Embed Size (px)
Citation preview

1 23
��������������� ����� ����� ��������� � ������������������������������ ���!"#��$����������!�!
���������� ������������������������ �������������������������������������� ������ ������������� ����������
��������� ��������������� ���������������������������������

1 23
Your article is protected by copyright andall rights are held exclusively by Springer-Verlag Berlin Heidelberg. This e-offprint isfor personal use only and shall not be self-archived in electronic repositories. If you wishto self-archive your article, please use theaccepted manuscript version for posting onyour own website. You may further depositthe accepted manuscript version in anyrepository, provided it is only made publiclyavailable 12 months after official publicationor later and provided acknowledgement isgiven to the original source of publicationand a link is inserted to the published articleon Springer's website. The link must beaccompanied by the following text: "The finalpublication is available at link.springer.com”.

1 3
PlantaDOI 10.1007/s00425-013-1997-7
ORIGINAL ARTICLE
Roles of ethylene and jasmonic acid in systemic induced defense in tomato (Solanum lycopersicum) against Helicoverpa zea
Donglan Tian · Michelle Peiffer · Consuelo M. De Moraes · Gary W. Felton
Received: 28 August 2013 / Accepted: 4 November 2013 © Springer-Verlag Berlin Heidelberg 2013
AbbreviationsNr Never ripe mutantdef1 Defenseless 1 mutantRU Rutgers wild-type plants of Nr mutantCM Castlemart, wild-type plants of def1 mutantMeJA Methyl jasmonatePIN2 Protease inhibitor 2ERF1 Ethylene response factorPR1 Pathogenesis related genePPOF Polyphenol oxidaseET EthyleneSA Salicylic acid
Introduction
Biotic and abiotic stresses negatively impact plant health and fitness. Plants respond to the biotic stress imposed by insect herbivores and pathogens by inducing systemic resistance pathways. Plant hormones play important roles in regulating this defensive process (Howe and Jander 2008). To date, jasmonic acid (JA), salicylic acid (SA), eth-ylene (ET) and abscisic acid have been intensively investi-gated in terms of plant defense against pathogen infection or insect herbivory (Abe et al. 2011; Fan et al. 2009; Leon-Reyes et al. 2009). Other plant hormones, such as brassi-nosteroids, gibberellins and auxins have also been reported as part of the defense-signaling network (Navarro 2006; Yasuda et al. 2003; Zsogon et al. 2008).
Jasmonic acid is a naturally occurring phytohormone that occurs ubiquitously in plant species and functions in regulation of seed germination, root growth, pollen devel-opment and fruit ripening (Zhang et al. 2011). In addi-tion to its role in plant growth development, JA and its precursors perform an important function in regulating
Abstract Inducible defenses that provide enhanced resistance to insect attack are nearly universal in plants. The defense-signaling cascade is mediated by the synthe-sis, movement, and perception of jasmonate (JA) and the interaction of this signaling molecule with other plant hor-mones and messengers. To explore how the interaction of JA and ethylene influences induced defenses, we employed the never-ripe (Nr) tomato mutant, which exhibits a partial block in ethylene perception, and the defenseless (def1) mutant, which is deficient in JA biosynthesis. The defense gene proteinase inhibitor (PIN2) was used as marker to compare plant responses. The Nr mutant showed a nor-mal wounding response with PIN2 induction, but the def1 mutant did not. As expected, methyl JA (MeJA) treatment restored the normal wound response in the def1 mutant. Exogenous application of MeJA increased resistance to Helicoverpa zea, induced defense gene expression, and increased glandular trichome density on systemic leaves. Exogenous application of ethephon, which penetrates tis-sues and decomposes to ethylene, resulted in increased H. zea growth and interfered with the wounding response. Eth-ephon treatment also increased salicylic acid in systemic leaves. These results indicate that while JA plays the main role in systemic induced defense, ethylene acts antagonisti-cally in this system to regulate systemic defense.
Keywords Methyl jasmonate · Ethephon · Induced systemic defense · Insect herbivore · Trichomes · Proteinase inhibitor · Hormones · Jasmonic acid · Salicylic acid
D. Tian · M. Peiffer · C. M. De Moraes · G. W. Felton (*) Center for Chemical Ecology, Department of Entomology, Penn State University, University Park, PA 16802, USAe-mail: [email protected]
Author's personal copy

Planta
1 3
host defensive responses. The regulation of plant defense by JA has been studied by observing the gene expression patterns following exogenous application of jasmonates or through the use of loss of function or gain of function mutants (Browse 2009; Howe 2004). Proteinase inhibitors (PINs) and polyphenol oxidases (PPO) are two of the best studied groups of JA-regulated defense proteins in veg-etative tissues. Wound-induced proteinase inhibitors have been shown to enhance the plant’s resistance to insects by inhibiting the proteolytic enzymes of the attacking insects (Koiwa et al. 1998). In some instances, JA partially regulates trichome production, which functions in plant defense (Halitschke et al. 2008; Li et al. 2004; Traw and Bergelson 2003; Tian et al. 2012b). In tomato, application of methyl jasmonate (MeJA) increases densities of glandu-lar trichomes on new leaves (Boughton et al. 2005). The inducibility of trichome density is ecologically significant because increased trichome density can negatively influ-ence herbivore populations (Agrawal 1999; Horgan et al. 2009; Kessler et al. 2011).
In addition to jasmonic acid, additional plant hor-mones are known to regulate plant defenses. Ethylene was the first identified gaseous hormone and it is involved in many plant physiological process throughout plant development including, seed germination, flowering, fruit ripening, organ senescence and response to stress (Zhang et al. 2010). Previous studies showed that the key compo-nents of ethylene biosynthesis, 1-aminocyclopropane—1 carboxylic acid synthesis and 1-aminocyclopropane-car-boxylic acid oxidase are critical for ethylene biosynthe-sis (An et al. 2006; Yang and Hoffman 1984). Regulation of the ethylene pathway is important in mediating plant development and stress response (Diaz et al. 2002; Man-telin et al. 2009; Zhang et al. 2011). Ethylene accumu-lation during pathogen infection up-regulates the expres-sion of many genes in plants (Diaz et al. 2002). Ethylene response factors, which represent the second largest tran-scription factor family in plants, have been shown to inte-grate signals from different plant hormone pathways and play important roles in stress response (Onate-Sanchez et al. 2007).
Hormone cross talk is a process where different hor-mone signaling pathways interact antagonistically or syn-ergistically to regulate responses to complex and changing environments (Verhage et al. 2010). There is ample evi-dence for cross talk among SA, JA and ET signaling path-ways in plant immunity (Leon-Reyes et al. 2009). Cross talk likely minimizes fitness costs and creates a flexible signaling network that allows the plant to fine-tune its defense response to the appropriate invaders (Bostock 2005; Thaler et al. 2012; Pieterse and Dicke 2007). In some instances, JA may collaborate with the SA response pathway to trigger an indirect defense against an insect
herbivore as observed in Arabidopsis (van Poecke and Dicke 2002). Walling (2000) found that phloem-feeding insects may induce both SA- and JA-dependent pathways. Simultaneous expression of SA and JA pathways was observed in the Mi-gene mediated resistance in tomato during aphid feeding (Martinez de Ilarduya et al. 2003), while in tobacco it was observed that there was an inverse relationship between endogenous concentrations of SA and JA (Felton et al. 1999). Exogenous application of SA has been shown to inhibit both JA synthesis and down-stream defenses in tomato (Chandok et al. 2004; Thaler et al. 2010). Moreover, it was shown that ET enhanced the plant response to SA, resulting in the expression of the SA-responsive marker gene pathogenesis-related-1 (Leon-Reyes et al. 2009). ET can synergistically or antagonistically interact with JA in the regulation of plant development and stress responses (O’Donnell et al. 1996; Zhao 2003). Many defense-related genes such as protein-ase inhibitors are regulated via a signaling pathway that requires both ET and JA (Adie et al. 2007a; Pieterse et al. 1998). In maize, the inhibition of ET biosynthesis or per-ception increased herbivore performance and damage in treated plants (Harfouche et al. 2006). However, ethylene acts antagonistically to suppress JA-induced expression of nicotine biosynthesis in tobacco (Rojo et al. 1999; Shoji et al. 2000). Exogenous ethylene application on Arabidop-sis increased the growth rate of the Egyptian cotton worm Spodoptera littoralis (Mantelin et al. 2009; Stotz et al. 2000). Bodenhausen and Reymond (2007) showed that the growth of the same caterpillar species on Arabidopsis was significantly lower on the ethylene-signaling mutant ein2-1 as compared to the wild-type control. These stud-ies on the interaction of plant hormones frequently show different impacts on plant defense, but some of the dif-ferences may be attributed to varied methodologies, treat-ments, etc. Furthermore, the interaction of plant hormones in defense depends on the specific plant-herbivore system studied.
