidence that dia-pause is an adaptation to avoid physically harsh environments (Hutchinson, 1967),only in a few papers (Strickler and Twombly, 1975; Nilssen, 1977,1980; Hairstonand Munns, 1984; Hairston, 1987) is predation considered as an ultimate factorfavoring diapause in copepods. Until the most recent paper by Slusarczyk (1995),it has not been shown convincingly that cladoceran diapause may also haveevolved as an adaptation to avoid predation.
In temperate zone lakes, timing of the cladoceran diapause which follows sexualreproduction would precede the period of adverse conditions in late fall andwinter; the reports on diapause during other times of the year are only anecdotal(FldBner, 1972; Frey, 1982).
Many studies reveal the significance of different environmental factors as stimulifor the onset of sexual reproduction in cladocerans: daylength (Stross and Hill, 1965,1968; Stross, 1969a,b), low temperature (Mortimer, 1935), low food (D'Abramo,1980; Carvalho and Hughes, 1983), various combinations of these factors (i.e. Berg,1934;Banta, 1939;Bunner and Halcrow, 1977),and the simultaneous action of short
O Oxford University Press 1407
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from

J.Pijanowska and G-Stolpe
day, low food and chemically mediated information on crowding (Kleiven el ai,1992).
There is increasing evidence that planktonic prey can directly assess predationrisk, via the recognition of specific signals released by a predator into the water.Many aspects of the morphology, behavior and life-history patterns of planktonicanimals have evolved as responses to important changes in selective pressures,induced by predators' exudates (for a review, see Larsson and Dodson, 1993). Wechecked whether ephippia formation in Daphnia could be directly induced bychemicals released into the environment by a fish predator. The clone used in theexperiment was isolated from the GroBer Binnensee (hypertrophic, brackish lakeinhabited by fish) and proved to be sensitive to predatory cues, both in terms ofbehavioral (Dawidowicz and Loose, 1992; Pijanowska, 1994) and life-historyresponses (Weider and Pijanowska, 1993). The chemical presence of fish wasassociated in our experiment with long day, high water temperature and high foodconcentration, to simulate a summer scenario.
Method
The experiment was run in two treatments, without (control treatment) and withfish chemicals (fish treatment); each of the treatments was replicated five times.The animals were fed with a suspension of the green alga Scenedesmus acutus,adjusted daily to 2.0 mg C I"1.
Initially,20 animals of 0.98 ± 0.14 mm initial size (mean ± 1 SD) were introducedto each of the 10 experimental glass jars, which contained 11 of the desired medium.All animals were born within 24 h, as second clutch offspring of 100 mothers whichwere kept under high food concentration (2.0 mg C H) from juveniles until theyreleased the experimental offspring. Aged lake water (aerated for 7 days and fil-tered through 0.45 (am filters prior to use) was used as a culture medium for bothcontrol and fish treatments.Two cyprinids (Leucaspius delineatus L.) of mean totallength 6.2 ± 0.2 cm (mean ± 1 SD) were allowed to swim freely for 24 h in 10 1 ofaged water medium, and they were fed daily, each with 100 Daphnia magna(>2 mm). Prior to use, water was filtered through 0.45 um.
For 45 days, animals were transferred daily to fresh (control and fish) media witha pipette. During transfer, they were counted, and the numbers of females carry-ing subitaneous and ephippial eggs were registered. The number of ephippiareleased into the water was also recorded daily. Exuviae, dead animals and releasedephippia were carefully removed to avoid transferring them to fresh media. Start-ing from the 30th day of the experiment, animals were still transferred to freshmedia daily, but were counted only every third day. Ephippia, if any were released,were still counted daily.
The study was conducted at 19°C, under a 17 h light: 7 h dark photoperiod (withsunset at 22:00 h and sunrise at 5:00 h). Oxygen concentration and pH wereoccasionally checked, and no differences were registered between treatments.
