Embed Size (px)
Citation preview

PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Effect of repeated in vitro sub-culturing on the virulence of Metarhiziumanisopliae against Helicoverpa armigera (Lepidoptera: Noctuidae)
Pallavi B. Nahara, Shuklangi A. Kulkarnia, Mahesh S. Kulyea,Santosh B. Chavana, Girish Kulkarnib, Armugham Rajendrana,Priya D. Yadava, Yogesh Shoucheb and Mukund V. Deshpandea,*
aBiochemical Sciences Division, National Chemical Laboratory, Pune, India; bMolecularBiology Unit, National Centre for Cell Science, Pune, India
(Received 26 October 2007; returned 31 December 2007; accepted 22 January 2008)
The effect of repeated conidial sub-culturing of Metarhizium anisopliae on itsvirulence against Helicoverpa armigera (Hubner) was studied. The LT50 observedagainst third instar larvae of H. armigera for the first sub-culture was 3.4 days; itincreased to 4.5 and 5.6 days for the 20th and the 40th sub-cultures, respectively.The LT50 values after passage of the 40th sub-culture on H. armigera decreased to4.4 and 3.7 days for the 40th (first in vivo) and the 40th (fifth in vivo) passages,respectively. Similarly, the LC50 of M. anisopliae towards third instar larvae of H.armigera increased from the first sub-culture (0.17!104) to (3.0!104) for the40th conidial transfers on potato dextrose agar and again decreased to 0.74!104
and 0.23!104 in the 40th (first in vivo) and the 40th (fifth in vivo) passage,respectively. Similar trends for LC50 and LT50 values were seen when sugarcanewoolly aphid, Ceratovacuna lanigera Zehntner was used as a host. Significantvariation in appressorium formation and cuticle-degrading enzyme productionsuch as chitinase, chitin deacetylase, chitosanase and protease during subsequentsub-culturing and passage through H. armigera was observed. Though there wasno effect on internal transcribed spacer (ITS) sequence pattern, interestingly, inrandomly amplified polymorphic DNA (RAPD), significant differences in theband intensities and in the banding pattern for different sub-cultures of M.anisopliae were observed. As stable virulence towards the insect pest is desirablefor commercialisation of a mycoinsecticide, such changes in virulence due torepeated in vitro transfer need to be monitored and minimised.
Keywords: in vitro sub-culturing; Metarhizium anisopliae; Helicoverpa armigera;appressorium formation; chitinase; protease; lipase; chitin deacetylase
Introduction
Commercial success of any mycoinsecticide depends significantly on the virulentnature of the infective propagules, such as conidia, that are sprayed in the field.Moreover, the stability of a fungal strain during repeated conidial sub-culturing onartificial media is very important for making it commercially feasible. Theentomopathogenic fungi degenerate as a result of loss of virulence and change inmorphology when successively sub-cultured on artificial media (Butt, Wang, Shah,and Hall 2006). However, the extent of the effect of repeated in vitro sub-culturing
*Corresponding author. Email: [email protected]
ISSN 0958-3157 print/ISSN 1360-0478 online
# 2008 Taylor & Francis
DOI: 10.1080/09583150801935650
http://www.informaworld.com
Biocontrol Science and Technology,Vol. 18, No. 4, 2008, 337"355
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

on viability and morphological, biochemical, and molecular characteristics, andmost importantly, on the virulence and host specificity, varies considerably withinentomopathogenic fungal strains. In this regard, several different observations of theeffect of repeated in vitro sub-culturing on the virulence of different entomopatho-gens have been reported. For instance, Morrow, Boucias, and Heath (1989) reporteda decrease in the ability of Nomuraea rileyi to produce yeast-like hyphal bodies aftersix conidial transfers on Sabouraud maltose and yeast extract agar. Furtherdevelopment of avirulent progeny towards velvetbean caterpillar, Anticarsiagemmatalis, after 10 conidial transfers was also observed. A strain of Beauveriabassiana had decreasing virulence against Leptinotarsa decemlineata after 16conidial transfers on an artificial medium (Schaerffenberg 1964). However, Brown-bridge, Costa, and Jaronski (2001) observed no decrease in the virulence of B.bassiana toward Bemisia argentifolii following repeated in vitro transfers. Aschersoniaaleyrodis lost its virulence towards greenhouse whitefly, Trialeurodes vaporariorumafter 19 sub-cultures on artificial medium (Fransen, Winkelman, and van Lenteren1987). On the other hand, Vandenberg and Cantone (2004) noted that the virulenceof Paecilomyces fumosoroseus towards Diuraphis noxia or Plutella xylostella did notchange even after 30 in vitro transfers. Interestingly, they further reported thatdifferent host passages had varying effects on virulence. For instance, the virulenceof P. fumosoroseus towards D. noxia after 15 passages in P. xylostella decreased andwas not regained even after five passages in D. noxia. Also, there was no change inthe virulence of P. fumosoroseus towards both the insect hosts, D. noxia andP. xylostella after 15 passages in D. noxia. Paecilomyces farinosus also apparentlyretained its virulence towards the English grain aphid, Sitobion avenae after 15repeated conidial transfers (Hayden, Bidochka, and Khachatourians 1992).Similarly, Hall (1980) reported no significant attenuation of Verticillium lecaniitowards an aphid host, Macrosiphoniella sanborni after repeated sub-culturing ondifferent solid media.
Different possible factors for the decrease in virulence were envisaged. Forinstance, Morrow et al. (1989) suggested that the attenuation process in the case ofN. rileyi was associated with the sporulation process and multiple vegetative transfersdid not decrease its virulence towards A. gemmatalis. However, conidia ofM. anisopliae lost their virulence towards fourth and fifth instar Galleria mellonellaand Tenebrio molitor after a conidial transfer on a medium with high C:N ratio. Thevirulent conidia had relatively low endogenous C:N ratios and high germinationrates (Shah, Wang, and Butt 2005). However, the gross morphological andphysiological changes seen during single- and multi-spore transfers could not becorrelated with virulence in all cases. For instance, while studying 44 isolates ofAschersonia against whiteflies B. argentifolii and T. vaporariorum, Meekes, Fransen,and van Lenteren (2002) did not find any correlation between sporulation capacityand virulence.
It has been reported that in fungus"insect interactions, different events such asgermination of conidia; formation of infection structures such as appressoria andyeast-like cells useful for penetration and/or colonisation, respectively; productionof cuticle-degrading enzymes such as endo-chitinase (EC 3.2.1.14), chitin deace-tylase (EC 3.5.1.41), chitosanase (EC 3.2.1.132), alkaline protease (EC 3.4.21.62)and lipase (EC 3.1.1.3), as well as non-enzymic killing components such assecondary metabolites, play a significant role (St. Leger, Cooper, and Charnley
338 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

1986a; Krieger de Moraes, Schrank, and Vainstein 2003; Nahar, Ghormade, andDeshpande 2004; Chavan, Ghormade, Nahar, and Deshpande 2005). Recently, itwas reported by Shakeri and Foster (2007) that the entomopathogenicity of amycoparasitic Trichoderma harzianum towards T. molitor larvae was due toenzymes such as chitinase and protease as well as the antibiotic peptaibol.
Helicoverpa armigera, a polyphagous pest, attacks more than 182 plant species,including cotton, chickpea, pigeonpea, maize, sorghum, sunflower, tomato andmany other vegetables. M. anisopliae M34412 was found to be most effectiveagainst H. armigera in chickpea as well as in pigeonpea fields (Nahar et al. 2003).Therefore, the objective of the current study was to assess the effect of repeated invitro sub-culturing on the virulence of M. anisopliae M34412 mainly againstH. armigera. Here we have described the formation of appressoria by M. anisopliaeon artificial surfaces and possible changes in appressorium formation as a resultof repeated in vitro sub-culturing. We have also measured extracellular cuticle-degrading enzyme activities of M. anisopliae produced in liquid media usingconidial and mycelial inocula. The median lethal time (LT50) and median lethalconcentration (LC50) were determined for the mortality of H. armigera byM. anisopliae. The phenotypic changes in M. anisopliae as a consequence ofrepeated in vitro sub-culturing were correlated with the possible genotypic(RAPD) changes.
Materials and methods
Organism
Metarhizium anisopliae M34412 (MTCC 5190 " Microbial Type Culture Collection,Chandigarh, India) was isolated using the soil dilution method (Nahar et al. 2003).The stock culture was maintained on PDA (potato dextrose agar) at 88C untiluse.
For repeated in vitro conidial transfers, M. anisopliae was grown on PDA at 288Cwith 70"80% relative humidity (RH) until sporulation was evident (approximately7 days). The conidia were harvested from PDA slants by scraping with a loop andsub-cultured to fresh PDA slants. This multi-spore in vitro transfer was repeated upto 40 sub-cultures. The suspensions of harvested conidia (1st, 10th, 20th, 30th and40th in vitro sub-cultures) were prepared in 10% glycerol, placed in cryo-vials, andstored at "808C until use. For in vivo passage in the insect host, conidial suspensions(1!107 conidia/mL) of the 40th sub-culture of M. anisopliae were prepared. Thirdinstar larvae of H. armigera were dipped in the conidial suspension for 5 s, and keptin vials with okra (Abelmoschus esculentus) pieces and incubated at 288C with 70"80% RH. The conidia from cadavers were harvested by scraping with a pointed metalneedle, suspended in 0.1% (w/v) Tween 80 and used to inoculate (1!107 conidia/mL)additional insects. This insect passage was repeated 5 times. After passage on theinsect host, conidial suspensions of all passaged cultures [40th (first in vivo) to 40th(fifth in vivo)] were made and stored as previously described.
