Embed Size (px)
Citation preview

SYNAPSE 6:328-337 (1990)
Effects of Short-Term Serotonin Depletion - on the Efficacy of Serotonin
Neurotransmission: Electrophysiological Studies in the Rat Central Nervous System
YVES CHAPUT, PHILIPPE LESIEUR, AND CLAUDE DE MONTIGNY Neurobiological Psychiatry Unit, De artment of Psychiatry, McGill University,
Montreal, Queb)ec, Canada H3A IAI
KEY WORDS p-Chlorophenylalanine, 5-HT, Dorsal hippocampus, Dorsal raphe, 5 , 7 -Dihydroxytryp tamine
ABSTRACT The effects of short-term serotonin (5-HT) depletion by p-chlorophenyl- alanine (PCPA) on the firing activity of dorsal raphe nucleus 5-HT neurons, on the responsiveness of dorsal hippocampus pyramidal neurons to microiontophoretically applied 5-HT and on the efficacy of the electrical stimulation of the ascending 5-HT pathway in suppressing the firing activity of CA, dorsal hippocampus pyramidal neurons were assessed in chloral hydrate-anesthetized rats. PCPA (250 mg/kg/day i.p. for 2 days) reduced the 5-HT content of the dorsal hippocampus by 90%. However, the number of spontaneously active 5-HT neurons per microelectrode trajectory through the dorsal raphe or their average rate of firing was unaltered. The effect of afferent 5-HT pathway stimulation was reduced in only 40% of treated rats, whereas the sensitivity of CA, pyramidal neurons to microiontophoretic 5-HT was not modified. The function of the terminal 5-HT autoreceptor was assessed using methiothepin, an autoreceptor antago- nist. Methiothepin (1 mg/kg, i.v.) significantly enhanced the efficacy of the stimulation in PCPA-treated rats, although the degree of enhancement was much less than in controls. A greater reduction of the effectiveness of the stimulation was obtained by increasing the dose of PCPA (350 mg/kg/day i.p. for 2 days). This regimen reduced the 5-HT content of the dorsal hippocampus by 95%. In these rats, the sensitivity of the terminal 5-HT autorecep- tor was assessed by increasing the frequency of the stimulation from 1 to 5 Hz. This procedure reduced to a similar extent the efficacy of the stimulation in treated and control rats, suggesting that the reduced effectiveness of methiothepin in enhancing 5-HT synaptic transmission in PCPA-treated rats is due to a lower degree of activation of the terminal 5-HT autoreceptor. The present results showing that the 350 mglkglday regimen of PCPA, but not the 250 mg/kg/day regimen, reduced the efficacy of the stimulation of the ascending 5-HT pathway suggest that a greater than 90% depletion is required to affect 5-HT neurotransmission significantly. The reduced level of activation of terminal 5-HT autoreceptors in rats treated with the lower dose of PCPA may facilitate the release of the remaining 5-HT per stimulation-triggered action potential, ensuring a virtually unaltered synaptic efficacy.
INTRODUCTION The clinical observation by Shopsin et al., (1975,
1976) of a rapid reversal of the therapeutic effect of two chemically distinct antidepressants (imipramine or tra- nylcypromine) by the short-term administration of the serotonin (5-HT) synthesis inhibitor para-chlorophenyl- alanine (PCPA) (Jequier et al., 1967; Koe and Weiss- man, 1966) to remitted patients constitutes strong evi- dence that the 5-HT system may play a pivotal role in mediating the therapeutic effect of antidepressant treatments. The recent report by the rapid mood lowering effect tryptophan in 0 1990 WILEY-LISS, INC.
patients oints to the same conclusion. There is a sub- stantial gody of clinical evidence in support of this contention, such as the therapeutic efficacy of selective 5-HT reuptake blockers and of 5-HT precursors in major depression (Asberg et al., 1986; Bech, 1988; Kaneko et al., 1979; Moller et al., 1980;Van Praag, 1981) as well as the potentiation of antidepressant treatments by L-tryptophan (Ayuso Guttirez and Lopez-Ibor, 1971; Coppen et al., 1963; Glassman and Platman, 1969; Pare, 1963; WAlinder et al., 1976) or by the addition of lithium
Received August 15,1989 accepted in revised form May 22,1990

EFFECT OF PCPA ON 5-HT NEUROTRANSMISSION 329
(de Montigny et al., 1981, 1983, 1985; Heninger et al., 1983; Nelson and Byck, 1982; Nelson and Mazure, 1986; Ryan et al., 1988).
Electrophysiological investi ations of the effects of
treatments in the rat suggest that an enhancement of 5-HT neurotransmission might be a common denomina- tor of treatments as diverse as tricyclic, tetracyclic, and atypical antidepressants; selective 5-HT reuptake blockers; monoamine oxidase inhibitors; and electro- convulsive shocks (for reviews, see Blier et al., 1987a; Willner, 1985). Furthermore, the time course of this potentiation of 5-HT neurotransmission is consistent with the delayed clinical efficacy of these treatments in ma'or de ression.
i h e efkcts of 5-HT depletion by the trypto han hy-
transmission in the rat hippocampus has never been extensively investigated. Segal(1975) reported that the short-term administration of PCPA to rats partially abolished one of two inhibitory effects of the electrical stimulation of either the dorsal or medial raphe nuclei on the firing activity of dorsal hip ocampus pyramidal
nent that was not modified by PCPA pretreatment was one characterized by a short latency and a short dura- tion (10 and 40 msec). This latter effect is similar in its characteristics to the suppression of the firing activity of hippocampus pyramidal neurons produced by the electrical activation of the ascending 5-HT athway in
shown to be a sensitive index of changes in the efficacy 5-HT synaptic transmission (Blier and de Montigny, 1983; Blier et al., 1987b; Chaput et al., 1986a,b). We therefore undertook to investigate, using an electro- physiological paradigm, the effects of short-term 5-HT depletion by PCPA on 5-HT neurotransmission in chlo- ral h drate-anesthetized rats. Three major constituents
activity of 5-HT-containing neurons of the dorsal raphe nucleus; 2) the efficacy of 5-HT synaptic transmission, as assessed by the effect of the electrical activation of the ascending 5-HT pathway on the firing activity of postsyna tic CA, dorsal hippocampus pyramidal neu-
5-HT applied microiontophoretically.
the repeated administration o 7 various antidepressant
droxylase inhibitor PCPA on the efficacy of 5- I! T neuro-
neurons. Interestingly, however, t K e inhibitory compo-
the ventromedial tegmentum (VMT), whic E has been
of 5- B T neurotransmission were examined: 1) the firing
rons; an Lp 3) the responsiveness of the same neurons to
MATERIAL AND METHODS Treatments
PCPA Male Sprague-Dawley rats (225-250 g) were admin-
istered either saline or PCPA (250 or 350 mgkglday i.p.) for 2 days. Electrophysiological experiments were car- ried out under chloral hydrate anesthesia (400 mgkg, i.p.), 24 hr after the second injection of saline or PCPA.
5,7-Dihydroxytryptamine Male Sprague-Dawley rats (200-225 g) were anesthe-
tized with chloral hydrate (400 mgkg i.p.) and mounted in a stereotaxic ap aratus, 1 hr following an intraperi-
tered to rotect noradrenergic neurons rom the neuro- f toneal injection o P desipramine (25 m kg), adminis-
toxic e ff ect of 5,7-dihydroxytryptamine (5,7-DHT). A
burr hole was drilled 1 mm lateral and 1 mm posterior to bre a and 200 pg free base of 5,7-DHT in 20 pl of 0.9%
lateral ventricle over a 5 min eriod. Control rats were
were carried out under chloral hydrate anest esia also, 10-14 days following the intracerebroventricular injec- tion of 5,7-DHT or saline.