1408
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from
Summer diapause in Daphnia
I
» » • • • » • • • * • •
days
day« days
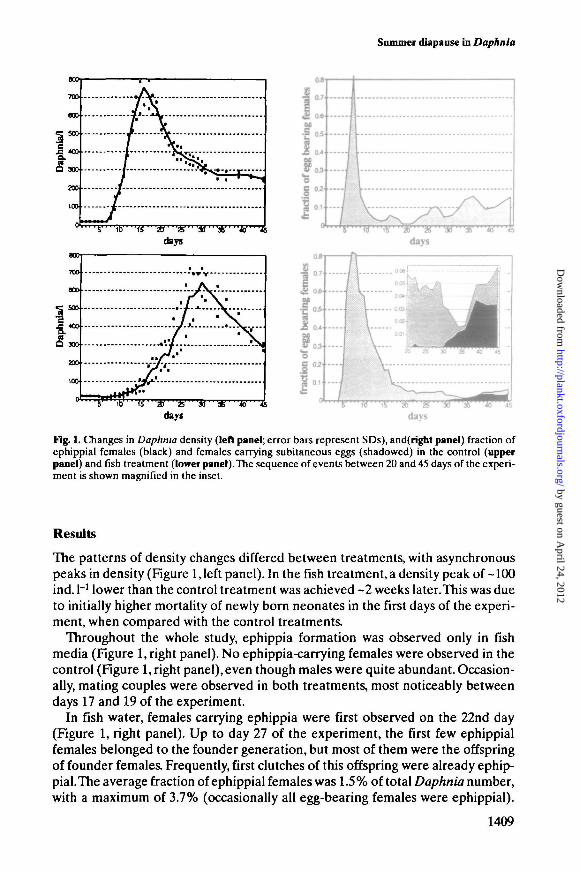
Fig. L Changes in Daphnia density (left panel; error bars represent SDs), and(right panel) fraction ofephippial females (black) and females carrying subitaneous eggs (shadowed) in the control (upperpanel) and fish treatment (lower panel). The sequence of events between 20 and 45 days of the experi-ment is shown magnified in the inset.
Results
The patterns of density changes differed between treatments, with asynchronouspeaks in density (Figure 1, left panel). In the fish treatment, a density peak of -100ind. I"1 lower than the control treatment was achieved ~2 weeks later. This was dueto initially higher mortality of newly born neonates in the first days of the experi-ment, when compared with the control treatments.
Throughout the whole study, ephippia formation was observed only in fishmedia (Figure 1, right panel). No ephippia-carrying females were observed in thecontrol (Figure 1, right panel), even though males were quite abundant. Occasion-ally, mating couples were observed in both treatments, most noticeably betweendays 17 and 19 of the experiment.
In fish water, females carrying ephippia were first observed on the 22nd day(Figure 1, right panel). Up to day 27 of the experiment, the first few ephippialfemales belonged to the founder generation, but most of them were the offspringof founder females. Frequently, first clutches of this offspring were already ephip-pial. The average fraction of ephippial females was 1.5 % of total Daphnia number,with a maximum of 3.7% (occasionally all egg-bearing females were ephippial).
1409
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from

J.Pijanowska and G£tolpe
The average number of ephippia released up to day 45 of the experiment was 34.4± 22.5 (mean ±1 SD) in the fish treatment.
Discussion
Of the two stimuli described by Hobaek and Larsson (1990) as necessary topromote a switch from female to male offspring production (short day, chemicallymediated crowding), and of the three invoked most recently by Kleiven etal. (1992)as necessary for the onset of sexual reproduction and subsequent formation ofresting eggs (short day, chemically mediated crowding and low food), populationdensities and resource level did not differ enough between control and fish treat-ments to generate such a substantial difference in Daphnia reproductive behavior.We can exclude a possibility that fish treatment provided a more 'crowded' situ-ation or more severe food depletion than controls, since population density in thefish treatment never reached the maximum control level, and was much lower thanin the control treatment until the onset of ephippia formation. The presence ofmales in control and fish media would also indicate that crowding 'perception' wasquite similar in both treatments. Apparently, the specific information of the chemi-cal presence of a predator, associated with the realistic approximation of summerlight and temperature regimes, provides an additional stimulus for limited sexualreproduction in planktonic cladocerans.
In the GroBer Binnensee, which was the source lake for our experimental clone,the permanent presence of males and the frequent appearance of ephippialfemales of D.magna in summer was reported by Lampert (1991) and by Stolpe(1993). The population consisted occasionally (in summer 1987) or permanently(in summer 1991) of as many as 20% males, while ephippial females contributedto total Daphnia densities at - 3 % (1991) on average, only sometimes exceeding10% (1987). The presence of males and ephippia production in summer in theGroBer Binnensee does not seem to result either from crowding or from lowresource levels. In summer 1987, throughout the period where both were moni-tored in the lake, Daphnia density averaged around 11 ind. I"1 (from Lampert,1991) and particulate organic carbon concentration in the fraction of seston par-ticles <30 urn was on average 0.6 mg C I"1 (Lampert and Rotthaupt, 1991). Insummer 1991, these values averaged 30 ind. H and 1.1 mg C I"1 respectively (Stolpe,1993); thus, food was less limiting in the lake than in our experiments.