Tomato has been a primary model system for elucidat-ing induced defenses and their signaling pathways against herbivores, yet most of the research has focused on signal-ing and rapidly-induced defenses in leaves (Melotto et al. 2008). Considerably less study has focused on systemic induction and the induction of defenses that require days to weeks to generations for expression (Rasmann et al. 2011; Tian et al. 2012a). In this study, we used tomato mutants and application of hormones to dissect the interactions of JA and ethylene that regulate plant systemic defense to a chewing herbivore, H. zea. Using the tomato never ripe (Nr) mutant, which is impaired in ethylene-mediated plant response, and the def1 mutant, which is deficient in JA bio-synthesis to study the hormone interactions, we employed a combination of insect bioassays, defense gene expression
Author's personal copy

Planta
1 3
assays, and plant hormone analyses to identify the key interactions between JA and ethylene.
Materials and methods
Plant materials and insects
Tomato (Lycopersicon esculentum) seeds of the def1 mutant and wild type Castlemart (CM) were kindly pro-vided by Dr. Gregg Howe (Michigan State University). Seeds for the Nr mutant (accession number LA3001) and the wild-type Rutgers (RU, accession number LA1090) were obtained from the Tomato Genetics Resource Center (University of California, Davis). All the plants were grown in Metromix 400 potting mix (Griffin Greenhouse & Nursery Supplies Tewksbury, MA) in a greenhouse at Pennsylvania State University, University Park, PA, USA). The greenhouse was maintained on a 16-h photo-period (Peiffer and Felton 2005). Tomato fruitworm (H. zea) eggs were purchased from BioServ (Frenchtown, NJ) and were reared on a wheat germ and casein-based artificial diet (Chippendale 1970) with ingredients from BioServ.
Plant treatments
To compare the effect of exogenous hormones on plant defense, tomato plants were sprayed with 2.5 mM methyl jasmonate (MeJA) in 0.8 % ethanol or 3 mM ethephon (2-chloroethylphosphonic acid, CEPA, Sigma, St. Louis, MO, USA) at the four-leaf-stage. The treated plants were allowed to grow in the greenhouse for another 2 weeks after treatment.
Two weeks after the spray treatments, the plant height was measured to compare how the exogenous hormones affected plant growth. Two leaf discs from the top fully expanded leaf were cut from each side of the mid-vein. Glandular trichome numbers were counted under a light microscope on each leaf disc. The density of trichomes was calculated as the number per disc/cm2.
To compare how plant hormones regulate the wound-ing response, 2 weeks after treatment, the newly emerged leaves on treated tomato plants were mechanically wounded by punching two ! in. diameter holes along the mid-vein of the terminal leaflet. 24 h after the wounding treatment, the wounded leaf was used for RNA extraction and gene expression assay.
To examine the effect of hormone treatment on seed-ling growth, we surface sterilized seeds and allowed them to germinate on agar plates with 100 µM MeJA or 100 µg/L ethephon in Murashige and Skoog (MS) medium (Sigma, St. Louis, USA). 1 week later, we
compared seedling growth by measuring the hypocotyl and root length. Also we collected the seedlings to com-pare the effect of hormone treatment on defense gene expression.
Insect bioassays
To determine if the hormones directly affected fruitworm growth, we used an artificial diet bioassay. Based on diet weight, 100 µg/g MeJA or 100 µg/g ethephon was added on the diet surface. After 20 min of drying, one neonate larva was placed in each cup. A total of 30 larvae were tested per treatment.
Detached leaf bioassays were conducted to compare how different plant treatments affected insect growth. Newly hatched H. zea larvae were reared on artificial diet for 1 day before transfer to the bioassay cups. A detached newly emerged leaf from plants was used for the bioassay. The treated leaves were put in a standard 1 oz cup (BioS-erv, Frenchtown, NJ) with 2 % agar on the bottom to main-tain moisture. Insects were kept at 27 °C, with a 16:8 L:D photoperiod in an incubator. 30 insects were used for each treatment. After 5 days, larval weights were recorded to the nearest 0.1 mg.
A choice test with MS medium grown seedling was con-ducted to compare how different treatments affect larval choice. The control and 2 treated seedlings were placed in the edge of a 150 ! 15 mm petri-dish and 15 first instar larvae were placed in the center. All experiments were repeated three times. After 24 h, we compared the number of caterpillars on each seedling.
RNA extraction and quantification analysis
To extract RNA, 100 mg leaf sample for each replicate was homogenized in liquid nitrogen and total RNA was purified with a RNeasy Plus Mini-kit (Qiagen, Valencia, CA, USA). High Capacity Transcription kit (Applied Biosystems, Fos-ter City, CA) was used for cDNA synthesis. All quantitative RT-PCR (qRT-PCR) reactions used Power SYBR Green PCR Master Mix and ran on 7500 Fast Real-Time PCR system (Applied Biosystems) following standard protocols (10 min at 95 °C, with 40 cycles of 15 s at 95 °C and 60 s at 60 °C). The ubiquitin gene was used as the housekeeping gene to normalize 2"!!C(T) (Rotenberg et al. 2006).
Plant hormone analysis
Plant hormone levels were determined 2 weeks after JA or ethephon treatment. To measure hormones, 100 mg of newly emerged leaves were collected to into FastPrep® tubes (Qbiogene, Carlsbad, CA) containing 1 g of Zirmil beads (1.1 mm; Saint-Gobain ZirPro, Mountainside, NJ),
Author's personal copy

Planta
1 3
and frozen at "80 °C until processing. (The newly emerged leaf would not have been directly exposed to the hormone treatments).
To extract and detect JA and SA, we used a previously described method (Tooker and De Moraes 2005), modi-fied slightly from that of Schmelz et al. (2003). Briefly, we derivatived the carboxylic acid to its methyl ester, which was isolated using vapor phase extraction and analyzed by GC–MS with isobutane chemical ionization using selected-ion monitoring.
Statistical analysis
Data were analyzed using general linear models for analy-sis of variance where appropriate with post hoc comparison of means using the Fisher-LSD means separation (Mintab Software, State College, PA).