Na t? 1 and 0.1% ascorbic acid was injected into the right
injected with saline. Electrop R ysiological ex eriments g
Recording from 5-HT-containing neurons of the nucleus raphe dorsalis
Sin le-barrelled glass micropipettes, preloaded with
ulled in the conventional manner and filled with a 2 M K aC1 solution saturated with fast green FCF (Fisher). A burr hole, 3 mm in diameter encompassing lambda, was drilled, and the micropipette was descended 1 mm anterior to lambda on the midline, into the dorsal raphe, usual1 attained at a depth of between 4.5 and 5.5 mm from t e surface of the brain.
Dorsal raphe 5-HT neurons were identified according to the criteria of Aghajanian (1978). Each 5-HT neuron encountered was recorded for a proximately 1 min in
electrode descent in the dorsal ra he consisted in a 1
electrode descents were performed: one 1 mm anterior to lambda on the midline, two 200 pm lateral to the first tract on each side, and two 200 pm rostra1 and caudal to the first electrode tract. A fast green deposit was left at the last recording site by passing a -26 pA current through the microelectrode for 15 min for subsequent histological verification of the location of recording sites.
fiberg 7 ass strands to promote capillary filling, were
order to obtain a measure of its 1 asal firing rate. Each
mm tract from the floor of the Sy P vius aqueduct. Five
Recording from CA, dorsal hippocampus pyramidal neurons
Five-barrelled glass micro ipettes, preloaded with
manner and their tips broken back to approximately 10 pm under microscopic control. The central barrel, used for extracellular unitary recordings, was filled with a 2 M NaCl solution saturated with fast green. The side barrels, used for microionto horesis, were filled with
40 mM in 200 mM NaC1, pH 4; Sigma), acetylcholine chloride (ACh; 20 mM in 200 mM NaC1, pH 4; Sigma). The fourth barrel, used for automatic current balancing, was filled with a 2 M NaCl solution.
The microelectrode was lowered 4.2 mm lateral and 4.2 mm anterior to lambda into the CA, region of the dorsal hip ocampus. Pyramidal neurons were identi- fied accorzing to the criteria of Kandel and Spencer (1961). Since these latter neurons demonstrate little spontaneous activity under chloral hydrate anesthesia, a leak or small ejection current of ACh (0-8 nA) was used to activate them within their hysiological range
midal neurons to 5-HT and to norepinephrine (NE) was assessed using the I.T,, method, i.e., by determining the charge in nC (current in nA X time in sec) required to obtain a 50% depression of firing rate from baseline (de Montigny and Aghajanian, 1977).
fiberglass strands, were pu P led in the conventional
the following solutions: 5-H 8 creatinine sulfate (0.5 or
(10-15 Hz). The responsiveness of K ippocampus pyra-

330 Y. CHAPUT ET AL
Electrical stimulation of the ascending 5-HT pathway
A bipolar concentric electrode NE-100 (David Kopf Instruments) was positioned with a lo" backward angle in the VMT, 1 mm anterior to lambda on the midline at a depth of 8 mm. Two stimulation paradigms were used. First, square 0.5 msec pulses of 300 p,A were delivered at a frequency of 1 Hz approximating that of the spon- taneous firing activity of dorsal raphe 5-HT neurons under chloral hydrate anesthesia while recording the firing activity of CA3 dorsal hippocampus pyramidal neurons. This control stimulation was chosen, because it has been shown to produce a detectable suppression of the firing activity of postsynaptic neurons in 93% of trials (Chaput et al., 1986a). These stimulation param- eters were chosen to carry out the present study because they result in a submaximal response of postsynaptic neurons. Indeed, the duration of the suppression of postsynaptic neuron firing activity can be prolonged by increasing the intensity of the current to 640 FA (Chaput et al., 1986a), by terminal 5-HT autoreceptor blockade (Blier et al., 1988; Chaput and de Montigny 1988; Chaput et al., 1986a,b), or by reducing the stimu- lation frequency from 1 to 0.5 Hz (Chaput and de Montigny, 1988). A second paradigm, whereby stimula- tions were carried out using a frequency of 5 Hz, was performed to assess the function of the terminal 5-HT autoreceptor. It has been observed in vivo that increas- ing the frequency of the stimulation of the 5-HT path- way from 1 to 5 Hz decreases its effectiveness (Blier et al., 1988; Chaput et al., 1986a). Presumably, the higher frequency of the stimulation results in a greater degree of activation of the terminal 5-HT autoreceptor at the time of arrival of the subsequent stimulation- triggered action potential. Therefore, each pulse a t the 5 Hz frequency releases a smaller amount of 5-HT. Fur- thermore, the effect of increasing the frequency of the stimulation from 1 to 5 Hz is reduced following long- term treatment of rats with drugs which decrease the function of the terminal 5-HT autoreceptor (Blier et al., 1988). This suggests that this approach provides a useful index of terminal 5-HT autoreceptor sensitivity.
Pulses were delivered through a direct-coupled isola- tion unit (SIU 478A, Grass). The unitary activity and the stimulation pulse were recorded on-line with an IBM-PC computer equipped with a Tecmar interface. Peristimulus time histograms of dorsal hippocampus pyramidal neuron firing activity were generated to measure the suppression of firing produced by the stimulation. The latter was quantified by computer as the absolute silence value (SIL), defined as the theoret- ical duration of a total suppression of firing that would represent the number of events suppressed by the stim- ulation. This latter value was calculated by computer by dividing the total number of events suppressed by the stimulation by the mean prestimulation frequency of firing of the neuron recorded. Thus the SIL value repre- sents the duration of the suppression of firing of these neurons normalized for their firing activity.
Biochemical analysis Brains were quickly removed and frozen on dry ice.
Coronal 300-~m-thick sections were cut in a cryostat at - 15°C. The CA, region was microdissected and punched
out. Tissue pellets were homogenized with a hand mi- crohomogenizer a t 4°C in 150 pl of 0.4 N HCl,,. Fifteen microliters were collected for protein determination (Lowry et al., 1951). The remaining homogenates were centrifu ed for 90 sec and the clear supernatant was collecte and analyzed for 5-HT by high- ressure liquid chromatography coupled with electroc emical detec- tion, as previously described (Chiew et al., 1982).
Statistical analysis One-way analysis of variance was used for the statis-
tical comparison of I.TSo values or SIL values obtained in PCPA- or 5,7-DHT-pretreated rats to those obtained in corresponding control rats. The statistical signifi- cance of the difference between the SIL values for the same hippocampus pyramidal neuron when the stimu- lation was carried out a t 1 and 5 Hz or prior to and following the acute intravenous injection methiothepin (MET) or 5-hydroxytryptophan (5-HTP) was assessed using the paired Student's t test. All results are ex- pressed as means ? SEM.
RESULTS Effect of PCPA treatment on 5-HT neuron
firing activity To obtain an index of the activity of 5-HT neurons
following PCPA, systematic electrode descents were performed into the dorsal raphe nucleus in control rats and in rats treated with PCPA (350 mg/kg/day for 2 days). The PCPA treatment modified neither the num- ber of spontaneously active 5-HT neurons encountered per tract nor their average discharge frequency (Tables I and 11). This confirms the finding of Wang and Aghaja- nian (1978) who observed a normal firing activity of dorsal raphe 5-HT neurons following a two-day PCPA pretreatment. Effect of PCPA treatments on the efficacy of 5-HT
synaptic transmission The 300 pA stimulation of the ascending 5-HT ath-
way at a frequency of 1 Hz in control rats resulte B in a suppression of hippocampus pyramidal neuron firing activity of 42 2 3.5 msec (Figs. 1A and 21, with a latency from the time of the stimulation to the onset of suppres- sion of 6 5 0.3 msec. The 5-HT content in the CA, region of the dorsal hippocampus of seven of these control animals was 10.3 2 0.6 ng/mg protein.