Growth, conidial germination and appressorium formation
To check the effect of repeated in vitro sub-culturing on growth, conidial suspensions(1!107 conidia/mL) of the first, 20th, 40th, 40th (first in vivo) and 40th (fifth in vivo)
Biocontrol Science and Technology 339
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

sub-cultures were inoculated on PDA plates. The PDA plates were incubated at 288Cfor 2 days. The mycelial colony (2 mm diameter) of 2-day-old culture was inoculatedat the centre of fresh PDA plates and incubated at 288C for 8 days. After 8 days, theplates were observed for difference in growth pattern.
For determining the effect of in vitro sub-culturing on conidial germination, theconidial suspensions (1!107 conidia/mL) of all the five [1st, 20th, 40th, 40th (first invivo) and 40th (fifth in vivo)] sub-cultures ofM. anisopliae were prepared in 0.1% (w/v)Tween 80 by scraping the conidia from the 10% glycerol stocks as described earlier(Nahar et al. 2003). The initial conidial count (1!107 conidia/mL) was adjustedusing a haemocytometer. The conidial suspensions that had been serially diluted to102 conidia/mL were inoculated on YPG agar (g/L): yeast extract, 3; peptone, 5;glucose, 10; agar, 20; pH, 6.8, slides. The plates containing YPG agar slides wereincubated at 288C with 70"80% RH for 24 h. The number of conidia forming germtubes was observed randomly in 10 different fields under a light microscope for 16 hat 2 h intervals. Approximately 30 conidia in each field were counted and eachexperiment was carried out in three different plates. For each plate, a minimum of300 conidia were recorded as germinated or ungerminated.
To study the appressorium formation on artificial surfaces, the conidialsuspensions (100 mL of 1!107 conidia/mL) of all the 5 sub-cultures of M. anisopliaeprepared in 0.1% (w/v) Tween 80 were inoculated separately in 5 mL YPG liquidmedium and incubated at 288C with 70"80% RH. Their growth was observed with alight microscope for the initiation of germination every 2 h. Once the germinationwas initiated, the nutrient source was depleted by centrifugation at 10,000!g for 10min as described by Xavier-Santos, Magalhaes, Elza, and Luna-Alves (1999). Thegerminated conidia were washed twice with sterile distilled water and the pellets weresuspended in 100-mL sterile distilled water. These suspensions were separately placedat the centre of polypropylene Petri plates which were sealed with parafilm andincubated at 288C with 70"80% RH. The plates were periodically observed under themicroscope for appressoria development over 24 h. The number of conidia producinggerm tubes and/or appressoria (minimum 30 conidia/field) was counted every 2 hrandomly in 10 different fields in each plate as described above. The experimentswere repeated twice using freshly prepared suspensions in Completely RandomizedBlock Design and the data of three experiments was pooled to get the average value.For statistical analysis, the data on percent appressoria formation was subjected toarcsine transformation to improve the homogeneity of variances and subjected toanalysis of variance. The means were compared by using an F-test at 5% level of acritical difference as described by Panse and Sukhatme (1989).
Production of cuticle-degrading enzymes
The YPG and the chitin medium containing (g/L): KH2PO4, 3; K2HPO4, 1;MgSO4, 0.7; (NH4)2SO4, 1.4; NaCl, 0.5; CaCl2, 0.5; yeast extract, 0.5; bacto-peptone, 0.5; urea, 0.3; oxgall, 1; trace metal solution, 1 mL (contained mg/mL:FeSO4, 5; MnSO4, 1.56; ZnSO4, 3.34; CoCl2, 2); chitin (Sigma, carbon source), 5;pH, 6 were prepared, for the extracellular constitutive and induced extracellularcuticle-degrading enzyme production experiments as described earlier (Vyas andDeshpande, 1989). The conidial suspensions (500 mL of 1!107 conidia/mL) of fivesub-cultures, viz. 1st, 20th, 40th, 40th (first in vivo) and 40th (fifth in vivo) prepared
340 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

in 0.1% (w/v) Tween 80 were inoculated in 50 mL YPG and chitin medium eachand incubated under shaking conditions (180 rpm) at 288C for 72"96 h. Theexperiments were carried out to check the levels of cuticle-degrading enzymesproduced when different in vitro and in vivo sub-cultured conidia were used as theinocula. All the experiments were carried out with the 1st, 20th, 40th, 40th (firstin vivo) and 40th (fifth in vivo) sub-cultures in triplicate unless otherwisementioned.
To ascertain the difference in the effect of single-spore and multi-spore transferon virulence factors, viz., cuticle-degrading enzyme production, if any, the conidiafrom the first and 40th sub-cultures were plated on PDA plates and incubated at288C. After 48 h, the single isolated colony was inoculated in YPG medium. Themycelium obtained after incubation at 288C for 48 h was added (5% amount) tofresh YPG and chitin medium for the extracellular cuticle-degrading enzymesproduction experiments as described earlier. The experiments were performed intriplicate.
The effect of serial transfer of mycelial inoculum on the production of cuticle-degrading enzyme activities was also studied. The vegetative mycelium was grown byinoculating conidia (1!107 conidia/100 mL medium) of the first sub-culture in YPGmedium. After every 24 h, the mycelium was transferred to YPG medium repeatedly10 times. The mycelium was harvested aseptically by filtration and stored in sterilevials at -208C until use. The 10% (w/v) first and 10th mycelial inocula wereinoculated in 50 mLYPG and chitin medium and incubated at 288C for 72"96 h. Forcomparison, the first and 10th sub-cultures of conidia were used as described above.
The chitinase, protease, and lipase activities were estimated after 96 h, while thechitin deacetylase, chitosanase activities were estimated after 72 h in YPG grownculture filtrate and after 96 h in chitin medium grown culture filtrate.
Enzyme assays
The total chitinase activity in the culture supernatant was estimated using acidswollen chitin as a substrate, the endo-chitinase with ethylene glycol chitin, 4-methyl-umbelliferyl-N?,Nƒ,Nƒ?-triacetylchitotrioside (4-MU-(GlcNAc)3), and N-acetylgluco-saminidase using 4-methylumbelliferyl-N-acetyl-b-d-glucosaminide (4-MU-GlcNAc)and p-nitrophenyl-N-acetyl-b-d-glucosaminide (pNP-GlcNAc) as described earlier(Ghormade, Lachake, and Deshpande 2000; Patil, Ghormade, and Deshpande 2000;Nahar et al. 2004). The chitosanase was measured using acid swollen chitosan as asubstrate. One unit of enzyme activity was expressed as 1 mmol N-acetylglucosamine(GlcNAc), 4-methyl umbelliferone, p-nitrophenol or glucosamine equivalentsproduced per minute. According to Nahar et al. (2004), the CDA activity wasestimated using ethylene glycol chitin as a substrate. One unit of enzyme released1 mmol of glucosamine from ethylene glycol chitosan formed per minute. Proteaseactivity was measured using Hammerstein casein as a substrate (Vyas andDeshpande 1989). One unit of enzyme liberated 1 mmol of tyrosine per minute.Lipase activity was determined as described by Pignede et al. (2000). The substrateemulsion was prepared with olive oil (20 mL) and gum arabic (165 mL, 10%, w/v,Sigma). One unit of lipase was the amount of enzyme that released 1 mmol of fattyacid per minute. Each experiment was carried out in three different flasks and theenzyme assays were carried out in triplicate with each flask. The experiment was
Biocontrol Science and Technology 341
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

repeated a further two times on different occasions using freshly prepared medium.The experiments were carried out in Completely Randomized Block Design and thedata of the three experiments was pooled to get the average value. For statisticalanalysis, the data was subjected to analysis of variance and the means were comparedby using an F-test at 5% level of critical difference as described by Panse andSukhatme (1989).
Insect rearing and bioassay
The initial culture of H. armigera was established by collecting larval and pupalstages of the insect from the fields. Larvae were reared individually in sterileautoclavable polypropylene vials (42!65 mm, 50 mL capacity; Laxbro Manufactur-ing Co., Pune, India) containing pieces of okra as a diet. Okra was first disinfectedfor 10 min with 0.5% sodium hypochlorite (Ignoffo, Futtler, Marston, Hostetter, andDickerson 1975). Insect eggs were also surface-sterilized with 0.5% sodiumhypochlorite. Conditions in the insect-rearing room were maintained at 25928Cand 6595%, respectively.
Third instar larvae of H. armigera were used for evaluating the effect of repeatedin vitro sub-culturing of M. anisopliae towards the susceptibility of the insect pest.The M. anisopliae sub-cultures*1st, 20th, 40th, 40th (first in vivo) and 40th (fifth invivo)*were grown on PDA at 288C for 7 days. Conidial suspensions of M. anisopliaesub-cultures were prepared by scraping conidia from 7-day-old culture, in 0.1% (w/v)Tween 80. Each bioassay was done with four concentrations of conidia (1!103, 1!105, 1!107 and 1!109 conidia/mL) to increase the possibility of identifying minorvariations in virulence, if any, that might go undetected if only a single dose was usedto determine median LC50. To determine median LT50 the day-wise mortality datawas used at the concentration of 1!107 conidia/mL. The experiments wereconducted in Completely Randomized Block Design with each treatment containingthe set of 30 larvae repeated three times. The set of 30 larvae with three replicationstreated with 0.1% (w/v) Tween 80 in sterile distilled water served as a control. Eachindividual larva was dipped in 10 mL conidial suspensions of M. anisopliae sub-cultures for 5 s. The same experiment was repeated a further two times using freshlyprepared conidial suspensions. After treatment, the individual larvae were trans-ferred to separate sterile vials containing moist Whatman No. 1 paper and a piece ofdisinfected okra. The okra pieces were changed every other day and the larvae werekept at 25928C, 6595% RH and 16L:8D for 14 days or until they died. Dead larvaewere transferred to sterile Petri plates containing moist cotton swabs and kept at28928C with 70"80% RH for at least 3"7 days to allow mycelial growth and conidiaformation over the cadavers. The data on percent mortality from three experimentswas pooled to get an average value, which was corrected by Abbott’s (1925) formula.The estimates of LT50 and LC50 for mortality at 14 days post-treatment were madeusing probit analysis (Finney 1981; Throne, Weaver, Chew, and Baker 1995).