Results
Hormone application with ethephon and MeJA affect tomato growth
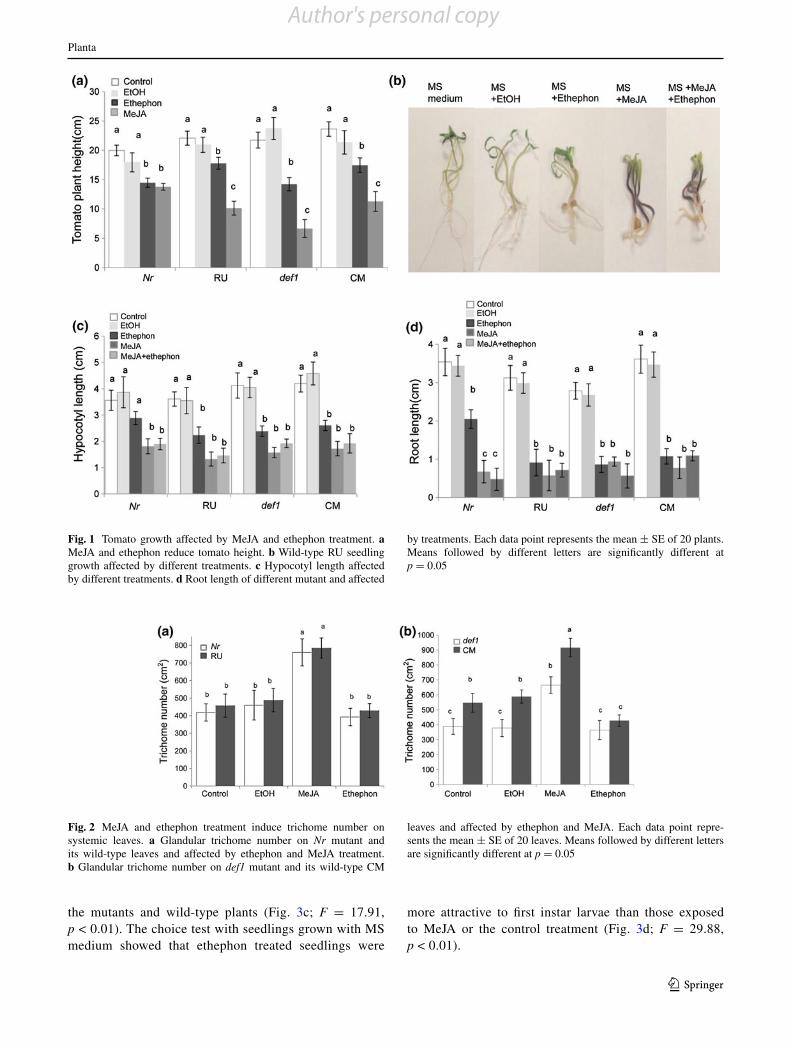
To examine how MeJA and ethephon affect plant growth, plant height was measured 2 weeks after treatments. The def1 and Nr mutant without hormone treatment grew nor-mally during this period and did not show any growth reduction compared to their respective wild-type plants. MeJA/ethephon treatments exhibited significant changes on tomato growth. Compared to the untreated plants, plant height in the Nr mutant was reduced 12.5 % by MeJA treatment and 31.0 % by the ethephon treatment. Whereas def1 mutant plant height was reduced 34.5 and 69.2 % by each treatment, respectively. Although Nr mutant displays reduced ethylene sensitivity, the ethephon treatment still reduced growth. Both wild-type plants showed significantly reduced plant growth after the treatment. After ethephon treatment, the height of RU and CM plants were reduced by 54.1 and 52.3 %, respectively. The MeJA treatment reduced the height by 22.6 and 26.1 % on both RU and CM plants (Fig. 1a, F = 97.48, p < 0.01).
When MeJA and ethephon were both added to MS medium plates at same time, the seedling growth was sig-nificantly affected by the treatment. Compared to control seedlings, both MeJA and ethephon significantly reduced hypocotyls length and root growth (Fig. 1b; F = 28.73, p < 0.001 and c; F = 7.01, p < 0.001).
Trichome production regulated by hormone application
A striking reduction in type VI glandular trichome den-sity was observed on JA deficient def1 mutant compared
to the wild-type CM trichome number (Fig. 2a; F = 27.18, p < 0.01). The trichome numbers on the def1 mutant were 40.9 % less than on CM. While the trichome numbers on the Nr mutant did not show significant differences com-pared to wild-type Rutgers (RU) plants (Fig. 2b; F = 0.15, p = 0.696).
MeJA and ethephon exhibited different effects on tri-chome production. Trichome density on both mutant and wild-type plants was significantly increased by MeJA treatment. The def1 mutant and CM plants showed a 67.5 and 71.4 % increase, respectively. Trichome density on Nr and RU was increased by 81.6 and 71.5 %, respectively as compared to the untreated control. Conversely the tri-chome density on ET-treated plants was decreased from 6.5 to 27 %, but with no significant reduction compared to the untreated mutants and wild-type plants (Fig. 2a, b). These results are consistent with a previous study in which MeJA induced trichomes on new-growth leaves and dem-onstrates the central role that JA plays in trichome forma-tion (Boughton et al. 2005). In previous studies, assays testing the effects of ethylene on trichome production have produced varying results, but this may be to due to differ-ences in application time, concentration, or host plant iden-tity (Gibson 2003; Plett et al. 2009). Our results did not indicate a significant interactive effect of JA and ET on tri-chome production.
Hormone application and systemic resistance against H. zea
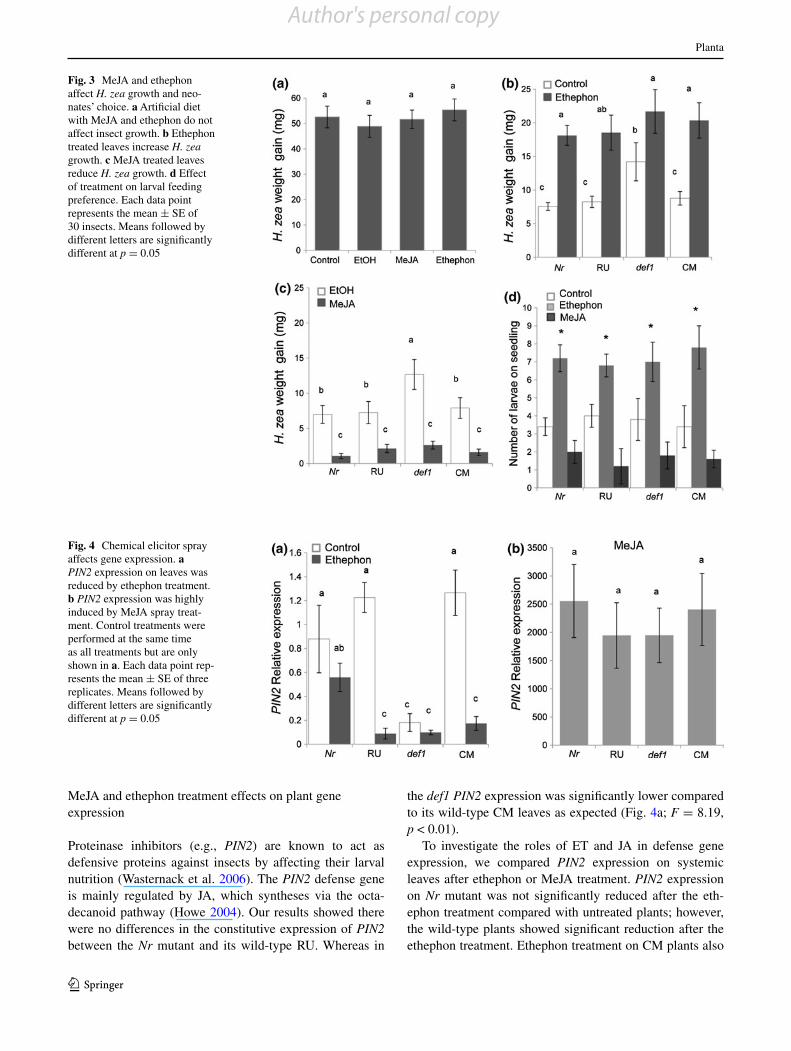
While various studies have shown that plant hormones such as JA and ET regulate plant defense to insect feed-ing (Adie et al. 2007a; Ankala et al. 2009), we examined whether the hormones had direct effects on the insect. Neither MeJA nor ethephon in diet affected larval growth rates over a 5-day feeding period (Fig. 3a; F = 0.23, p = 0.799).
We then tested the growth of larvae on the ethylene and JA mutants. Bioassays with leaves from Nr mutant and RU wild-type plants showed that H. zea growth was not significantly different between the two genotypes (Fig. 3b; F = 9.42, p < 0.01). However, the growth of H. zea significantly increased on the def1 mutant compared to its wild-type CM (Fig. 3b). To assess how JA and ET treatments of plants affected insect growth, the new-growth leaves were removed for bioassay. The results showed that treatment of plants with ethephon signifi-cantly increased caterpillar growth on both mutants and wild-type plants compared to control plants (Fig. 3b). Because a small amount of ethanol was added to dis-solve MeJA, we also used an ethanol control treatment; nevertheless MeJA significantly reduced the caterpil-lar growth compared with the ethanol treatment in all
Author's personal copy
Planta
1 3
the mutants and wild-type plants (Fig. 3c; F = 17.91, p < 0.01). The choice test with seedlings grown with MS medium showed that ethephon treated seedlings were
more attractive to first instar larvae than those exposed to MeJA or the control treatment (Fig. 3d; F = 29.88, p < 0.01).