R f
TABLE I . Number of spontaneously active 5-HT neurons (k SEM) per 1 mm microelectrode track through the dorsal raphe nucleus
No. of 5-HT neurondtract No. of tracts
Control PCPA'
6.1 i 0.5 6.5 + 0.5
18 23
'Rats treated for 2 days with p-chlorophenylalanine (PCPA) at dose of 250 mg/kg/day.
TABLE II. Spontaneous firing activity (Hz f SEM) of 5-HT neurons in the dorsal raphe nucleus
No. of neurons recorded Firing activity
Control 1.07 i 0.07 109 PCPA' 1.12 i 0.08 148 'Rats treated for 2 days with PCPA at a dose of 250 mg/kg/day.
EFFECT OF PCPA ON 5-HT NEUROTRANSMISSION
100-
80 - - 5 60-
+I
J - 40-
07
v : 07
20 -
33 1
A - CONTROL
161
- PCPA (350 mglkgldayX2 days)
I
C - 5,7-DHT (200 pg, i.c.v.)
-50 -25 25 50 75 100
TIME (ms)
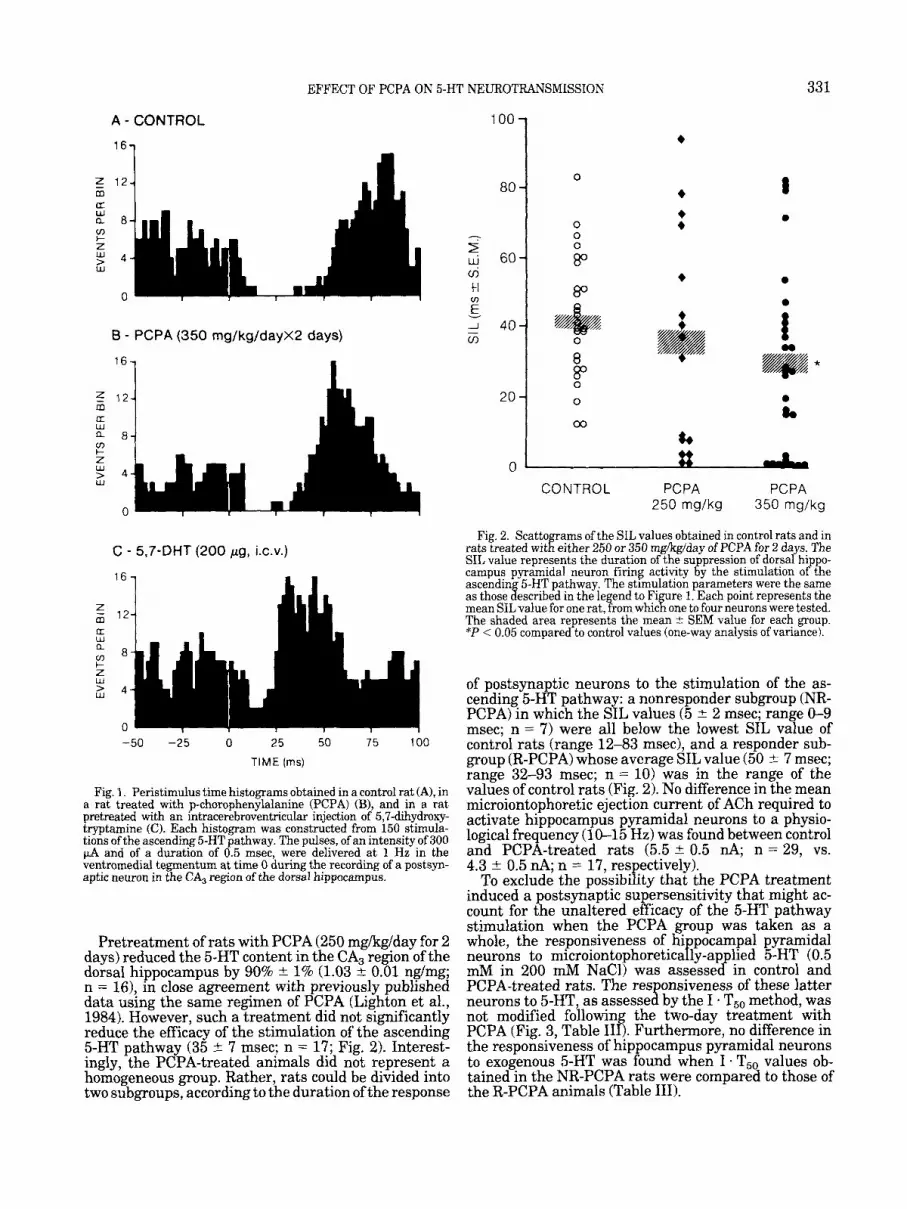
Fig. 1. Peristimulus time histograms obtained in a control rat (A), in a rat treated with p-chorophenylalanine (PCPA) (B), and in a rat pretreated with an intracerebroventricular injection of 5,7-dihydroxy- tryptamine (C). Each histogram was constructed from 150 stimula- tions of the ascending 5-HT pathway. The pulses, of an intensity of 300 p4 and of a duration of 0.5 msec, were delivered at 1 Hz in the ventromedial tegmentum at time 0 during the recording of a postsyn- aptic neuron in the CA, region of the dorsal hippocampus.
Pretreatment of rats with PCPA (250 mgkglday for 2 days) reduced the 5-HT content in the CA, region of the dorsal hippocampus by 90% 2 1% (1.03 2 0.01 nglmg; n = 16), in close agreement with previously published data using the same regimen of PCPA (Lighton et al., 1984). However, such a treatment did not significantly reduce the efficacy of the stimulation of the ascending 5-HT pathway (35 2 7 msec; n = 17; Fig. 2). Interest- ingly, the PCPA-treated animals did not represent a homogeneous group. Rather, rats could be divided into two subgroups, according to the duration of the response
+
+ + +
+
+ * k
0 @ CONTROL PCPA PCPA
250 m g l k g 350 m g l k g
Fig. 2. Scatto ams of the SIL values obtained in control rats and in rats treated witreither 250 or 350 mg/kg/day of PCPA for 2 days. The SIL value represents the duration of the su pression of dorsal hi PO campus pyramidal neuron firing activity !y the stimulation o?the ascendinf 5-HT pathway. The stimulation parameters were the same as those escribed in the legend to Fi re 1 Each point represents the mean SILvalue for one rat, from whicrone to four neurons weretested. The shaded area represents the mean k SEM value for each group. *P < 0.05 compared to control values (one-way analysis of variance).
of postsynaptic neurons to the stimulation of the as- cending 5-HT pathway: a nonresponder subgroup (NR- PCPA) in which the SIL values (5 * 2 msec; range 0-9 msec; n = 7) were all below the lowest SIL value of control rats (range 12-83 msec), and a responder sub- group (R-PCPA) whose average SIL value (50 % 7 msec; range 32-93 msec; n = 10) was in the range of the values of control rats (Fig. 2). No difference in the mean microiontophoretic ejection current of ACh required to activate hippocampus pyramidal neurons to a physio- logical frequency (10-15 Hz) was found between control and PCPA-treated rats (5.5 & 0.5 nA; n = 29, vs. 4.3 t 0.5 nA; n = 17, respectively).