To check the effect of repeated sub-culturing on the host specificity, if any, thebioassays were also carried out against third instar nymphs of the sugarcane woollyaphid, C. lanigera. In each set of experiments, sugarcane leaf discs (5!5 cm) with100 third instar nymphs were treated with conidial suspensions (1!103, 1!105, 1!107, 1!109 conidia/mL) of the 1st, 20th, 40th (first in vivo) and 40th (fifth in vivo)M. anisopliae cultures. The control was maintained with 100 nymphs treated with
342 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

sterile distilled water containing 0.1% (w/v) Tween 80. To determine LT50 the day-wise mortality data was used at a concentration of 1!107 conidia/mL. Spraying wascarried out using an atomiser (hand-spraying device for laboratory bioassay). Theexperiments were conducted in Completely Randomized Block Design with eachtreatment containing the set of 100 nymphs repeated three times. The sameexperiments were repeated a further two times using freshly prepared conidialsuspensions. After treatment, sugarcane leaf discs were kept in sterile Petri platescontaining moist filter paper and were incubated at 25928C with 70"80% RH for 8days. The nymphs on each leaf disc were monitored for the development of mycosison each individual insect. The mortality was recorded using a dissection microscopeat 4"64! magnification. The data was analysed to obtain LT50 and LC50 values asdescribed earlier.
ITS and RAPD analysis
Conidia from the 1st, 10th, 20th, 30th, 40th and 40th (first in vivo) sub-cultures ofM. anisopliae were selected for determining genotypic changes, if any, as a result of invitro repeated sub-culturing. Conidial suspensions (1!107 conidia/100 mL ofmedium) of these M. anisopliae sub-cultures were inoculated in a semi-syntheticcomplete medium containing (g/L): KH2PO4, 0.36; Na2HPO4, 1.6; KCl, 1; MgSO4,0.6; NH4NO3, 0.7; yeast extract, 5; glucose, 10; pH, 7 and incubated at 288C undershaking conditions (180 rpm) for 48 h. The mycelium from each flask was harvestedseparately by vacuum filtration and the genomic DNA was extracted using QiagenDneasy Plant Mini kit. The quantity and quality of DNA was assessed on a 1%agarose gel.
The genomic DNA was amplified by ITS 1(TCCGTAGGTGAACCTGCGG)and ITS 4 (TCCTCCGCTTATTGATATGC) primers in PE 9700 thermocycler. Thereaction mixture (25 mL) consisted of 10! Taq polymerase buffer (New EnglandBiolabs), 2 mM dNTPs, 10 pM primers, 1 unit Taq polymerase (New EnglandBiolabs), and 10 ng DNA. The PCR conditions were: an initial denaturation at 948Cfor 2 min, followed by 35 cycles of denaturation at 948C for 1 min, annealing at 508Cfor 1 min and extension at 728C for 1 min followed by final extension at 728C for 10min. The positive amplicons were purified by PCRCleanup Qiagen Kit according tothe instruction manual. Purified PCR products were sequenced on both the strandson an Ab3730DNA analyzer using the Big Dye terminator kit (Applied Biosystems,Inc. Foster City, CA). The sequences were analyzed at the National Center forBiotechnology Information (Bethesda, MD) (http//: www.ncbi.nlm.nih.gov/BLAST)for closed homology.
For RAPD analysis, five individual 10-nucleotide random primers were used forDNA amplification (947G, 5?-GGTTGGTGGG-3?; OPA13, 5?-CACCACCCAC-3?;OPA07, 5?-GAAACGGGTG-3?; OPA03, 5?-AGTCAGCCAC-3?; 935G, 5?-GGGTTGTGGG-3?). The reaction mixtures (20 mL) consisted of 10! Taqpolymerase buffer (New England Biolabs), 2mM dNTPs, 10 pM primers, 1 unitTaq polymerase (New England Biolabs), and 10 ng DNA. The PCR conditions were:an initial denaturation at 948C for 4 min, followed by 40 cycles of denaturation at948C for 1 min, annealing at 508C for 1 min and extension at 728C for 1 min followedby final extension at 728C for 10 min. The RAPD PCR amplicons were resolved in2.5% agarose gels in 1! Tris borate EDTA (TBE) buffer (pH 7.5) and stained with
Biocontrol Science and Technology 343
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

ethidium bromide. Visual analysis of stained gel photographs was used to detect anychange in banding patterns. The RAPD gels were replicated three times fordetermining the consistency of the results.
Results
Growth, conidial germination and appressorium formation
Vegetative growth of M. anisopliae on PDAwas observed at 288C for 8 days. All thesub-cultures grew profusely on PDA plates with dark green sporulation without anysectoring. The average size of the conidia was 7.1190.02!3.0590.03 mm. Repeatedin vitro sub-culturing of M. anisopliae did not affect the viability of the conidia. Allthe sub-cultures (1st, 20th, 40th) and passaged cultures (40th, 1st in vivo passage and40th, fifth in vivo passage) of M. anisopliae showed !90% germination on YPG agarwithin 24 h (data not shown). However, appressorium formation was significantlyaffected by repeated in vitro sub-culturing on PDA. The appressorium formation inthe first sub-culture was 96.990.46%. Repeated in vitro sub-culturing reduced it to86.695.12% and 76.793.36% for the 20th and 40th sub-cultures, respectively, whilethe passage of M. anisopliae 40th sub-culture in H. armigera resulted in increasedappressorium formation to 82.297.76 and 89.895.47% in the first and fifthpassages, respectively.
Production of cuticle-degrading enzymes
Sub-cultures of conidia, when used as an inoculum, showed significant variation inthe constitutive production of cuticle-degrading enzymes, viz., chitinase, chitosa-nase, CDA and protease in YPG medium (Table 1). The chitinase, chitosanase,CDA and protease activities of the first sub-culture decreased significantly (20"28%)after the 40th in vitro sub-culturing. The in vivo passage of the 40th sub-culture inH. armigera for 5 times resulted in a gradual increase in the cuticle-degradingenzyme activities. A similar trend for the production of cuticle-degrading enzymes,using different sub-cultures of conidia as the inoculum, was observed in a chitinmedium (Table 2). There was 19"29% decrease in the enzyme activities of the 40thsub-culture in chitin medium as compared to the activities of the first sub-culture.
Table 1. Effect of repeated in vitro sub-culturing and passage in insect host on theextracellular production of cuticle-degrading enzyme activities (mean9SD) in YPG mediumby Metarhizium anisopliae.
M. anisopliaesub-culture
Chitinase(!10#3 U/mL)
Chitosanase(!10#3 U /mL)
CDA(!10#3 U/mL)
Protease(U/mL)
Lipase(U/mL)
1st 0.51790.01d 1.7890.03d 1.2190.05e 0.9690.02c 0.27090.03a
20th 0.45390.01b 1.5590.03c 1.0690.04c 0.8190.03b 0.26390.02a
40th 0.37390.02a 1.3890.03a 0.9090.01a 0.7590.01a 0.23590.02a
40th (1st in vivo) 0.43390.03b 1.5290.04b 0.9790.01b 0.8490.02b 0.23590.02a
40th (5th in vivo) 0.47090.02c 1.6990.02c 1.1590.03d 0.9390.01c 0.26390.02a
Numbers denoted by same letter within the column are statistically not significant. 40th (1st in vivo) and40th (5th in vivo) " passaged cultures as described under Materials and methods. SD, standard deviation.