Fig. 1 Tomato growth affected by MeJA and ethephon treatment. a MeJA and ethephon reduce tomato height. b Wild-type RU seedling growth affected by different treatments. c Hypocotyl length affected by different treatments. d Root length of different mutant and affected
by treatments. Each data point represents the mean ± SE of 20 plants. Means followed by different letters are significantly different at p = 0.05
Fig. 2 MeJA and ethephon treatment induce trichome number on systemic leaves. a Glandular trichome number on Nr mutant and its wild-type leaves and affected by ethephon and MeJA treatment. b Glandular trichome number on def1 mutant and its wild-type CM
leaves and affected by ethephon and MeJA. Each data point repre-sents the mean ± SE of 20 leaves. Means followed by different letters are significantly different at p = 0.05
Author's personal copy
Planta
1 3
MeJA and ethephon treatment effects on plant gene expression
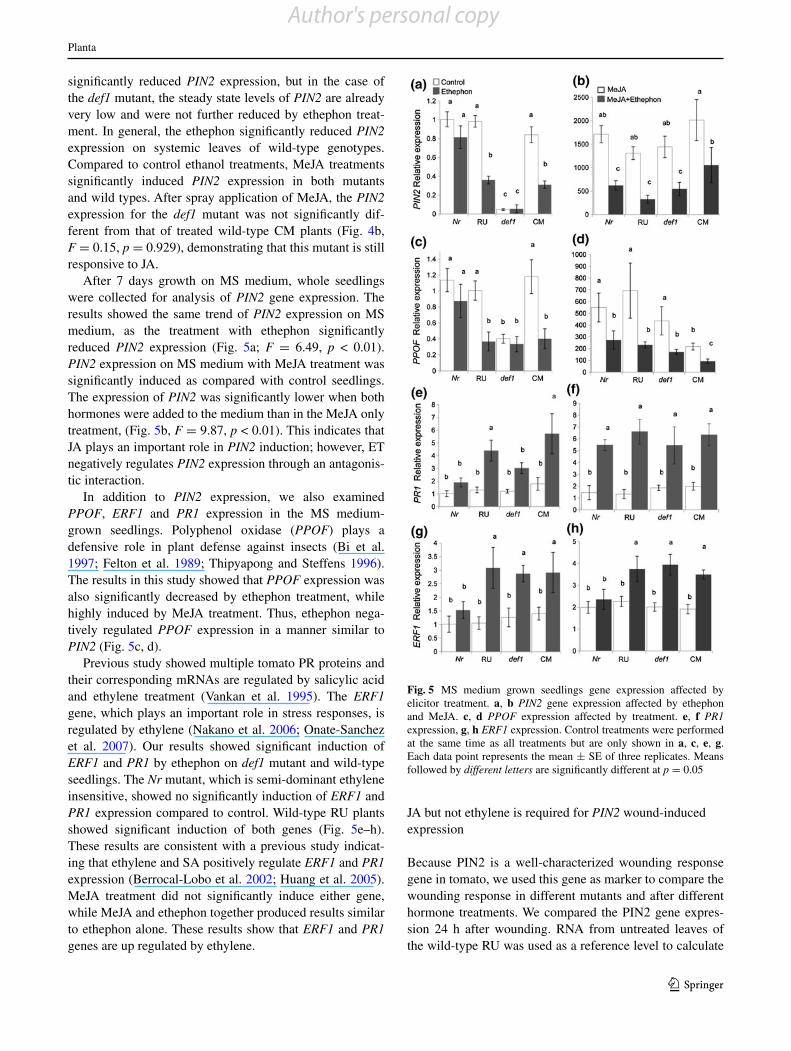
Proteinase inhibitors (e.g., PIN2) are known to act as defensive proteins against insects by affecting their larval nutrition (Wasternack et al. 2006). The PIN2 defense gene is mainly regulated by JA, which syntheses via the octa-decanoid pathway (Howe 2004). Our results showed there were no differences in the constitutive expression of PIN2 between the Nr mutant and its wild-type RU. Whereas in
the def1 PIN2 expression was significantly lower compared to its wild-type CM leaves as expected (Fig. 4a; F = 8.19, p < 0.01).
To investigate the roles of ET and JA in defense gene expression, we compared PIN2 expression on systemic leaves after ethephon or MeJA treatment. PIN2 expression on Nr mutant was not significantly reduced after the eth-ephon treatment compared with untreated plants; however, the wild-type plants showed significant reduction after the ethephon treatment. Ethephon treatment on CM plants also
Fig. 3 MeJA and ethephon affect H. zea growth and neo-nates’ choice. a Artificial diet with MeJA and ethephon do not affect insect growth. b Ethephon treated leaves increase H. zea growth. c MeJA treated leaves reduce H. zea growth. d Effect of treatment on larval feeding preference. Each data point represents the mean ± SE of 30 insects. Means followed by different letters are significantly different at p = 0.05
Fig. 4 Chemical elicitor spray affects gene expression. a PIN2 expression on leaves was reduced by ethephon treatment. b PIN2 expression was highly induced by MeJA spray treat-ment. Control treatments were performed at the same time as all treatments but are only shown in a. Each data point rep-resents the mean ± SE of three replicates. Means followed by different letters are significantly different at p = 0.05
Author's personal copy
Planta
1 3
significantly reduced PIN2 expression, but in the case of the def1 mutant, the steady state levels of PIN2 are already very low and were not further reduced by ethephon treat-ment. In general, the ethephon significantly reduced PIN2 expression on systemic leaves of wild-type genotypes. Compared to control ethanol treatments, MeJA treatments significantly induced PIN2 expression in both mutants and wild types. After spray application of MeJA, the PIN2 expression for the def1 mutant was not significantly dif-ferent from that of treated wild-type CM plants (Fig. 4b, F = 0.15, p = 0.929), demonstrating that this mutant is still responsive to JA.
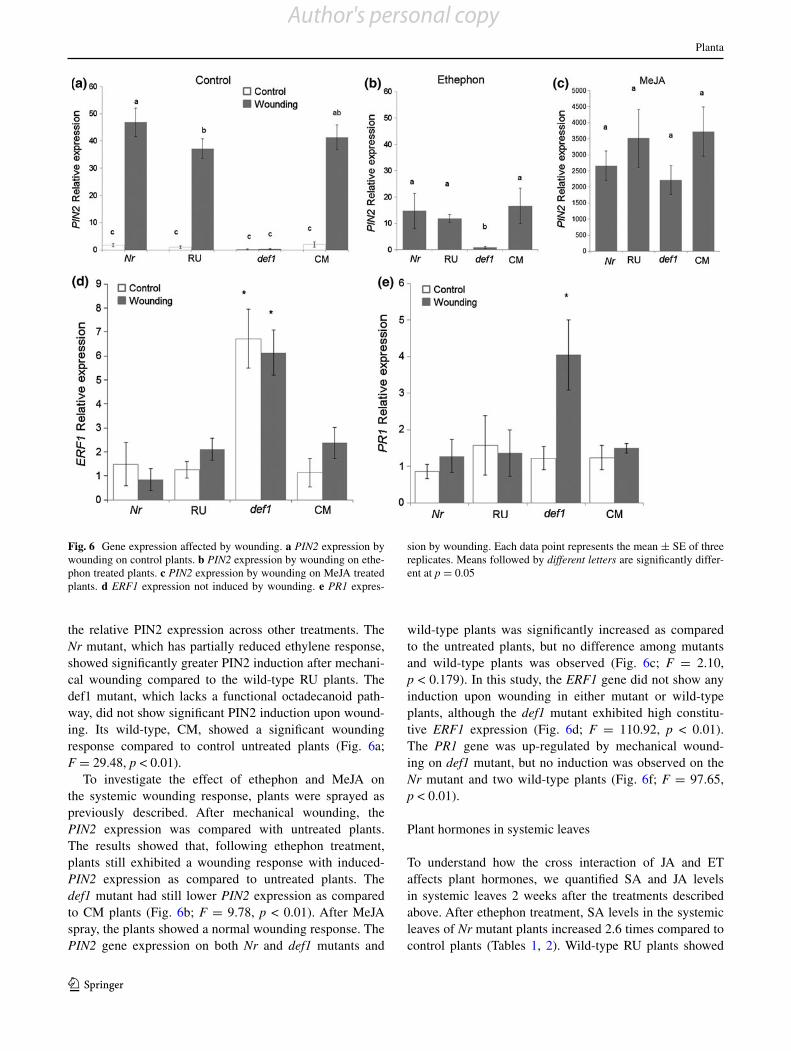
After 7 days growth on MS medium, whole seedlings were collected for analysis of PIN2 gene expression. The results showed the same trend of PIN2 expression on MS medium, as the treatment with ethephon significantly reduced PIN2 expression (Fig. 5a; F = 6.49, p < 0.01). PIN2 expression on MS medium with MeJA treatment was significantly induced as compared with control seedlings. The expression of PIN2 was significantly lower when both hormones were added to the medium than in the MeJA only treatment, (Fig. 5b, F = 9.87, p < 0.01). This indicates that JA plays an important role in PIN2 induction; however, ET negatively regulates PIN2 expression through an antagonis-tic interaction.