To exclude the possibility that the PCPA treatment induced a postsynaptic supersensitivity that might ac- count for the unaltered efficacy of the 5-HT pathway stimulation when the PCPA group was taken as a whole, the responsiveness of hi pocampal pyramidal
mM in 200 mM NaC1) was assessed in control and PCPA-treated rats. The res onsiveness of these latter
not modified following the two-day treatment with PCPA (Fig. 3, Table 111). Furthermore, no difference in the responsiveness of hippocampus pyramidal neurons to exogenous 5-HT was found when I . T,, values ob- tained in the NR-PCPA rats were compared to those of the R-PCPA animals (Table 111).
neurons to microiontophoretica P ly-applied 5-HT (0.5
neurons to 5-HT, as assesse a by the I . T,, method, was

200 -
ln 0 --. 2 100- Y - 0
- a
0-
53 17 107 + 11 PCPA' NR-PCPA3 108 i 20 25 7 R-PCPA3 103 i 15 28 10
'The I.T5o value represents the charge (in nC) required to depress by 50W the firing activity of the neuron recorded. 2Rats treated for 2 days with PCPA a t a dose of 250 mg/kg/day. 3PCPA-treated rats were divided a posteriori in two subgroups: the NR-PCPA subgroup showing a markedly reduced response to electrical stimulation of the ascending 5-HT pathway and the R-PCPA showinga normal response (see text for details).
200 -)
ln 0 7
LLI 100- Y a m -
0-
(I) '' 40 2 - i (I) -
20
0 CONTROL PCPA
250 mg/kg

EFFECT OF PCPA ON 5-HT NEUROTRANSMISSION 333
mg/kg/day, i.p. for 2 days (n = 29). The 5-HT content of the CA, hippocam a1 region was determined in eight of
manipulation during the electrophysiological experi- ment: There was a 95% reduction of the 5-HT content (0.5 2 0.1 ng/mg rotein). This regimen of PCPA si if-
ascending 5-HT pathway (Figs. 1B and 2). However, as can be appreciated from the scattograms of Figure 2, there was a large variation in the efficacy of the stimu- lation of the ascending 5-HT pathway in this group of PCPA-treated rats also.
To assess terminal 5-HT autorece tor function follow-
increasin the frequency of the stimulation (from 1 to 5
sion was assessed in treated rats, where 5-HT pathway stimulation had yielded SIL values comparable to those obtained in the control rats (SIL values 3 30 msec). This ost-hoc analysis was based on the assumption
responsible for the unaltered efficacy of the stimulation in these treated rats, then increasing the frequency of the stimulation from 1 to 5 Hz should no longer reduce its efficacy, as it does in control animals as a result of an increased level of activation of the 5-HT terminal au- torece tor by endogenously released 5-HT (Blier et al.,
data, increasing the stimulation frequency from 1 to 5 Hz in control rats significantly reduced its efficacy (Fig. 5). A comparable degree of reduction was obtained at 5 Hz in the subgroup of PCPA-treated rats, which had a normal response to the 1 Hz stimulation. This suggests that the unaltered res onse to afferent 5-HT pathway stimulation in this su ! group of rats could not be ex- plained on the basis of an alteration in 5-HT terminal autoreceptor sensitivity (Fig. 5).
these rats in whic R there had been no pharmacological
icantly decrease cp the efficacy of the stimulation o F-l the
ing treatment with 350 mgkg of f: CPA, the effect of
Hz) in re P ucing the efficacy of 5-HT synaptic transmis-
that i P 5-HT terminal autoreceptor desensitization were
1988; 8 haput et al., 1986a). In agreement with previous
r +I E v)
1 20 cn
0 CONTROL PCPA
350 rnglkg
0 1 Hz 5 Hz
Fig. 5. Degree of suppression of CA, dorsal hippocampus pyramidal neuron firing activity by stimulations at 1 and 5 Hz of the ascending 5-HT athway in control and in rats treated with PCPA for 2 days. The stimufat ion parameters were the same as those presented in legend to Figure 1. The number at the bottom of each pair of columns represents the number of neurons tested. *P < 0.05, **P < 0.005 compared to corresponding value at 1 Hz (paired Student’s t test).
To determine whether, in rats treated with PCPA (350 mgkg) in which the stimulation of the ascending 5-HT pathway resulted in a markedly reduced SIL value (8 ? 2 msec; n = 9), the efficacy of the stimulation could be augmented by the immediate 5-HT precursor 5-hydroxyt tophan (5-HTP), stimulations of the as-
following, the intravenous injection of 5-H P (combined with carbidopa, 1.5 mgkg, to prevent its peripheral decarboxylation). 5-HTP (5 mgkg i.v.) produced a two- fold increase of SIL values (Fi . 6), the maximal effect
In an attempt to determine whether a comparable increase in the SIL value could be obtained by applying 5-HT in the vicinity of the soma of CA, dorsal hippocam- pus yramidal neurons, stimulations of the ascending
the microiontophoretic application of 5-HT with an avera e ejection current of 55 nA, for an average dura- tion of255 sec (range 150-1000 sec), using iontophoretic solutions of 5-HT of 0.5 or 40 mM in 200 mM NaCl. No modification of the SIL value could be detected (prior to 7 i 1 msec; following 9 2 2 msec; n = 15).
Effect of 5,7-dihydroxytryptamine To compare the effect of the PCPA-induced depletion
of 5-HT to that of lesioning 5-HT neurons, the response of hippocampus pyramidal neurons to the microionto- phoretic a plication of 5-HT and to the stimulation of
following the intracerebroventricular administration of
.F cending 5-H ?l! pathway were carried out rior to, and
being observed about 20 min a fg ter the injection.
5-H 4 pathway were carried out prior to, and following,
the ascen B ing 5-HT pathway was assessed 10-14 days
0
PRIOR TO 5-HTP FOLLOWING 5-HTP (5 mg/kg, i.v.)
Fig. 6. Degree of suppression of the firing activity of CA, dorsal hi ocampus pyramidal neurons by the stimulation of the ascending 5% pathway prior to, and following, the intravenous administration of 5-hydroxyt to han (5-HTP) in rats treated for 2 days with PCPA (350 mg/kg/d3. !. timulation parameters were the same as those described in the legend to Fi re 1 The number at the bottom of the columns represents the numgr ofneurons tested. *P < 0.05 (paired Student’s t test).

334 Y. CHAPUT ET AL.
either 5,7-DHT7 a 5-HT neurotoxin, or saline. The 5,7- DHT pretreatment reduced the 5-HT content in the CA, region of the dorsal hippocampus by 90% (1 t 0.1 ng/mg protein; n = 10). As previously observed (de Montigny et al., 1980), there was no difference in the I * T,, value for the microionto horetic application of 5-HT between
indicating that the latter pretreatment had not induced a postsynaptic supersensitivity in this region of the brain. The efficacy of the stimulation of the ascending 5-HT pathway was reduced by 60% relative to control rats (Fig. 7).
DISCUSSION The present results showing that the 350 mg/kg/day
regimen of PCPA, but not the 250 mg/kg/day regimen, reduced significantly the efficacy of the stimulation of the ascending 5-HT pathway (Fig. 2) suggest that a greater than 90% depletion of the dorsal hippocampus 5-HT content is required to affect the 5-HT synaptic efficacy in this region of the brain. Interestingly, this conclusion is similar to that reached by Curet and de Montigny (1988) concerning the degree of depletion of the dorsal hippocampus NE content to produce a de- crease of the efficacy of the stimulation of the locus coeruleus. However, before considering the possible substratum for this apparent remarkable homeostatic capacity of the 5-HT synapse, several other possibilities must be considered.