344 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010
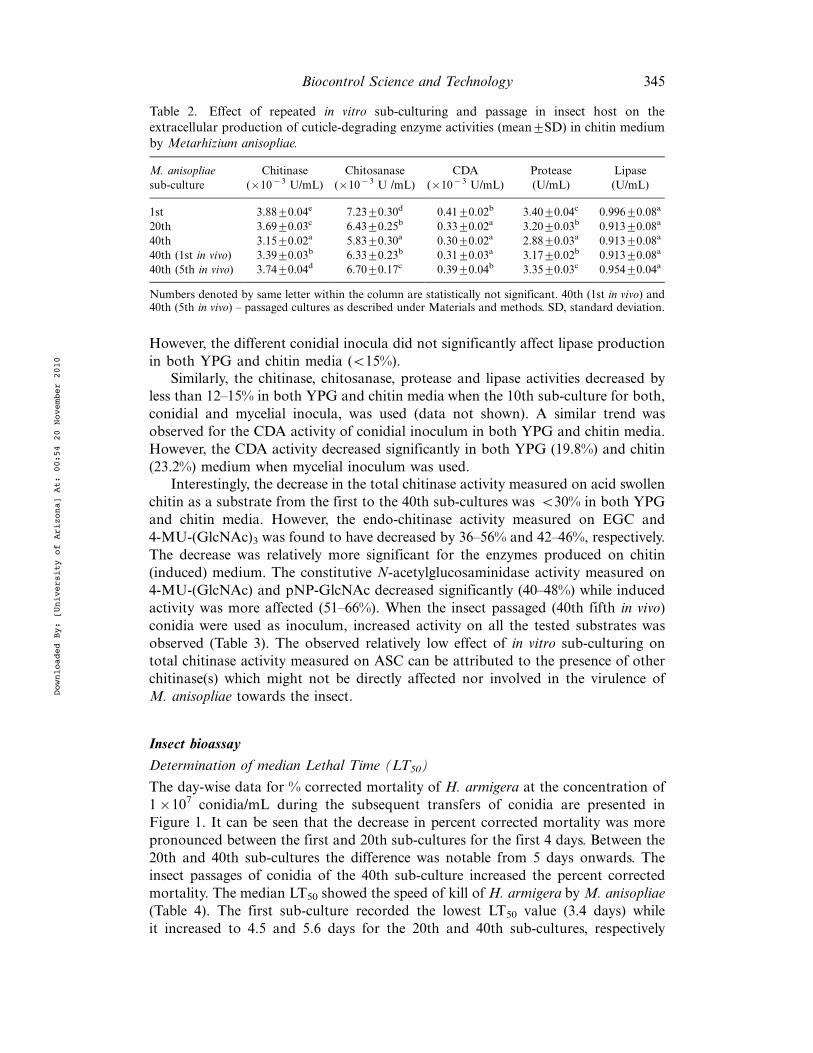
However, the different conidial inocula did not significantly affect lipase productionin both YPG and chitin media (B15%).
Similarly, the chitinase, chitosanase, protease and lipase activities decreased byless than 12"15% in both YPG and chitin media when the 10th sub-culture for both,conidial and mycelial inocula, was used (data not shown). A similar trend wasobserved for the CDA activity of conidial inoculum in both YPG and chitin media.However, the CDA activity decreased significantly in both YPG (19.8%) and chitin(23.2%) medium when mycelial inoculum was used.
Interestingly, the decrease in the total chitinase activity measured on acid swollenchitin as a substrate from the first to the 40th sub-cultures was B30% in both YPGand chitin media. However, the endo-chitinase activity measured on EGC and4-MU-(GlcNAc)3 was found to have decreased by 36"56% and 42"46%, respectively.The decrease was relatively more significant for the enzymes produced on chitin(induced) medium. The constitutive N-acetylglucosaminidase activity measured on4-MU-(GlcNAc) and pNP-GlcNAc decreased significantly (40"48%) while inducedactivity was more affected (51"66%). When the insect passaged (40th fifth in vivo)conidia were used as inoculum, increased activity on all the tested substrates wasobserved (Table 3). The observed relatively low effect of in vitro sub-culturing ontotal chitinase activity measured on ASC can be attributed to the presence of otherchitinase(s) which might not be directly affected nor involved in the virulence ofM. anisopliae towards the insect.
Insect bioassay
Determination of median Lethal Time (LT50)
The day-wise data for % corrected mortality of H. armigera at the concentration of1!107 conidia/mL during the subsequent transfers of conidia are presented inFigure 1. It can be seen that the decrease in percent corrected mortality was morepronounced between the first and 20th sub-cultures for the first 4 days. Between the20th and 40th sub-cultures the difference was notable from 5 days onwards. Theinsect passages of conidia of the 40th sub-culture increased the percent correctedmortality. The median LT50 showed the speed of kill of H. armigera by M. anisopliae(Table 4). The first sub-culture recorded the lowest LT50 value (3.4 days) whileit increased to 4.5 and 5.6 days for the 20th and 40th sub-cultures, respectively
Table 2. Effect of repeated in vitro sub-culturing and passage in insect host on theextracellular production of cuticle-degrading enzyme activities (mean9SD) in chitin mediumby Metarhizium anisopliae.
M. anisopliaesub-culture
Chitinase(!10#3 U/mL)
Chitosanase(!10#3 U /mL)
CDA(!10#3 U/mL)
Protease(U/mL)
Lipase(U/mL)
1st 3.8890.04e 7.2390.30d 0.4190.02b 3.4090.04c 0.99690.08a
20th 3.6990.03c 6.4390.25b 0.3390.02a 3.2090.03b 0.91390.08a
40th 3.1590.02a 5.8390.30a 0.3090.02a 2.8890.03a 0.91390.08a
40th (1st in vivo) 3.3990.03b 6.3390.23b 0.3190.03a 3.1790.02b 0.91390.08a
40th (5th in vivo) 3.7490.04d 6.7090.17c 0.3990.04b 3.3590.03c 0.95490.04a
Numbers denoted by same letter within the column are statistically not significant. 40th (1st in vivo) and40th (5th in vivo) " passaged cultures as described under Materials and methods. SD, standard deviation.
Biocontrol Science and Technology 345
Downloaded By: [University of Arizona] At: 00:54 20 November 2010
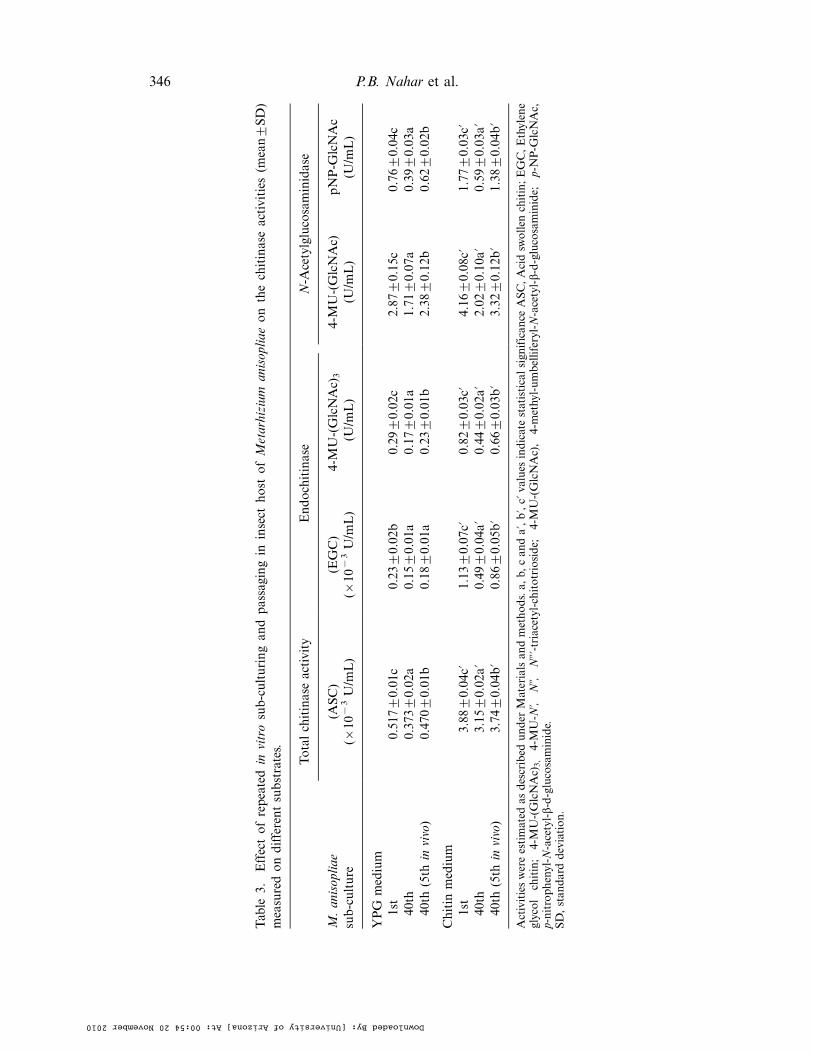
Table
3.Effectofrepeatedin
vitrosub-culturingan
dpassagingin
insect
host
ofMetarhizium
anisopliae
onthechitinaseactivities
(mean9SD)
measuredondifferentsubstrates.
Totalchitinaseactivity
Endochitinase
N-A
cetylglucosaminidase
M.anisopliae
sub-culture
(ASC)
(!10
#3U/m
L)
(EGC)
(!10
#3U/m
L)
4-MU-(GlcNAc)
3
(U/m
L)
4-MU-(GlcNAc)
(U/m
L)
pNP-G
lcNAc
(U/m
L)
YPG
medium
1st
0.5179
0.01c
0.23
90.02b
0.29
90.02c
2.87
90.15c
0.76
90.04c
40th
0.3739
0.02a
0.15
90.01a
0.17
90.01a
1.71
90.07a
0.39
90.03a
40th
(5th
invivo)
0.4709
0.01b
0.18
90.01a
0.23
90.01b
2.38
90.12b
0.62
90.02b
Chitin
medium
1st
3.88
90.04c?
1.13
90.07c?
0.82
90.03c?
4.16
90.08c?
1.77
90.03c?
40th
3.15
90.02a?
0.49
90.04a?
0.44
90.02a?
2.02
90.10a?
0.59
90.03a?
40th
(5th
invivo)
3.74
90.04b?
0.86
90.05b?
0.66
90.03b?
3.32
90.12b?
1.38
90.04b?
Activitieswereestimated
asdescribed
under
Materialsan
dmethods.a,
b,can
da?,b?,c?
values
indicatestatisticalsign
ifican
ceASC,Acidsw
ollen
chitin;EGC,Ethylene
glycol
chitin;
4-MU-(GlcNAc)
3,
4-MU-N
?,Nƒ,
Nƒ?-triacetyl-chitotrioside;
4-MU-(GlcNAc),
4-methyl-umbelliferyl-N-acetyl-b-d-glucosaminide;
p-NP-G
lcNAc,
p-nitrophenyl-N
-acetyl-b-d-glucosaminide.