In addition to PIN2 expression, we also examined PPOF, ERF1 and PR1 expression in the MS medium-grown seedlings. Polyphenol oxidase (PPOF) plays a defensive role in plant defense against insects (Bi et al. 1997; Felton et al. 1989; Thipyapong and Steffens 1996). The results in this study showed that PPOF expression was also significantly decreased by ethephon treatment, while highly induced by MeJA treatment. Thus, ethephon nega-tively regulated PPOF expression in a manner similar to PIN2 (Fig. 5c, d).
Previous study showed multiple tomato PR proteins and their corresponding mRNAs are regulated by salicylic acid and ethylene treatment (Vankan et al. 1995). The ERF1 gene, which plays an important role in stress responses, is regulated by ethylene (Nakano et al. 2006; Onate-Sanchez et al. 2007). Our results showed significant induction of ERF1 and PR1 by ethephon on def1 mutant and wild-type seedlings. The Nr mutant, which is semi-dominant ethylene insensitive, showed no significantly induction of ERF1 and PR1 expression compared to control. Wild-type RU plants showed significant induction of both genes (Fig. 5e–h). These results are consistent with a previous study indicat-ing that ethylene and SA positively regulate ERF1 and PR1 expression (Berrocal-Lobo et al. 2002; Huang et al. 2005). MeJA treatment did not significantly induce either gene, while MeJA and ethephon together produced results similar to ethephon alone. These results show that ERF1 and PR1 genes are up regulated by ethylene.
JA but not ethylene is required for PIN2 wound-induced expression
Because PIN2 is a well-characterized wounding response gene in tomato, we used this gene as marker to compare the wounding response in different mutants and after different hormone treatments. We compared the PIN2 gene expres-sion 24 h after wounding. RNA from untreated leaves of the wild-type RU was used as a reference level to calculate
Fig. 5 MS medium grown seedlings gene expression affected by elicitor treatment. a, b PIN2 gene expression affected by ethephon and MeJA. c, d PPOF expression affected by treatment. e, f PR1 expression, g, h ERF1 expression. Control treatments were performed at the same time as all treatments but are only shown in a, c, e, g. Each data point represents the mean ± SE of three replicates. Means followed by different letters are significantly different at p = 0.05
Author's personal copy
Planta
1 3
the relative PIN2 expression across other treatments. The Nr mutant, which has partially reduced ethylene response, showed significantly greater PIN2 induction after mechani-cal wounding compared to the wild-type RU plants. The def1 mutant, which lacks a functional octadecanoid path-way, did not show significant PIN2 induction upon wound-ing. Its wild-type, CM, showed a significant wounding response compared to control untreated plants (Fig. 6a; F = 29.48, p < 0.01).
To investigate the effect of ethephon and MeJA on the systemic wounding response, plants were sprayed as previously described. After mechanical wounding, the PIN2 expression was compared with untreated plants. The results showed that, following ethephon treatment, plants still exhibited a wounding response with induced-PIN2 expression as compared to untreated plants. The def1 mutant had still lower PIN2 expression as compared to CM plants (Fig. 6b; F = 9.78, p < 0.01). After MeJA spray, the plants showed a normal wounding response. The PIN2 gene expression on both Nr and def1 mutants and
wild-type plants was significantly increased as compared to the untreated plants, but no difference among mutants and wild-type plants was observed (Fig. 6c; F = 2.10, p < 0.179). In this study, the ERF1 gene did not show any induction upon wounding in either mutant or wild-type plants, although the def1 mutant exhibited high constitu-tive ERF1 expression (Fig. 6d; F = 110.92, p < 0.01). The PR1 gene was up-regulated by mechanical wound-ing on def1 mutant, but no induction was observed on the Nr mutant and two wild-type plants (Fig. 6f; F = 97.65, p < 0.01).
Plant hormones in systemic leaves
To understand how the cross interaction of JA and ET affects plant hormones, we quantified SA and JA levels in systemic leaves 2 weeks after the treatments described above. After ethephon treatment, SA levels in the systemic leaves of Nr mutant plants increased 2.6 times compared to control plants (Tables 1, 2). Wild-type RU plants showed
Fig. 6 Gene expression affected by wounding. a PIN2 expression by wounding on control plants. b PIN2 expression by wounding on ethe-phon treated plants. c PIN2 expression by wounding on MeJA treated plants. d ERF1 expression not induced by wounding. e PR1 expres-
sion by wounding. Each data point represents the mean ± SE of three replicates. Means followed by different letters are significantly differ-ent at p = 0.05
Author's personal copy
Planta
1 3
a 6.4-fold increase compared to the control. Furthermore, SA levels in both the def1 mutant and wild-type CM plants increased ca. tenfold after ethephon treatment as compared to control plants. In contrast, MeJA did not significantly increase SA levels in either mutant or wild-type systemic leaves. As expected, JA levels were significantly increased on systemic leaves as compared to ethanol treatment after MeJA treatment. The ethanol treatment did not show any effect on JA levels. JA levels in thesystemic leaves of the Nr and def1 mutants showed no significant decrease after ethephon treatment; however, in RU plants, JA was reduced after ethephon treatment as compared to controls. However, JA in CM plants was not reduced by ethephon treatment (Table 2).
Discussion
The plant hormones SA, JA and ET are known to play important signaling roles in local and systemic plant defense against insect herbivores (Howe 2004; Onkokesung et al. 2010). JA signaling in particular plays a prominent role in promoting plant defense responses to many her-bivores (Bodenhausen and Reymond 2007; Cooper and Goggin 2005; McConn et al. 1997), while SA and ET fre-quently interact with JA in regulating such defenses (Adie et al. 2007a; Beckers and Spoel 2006; Campos et al. 2009; Wasternack et al. 2006). Both positive and negative effects of ET on JA-regulated responses have previously been observed (O’Donnell et al. 1996; Stotz et al. 2000).
Our results demonstrate the role of JA in defense against the caterpillar H. zea, but also show the potential nega-tive cross talk between JA and ET. Pretreatment of tomato plants with MeJA significantly decreased H. zea growth on both mutants and wild-type plants. Larval growth on the Nr mutant was not affected as compared to the wild-type plants, but pretreatment with ethephon compromised resist-ance to H. zea in both Nr and wild-type plants. This antag-onistic interaction has also been reported in other plants, including one previous study showing that pretreatment of Arabidopsis with ethephon elevated susceptibility to gen-eralist insects including the Egyptian cotton worm (Stotz et al. 2000). Ethylene negatively regulated local expression of lectin GS-II in Griffonia simplicifolia, which inhibited insect growth and development (Zhu-Salzman et al. 1998). Ethylene was previously reported to negatively regulate JA-induced nicotine biosynthesis and reduce direct plant defenses (Kahl et al. 2000). In contrast, blocking ethylene synthesis and perception in maize resulted in enhanced plant susceptibility to caterpillar feeding (Harfouche et al. 2006; Ankala et al. 2009). These studies suggest that the role of ET in defense may depend on the specific plant-her-bivore system being studied.
Glandular trichomes are an important component of the defense phenotype of many plants. Trichome density is regulated by jasmonates, brassinosteroids and ethyl-ene (Boughton et al. 2005; Campos et al. 2009; Tian et al. 2012a, b). And JA plays a positive role in glandular tri-chome induction in many plant species (Peiffer et al. 2009; Rasmann et al. 2011; Traw and Bergelson 2003; van Schie
Table 1 Primers used for real-time PCR assays of relative expression
Primer name Gene name/accession number Primer sequence (5!–3!)