First, the limited efficacy of PCPA in reducing the effect of the stimulation of the VMT on the firing activity of dorsal hippocampus pyramidal neurons raises the issue of the validity of this paradigm as an index of the
control and 5,7-D H T-pretreated rats (data not shown),
0
' 0 ° 1
0 I 0 0
0
AAA A
0 CONTROL 5, 7-DHT
Fi 7 Scattograms of the degree of suppression of the firing activity of C f 3 dorsal hi ocampus pyramidal neurons by the stimulation of the ascending5-% pathway in control rats and in rats pretreated with 5,7-DHT. Stimulation parameters were the same as those described in the legend to Figure 1. Each value represents the mean value for one rat, from which two to four neurons were tested. The shaded area represents the mean ? SEM value for the group. *P < 0.01 (one-way analysis of variance).
function of 5-HT synaptic transmission. That this para- digm could be unsuitable to assess 5-HT synaptic func- tion appears unlikely in light of the following electro- physiological observations: 1) The electrical stimulation the VMT antidromically activates 5-HT-containing neu- rons of the dorsal and median raphe nuclei, an effect blocked by a 5,7-DHT, but not a 6-hydroxydopamine, pretreatment into the VMT (Wang and Aghajanian, 197713,1978); 2) the latency of the effect of the stimula- tion on postsynaptic neurons is entirely compatible with the slow conduction velocity of small unmyelinated 5-HT axons and the distance from the tip of the stimu- lation electrode to the hippocampus (Blier and de Mon- tigny, 1983,1985; Chaput et al., 1986a); 3) the effect of the stimulation is markedly reduced by the systemic administration of a selective 5-HTlA receptor antago- nist (Chaput and de Montigny, 1988), the 5-HTIA recep- tor being in large part responsible for the suppressant effect of 5-HT in the CA1 and CA, regions of the rat hippocampus (Andrade and Nicoll, 1987; Beck, 1989; Chaput and de Montigny, 1988; Colino and Halliwell, 1987); 4) the terminal 5-HT autoreceptor blockade, which augments the stimulus-evoked release of F3H15- HT in vitro (Moret, 1985), enhances the efficacy of the stimulation in vivo (Blier et al., 1988; Chaput and de Montigny, 1988; Chaput et al., 1986a,b; present re- sults), and, to the contrary, terminal 5-HT autoreceptor activation, which has been shown to decrease 5-HT release in vivo (Martin and Marsden, 19861, decreases the SIL value (Chaput and de Montigny, 1988). Taken together, these data strongly suggest that the SIL value obtained with the VMT stimulation is a sensitive and accurate index of 5-HT synaptic transmission. That the effect of the stimulation was virtually abolished in more than one-half of the 5,7-DHT-treated rats (Fig. 7), in keeping with the report of Blier et al. (1987b), and its enhancement by small doses of 5-HTP in PCPA-treated animals (Fig. 6) support this contention. The inability of prolonged microiontophoretic applications of 5-HT onto CA, pyramidal neurons to restore the efficacy of the stimulation in PCPA-treated rats is most likely due to fact that most 5-HT terminals are found within the stratum radiatum (Azmitia and Segal, 1978), whereas electrophysiological recordings are obtained some dis- tance away, in the stratum pyramidale. Alternatively, that this failure might have been due to the activation, by microiontophoresed 5-HT7 of 5-HT terminal autore- ceptors would appear less likely in that prolonged mi- croiontophoretic applications of 5-HT does not alter the efficacy of the stimulation in control animals (unpub- lished observations).
A second possibility is that 5-HT depletion by PCPA resulted in postsynaptic 5-HT receptor supersensitivity, cancelling out the reduction of the synaptic efficacy expected to result from the depletion. Indeed, an in- crease in [3H15-HT specific binding to crude rat fore- brain membrane preparations has been reported to occur following short-term PCPA administration (Bennet and Snyder, 1976; Fleisher et al., 1979; Steigrad et al., 1978). In addition, Fleisher et al. (1979) have reported that PCPA, as well as its metabolite p-chlorophenyl- acetic acid, can increase the affinity of L3H]5-HT binding in vitro. However, this increased binding, as Steigrad et al. (1978) proposed, is more likely the result of a decreased 5-HT content leading to a reduced competi-

EFFECT OF PCPA ON 5-HT NEUROTRANSMISSION 335
tion for the binding sites rather than of an increased number of receptors. This is supported by the finding of a normal sensitivity of postsynaptic 5-HT receptors following PCPA in both behavioral (Dourish et al., 1986; Stewart et al., 1976; Trulson et al., 1976) and electro- physiological paradigms (Ferron et al., 1982; Wang et al., 1979; present study).
A third possibility for interpreting the rather limited efficacy of PCPA in reducing the effect of the VMT stimulation despite a marked 5-HT depletion may per- tain to either the coexistence of another neurotransmit- ter within 5-HT-containing neurons or the existence of non-5-HT fibers emanating from both the dorsal and median raphe nuclei. Indeed, it has been suggested that nond-HT neurons [possibly catecholaminergic or pepti- dergic (Steinbush and Nieuwenhuys, 1983)l in the dorsal and median raphe nuclei contribute to their ascending fiber tracts, including those afferent to the hippocampus (Kohler and Steinbush, 1982; Kohler et al., 1982; Kosofsky and Molliver, 1987; O’Hearn and Molliver, 1984). Therefore, the possibility that the stim- ulation of the ascending 5-HT tracts within the VMT might also activate nond-HT axons cannot be excluded. On the other hand, coexistence has been most exten- sively investigated in the rat medullary raphe nuclei projecting to the spinal cord, where substance P (SP) and thyrotropin-releasing hormone (TRH) immunore- activity have been detected in 5-HT-containing soma (for review, see Hokfelt et al., 1987). Prior treatment of cats with PCPA has been shown to enhance markedly the excitatory effect of the microiontophoretic applica- tion of SP and as well as that of 5-HT onto dorsal horn neurons, without altering SP content of the spinal cord (Jeftinija et al., 1986). In the rat, an increase in SP content of the midbrain raphe nuclei has been reported following short-term PCPA administration (Savard et al., 1984), although in these latter regions SP immu- nostaining may reflect terminal projections of SP-con- taining neurons originating from outside the raphe (Hokfelt et al., 1987; Neckers et al., 1979). Both the midbrain raphe nuclei and the hippocampus of the rat have been shown by radioimmunoassay to contain mod- erate amounts of TRH (Lighton et al., 1984). Further- more, TRH recognition sites have been reported in the hip ocampus (Funatsu et a]., 19851, and microinjection
PCPA-induced muricidal behavior in rats (Pucilowski et al., 1988). However, lesioning with 5,7-DHT mark- edly reduced the 5-HT, but not TRH, content of the median and dorsal raphe nuclei (Lighton et al., 1984). Given both in vivo and in vitro evidence in the rat spinal cord that TRH exerts a postsynaptic effect similar to that of 5-HT and that SP enhances 5-HT release via terminal 5-HT autoreceptor blockade (Barbeau and Be- dard, 1981; Mitchell and Fleetwood-Walker, 1981; Hansen et al., 1983), it could be speculated that an analogous situation exists in the hippocampus account- ing for the maintenance of 5-HT transmission despite severe 5-HT depletion. However, such an interpretation would ap ear premature given the paucity of evidence
projections of the midbrain raphe nuclei to the hippo- campus.