SD,stan
darddeviation.
346 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010
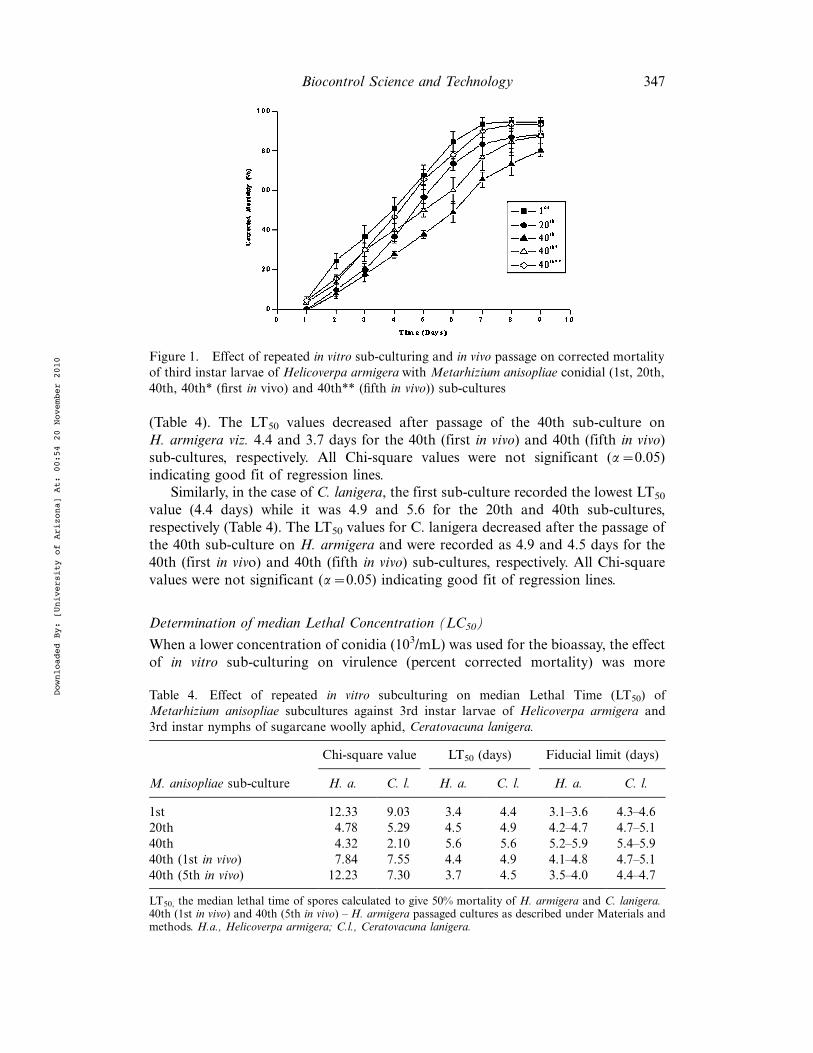
(Table 4). The LT50 values decreased after passage of the 40th sub-culture onH. armigera viz. 4.4 and 3.7 days for the 40th (first in vivo) and 40th (fifth in vivo)sub-cultures, respectively. All Chi-square values were not significant (a$0.05)indicating good fit of regression lines.
Similarly, in the case of C. lanigera, the first sub-culture recorded the lowest LT50
value (4.4 days) while it was 4.9 and 5.6 for the 20th and 40th sub-cultures,respectively (Table 4). The LT50 values for C. lanigera decreased after the passage ofthe 40th sub-culture on H. armigera and were recorded as 4.9 and 4.5 days for the40th (first in vivo) and 40th (fifth in vivo) sub-cultures, respectively. All Chi-squarevalues were not significant (a$0.05) indicating good fit of regression lines.
Determination of median Lethal Concentration (LC50)
When a lower concentration of conidia (103/mL) was used for the bioassay, the effectof in vitro sub-culturing on virulence (percent corrected mortality) was more
Figure 1. Effect of repeated in vitro sub-culturing and in vivo passage on corrected mortalityof third instar larvae of Helicoverpa armigera with Metarhizium anisopliae conidial (1st, 20th,40th, 40th* (first in vivo) and 40th** (fifth in vivo)) sub-cultures
Table 4. Effect of repeated in vitro subculturing on median Lethal Time (LT50) ofMetarhizium anisopliae subcultures against 3rd instar larvae of Helicoverpa armigera and3rd instar nymphs of sugarcane woolly aphid, Ceratovacuna lanigera.
Chi-square value LT50 (days) Fiducial limit (days)
M. anisopliae sub-culture H. a. C. l. H. a. C. l. H. a. C. l.
1st 12.33 9.03 3.4 4.4 3.1"3.6 4.3"4.620th 4.78 5.29 4.5 4.9 4.2"4.7 4.7"5.140th 4.32 2.10 5.6 5.6 5.2"5.9 5.4"5.940th (1st in vivo) 7.84 7.55 4.4 4.9 4.1"4.8 4.7"5.140th (5th in vivo) 12.23 7.30 3.7 4.5 3.5"4.0 4.4"4.7
LT50, the median lethal time of spores calculated to give 50% mortality of H. armigera and C. lanigera.40th (1st in vivo) and 40th (5th in vivo) " H. armigera passaged cultures as described under Materials andmethods. H.a., Helicoverpa armigera; C.l., Ceratovacuna lanigera.
Biocontrol Science and Technology 347
Downloaded By: [University of Arizona] At: 00:54 20 November 2010
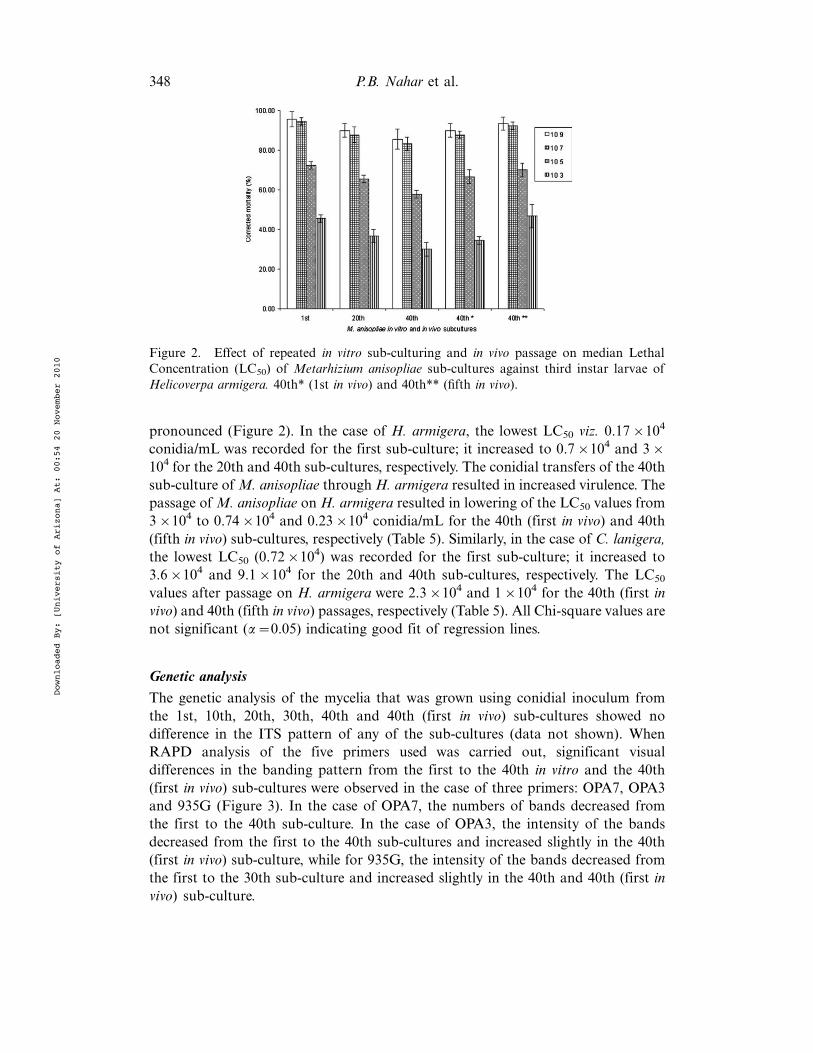
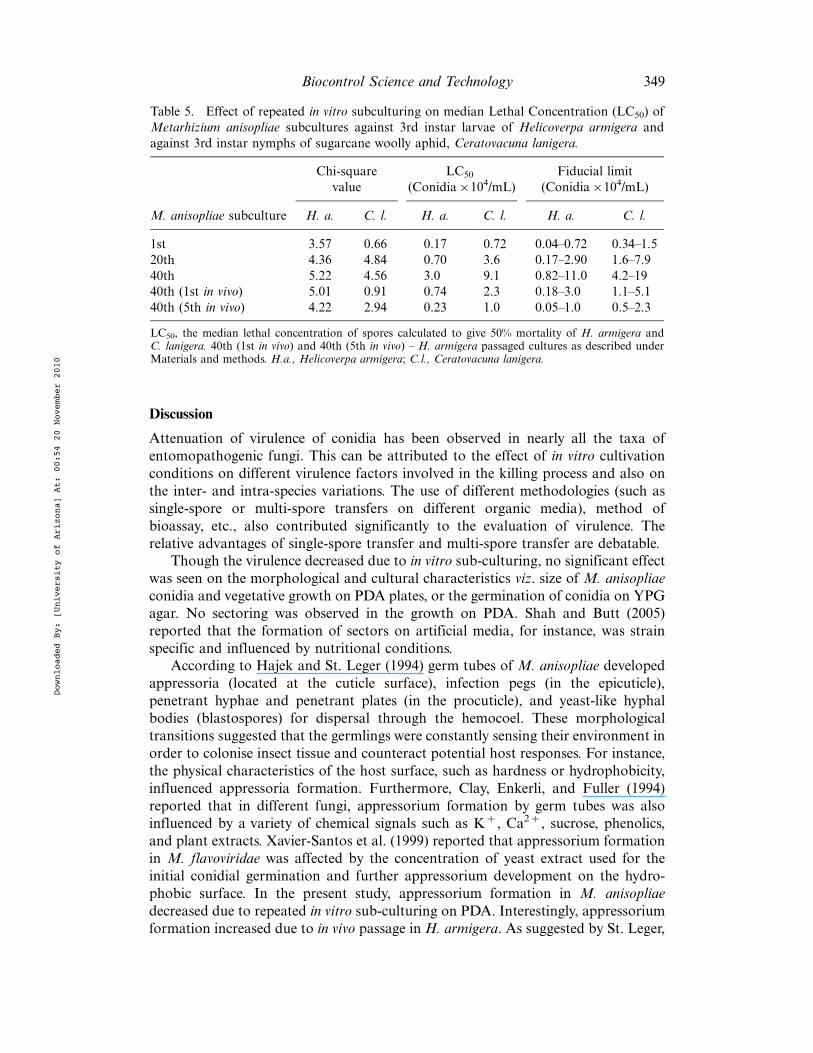
pronounced (Figure 2). In the case of H. armigera, the lowest LC50 viz. 0.17!104
conidia/mL was recorded for the first sub-culture; it increased to 0.7!104 and 3!104 for the 20th and 40th sub-cultures, respectively. The conidial transfers of the 40thsub-culture ofM. anisopliae throughH. armigera resulted in increased virulence. Thepassage of M. anisopliae on H. armigera resulted in lowering of the LC50 values from3!104 to 0.74!104 and 0.23!104 conidia/mL for the 40th (first in vivo) and 40th(fifth in vivo) sub-cultures, respectively (Table 5). Similarly, in the case of C. lanigera,the lowest LC50 (0.72!104) was recorded for the first sub-culture; it increased to3.6!104 and 9.1!104 for the 20th and 40th sub-cultures, respectively. The LC50
values after passage on H. armigera were 2.3!104 and 1!104 for the 40th (first invivo) and 40th (fifth in vivo) passages, respectively (Table 5). All Chi-square values arenot significant (a$0.05) indicating good fit of regression lines.