PIN2 Wound inducible proteinase inhibitor 2/K03291 GGA TTT AGC GGA CTT CCT TCT G ATG CCA AGG CTT GTA CTA GAG AAT G
PPOF Polyphenol oxidase/z12837 ACT TTC ACG CGG TAA GGG TTA CGA ATG TGG ACA GGA TGT GGA ACG AGT
PR1 Pathogenesis related gene1/NM 001247429 GTC CCC AGC TGT TTG ATT GG TTG GAT TCG GTC CTA TGA CAT G
ERF1 Ethylene response factor1/EU395634 AATGGAATTAGGGTTTGGTTAGGAA GACCAAGGACCCCTCATTGA
Ubi Ubiquitin/X58253 GCC AAG ATC CAG GAC AAG GA GCT GCT TTC CGG CGA AA
Table 2 Salicylic acid and JA affected by MeJA and Ethephon
Means followed by different letters are significantly different at p = 0.05
Salicylic acid (ng/g fresh weight) JA (ng/g fresh weight)
Control Ethephon EtOH MeJA Control Ethephon EtOH MeJA
Nr 11.33 ± 2.27cd 29.83 ± 7.67b 10.70 ± 2.32cd 9.58 ± 2.95cd 23.76 ± 1.94c 15.25 ± 4.12c 21.50 ± 1.61c 283.63 ± 45.26b
RU 12.54 ± 3.92c 73.95 ± 8.14a 8.64 ± 2.76d 15.80 ± 1.09c 20.34 ± 2.88c 8.70 ± 1.78c 19.98 ± 2.12c 278.59 ± 32.32b
def1 6.32 ± 2.85d 66.97 ± 11.36a 9.41 ± 1.23cd 10.28 ± 3.58cd 11.13 ± 1.14c 6.44 ± 1.87c 12.38 ± 1.31c 384.62 ± 43.67a
CM 8.90 ± 1.38d 67.72 ± 13.02a 7.22 ± 2.21d 12.71 ± 4.57c 19.17 ± 4.32c 7.28 ± 2.24c 19.37 ± 2.39c 490.62 ± 51.78a
Author's personal copy

Planta
1 3
et al. 2007). In this study, the def1 mutant with JA defi-ciency showed significantly reduced trichome induction as compared to its wild-type, CM. In contrast, the Nr mutant did not show any trichome reduction ascompared to wild-type RU plants, although ethylene perception was partially blocked. There are varying reports about ET function in regulating trichome formation; for example, in Arabidop-sis, ET was found to inhibit trichome formation on stems and leaves (Gibson 2003) but, in another study, ET posi-tively regulated plant cell division and increased trichome initiation and development (Kazama et al. 2004). Exog-enous application of ET has been found to increase branch number in cucumber trichomes, and increased ethylene synthesis has been correlated with trichome branching (Plett et al. 2009). The different findings observed in these studies may be due to variation among plant species or to differences in ET application rates and timing. Ethylene may also regulate non-glandular trichome branching, but we did not examine its possible effect on these trichomes.
Plants respond to wounding or insect herbivory by acti-vating a series of wound-responsive defense genes (Bald-win et al. 2001; Bergey et al. 1999; Chen et al. 2004; Heinrich et al. 2011). Jasmonates are essential signaling molecules in this process (Gfeller et al. 2010). As expected, we observed lower expression of PIN2 in the def1 mutant as compared to CM plants. Wounding did not induce sig-nificant PIN2 expression in the def1 mutant as compared to wild-type CM plants. The ethylene insensitive mutant Nr showed normal constitutive and wound-induced PIN2 expression, similar to that of wild-type RU plants. This shows that JA, but not ET is required for the wounding response. The wounding response in systemic leaves was also affected by ethephon and MeJA treatment. MeJA treat-ment restored normal wounding responses in def1 mutants. The ethephon treatment decreased wound-inducible PIN2 gene expression in systemic leaves.
A previous study also reported that ethylene treatment reduced proteinase inhibitor 1(PIN1) gene expression in tomato (Diaz et al. 2002). However, a synergistic interac-tion of ET and JA in the regulation of proteinase inhibitor genes has been shown in some instances (Adie et al. 2007b; O’Donnell et al. 1996). Our study showed MeJA applica-tion induced PIN2 gene expression in systemic leaves, while ethephon treatment decreased PIN2 gene expres-sion. Our results contradict previous reports that wound-induced PIN2 gene expression depends on both JA and ET (O’Donnell et al. 1996). In those experiments, spe-cific inhibitors or elicitors, which regulate ET on plants, were applied to leaves and gene expression on local leaves was quantified after wounding, whereas in our study we focused on PIN2 expression in systemic leaves. The meth-ods employed also differed in some ways: our study used 3 mM ethephon spray on the plants, whereas some previous
studies incubated plants in airtight chambers with ethylene for a short time before wounding plants; furthermore we used quantitative RT-PCR to quantify the gene expression change upon treatment rather than northern blots or semi-quantitative RT-PCR.
The ethylene response factor 1 (ERF1) acts as a posi-tive regulator of JA and ET signaling and has been shown to play an important role in mediating defense responses in Arabidopsis (Lorenzo et al. 2003). Expression of ERF1 and the SA-regulated PR1 genes in the MS media-grown seedlings were not significantly induced by MeJA treat-ment, but were significantly induced by ethephon treat-ment. We did not find ERF1 to be induced by our method of mechanical wounding in either plant tested. The def1 mutant showed high constitutive ERF1 expression com-pared with wild-type plants. The ethephon treatment induced ERF1 expression and decreased the PIN2 wound-ing response. This reduction is consistent with a previ-ous report that ERF1 represses the wound-responsive PDF1.2 gene in Arabidopsis (Lorenzo et al. 2003). PR1 transcripts are regulated by high levels of SA and patho-gen attack in tomato plants (Tornero et al. 1997), but the effect of wounding on PR1 expression has not previously been reported. In this study, the Nr mutant and wild-type plants did not show PR1 induction by mechanical wound-ing, while this gene was induced by wounding in the def1 mutant. Because the def1 mutant is deficient in JA bio-synthesis, the lack of JA as a negative SA regulator may explain the induction of PR1. Our findings are consistent with previous reports that ERF1 and PR1 genes can be up-regulated by ethylene response (Cole et al. 2004; Lor-enzo et al. 2003).
Significant progress has been made in elucidating the interactive roles of SA, JA and ET in plant defense responses (Abe et al. 2011; Kniskern et al. 2007; Leon-Reyes et al. 2009). The current findings show that eth-ephon treatment significantly increases SA levels in sys-temic leaves while decreasing JA levels. Significant ERF1 and PR1 gene induction was observed when SA levels were increased by ethephon treatment. JA positively regulated PIN2 gene expression and glandular trichome density, but ET positively regulated SA biosynthesis and response with the induction of ERF1 and PR1 gene expression. While ET frequently interacts synergistically with JA to mediate insect resistance in some plant species, we found that ET inhibited induced resistance against the tomato fruitworm, decreasing the JA response as indi-cated by decreased PIN2 and PPOF gene expression and insect bioassays.
Acknowledgments Dawn S. Luthe reviewed an earlier draft of the manuscript and provided helpful criticism. The work was supported in part by the United States Department of Agriculture–Agricultural and Food Research Initiative grant 2011-67013-30352.