Although the uptake of L3H]5-HT rat hypothalamic synaptosomes does not appear to be altered by pretreat-
of f RH into this region has been shown to inhibit
for the co Y ocalization of either neuropeptide within the
ment of rats with PCPA (Pazdernik and Tuomisto, 19831, McBride et al. (1976) have reported a marked increase in the specific activity of 5-hydroxyindole ace- tic acid (5-HIAA) over that of 5-HT following D,Ld-HTP administration in PCPA-pretreated rats, suggesting an increased release of 5-HT. Galzin et al. (1985), using rat hypothalamic slices, have shown that the electrically evoked release of [3H]5-HT is enhanced by the short- term treatment of rats with PCPA. Furthermore, in this latter study, the efficacy of MET in enhancing, but not that of terminal 5-HT autoreceptor agonists in decreas- ing, the stimulus-evoked release of L3H15-HT was re- duced, suggesting that the enhanced release of L3H15- HT was due to a lesser degree of activation of the terminal 5-HT autoreceptor. Consistent with these data, in the present study, the effectiveness of MET in enhancing the efficacy of the VMT stimulation in rats treated with PCPA was reduced, whereas there was no reduction in the effectiveness of increasing the fre- quency of the stimulations from 1 to 5 Hz. These data suggest that the terminal 5-HT autoreceptor is acti- vated to a lesser degree in rats pretreated with PCPA and that this degree of activation can be readily in- creased by reducing the time interval between stimula- tion-trig ered action potentials. Therefore, the possibil-
R-PCPA animals derives from a lower level of activation of the 5-HT terminal autoreceptors by endogenous 5-HT as a result of neurotransmitter de letion within 5-HT
mechanism might explain, a t least partly, the normal response to afferent 5-HT pathway stimulation of most pyramidal neurons in rats treated with the low dose (250 mg/kg/day) of PCPA. Indeed, a lower level of tonic activation of 5-HT autoreceptors prior to the beginning of the stimulation would allow a relatively greater release of neurotransmitter per stimulation pulse.
Numerous methodological differences (stimulation sites; pulse duration, intensity, and frequency, pooling of the responses of CA, and CAB neurons) exist between the present study and that of Segal (19751, which ren- ders a comparison of the effects of PCPA treatment on 5-HT synaptic transmission difficult. In the latter study, the responses of pyramidal neurons to dorsal or median raphe stimulation were much more heteroge- neous than those in the present study. However, the short-latency inhibition of pyramidal neurons predomi- nantly observed by Segal(1975) with the median raphe stimulation closely resembles the effect studied here in both its temporal characteristics (latency, duration) and its resistance to PCPA treatment. In contrast, Wang and Aghajanian (1977a) reported that, in the rat, the inhibitory response of amygdaloid neurons to electrical stimulation of the dorsal raphe nucleus in vivo was reduced by 75% by a single dose of PCPA (400 mgkg). A possible explanation for the discrepancy between the effectiveness of PCPA in the latter and present study may pertain to differences in the degree of 5-HT deple- tion obtained between discrete regions of the rat CNS (Fleisher et al., 1979; Reader and Gauthier, 1984; Val- zelli et al., 1983).
The present observation that the 5,7-DHT pretreat- ment, but not the regimen of 250 mg/kg/day of PCPA, significantly reduced the efficacy of the stimulation of the ascending 5-HT pathway (Figs. 2 and 71, despite the
ity that t a e unaltered efficacy of the stimulations in the
terminals must be entertained. 8 uch a homeostatic

336 Y. CHAPUT ET AL.
fact that both produced a similar (90%) depletion of the 5-HT content in the dorsal hippocampus. It might be speculated that the PCPA treatment most likely de- creases the 5-HT content per terminal, whereas the 5,7-DHT neurotoxine reduces the number of terminals but presumably not the 5-HT content of the few remain- ing terminals. If this is the case, the mechanism whereby the 5-HT synapse might maintain its homeo- stasis in PCPA-treated rats (i.e., a reduced activation of terminal 5-HT autorece tors) might not be operative in 5,7-DHT-treated rats. T K is would account for the differ- ential effect of the PCPA and the 5,7-DHT pretreat- ments on the efficacy of the stimulation of the ascending 5-HT pathway.
The present results demonstrate that the efficacy of 5-HT synaptic transmission is significantly reduced following short-term administration of a high dose of PCPA in the rat. The rapid reversal of pharmacological remission by PCPA, at a dose of approximately 40 mgkg, in patients treated with various antidepressants (Shopsin et al., 1975, 19761, and the rapid mood-low- ering effect of decreasing plasma tryptophan in de- pressed patients in pharmacological remission (Delgado et al., 1990) suggest a role for 5-HT in mediating the therapeutic effect of antidepressant drugs in major affective disorders. However, there is an apparent dis- crepancy between the dose of PCPA required in the present study to affect 5-HT neurotransmission in the rat and that inducing a relapse in pharmacologically remitted depressed patients. Obviously, a s ecies differ- ence might well exist. Nonetheless, in lig R t of the en- hancement of 5-HT neurotransmission by various types of antidepressant treatments in the rat (see Blier et al., 1987a; Willner, 19851, it would be most interesting to investigate the possibility that the 5-HT system is much more sensitive to the disruptive effect of PCPA following an antidepressant treatment as a result of an alteration of the properties of the 5-HT synapse.
ACKNOWLEDGMENTS This work was supported in part by the Canadian
Medical Research Council (MRC) grant MT-6444 (to C.de M.) and by a Mental Health Research Network Grant from the Fonds de la Recherche en Sante du Quebec. Y.C. is in receipt of an MRC Fellowship. We thank M. Boulay for his technical assistance. L. Caille for writing the programs, G. Filosi for the illustrations, and T. Romano for her secretarial assistance.
REFERENCES Aghajanian, G.K. (1978) Feedback regulation of central monoaminer-
gic neurons: evidence from single-cell recording studies. In: Essays in Neurochemistry and Neuropharmacology, Vol3. M.B.H. Youdim, W. Lovenberg, J.R. Lagnado, and D.F. Sharman, eds. John Wiley, New York, pp. 1-32.
Andrade, R., and Nicoll, R.A. (1987) Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro. J . Physiol., 394:99-124.
Asberg, M., Ericksson, B., Matensson, B., Traskmen-Bendz, L., and Wagner, A. (1986) Therapeutic effects of serotonin uptake inhibitors in depression. J. Clin. Psychiatry, 47:23-35.
Ayuso Guttirez, J.L., and Lopez-Ibor, A.J.J. (1971) Tryptophan and a MA01 (nialamide) in the treatment of major depression. A double- blind study. Int. J. Pharmacopsychiatry, 7:92-97.
Azmitia, E.C., and Segal, M. (1978) An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. J. ComD. Neurol.. 179:641-667.
Barbeau, H., and Bedard, P. (1981) Similar motor effects of 5-HT and TRH in rats following chronic spinal transection and 5,7-dihydroxy- tryptamine injection. Neuropharmacology, 20:477481.
Baumann, P.A., and Waldmeier, P.C. (1984) Negative feedback control of serotonin release in vivo: comparison of 5-hydroxyindolacetic acid levels measured by voltammetry in conscious rats and by biochemi- cal techniques. Neuroscience, 11:195-204.
Bech, P. (1988) A review of the antide ressant properties of serotonin reuptake inhibitors. Adv. Biol. PsycEiatry, 17:58-69.
Beck, S.G. (1989) 5-carboxyamidotryptamine mimics only the 5-HT elicited hyperpolarization of hippocampal pyramidal neurons via 5-HTIA receptor. Neurosci. Lett., 99:lOl-106.
Bennet, J.P., and Snyder, S.H. (1976) Serotonin and lysergic acid diethylamide binding in rat brain membranes: relationship to postsynaptic serotonin receptors. Mol. Pharmacol., 12:373-389.
Blier, P., Chaput, Y., and de Montigny, C. (1988) Long-term 5-HT reuptake blockade, but not monoamine oxidase inhibition, decreases the function of terminal 5-HT autoreceptors: An electrophysiological study in the rat brain. Naunyn-Schmiedebergs Arch. Pharmacol., 337:246-254.
Blier, P., and de Montigny, C. (1983) Eledrophysiological investiga- tions of the effect of repeated zimelidine administration on seroton- ergic neurotransmission in the rat. J. Neurosci., 3:127&1278.