Genetic analysis
The genetic analysis of the mycelia that was grown using conidial inoculum fromthe 1st, 10th, 20th, 30th, 40th and 40th (first in vivo) sub-cultures showed nodifference in the ITS pattern of any of the sub-cultures (data not shown). WhenRAPD analysis of the five primers used was carried out, significant visualdifferences in the banding pattern from the first to the 40th in vitro and the 40th(first in vivo) sub-cultures were observed in the case of three primers: OPA7, OPA3and 935G (Figure 3). In the case of OPA7, the numbers of bands decreased fromthe first to the 40th sub-culture. In the case of OPA3, the intensity of the bandsdecreased from the first to the 40th sub-cultures and increased slightly in the 40th(first in vivo) sub-culture, while for 935G, the intensity of the bands decreased fromthe first to the 30th sub-culture and increased slightly in the 40th and 40th (first invivo) sub-culture.
Figure 2. Effect of repeated in vitro sub-culturing and in vivo passage on median LethalConcentration (LC50) of Metarhizium anisopliae sub-cultures against third instar larvae ofHelicoverpa armigera. 40th* (1st in vivo) and 40th** (fifth in vivo).
348 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010
Discussion
Attenuation of virulence of conidia has been observed in nearly all the taxa ofentomopathogenic fungi. This can be attributed to the effect of in vitro cultivationconditions on different virulence factors involved in the killing process and also onthe inter- and intra-species variations. The use of different methodologies (such assingle-spore or multi-spore transfers on different organic media), method ofbioassay, etc., also contributed significantly to the evaluation of virulence. Therelative advantages of single-spore transfer and multi-spore transfer are debatable.
Though the virulence decreased due to in vitro sub-culturing, no significant effectwas seen on the morphological and cultural characteristics viz. size of M. anisopliaeconidia and vegetative growth on PDA plates, or the germination of conidia on YPGagar. No sectoring was observed in the growth on PDA. Shah and Butt (2005)reported that the formation of sectors on artificial media, for instance, was strainspecific and influenced by nutritional conditions.
According to Hajek and St. Leger (1994) germ tubes of M. anisopliae developedappressoria (located at the cuticle surface), infection pegs (in the epicuticle),penetrant hyphae and penetrant plates (in the procuticle), and yeast-like hyphalbodies (blastospores) for dispersal through the hemocoel. These morphologicaltransitions suggested that the germlings were constantly sensing their environment inorder to colonise insect tissue and counteract potential host responses. For instance,the physical characteristics of the host surface, such as hardness or hydrophobicity,influenced appressoria formation. Furthermore, Clay, Enkerli, and Fuller (1994)reported that in different fungi, appressorium formation by germ tubes was alsoinfluenced by a variety of chemical signals such as K%, Ca2%, sucrose, phenolics,and plant extracts. Xavier-Santos et al. (1999) reported that appressorium formationin M. flavoviridae was affected by the concentration of yeast extract used for theinitial conidial germination and further appressorium development on the hydro-phobic surface. In the present study, appressorium formation in M. anisopliaedecreased due to repeated in vitro sub-culturing on PDA. Interestingly, appressoriumformation increased due to in vivo passage in H. armigera. As suggested by St. Leger,
Table 5. Effect of repeated in vitro subculturing on median Lethal Concentration (LC50) ofMetarhizium anisopliae subcultures against 3rd instar larvae of Helicoverpa armigera andagainst 3rd instar nymphs of sugarcane woolly aphid, Ceratovacuna lanigera.
Chi-squarevalue
LC50
(Conidia!104/mL)Fiducial limit
(Conidia!104/mL)
M. anisopliae subculture H. a. C. l. H. a. C. l. H. a. C. l.
1st 3.57 0.66 0.17 0.72 0.04"0.72 0.34"1.520th 4.36 4.84 0.70 3.6 0.17"2.90 1.6"7.940th 5.22 4.56 3.0 9.1 0.82"11.0 4.2"1940th (1st in vivo) 5.01 0.91 0.74 2.3 0.18"3.0 1.1"5.140th (5th in vivo) 4.22 2.94 0.23 1.0 0.05"1.0 0.5"2.3
LC50, the median lethal concentration of spores calculated to give 50% mortality of H. armigera andC. lanigera. 40th (1st in vivo) and 40th (5th in vivo) " H. armigera passaged cultures as described underMaterials and methods. H.a., Helicoverpa armigera; C.l., Ceratovacuna lanigera.
Biocontrol Science and Technology 349
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

Butt, Goettel, Staples, and Roberts (1989), a nutrient gradient associated with thegerm tube contact with a rigid insect surface may be the main stimulus triggeringappressorium formation. Vestergaard, Butt, Bresciani, Gillespie, and Eilenberg(1999) observed that appressorium formation in M. anisopliae was one of theimportant factors contributing to the mortality of western flower thrip, Frankliniellaoccidentalis.
The effect of repeated transfers on the production of virulence factors, viz.cuticle-degrading enzymes and toxins, in the fungus"insect interaction was alsosignificant. Several studies utilising B. bassiana, N. rileyi, and other fungi suggestedthat virulence was correlated, at least in part, with chitinase activity (El-Sayed,Coudron, and Ignoffo 1989; El-Sayed, Ignoffo, Leathers, and Gupta 1993; St. Leger,Joshi, Bidochka, Rizzo, and Roberts 1996). The studies by Binod, Sukumaran,Shirke, Rajput, and Pandey (2007) showed that the culture filtrate containingchitinase from T. harzianum was capable of negatively affecting growth andmetamorphosis of Heliothis larvae, while St. Leger, Cooper, and Charnley (1986b)reported that extracellular proteases and chitinases produced by M. anisopliae,M. flavoviride and B. bassiana were important for insect penetration. Nahar et al.(2004) suggested the importance of constitutively produced chitin deacetylase andchitosanase in the virulence of M. anisopliae towards H. armigera.
In a number of studies with plant, animal, and fungal pathogens, the roles ofconstitutive as well as induced enzymes in growth and virulence have been reported(Inbar and Chet 1995; Weber and Pitt 1997; Kapat, Zimand, and Elad 1998;Sebbane, Jarrett, Linkenhoker, and Hinnebusch 2004). Inbar and Chet (1995)reported induction of specific chitinases in T. harzianum during mycoparasitism of
Figure 3. Representative agarose gel electrophoresis of RAPD-PCR products for in vitroconidial transfers (1st, 10th, 20th, 30th and 40th) and in vivo passage of 40th sub-culture(40th first in vivo) of M. anisopliae. Lanes 1"6, Primer 947G; Lanes 7"12, Primer OPA13;Lanes 13"18, Primer OPA07; Lane 19, DNA ladder; Lanes 20"25, Primer OPA03; and Lanes26"31, Primer 935G.