Author's personal copy

Planta
1 3
References
Abe H, Tomitaka Y, Shimoda T, Seo S, Sakurai T, Kugimiya S, Tsuda S, Kobayashi M (2011) Antagonistic plant defense system regu-lated by phytohormones assists interactions among vector insect, thrips, and a tospovirus. Plant Cell Physiol 10:1093
Adie B, Chico JM, Rubio-Somoza I, Solano R (2007a) Modulation of plant defenses by ethylene. J Plant Growth Regul 26:160–177
Adie B, Pérez-Pérez J, Pérez-Pérez MM, Godoy M, Sánchez-Serrano J-J, Schmelz EA, Solano R (2007b) ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 19:1665–1681
Agrawal AA (1999) Induced responses to herbivory in wild rad-ish: effects on several herbivores and plant fitness. Ecology 80:1713–1723
Ament K, Kant MR, Sabelis MW, Haring MA, Schuurink RC (2004) Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato. Plant Physiol 135:2025–2037
An LZ, Xu XF, Tang HG, Zhang MX, Hou ZD, Liu YH, Zhao ZG, Feng HY, Xu SJ, Wang XL (2006) Ethylene production and 1-ami-nocyclopropane-1-carboxylate (ACC) synthase gene expression in tomato (Lycopersicon esculentum Mill.) leaves under enhanced UV-B radiation. J Integr Plant Biol 48:1190–1196
Ankala A, Luthe DS, Williams WP, Wilkinson JR (2009) Integration of ethylene and jasmonic acid signaling pathways in the expres-sion of maize defense protein Mir 1-CP. Mol Plant Microbe Inter-act 22:1555–1564
Baldwin IT, Halitschke R, Kessler A, Schittko U (2001) Merging molecular and ecological approaches in plant-insect interactions. Curr Opin Plant Biol 4:351–358
Beckers GJM, Spoel SH (2006) Fine-tuning plant defence signalling: Salicylate versus jasmonate. Plant Biol 8:1–10
Bergey DR, Orozco-Cardenas M, de Moura DS, Ryan CA (1999) A wound- and systemin-inducible polygalacturonase in tomato leaves. Proc Natl Acad Sci USA 96:1756–1760
Berrocal-Lobo M, Molina A, Solano R (2002) Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. Plant J 29:23–32
Bi JL, Murphy JB, Felton GW (1997) Antinutritive and oxidative components as mechanisms of induced resistance in cotton to Helicoverpa zea. J Chem Ecol 23:97–117
Bodenhausen N, Reymond P (2007) Signaling pathways controlling induced resistance to insect herbivores in Arabidopsis. Mol Plant Microbe Interact 20:1406–1420
Bostock RM (2005) Signal crosstalk and induced resistance: strad-dling the line between cost and benefit. Annu Rev Phytopathol 43:545–580
Boughton AJ, Hoover K, Felton GW (2005) Methyl jasmonate appli-cation induces increased densities of glandular trichomes on tomato, Lycopersicon esculentum. J Chem Ecol 31:2211–2216
Browse J (2009) Jasmonate passes muster: a receptor and targets for the defense hormone. Annu Rev Plant Biol 60:183–205
Campos ML, de Almeida M, Rossi ML, Martinelli AP, Litholdo CG, Figueira A, Rampelotti-Ferreira FT, Vendramim JD, Benedito VA, Peres LEP (2009) Brassinosteroids interact negatively with jasmonates in the formation of anti-herbivory traits in tomato. J Exp Bot 60:4346–4360
Chandok MR, Ekengren SK, Martin GB, Klessig DF (2004) Sup-pression of pathogen-inducible NO synthase (iNOS) activity in tomato increases susceptibility to Pseudomonas syringae. Proc Natl Acad Sci USA 101:8239–8244
Chen H, McCaig BC, Melotto M, He SY, Howe GA (2004) Regu-lation of plant arginase by wounding, jasmonate, and the phyto-toxin coronatine. J Biol Chem 279:45998–46007
Chippendale GM (1970) Metamorphic changes in fat body proteins of the southwestern corn borer, Diatraea grandiosella. J Insect Physiol 16:1057–1068
Cole AB, Kiraly L, Lane LC, Wiggins BE, Ross K, Schoelz JE (2004) Temporal expression of PR-1 and enhanced mature plant resist-ance to virus infection is controlled by a single dominant gene in a new Nicotiana hybrid. Mol Plant Microbe Interact 17:976–985
Cooper WR, Goggin FL (2005) Effects of jasmonate-induced defenses in tomato on the potato aphid, Macrosiphum euphor-biae. Entomologia Experimentalis et Applicata 115:107–115
Diaz J, ten Have A, van Kan JAL (2002) The role of ethylene and wound signaling in resistance of tomato to Botrytis cinerea. Plant Physiol 129:1341–1351
Duffey SS (1986) Plant glandular trichomes: their partial role in defence against insects. In: Juniper BE, Southwood TE (eds) Insects and the plant surface. Arnold, London, pp 151–172
Fan J, Hill L, Crooks C, Doerner P, Lamb C (2009) Abscisic acid has a key role in modulating diverse plant-pathogen interactions. Plant Physiol 150:1750–1761
Felton GW, Broadway RM, Duffey SS (1989) Inactivation of protease inhibitor activity by plant-derived quinones complications for host-plant resistance against noctuid herbivores. J Insect Physiol 35:981–990
Felton GW, Korth KL, Bi JL, Wesley SV, Huhman DV, Mathews MC, Murphy JB, Lamb C, Dixon RA (1999) Inverse relationship between systemic resistance of plants to microorganisms and to insect herbivory. Curr Biol 9:317–320
Gfeller A, Liechti R, Farmer EE (2010) Arabidopsis jasmonate signal-ing pathway. Sci Signal 3: cm3
Gibson S (2003) Ethylene inhibits trichome formation on stems and brancing on leaves. Int Conf 14:501707200
Halitschke R, Stenberg JA, Kessler D, Kessler A, Baldwin IT (2008) Shared signals—alarm calls’ from plants increase apparency to herbivores and their enemies in nature. Ecol Lett 11:24–34
Harfouche AL, Shivaji R, Stocker R, Williams PW, Luthe DS (2006) Ethylene signaling mediates a maize defense response to insect herbivory. Mol Plant Microbe Interact 19:189–199
Heinrich M, Baldwin IT, Wu J (2011) Two mitogen-activated protein kinase kinases, MKK1 and MEK2, are involved in wounding- and specialist lepidopteran herbivore Manduca sexta-induced responses in Nicotiana attenuata. J Exp Bot 62:4355–4365
Horgan FG, Quiring DT, Lagnaoui A, Pelletier Y (2009) Effects of altitude of origin on trichome-mediated anti-herbivore resistance in wild Andean potatoes. Flora 204:49–62
Howe GA (2004) Jasmonates as signals in the wound response. J Plant Growth Regul 23:223–237
Howe GA, Jander G (2008) Plant immunity to insect herbivores. Annu Rev Plant Biol 59:41–66
Huang Z, Yeakley JM, Garcia EW, Holdridge JD, Fan JB, Whitham SA (2005) Salicylic acid-dependent expression of host genes in compat-ible Arabidopsis-virus interactions. Plant Physiol 137:1147–1159
Kahl J, Siemens DH, Aerts RJ, Gabler R, Kühnemann F, Preston CA, Baldwin IT (2000) Herbivore-induced ethylene suppresses a direct defense but not a putative indirect defense against an adapted herbivore. Planta 210:336–342
Kang JH, Shi F, Jones AD, Marks MD, Howe GA (2010) Distor-tion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry. J Exp Bot 61:1053–1064
Kazama H, Dan H, Imaseki H, Wasteneys GO (2004) Transient exposure to ethylene stimulates cell division and alters the fate and polarity of hypocotyl epidermal cells. Plant Physiol 134:1614–1623
Kessler A, Halitschke R, Poveda K (2011) Herbivory-mediated pol-linator limitation: negative impacts of induced volatiles on plant-pollinator interactions. Ecology 92:1769–1780
Author's personal copy

Planta
1 3
Kniskern JM, Traw MB, Bergelson J (2007) Salicylic acid and jas-monic acid signaling defense pathways reduce natural bacterial diversity on Arabidopsis thaliana. Mol Plant Microbe Interact 20:1512–1522
Koiwa H, Shade RE, Zhu-Salzman K, Subramanian L, Murdock LL, Nielsen SS, Bressan RA, Hasegawa PM (1998) Phage display selection can differentiate insecticidal activity of soybean cysta-tins. Plant J 14:371–379
Leon-Reyes A, Spoel SH, De Lange ES, Abe H, Kobayashi M, Tsuda S, Millenaar FF, Welschen RAM, Ritsema T, Pieterse CMJ (2009) Ethylene modulates the role of NPR1 in cross-talk between salic-ylate and jasmonate signaling. Plant Physiol 108:133926
Li L, Zhao YF, McCaig BC, Wingerd BA, Wang JH, Whalon ME, Pichersky E, Howe GA (2004) The tomato homolog of CORO-NATINE-INSENSITIVE1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glan-dular trichome development. Plant Cell 16:126–143