Blier, P., and de Montigny, C. (1985) Short-term lithium administra- tion enhances serotonergic neurotransmission: eledrophysiological evidence in the rat CNS. Eur. J. Pharmacol., 113:69-77.
Blier, P., de Montigny, C., and Chaput, Y. (1987a) Modifications of the serotonin system by antidepressant treatments. Implications for the therapeutic response in major depression. J. Clin. Psychopharma- col., 6:24%35S.
Blier, P.. de Montigny, C., and Tardif, 1). i1987b1 Short-term lithium treatment enhances res onsiveness of postsynaptic 5-HT,, recep- tors without altering 5-8T autoreceptor sensitivity: an electrophys- iological study in the rat. Synapse, 1225-232.
Cha ut, Y , Blier, P., and de Montigny, C. (1986ar In vivo electrophys- iotgical evidence for the regulatory role of autoreceptors on seroton- erac terminals. J. Neurosci., 6:279W2801.
Chaput, Y., and de Montigny, C . (1988) Effects of the 5-HT, receptor antagonist, BMY 7378, on 5-HT neurotransmission: Electrophysio- logwal studies in the rat CNS. J. Pharmacol. Exp. Ther., 246:359- 370.
y, C , and Blier, P. 11986bi Effect ofa selective 5-RTreu take blocf%,citalo ram, on the sensitivityof5-IITautore- ce tors [lectro hysiologicaf studies in the rat brain. Saunyn- Scimiedebergs L c h . Pharmacol., 333:342-348.
Chiew, C.C., Zukowska-Grojec, Z., Kirk, K.L., and Kopin, I.J. (1982) 6-Fluorocathecolamines as false adrenergic neurotransmitters. J. Pharaarol. Ex/. Ther., 225529433,
C o h o . A,, and I alliwell, J.V 11987) Differential modulation of three
Cha ut, Y,, de Monti
separate K+-conductances in hippocampal CA, neurons by seroto- nin. Nature, 328:73-77.
Coppen, A,, Shaw, D.M., and Farrell, J.P. (1963) Potentiation of the antidepressive effects of a monoamine oxidase inhibitor by tryp- tophan. Lancet 1:79-81.
Cyret, O., and de Montigny, C. (1988) Electrophysiological character- ization to adrenoceptors in the rat dorsal hipgocampus. 11. Receitors mediating the effect of synaptically release norepinephrine. rain Res., 475:47-57.
Delgado, P., Charne , D.S., Price, L.H., Aghajanian, G.K., Landis H., and Heninger, G . 2 (1990) Serotonin function and the mechanism of antidepressant action: reversal of antidepressant-induced remission by rapid depletion of plasma tryptophan. Arch. Gen. Psychiatry, 47:411418.
de Montigny, C., and Aghajanian, G.K., (1977) Preferential action of 5-methoxytrypam~ne and 5-methoxvdimethvltrv~tamine on Dresvn- aptic serotonh rece tors: A comparative iont6phoretic study with LSD and serotonin. keuro harmacology, 162311-818.
deMontigny, C., Cournoyer, E., Morissette, R., Lanflois, R,., andcaille, G. (1983) Lithium carbonate addition in tricyc ic antidepressant- resistant uni olar depression. Arch. Gen. Psychiatry, 40:1327-1334.
de Montigny, 8., El-, .R.? Caille, G. (1985). Rapid response to the addition of lithium in iprindole-resistant unipolar depression: a pilot study. Am. J. Ps chiatry, 142:22@-223.
de Montigny, C., dmnberg, F., Mayer, A,, and Deschenes, J.P. (1981) Lithium induces rapid relief of de ression on tricyclic antidepressant drugnon-res onders. Br. J. Ps cgiatry, 138:252-256.
de Monti y, e., Wang, R.Y., Jeader, T.A., and Aghajanian, G.K. (1980) Enoaminergc denervation of the rat hippocampus: Micro- iontophoretic studies on pre- and postsynaptic supersensitivity to nore ine hrine and serotonin. Brain Res., 200:363-376.
Dourisg, C%., Hutson, P.H., and Curzon, G. (1986) Para-chlorophenyl-

EFFECT OF PCPA ON 5-HT NEUROTRANSMISSION 337
alanine prevents feeding induced by the serotonin agonist 8-hy- droxy-2-(di-n-propylamino)tetralin (8-OH-DPAT). Psychopharma- cology, 89:467471.
Ferron, A., Descarries, L., and Reader, T. (1982) Altered neuronal responsiveness to biogenic amines in rat cerebral cortex after seroto- nin denervation or depletion. Brain Res., 231:93-108.
Fleisher, L.N., Simon. J.R., and Aurison. M.H. (1979) A biochemical-
Neckers, L.M., Schwartz, J.P., Wyatt, R.J., and Speciale, S.G. (1979) Substance P afferents from the habenula innervate the dorsal raphe nucleus. Exp. Brain Res., 37:619-623.
Nelson, C.J., and Byck, R. (1982) Rapid response to lithium in phenelzine non-responders. Br. J. Psychiatry, 141:85-86.
Nelson, J.C., and Mazure, C.M. (1986) Lithium augmentation in usvchotic deuression refractorv to combined drua treatment. Am. J k&chiatrv. i43:363-366. "
- behavioral model for stud$ng serotonergic supersensitivity in brain. J. Neurochem., 32:1613-1619.
Funatsu, K., Teshima, S., and Inanaga, K. (1985) Various types of thyrotropin releasing hormone receptors in discrete brain regions and the pituitary of the rat. J. Neurochem., 45:390-397.
Galzin, A.M., Moret, C., and Langer, S.Z. (1985) Interaction between tri clic and non tricyclic 5-hydroxytryptamine uptake inhibitors anythe presynaptic 5-hydroxytryptamine inhibitory autoreceptors in the rat hypothalamus. J. Pharmacol. Exp. Ther., 235200-211.
Glassman, A., and Platman, S.R. (1969) Potentiation of a monoamine oxidase inhibitor by tryptophan. J. Psychiatr. Res., 7:83-89.
Hansen, S., Svensson, L., Hokfelt, T., and Everitt, B.J. (1983) 5- Hydroxytryptamine-thyrotropin releasing hormone interactions in the spinal cord: effects on parameters of sexual behavior in the male rat. Neurosci. Lett., 42:299-304.
Heninger, G.R., Charney, D.S., and Sternberg, D.E. (1983) Lithium carbonate augmentation of antidepressant treatments. Arch. Gen. Psychiatry, 40:1335-1342.
Hokfelt, T., Johansson, O., Holets, V., Meister, B., and Melander, T. (1987) Distribution of neuropeptides with special reference to their coexistence with classical transmitters. In Psychopharmacology: The Third Generation of Progress. H.Y. Meltzer, ed. Raven Press, New York, pp. 401416.
Jeftinija, S., Raspantini, C., Randic, M., Yaksh, T.L., Go, V.L.W., and Larson, A.A. (1986) Altered responsiveness to substance P and 5-hydroxytryptamine in cat dorsal horn neurons after 5-HT deple- tion with p-chlorophenylalanine. Brain Res., 368:107-115.
Jequier, E., Lovenberg, W., and Sjoerdsma, A. (1967) Tryptophan hydroxylase inhibition: The mechanism by which chlorophenylala- nine depletes rat brain serotonin. Mol. Pharmacof-, 3:274-278.
Kandel. E.R.. and Suencer. W.A. (1961) Electrouhvsiolom of hiDDo-
O'HLarn, E.; and Molliver, M.E. (1984) Organization of raphe-cortical projections in the rat: A quantitative retrograde study. Brain Res. Bull., 13:709-726.
Pare, C.M.B. (1963) Potentiation of monoamine oxidase inhibitors by tryptophan. Lancet, 2:527-528.