350 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

Sclerotium rolfsii. Only live mycelium of S. rolfsii was found to induce enzymeproduction. In our study, we observed a gradual decrease in the constitutive (YPGmedium) and induced (chitin medium) activities of chitinase, chitosanase, CDA andprotease with repeated conidial transfers on artificial media. These activities wereseen to increase after insect passage (Tables 1 and 2). Similarly, the conidia producedon insect cadavers showed higher levels of Pr1 (an important cuticle degradingprotease bound to the spore wall) than the conidia produced in vitro (St. Leger,Goettel, Roberts, and Staples 1991; Shah et al. 2005).
It was reported by Morrow et al. (1989) that in N. rileyi repeated vegetative(mycelial) transfer did not affect the virulence to A. gemmatalis. In the case ofM. anisopliae, when mycelial inoculum from the 10th serial mycelial transfer wasused, the decrease in the cuticle-degrading enzyme activities such as chitinase,chitosanase, protease and lipase was similar to the activities produced by inoculatingconidia of the 10th sub-culture. However, the chitin deacetylase activity in both YPGand chitin media was found to decrease significantly as compared to the 40thconidial sub-culture used as an inoculum. This can be attributed to the role of chitindeacetylase in the penetration of insect cuticle as well as for self-defense, as suggestedby Nahar et al. (2004). Their investigations suggested that constitutive chitindeacetylase from M. anisopliae, in the absence of induced chitinase, convertedcuticular chitin to chitosan and thus played an important role in cuticle softeningand pathogenesis. Chitin deacetylase deacetylates the cell wall chitin of thepenetrating fungal hypha for protection against plant endochitinases. Insectsproduce chitinase to degrade old cuticle during moulting, which can also act onfungal cell wall chitin. To combat insect chitinases, entomopathogenic fungi can alsoadopt this as a defense mechanism (Nahar et al. 2004).
Attenuated conidia may germinate and infect their hosts marginally slower thannon-attenuated strains. This may be partly due to the attenuated conidia lacking theright set of enzymes to facilitate host penetration. For example,M. anisopliae conidiaproduced in vitro had lower levels of Pr1 (an important cuticle-degrading protease,bound to the spore wall) than conidia produced on insect cadavers (St. Leger et al.1991; Shah et al. 2005). Furthermore, attenuated culture may not differentiate intothe form adapted for the hemocoel or produce the metabolites that normallysuppress the host defense. Wang, Skrobek, and Butt (2003) reported that attenuatedcultures did not produce destruxins, the secondary metabolites harmful to the host’simmune system. Butt et al. (2006) reported that Pr1 and destruxins could be used asmarkers to monitor the virulence of entomopathogenic fungi during mass produc-tion. The present investigations suggest that cuticle-degrading enzymes such aschitinase, CDA, chitosanase and protease could be used as markers to monitor thevirulence of entomopathogenic fungi during mass production.
Wang, Hu, and St. Leger (2005) reported that, in M. anisopliae, the differentialtranscription regulation of gene expression under saprophytic phase, in the presenceof host cuticle or hemolymph, was just a physiological adaptation in theenvironment. Weber and Pitt (1997) reported the constitutive presence of two formsof acid phosphatases in B. cinerea with strikingly different properties. In the case ofentomopathogens like Metarhizium species, which can also grow saprophytically, theconstitutive activities of the 40th sub-culture may be mainly responsible for thegrowth. Therefore, in the present case, decrease in the levels of enzyme activities
Biocontrol Science and Technology 351
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

during in vitro repeated transfers from the first to the 40th sub-culture can beattributed to the decrease in the activities which are involved in some other functions,such as virulence. This could be one of the reasons for the increase in the activitiesafter insect passages. Furthermore, specific substrates used to measure endo-chitinase and N-acetylglucosaminidase activities distinctly indicated the role ofsome of the specific chitinolytic enzymes in virulence and/or growth (Table 3). It willbe interesting to find out the presence of different isozymes, if any, involved in thegrowth and virulence of our strain M. anisopliae towards H. armigera.
The virulence of Paecilomyces farinosus towards the English grain aphid, Sitobionavenae (F.) increased after host passage as well as when the fungus was grown in themedium containing S. avenae cuticle as the only nutrient source (Hayden et al. 1992).In both methods, after three selective sub-cultures of P. farinosus, LT50 decreasedfrom 11 to 5 days. The changes in the LT50 of the present strain M. anisopliae afterpassage through H. armigera were in accordance with the earlier reports (Table 4).There are several reports that passaging did not enhance virulence, presumablybecause the strains were at their full insecticidal potential or because the strainschanged irreversibly physiologically (Butt et al. 2006). In the present study, the in vivopassage (5 times) of the M. anisopiae through H. armigera increased the levels ofvirulence factors such as cuticle-degrading enzyme activities as well as virulenceagainst H. armigera and C. lanigera. The LC50 values being higher for C. lanigerathan for H. armigera can be attributed to the host specificity, as well as to the waxyfluffy coating of the insect or method of bioassay (Table 5).
The effectiveness of the biocontrol preparation depends on the number of conidiarequired to kill individual larva. As seen from Table 5, the LC50 value of the 40thsub-culture increased as compared to that of the first sub-culture. This will affect thefield performance of the 40th sub-culture. The LC50 can be decreased by passagingM. anisopliae through H. armigera. Alternatively, the same level of efficacy can beobtained by applying !20% conidial inoculum under field conditions. However, theselection of either of the above strategies would greatly depend on the cost factor.
Genetic analysis of M. anisopliae in vitro and in vivo sub-cultures showedcorrelation with the biochemical data. In RAPD analysis for three primers (OPA7,OPA3 and 935 G) significant visual differences were observed in the banding patternfrom the first to the 40th in vitro and the 40th in vivo passage ofM. anisopliae (Figure3). The NCBI blast results of OPA7 showed significant alignment with serinepeptidase from Aspergillus fumigatus. While OPA 3 primer was reported to be usedfor the amplification of lysophospholipase, it was also used for the amplification ofthe calmodulin gene. The biochemical processes controlled by the Ca-calmodulinsignalling system were involved in the induction of appressorium in Colletotrichumtrifolii pathogenic on red pepper (Warwar and Dickman 1996). These observationsindicated the phenotypic changes with respect to the virulence of sub-culturedconidia. However, further experimentation is necessary to identify explicitly theeffect of sub-culturing on the virulence factors. Understanding of the biochemicalmechanism of fungus interaction with respect to the virulence and growth of theorganism on artificial media is necessary for the cost-effective commercial exploita-tion of the entomopathogens.
352 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

Acknowledgements
The authors are grateful to the Indo-Swiss Collaboration in Biotechnology (ISCB) programmefor financial support.
References
Abbott, W.S. (1925), ‘‘A Method for Computing the Effectiveness of an Insecticide,’’ Journal ofEconomic Entomology, 18, 265"267.
Binod, P., Sukumaran, R.K., Shirke, S.V., Rajput, J.C., and Pandey, A. (2007), ‘‘Evaluation ofFungal Culture Filtrate Containing Chitinase as a Biocontrol Agent Against Helicoverpaarmigera,’’ Journal of Applied Microbiology, 103, 1845"1852.
Brownbridge, M., Costa, S., and Jaronski, S.T. (2001), ‘‘Effects of In Vitro Passage ofBeauveria bassiana on Virulence to Bemisia argentifolii,’’ Journal of Invertebrate Pathology,77, 280"283.
Butt, T.M., Wang, C.S., Shah, F.A., and Hall, R. (2006), ‘‘Degeneration of EntomogenousFungi,’’ in An Ecological and Societal Approach to Biological Control, eds J. Eilenberg andH.M.T. Hokkanen, Amsterdam: Springer, pp. 213"226.
Chavan, S., Ghormade, V., Nahar, P., and Deshpande, M.V. (2005), ‘‘EntomopathogenicFungi: A Valuable Tool to Fight against Insect Pests,’’ in Plant Protection in NewMillennium, eds. A.V. Gadewar and B.P. Singh, New Delhi: Satish Serial Publishing House,pp. 227"243,.
Clay, R.P., Enkerli, J., and Fuller, M.S. (1994), ‘‘Induction and Formation of Cochliobolussativus Appressoria,’’ Protoplasma, 178, 34"47.
El-Sayed, G.N., Coudron, T.A., and Ignoffo, C.M. (1989), ‘‘Chitinolytic Activity andVirulence Associated with Native and Mutant Isolates of an Entomopathogenic FungusNomuraea rileyi,’’ Journal of Invertebrate Pathology, 54, 394"403.
El-Sayed, G.N., Ignoffo, C.M., Leathers, T.D., and Gupta, S.C. (1993), ‘‘Cuticular and Non-Cuticular Substrate Influence on Expression of Cuticle-Degrading Enzymes from Conidiaof an Entomopathogenic Fungus, Nomuraea rileyi,’’ Mycopathologia, 122, 79"87.
Finney, D.J. (1981), Probit Analysis (3rd ed., Pub. 5), New Delhi: Chand and Co. Ltd, pp. 19"76.
Fransen, J.J., Winkelman, K., and Van Lenteren, J.C. (1987), ‘‘The Differential Mortality atVarious Life Stages of the Greenhouse Whitefly, Trialeurodes vaporariorum (Homoptera:Aleyrodidae), by Infection with the Fungus Aschersonia aleyrodis (Deuteromycotina:Coelomycetes),’’ Journal of Invertebrate Pathology, 50, 158"165.
Ghormade, V.S., Lachake, S.A., and Deshpande, M.V. (2000), ‘‘Dimorphism in Benjaminiellapoitrasii: Involvement of Intracellular Endochitinase and N-Acetylglucosaminidase Activ-ities in the Yeast-Mycelium Transitions,’’ Folia Microbiology, 45, 231"238.