Lorenzo O, Piqueras R, Sánchez-Serrano JJ, Solano R (2003) ETH-YLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 15:165–178
Mantelin S, Kishor KB, Isgouhi K (2009) Ethylene contributes to potato aphid susceptibility in a compatible tomato host. New Phytol 183:444–456
Martinez de Ilarduya O, Xie Q, Kaloshian I (2003) Aphid-induced defense responses in Mi-1-mediated compatible and incompat-ible tomato interactions. Mol Plant Microbe Interact 16:699–708
McConn M, Creelman RA, Bell E, Mullet JE, Browse J (1997) Jas-monate is essential for insect defense Arabidopsis. Proc Natl Acad Sci USA 94:5473–5477
Melotto M, Mecey C, Niu Y, Chung HS, Katsir L, Yao J, Zeng WQ, Thines B, Staswick P, Browse J, Howe GA, He SY (2008) A criti-cal role of two positively charged amino acids in the Jas motif of Arabidopsis JAZ proteins in mediating coronatine- and jas-monoyl isoleucine-dependent interactions with the COI1F-box protein. Plant J 55:979–988
Nakano T, Suzuki K, Fujimura T, Shinshi H (2006) Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol 140:411–432
Navarro L (2006) A plant miRNA contributes to antibacterial resist-ance by repressing auxin signaling. Science 312:436–439
O’Donnell PJ, Calvert C, Atzorn R, Wasternack C, Leyser HMO, Bowles DJ (1996) Ethylene as a signal mediating the wound response of tomato plants. Science 274:1914–1917
Onate-Sanchez L, Anderson JP, Young J, Singh KB (2007) AtERF14, a member of the ERF family of transcription factors plays a non-redundant role in plant defense. Plant Physiol 143:400–409
Onkokesung N, Galis I, von Dahl CC, Matsuoka K, Saluz H-P, Baldwin IT (2010) Jasmonic acid and ethylene modulate local responses to wounding and simulated herbivory in Nicotiana attenuata leaves. Plant Physiol 153:785–798
Peiffer M, Felton GW (2005) The host plant as a factor in the syn-thesis and secretion of salivary glucose oxidase in larval Helicov-erpa zea. Arch Insect Biochem Physiol 58:106–113
Peiffer M, Tooker JF, Luthe DS, Felton GW (2009) Plants on early alert: glandular trichomes as sensors for insect herbivores. New Phytol 184:644–656
Pieterse CMJ, Dicke M (2007) Plant interactions with microbes and insects: from molecular mechanisms to ecology. Trends Plant Sci 12:564–569
Pieterse CM, van Wees SC, van Pelt JA, Knoester M, Laan R, Gerrits H, Weisbeek PJ, van Loon LC (1998) A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 10:1571–1580
Plett JM, Mathur J, Regan S (2009) Ethylene receptor ETR2 controls trichome branching by regulating microtubule assembly in Arabi-dopsis thaliana. J Exp Bot 60:3923–3933
Rasmann S, De Vos M, Casteel CL, Tian D, Halitschke R, Sun JY, Agrawal AA, Felton GW, Jander G (2011) Herbivory in the previ-ous generation primes plants for enhanced insect resistance. Plant Physiol 158(2):854–863
Reymond P, Weber H, Damond M, Farmer EE (2000) Differential gene expression in response to mechanical wounding and insect feeding in Arabidopsis. Plant Cell 12:707–720
Rojo E, León J, Sánchez-Serrano JJ (1999) Cross-talk between wound signalling pathways determines local versus systemic gene expression in Arabidopsis thaliana. Plant J 20:135–142
Rotenberg D, Thompson TS, German TL, Willis DK (2006) Methods for effective real-time RT-PCR analysis of virus-induced gene silencing. J Virological Meth 138:49–59
Schmelz EA, Engelberth J, Alborn HT, O’Donnell P, Sammons M, Toshima H, Tumlinson JH 3rd (2003) Simultaneous analysis of phytohormones, phytotoxins, and volatile organic compounds in plants. Proc Natl Acad Sci USA 100:10552–10557
Shoji T, Nakajima K, Hashimoto T (2000) Ethylene suppresses jas-monate-induced gene expression in nicotine biosynthesis. Plant Cell Physiol 41:1072–1076
Stotz HU, Pittendrigh BR, Kroymann J, Weniger K, Fritsche J, Bauke A, Mitchell-Olds T (2000) Induced plant defense responses against chewing insects. Ethylene signaling reduces resistance of Arabidopsis against Egyptian cotton worm but not diamondback moth. Plant Physiol 124:1007–1017
Thaler JS, Agrawal AA, Halitschke R (2010) Salicylate-medi-ated interactions between pathogens and herbivores. Ecology 91:1075–1082
Thaler JS, Humphrey PT, Whiteman NK (2012) Evolution of jasmonate and salicylate signal cross-talk. Trends Plant Sci 17:260–270
Thipyapong P, Steffens JC (1996) Defensive role of polyphenol oxi-dases against Pseudomonas syringae pv tomato. Plant Physiol 111:791
Tian D, Peiffer M, Shoemaker E, Tooker J, Haubruge E, Francis F, Luthe DS, Felton GW (2012a) Salivary glucose oxidase from caterpillars mediates the induction of rapid and delayed-induced defenses in the tomato plant. PLoS ONE 7:e36168
Tian D, Tooker J, Peiffer M, Chung S, Felton G (2012b) Role of tri-chomes in defense against herbivores: comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 236:1053–1066
Tooker JF, De Moraes CM (2005) Jasmonate in lepidopteran eggs and neonates. J Chem Ecol 31:2753–2759
Tornero P, Gadea J, Conejero V, Vera P (1997) Two PR-1 genes from tomato are differentially regulated and reveal a novel mode of expression for a pathogenesis-related gene during the hypersen-sitive response and development. Mol Plant Microbe Interact 10:624–634
Traw MB, Bergelson J (2003) Interactive effects of jasmonic acid, salicylic acid, and gibberellin on induction of trichomes in Arabi-dopsis. Plant Physiol 133:1367–1375
van Poecke RMP, Dicke M (2002) Induced parasitoid attraction by Arabidopsis thaliana: involvement of the octadecanoid and the salicylic acid pathway. J Exp Bot 53:1793–1799
van Schie CCN, Haring MA, Schuurink RC (2007) Tomato linalool synthase is induced in trichomes by jasmonic acid. Plant Mol Biol 64:251–263
Vankan JAL, Cozijnsen T, Danhash N, Dewit P (1995) Induction of tomato stress protein mRNAs by ethephon, 2,6-dichloroisonico-tinic acid and salicylate. Plant Mol Biol 27:1205–1213
Vera P, Conejero V (1990) Effect of ethephon on protein degradation and the accumulation of ‘pathogenesis-related’ (PR) proteins in tomato leaf discs. Plant Physiol 92:227–233
Verhage A, van Wees SCM, Pieterse CMJ (2010) Plant immunity: it’s the hormones talking, but what do they say? Plant Physiol 154:536–540
Author's personal copy

Planta
1 3
Walling LL (2000) The myriad plant responses to herbivores. J Plant Growth Regul 19:195–216
Wasternack C, Stenzel I, Hause B, Hause G, Kutter C, Maucher H, Neumerkel J, Feussner I, Miersch O (2006) The wound response in tomato—role of jasmonic acid. J Plant Physiol 163:297–306
Yang SF, Hoffman NE (1984) Ethylene biosynthesis and its regulation in higher plants. Annu Rev Plant Physiol 35:155–189
Yasuda M, Nakashita H, Hasegawa S, Nishioka M, Arai Y, Uramoto M, Yamaguchi I, Yoshida S (2003) N-cyanomethyl-2-chloroison-icotinamide induces systemic acquired resistance in Arabidopsis without salicylic acid accumulation. Biosci Biotechnol Biochem 67:322–328
Zhang Y, Xu S, Ding P, Wang D, Cheng YT, He J, Gao M, Xu F, Li Y, Zhu Z, Li X, Zhang Y (2010) Control of salicylic acid synthe-sis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc Natl Acad Sci USA 107:18220–18225
Zhang L, Jia C, Liu L, Zhang Z, Li C, Wang Q (2011) The involve-ment of jasmonates and ethylene in Alternaria alternata f. sp. lycopersici toxin-induced tomato cell death. J Exp Bot 62:5405–5418
Zhao Y (2003) Virulence systems of Pseudomonas syringae pv. tomato promote bacterial speck disease in tomato by targeting the jasmonate signaling pathway. Plant J 36:485–499
Zhu-Salzman K, Salzman RA, Koiwa H, Murdock LL, Bressan RA, Hasegawa PM (1998) Ethylene negatively regulates local expression of plant defense lectin genes. Physiologia Plantarum 104:365–372
Zsogon A, Lambais MR, Benedito VA, Figueira AVD, Peres LEP (2008) Reduced arbuscular mycorrhizal colonization in tomato ethylene mutants. Scientia Agricola 65:259–267
Author's personal copy





