Pazdernik, T.L., and Tuomisto, J. (1983) Do amine and 5-HT uptake by synaptosomes after pretreatment wit! p-chlorophenylalanine. Med. Biol., 61:264-267.
Pucilowski, O., Trzaskowska, E., Kotostowski, W., Jankowska, E., and K u p y w s k i , G. (1988) Inhibition ofg-chlorophenylalanine-in- duce muricide behanor following TR microinjection into the limbic structures in the rat. J. Neural Transmiss., 74:175-180.
Reader, T.A., and Gauthier, P. (1984) Catecholamines and serotonin in the rat central nervous system after 6-OHDA and PCPA. J . Neural Transmiss., 59:207-227.
Ryan, N.D., Meyer, V., Dochille, S., Mazzie, D., and Puig-Antich, J . (1988) Lithium antidepressant augmentation in TCA-refractory de- pression in adolescents. J . Am. Acad. Child Adolesc. Psychiatry, 27:371-376.
Savard, P., Merand, Y., and Dupont, A. (1984) The effects of p- chlorophenylalanine (PCPA) treatment on the substance P content measured in discrete brain nuclei of normal and neonatally-induced hypothyroidrats. Brain Res., 292:349-355.
Segal, M. (1975) Physiological and pharmacological evidence for a serotonergic projection to the hippocampus. Brain Res., 94:115-131.
Sho sin, B., Friedman, E., and Gershon, S. (1976) Para-chloropheny- lapanine reversal of tranylcypromine effects in depressed patients. Arch. Gen. Psychiatry, 33:811-819.
Shopsin, B., Gershon, M., Goldstein, M., Friedman, E., and Wilk, S. (1975) Use of svnthesis inhibitors in defining a role for bioeenic
campal neurons. 11. After-potentials and repeiitcve firGg. Psycho- pharmacology, 83:20-27. chopharmacol. Commun., 1:239-249.
Keneko, M., Kumashiro, H., Takahashi, Y., and Hoshino, Y. (1979) L-5-HTP treatment of and serum 5-HT level after L-5-HTP loading in depressed patients. Neuropsychobiology, 5:232-240.
Koe, B.E., and Weissman, A. (1966) Parachlorophenylalanine: a spe- cific deuletor of brain serotonin. J . Pharmacol. Exu. Ther.. 154:
amines during imipramine treatment in depFessed patients.-Psy-
Steigrad, P., Tobler, I., Waser, P.J., and Borbely, A.A. (1978) Effect of p-chlorophenylalanine on cerebral serotonin binding, serotonin con- centration and motor activity in the rat. Naunyn-Schmiedebergs Arch. Pharmacol., 305143-148.
Steinbush. H.W.M.. and Nieuwenhuvs. R. (1983) The rauhe nuclei of 499-5 16.
Kohler, C., and Steinbush, H.W.M. (1982) Identification of serotonin and non-serotonin containing neurons of the midbrain raphe project- ing to the entorhinal area and hippocampal formation. A combined immunohistochemical and fluorescent retrograde tracing study in the rat brain. Neuroscience, 7:951-975.
Kohler, C., Chan-Palay, V., and Steinbush, H.W.M. (1982) The distri- bution and origin of serotonin-containing fibers in the septa1 area: A combined immunohistochemical and fluorescent retrograde tracing study in the rat. J. Comp. Neurol., 209:91-111.
Kosofsky, B.E., and Molliver, M.E. (1987) The serotoninergic innerva- tion of the cerebral cortex: Different classes of axon terminals arise from dorsal and median raphe nuclei. Synapse, 1:153-168.
Lighton, C., Marsden, C.A., and Bennett, G.W. (1984) The effects of 5,7-dihydroxytryptamine and p-chlorophenylalanine on th otro pin-releasing hormone in regions of the brain and spinal corgf the rat. Neuropharmacology, 23:55-60.
Lowry, O.H., Rosebrough, M.J., Farr, A.L., and Randall, R.J. (1951) Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265-275.
Martin, K.F., and Marsden, C.A. (1986) In vivo voltammetry in the suprachiasmatic nucleus of the rat: effects of RU 24969, me- thiothepin and ketanserin. Eur. J. Pharmacol., 121335-140.
McBride, W.J., Penn, P.E., Hyde, T.P., Lane, J.D., and Smith, J.E. (1976) Effects of p-chlorophenylalanine on the metabolism of seroto- nin from 5-hydroxytrypto han Neurochem. Res., 1:437-449.
Mitchell, R., and FleetwoolWaiker, S. (1981) Substance P, but not TRH, modulates the 5-HT autoreceptor in ventral lumbar spinal cord. Eur. J . Pharmacol., 76:119-120.
Moller, S.E., Kirk, L., and Honore, P. (1980) Relationship between plasma ratio of tryptophan to competing amino acids and the re- sponse to L-tryptophan treatment in endogenously-depressed pa- tients. J . Affect. Disorders, 2:47-59.
Moret, C. (1985) Pharmacology of the serotonin autoreceptor. In: Neuropharmacology of Serotonin. A.R. Green, ed. Oxford University Press, Oxford, pp. 21-49.
the rat brainstem: A cvtoarchitekonic and immunohtochemical study. In: Chemical NeGroanatomy. Emson, C.P., ed. Raven Press, New York, pp. 131-207.
Stewart,R.M., Growdon, J.H., Cancian, D., andBaldessarini, R. (1976) 5-Hydroxytryptophan-induced myoclonus: Increased sensitivity to serotonin after intracranial 5.7-dihydroxytrvptamine in the adult - - _ rat. Neuro harmacology, 15:449-455.
Trulson, M.E., Eubanks, E.E., and Jacobs, B.L. (1976) Behavioral evidence for suuersensitivitv followine destruction of central sero- toner 'c nerve tkninals by 5,7-dihydr&ytryptamine. J . Pharmacol. Exp. Fher. 198:23-32.
Valzelli, L., Bernasconi, S., and Dalessandro, M. (1983) Time-course of PCPA-induced depletion of brain serotonin and muricidial aggres- sion in the rat. Pharmacol. Res. Comm., 15:387-395.
Van Praag, H.M. (1981) Management of depression with serotonin precursors. Biol. Psychiatry, 16:291-310.
Whlinder, J., Skott, A,, Carlsson, A. Nagy, A., and Roos, B.-E. (1976) Potentiation to the antidepressant action of clomipramine by tryp- tophan. Arch. Gen. Psychiatry, 33:1384-1389.
Wang, R.Y., and Aghajanian, G. (1977a) Inhibition of neurons in the amygdala by dorsal raphe stimulation: mediation through a direct serotonergic pathway. Brain Res., 120:85-102.
Wang, R.Y., and Aghajanian, G. (1977b) Antidromically identified serotonergic neurons in the rat midbrain raphe: evidence for collat- eral inhibition. Brain Res., 132:186-193.
Wang, R.Y., and Aghajanian, G. (1978) Collateral inhibition of seroton- ergx neurones in the rat dorsal raphe nucleus: Pharmacological evidence. Neuropharmacology, 17:819-825.
Wang, R.Y., de Montigny, C., Gold, B.I., Roth, R.H., and Aghajanian, G.K. (1979) Denervation supersensitivity to serotonin in rat fore- brain: single cell studies. Brain Res., 178:479-497.
Willner, P. (1985) Antidepressants and serotonergic neurotransmis- sion: A n integrative review. Psychopharmacology, 85387404.




![Glutamatergic neurotransmission and synaptic plasticity: molecular, clinical, and phylogenetic aspects [Portuguese]](https://img.pdfslide.net/doc/110x75/634c6757738f1906320e170a/glutamatergic-neurotransmission-and-synaptic-plasticity-molecular-clinical-and.jpg)
