Hall, R.A. (1980), ‘‘Effect of Repeated Sub-Culturing on Agar and Passaging through anInsect Host on Pathogenicity, Morphology, and Growth Rate of Verticillium lecanii,’’Journal of Invertebrate Pathology, 36, 216"222.
Hajek, A.E., and St. Leger, R.J. (1994), ‘‘Interactions between Fungal Pathogens and InsectHosts,’’ Annual Review of Entomology, 39, 293"322.
Hayden, T.P., Bidochka, M.J., and Khachatourians, G.G. (1992), ‘‘Entomopathogenicity ofSeveral Fungi toward the English Grain Aphid (Homoptera: Aphididae) and Enhancementof Virulence with Host Passage of Paecilomyces farinosus,’’ Journal of EconomicEntomology, 85, 58"64.
Ignoffo, C.M., Futtler, B., Marston, N.L., Hostetter, D.L., and Dickerson, W.A. (1975),‘‘Seasonal Incidence of the Entomopathogenic Fungus, Spicaria rileyi Associated withNoctuid Pests of Soybeans,’’ Journal of Invertebrate Pathology, 25, 135"137.
Biocontrol Science and Technology 353
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

Inbar, J., and Chet, I. (1995), ‘‘The Role of Recognition in the Induction of Specific Chitinasesduring Mycoparasitism by Trichoderma harzianum,’’ Microbiology, 141, 2823"2829.
Kapat, A., Zimand, G., and Elad, Y. (1998), ‘‘Biosynthesis of Pathogenecity HydrolyticEnzymes by Botrytis cinerea during Infection of Bean Leaves and In Vitro,’’ MycologicalResearch, 102, 1017"1024.
Krieger de Moraes, C., Schrank, A., and Vainstein, M.H. (2003), ‘‘Regulation of ExtracellularChitinases and Proteases in the Entomopathogen and Acaricide Metarhizium anisopliae,’’Current Microbiology, 46, 205"210.
Meekes, E.T.M., Fransen, J.J., and van Lenteren, J.C. (2002), ‘‘Pathogenicity of Aschersoniaspp. against Whiteflies Bemisia argentifolii and Trialeurodes vaporariorum,’’ Journal ofInvertebrate Pathology, 81, 1"11.
Morrow, B.J., Boucias, D.G., and Heath, M.A. (1989), ‘‘Loss of Virulence in an Isolate of anEntomopathogenic Fungus, Nomuraea rileyi, after Serial In Vitro Passage,’’ Journal ofEconomic Entomology, 82, 404"407.
Nahar, P., Kulye, M., Yadav, P., Hassani, M., Tuor, U., Keller, S., and Deshpande, M.V. (2003),‘‘Comparative Evaluation of Indigenous Fungal Isolates, Metarhizium anisopliae M34412,Beauveria bassiana B3301 and Nomuraea rileyi N812 for the Control of Helicoverpaarmigera (Hub) on Chickpea,’’ Journal of Mycology and Plant Pathology, 33, 372"377.
Nahar, P., Ghormade, V., and Deshpande, M.V. (2004), ‘‘The Extracellular ConstitutiveProduction of Chitin Deacetylase in Metarhizium anisopliae: Possible Edge to Entomo-pathogenic Fungi in the Biological Control of Insect Pests,’’ Journal of InvertebratePathology, 85, 80"88.
Panse, V.G., and Sukhatme, D.V. (1989), Statistical Methods for Horticultural Workers(4th ed.), New Delhi: Japan Art Press.
Patil, R.S., Ghormade, V., and Deshpande, M.V. (2000), ‘‘Chitinolytic Enzymes: Anexploration,’’ Enzyme Microbial Technolog,y, 26, 473"483.
Pignede, G., Wang, H., Fudalej, F., Gaillardin, C., Seman, M., and Nicaud, J.M. (2000),‘‘Characterization of an Extracellular Lipase Encoded by LIP2 in Yarrowia lipolytica,’’Journal of Bacteriology, 182, 2802"2810.
Schaerffenberg, B. (1964), ‘‘Biological and Environmental Conditions for the Development ofMycoses Caused by Beauveria and Metarhizium,’’ Journal of Insect Pathology, 6, 8"20.
Sebbane, F., Jarrett, C.O., Linkenhoker, J.R., and Hinnebusch, B.J. (2004), ‘‘Evaluation of theRole of Constitutive Isocitrate Lyase Activity in Yersinia pestis Infection of the Flea Vectorand Mammalian Host,’’ Infection Immunity, 72, 7334"7337.
Shah, F.A., and Butt, T.M. (2005), ‘‘Influence of Nutrition on the Production and Physiologyof Sectors Produced by the Insect Pathogenic Fungus Metarhizium anisopliae,’’ FEMSMicrobiology Letters, 250, 201"207.
Shah, F.A., Wang, C.S., and Butt, T.M. (2005), ‘‘Nutrition Influences Growth and Virulenceof the Insect-Pathogenic FungusMetarhizium anisopliae,’’ FEMSMicrobiology Letters, 251,259"266.
Shakeri, J., and Foster, H.A. (2007), ‘‘Proteolytic Activity and Antibiotic Production byTrichoderma harzianum in Relation to Pathogenicity to Insects,’’ Enzyme and MicrobialTechnology, 40, 961"968.
St. Leger, R.J., Cooper, R.M., and Charnley, A.K. (1986a), ‘‘Cuticle-Degrading Enzymes ofEntomopathogenic Fungi: Regulation of Production of Chitinolytic Enzymes,’’ Journal ofGeneral Microbiology, 132, 1509"1517.
St. Leger, R.J., Cooper, R.M., and Charnley, A.K. (1986b), ‘‘Cuticle-Degrading Enzymes ofEntomopathogenic Fungi: Cuticle-Degradation In Vitro by Enzymes from Entomopatho-gens,’’ Journal of Invertebrate Pathology, 47, 167"177.
St. Leger, R.J., Butt, T.M., Goettel, M.S., Staples, R.C., and Roberts, D.W. (1989),‘‘Production of Appressoria by the Entomopathogenic Fungus Metarhizium anisopliae,’’Experimental Mycology, 13, 274"288.
354 P.B. Nahar et al.
Downloaded By: [University of Arizona] At: 00:54 20 November 2010

St. Leger, R.J., Goettel, M., Roberts, D.W., and Staples, R.C. (1991), ‘‘Prepenetration Eventsduring Infection of Host Cuticle by Metarhizium anisopliae,’’ Journal of InvertebratePathology, 58, 168"179.
St. Leger, R.J., Joshi, L., Bidochka, M.J., Rizzo, N.W., and Roberts, D.W. (1996),‘‘Characterization and Ultrastructural Localization of Chitinases from Metarhiziumanisopliae,M. flavoviride, and Beauveria bassiana during Fungal Invasion of Host (Manducasexta) Cuticle,’’ Applied and Environmental Microbiology, 62, 907"912.
Throne, J.E., Weaver, D.K., Chew, V., and Baker, J.E. (1995), ‘‘Probit Analysis of CorrelatedData: Multiple Observations over Time at one Concentration,’’ Journal of EconomicEntomology, 88, 1510"1512.
Vandenberg, J.D., and Cantone, F.A. (2004), ‘‘Effect of Serial Transfer of Three Strains ofPaecilomyces fumosoroseus on Growth In Vitro, Virulence, and Host Specificity,’’ Journal ofInvertebrate Pathology, 85, 40"45.
Vestergaard, S., Butt, T.M., Bresciani, J., Gillespie, A.T., and Eilenberg, J. (1999), ‘‘Light andElectron Microscopy Studies of the Infection of the Western Flower Thrips Frankliniellaoccidentalis (Thysanoptera: Thripidae) by the Entomopathogenic Fungus Metarhiziumanisopliae,’’ Journal of Invertebrate Pathology, 73, 25"33.
Vyas, P.R., and Deshpande, M.V. (1989), ‘‘Chitinase Production by Myrothecium verrucariaand its Significance for Fungal Mycelia Degradation,’’ Journal of General and AppliedMicrobiology, 35, 343"350.
Wang, C., Skrobek, A., and Butt, T.M. (2003), ‘‘Concurrence of Losing a Chromosome andthe Ability to Produce Destruxins in a Mutant of Metarhizium anisopliae,’’ FEMSMicrobiology Letters, 226, 373"378.
Wang, C., Hu, G., and St. Leger, R.J. (2005), ‘‘Differential Gene Expression by Metarhiziumanisopliae Growing in Root Exudates and Host (Manduca sexta) Cuticle or HemolymphReveals Mechanisms of Physiological Adaptation,’’ Fungal Genetics and Biology, 42, 704"718.
Warwar, V., and Dickman, M.B. (1996), ‘‘Effects of Calcium and Calmodulin on SporeGermination and Appressorium Development in Colletotrichum trifolii,’’ Applied andEnvironmental Microbiology, 62, 74"79.
Weber, R.W.S., and Pitt, D. (1997), ‘‘Purification, Characterization and Exit Routes of TwoAcid Phosphatases Secreted by Botrytis cinerea,’’ Mycological Research, 101, 1431"1439.
Xavier-Santos, S., Magalhaes, B., Elza, A., and Luna-Alves., L. (1999), ‘‘Differentiation of theEntomopathogenic Fungus Metarhizium flavoviride (Hyphomycetes),’’ Revista de Micro-biologia, 30, 47"51.
Biocontrol Science and Technology 355
Downloaded By: [University of Arizona] At: 00:54 20 November 2010





