In cladocerans, the possibility of predator-induced sexual reproduction, fol-lowed by diapausing of fertilized eggs, has been proposed only recently by Slusar-czyk (1995). Other evidence is scarce and comes from anecdotal observations(J.Pijanowska, LJ.Weider and A.Howe, unpublished data; J.Luning, unpublisheddata).
Ephippia formation in summer under heavy predation can be adaptive. In theGroBer Binnensee, many females are eaten before they manage to release theirfirst offspring, even before attaining the size at which they could reproduce. Onlyseldom, if ever, do they manage to release later clutches (Lampert, 1991). There-fore, fitness gain from survival as resting eggs can be higher than that achievedfrom instantaneous reproduction, which is small anyway. Finally, even if an
1410
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from

Summer diapause in Daphnia
ephippial female is eaten by fish, eggs can remain viable after passing through afish gut (Mellors, 1975). Under heavy predation, when the survival probability ofparthenogenetic females is low (or Daphnia assess it as low from the perceptionof their chemical environment), higher fitness can be achieved through survival ina diapausing state during adverse periods than through immediate reproductionvia parthenogenesis (Levins, 1969; Cohen, 1970; Nunney, 1989; Roughgarden,1991). Ensuring the survival of even a few propagules can provide 'clonal safety'against adverse events, which could otherwise endanger, if not wipe out, the wholelineage (Frey, 1982). A small fraction of individuals entering diapause, and thesimultaneous presence of parthenogenetically reproducing females, can provideadvantages from sexual reproduction once the predation risk is high, and allowthe immediate gain via asexual reproduction once the predation risk falls again.The obvious advantage of plasticity in response to a stimulus from a predator isto maximize fitness if predatory events are unpredictable, and costs of sexualreproduction and of withdrawing from active stages are presumably high.
Under a predictable predation regime, the evolutionary stable strategy ESS(Maynard Smith, 1982) for initiating diapause would be to produce ephippialclutches to match the first meaningful onset of increased predation risk (Hairston,1987). In our experimental conditions, however, the response to a stimulus was notimmediate. In contrast to most founder females, their offspring from early clutchesresponded to the stimulus. The same delay was reported by Carvalho and Hughes(1983) and Slusarczyk (1995) for D.magna, and by Stross and Hill (1965) in theresponse of Daphnia pulex to short daylength. Under our experimental conditions,it happened very often that the first broods of primiparous females were alreadyephippial. The sensitivity to the stimulus, therefore, and a correct assessment ofthe environmental situation must take place early during ontogenesis.
Acknowledgements
Many thanks go to Winfried Lampert and Maciej Gliwicz for their inspiring com-ments at all stages of our study. Herwig Stibor is kindly acknowledged for scien-tific, and Larry Weider for scientific and linguistic review of the manuscript. PetterLarsson,Esa Ranta and Raymond Stross kindly provided critical comments on anearlier version of the manuscript.
ReferencesBanta^A.M. (1939) Studies on the Physiology, Genetics and Evolution of Some Cladocera. Publication
513, Carnegie Department of Genetics, Paper 39. Institution of Washington, Washington, DC.Berg,K- (1934) Cyclic reproduction, sex determination and depression in the Cladocera. BioL Rev., 9,
139-174.Bunner,H.C. and Halcrow.K. (1977) Experimental induction of the production of ephippia by Daphnia
magna Straus (Cladocera). Crustaceana, 32,77-86.Carvalho,G.R. and Hughes,R.N. (1983) The effect of food availability, female culture-density and
photoperiod on ephippia production in Daphnia magna Straus (Crustacea: Cladocera). FreshwaterBioL, 13,37-46.
Cohen JD. (1970) A theoretical model for the optimal timing of diapause. Am. Nat,, 104,389-400.
1411
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from

J.Pijanowska and GJStorpe
D'Abramo.L.R. (1980) The stimulus for the asexual-sexual switch in Moma macrocopa. Limnol.Occanogr.,2S,422-A29.
Dawidowicz,P. and Loose,C. (1992) Metabolic costs during predator-induced diel vertical migration ofDaphnia. Limnol. Oceanogr., 37,1589-1595.
FloBner.D. (1972) Krebstiere, Crustacea. Kiemen und Blatfiifier, Branchiopoda, FischlOuse, Branchiura.Die Tierwell Deutschlands, 60 Teil. Gustav Fischer Verlag, Jena.
Frey.D.G. (1982) Contrasting strategies of gamogenesis in northern and southern populations ofCladocera. Ecology, 63,223-241.
Hairston.N.G.Jr (1987) Diapause as a predator-avoidance adaptation. In Kerfoot.W.C. and Sih,A.(eds), Predation. Direct and Indirect Impacts on Aquatic Communities. University Press of NewEngland, Hanover, NH, pp. 281-290.
Hairston,N.G.,Jr and Munns,W.R.rlr (1984) The timing of copepod diapause as an evolutionary stablestrategy. Am. Nat., 123,733-751.
Hobaek.A. and Larsson.P. (1990) Sex determination in Daphnia magna. Ecology, 7,2255-2268.Hutchinson,G.E. (1967) A Treatise on Limnology. VoL 2: Introduction to Lake Biology and the Limno-
plankton. John Wiley & sons. New York.Kleiven.O.T., Larsson,P. and Hobaek,A. (1992) Sexual reproduction in Daphnia magna requires three
stimuli. Oikos, 65,197-206.Lampert,W. (1991) The dynamics of Daphnia magna in a shallow lake. Verh. Int. Ver. Limnol., 24,
795-798.Lampert.W. and Rothhaupt.K.O. (1991) Alternating dynamics of rotifers and Daphnia magna in a
shallow lake. Arch. Hydrobiol., 120,447-456.Larsson^R and Dodson,S.I. (1993) Chemical communication in planktonic animals. Arch. Hydrobiol,
129,129-155.Levins,R. (1969) Dormancy as an adaptive strategy. Symp. Soc. Exp. Bioi, 23,1-10.Maynard SmithJ. (1982) Evolution and the Theory of Games. Cambridge University Press, Cambridge.Mellors,W.R. (1975) Selective predation of ephippial Daphnia and the resistance of ephippial eggs to
digestion. Ecology, 56,974-980.Mortimer,C.H. (1935) Untersuchungen flber den Generationswechsel der Qadoceren. Natunvis-
NilssenJ.P. (1977) Cryptic predation and the demographic strategy of two limnetic cyclopoid cope-pods. Mem. 1st. hot IdrobioL, 34,187-196.
NilssenJ.P. (1980) When and how to reproduce: a dilemma for limnetic cyclopoid copepods. InKerfoot,W.C. (ed.), Evolution and Ecology of Zooplankton Communities. University Press of NewEngland, Hanover, NH, pp. 418-426.
Nunney.L. (1989) The maintenance of sex by group selection. Evolution, 43,245-257.PijanowskaJ. (1994) Fish-enhanced patchiness in Daphnia distribution. Verh. Int. Ver. Limnol, 25,
2366-2368.Roughgarden J. (1991) The evolution of sex. Am. Nat, 138,934-953.Slusarczyk,M. (1995) Predator-induced diapause in Daphnia. Ecology, 76,1008-1013.StolpcG. (1993) Populationsdynamik und biotische Interaktionen von Daphnia magna im Grossen
Binnensee. MSc Thesis, Max-Planck Institut fuer Limnologie, Abteilung Oekophysiologie, Ploen,FRG.
StricklerJ.R. and Twombly,S. (1975) Reynolds number, diapause, and predatory copepods. Int. Ver.Theor. Angew. Limnol Verh., 19,2943-2950.
Stross,R.G. (1969a) Photoperiod control of diapause in Daphnia. II. Induction of winter diapause inthe arctic BioL Bull., 136,264-273.
StrossJt.G. (1969b) Photoperiod control of diapause in Daphnia. III.Two-stimulus control of long-day,short-day induction. BioL Bull, 137,359-374.
Stross,R.G. and HillJ.G. (1965) Diapause induction in Daphnia requires two stimuli. Science, 150,1462-1464.
Stross^-G. and HillJ.G. (1968) Photoperiod control of winter diapause in the freshwater crustaceanDaphnia. BioL Bull, 134,176-198.
WeiderJ-J. and PijanowskaJ. (1993) Plasticity of Daphnia life histories in response to chemical cuesfrom predators Oikos, 67,385-392.
Received on September 19,1995; accepted on March 18,1996
1412
by guest on April 24, 2012
http://plankt.oxfordjournals.org/D
ownloaded from





























