Embed Size (px)
Citation preview

Primary Research Paper
Effects of the sand bar breaching on Typha domingensis (PERS.) in a tropical
coastal lagoon
Anderson M. dos Santos1,2, Andre M. Amado1, Murilo Minello1, Vinicius F. Farjalla1,*& Francisco A. Esteves11Laboratorio de Limnologia, Depto. de Ecologia, Instituto de Biologia, Universidade Federal do Rio de Janeiro.Av. Brigadeiro Trompowski s/n, Predio CCS, bloco A, sub-solo, sala A0-008 – Ilha do Fundao, P.O. Box 6802, 21941-590,
Rio de Janeiro RJ, Brazil2Instituto de Ciencias Biologicas, BL I-3, sala 163, Universidade Federal de Minas Gerais, Av. Antonio Carlos,6627 – Pampulha, P.O. Box 486, 31270-901, Belo Horizonte, Brazil
(*Author for correspondence: E-mail: [email protected])
Received 26 November 2004; in revised form 13 June 2005; accepted 24 June 2005
Key words: coastal lagoon, aquatic macrophytes, ecosystem management, eutrophication
Abstract
Coastal lagoons are usually subjected to several kinds of human impacts, especially eutrophication. Thebreaching of the sand bar, which separates the lagoon from the ocean, by human action, is a commonprocess used to decrease the negative effects of eutrophication. The aims of this research were to evaluatethe effects of the artificial sand bar breaching on the populations of the aquatic macrophyte Typhadomingensis and the subsequent effects on nutrients concentration in a tropical coastal lagoon. Samplingswere carried out monthly from February/01 to January/02 in a monospecific stand of T. domingensis atImboassica lagoon (Rio de Janeiro/Brazil). Two sampling sites, in the middle and in the border of thestand, were marked in three different transects. Water depth was measured and the aerial biomass sampledwith a 0.25 m2 quadrat. The macrophyte samples were separated into live and dead material and the shootlength, shoot density and number of leaves were analyzed. All plant material was oven dried till constantweight and net primary production, dead stand crop production and the nutrients release throughdecomposition were estimated at each site. The decrease in water level due to sand bar opening affectednegatively T. domingensis populations, but the most intense effects were observed in the middle of the stand.The shoot mortality was highly enhanced after the sand bar breaching and the nutrients were releasedthrough decomposition to the water column. It reflected on an input of 11.5 kg C m)2, 0.22 kg N m)2 and0.13 kg P m)2 into the lagoon, which represent from 22.5 to 44.8 ton P and 35.9 to 71.8 ton N to thelagoon. The decrease of nutrients concentration after the sand bar breaching was not successfullyaccomplished. The decay of T. domingensis stands due to the sand bar breaching neutralized the expor-tation of nutrients to the ocean, and contributed to the phosphorous increase in the water column. Thus, toa better management of aquatic ecosystems subjected to human eutrophication, the role of aquatic mac-rophytes decomposition on internal fertilization of aquatic ecosystems should be accounted.
Introduction
Tropical coastal lagoons are usually shallow eco-systems characterized by a high perimeter/volume
ratio and broad littoral areas. These ecosystemsshow high primary productivity mainly related tothe aquatic macrophytes community (Knoopers,1994). Aquatic macrophytes also participate in the
Hydrobiologia (2006) 556:61–68 � Springer 2006DOI 10.1007/s10750-005-1084-6

nutrient cycling, pumping nutrients from sedimentto water column or releasing nutrients during thedecomposition process (Wetzel, 1969). In this way,aquatic macrophytes play an important role tonutrient cycling in coastal lagoons.
Coastal lagoons are usually subjected to humanimpacts once coastal areas are the most denselyhuman-populated areas in the world (Vitousek &Mooney, 1997). Domestic and industrial sewagedisposal, embankment of the margins, introduc-tion of non-indigenous species and the artificialopening of the sand bar are the main examples ofhuman interference in those ecosystems (Esteves,1998). The sand bar breaching consists in theartificial removal of part of the sand bar, whichisolates the lagoon from the ocean. This processpromotes temporary communication and waterchange between these two aquatic ecosystems. It isusually observed the decrease on water level andnutrient concentration (exported to the sea) as faras changes in biological communities (Suzuki,1995; Branco et al., 1998; Panosso & Esteves,2000; Santos & Esteves, 2002; Amado et al., 2005).
The aims of this research were to evaluate theeffects of abrupt water level variation on Typhadomingensis aquatic macrophyte populations andthe subsequent effects to nutrients concentration ina tropical coastal lagoon.
Study area
This study was performed at Imboassica lagoon, atropical coastal lagoon Rio de Janeiro state/Brazil(area of 3.26 km2), isolated from the ocean by anarrow sand bar, located in the north of Rio deJaneiro State (22� 30¢ S; 42� 00¢ W). The regionalclimate is warm-humid, and the mean annualtemperature varies between 18 and 27 �C. Meanrelative humidity is 83%, and mean annual rainfallis 1300 mmwith well-defined dry and rainy seasons(Fiderj, 1977). This ecosystem is densely colonizedmainly by emergent aquatic macrophytes(T. domingensis and Eleocharis mutata) and even-tually by submerged species (Chara sp.) (Palma-Silva et al., 2002). We estimate that T. domingensiscolonizes about 5–10% of Imboassica lagoon totalarea.
Imboassica lagoon receives continuouslynon-treated domestic sewage disposal (from
neighborhood houses). In February, 8th, 2001, theMacae City Hall, intending to decrease the eco-logical and sanitary degradation, managed a sandbar breaching event. The sand bar remove pro-moted partial water change between the lagoonand the ocean for about 2 weeks, when it wasnaturally replaced.
Methodology
Sampling and biometric parameters
Samplings were performed monthly fromFebruary/01 to January/02. Typha domingensiswere sampled in a large stand, in three paralleltransects from the margin to inside the lagoon.Two sampling sites, in the central part (middle)and in the border of the stand, were marked ineach transect performing six sampling sites. Thewater depth and the aerial biomass (with a 0.25 m2
quadrat) were sampled in each site. The shootswere immediately separated into live (more than50% of green tissue) and dead (less than 50% ofgreen tissue) material. The living shoots had thelength and density measured at each sampling site.All samples (live and dead leaves) were oven dried(around 60 �C) till reaching constant weight andthe biomass was estimated by gravimetric scale(0.01 precision).
Necromass production
Necromass production was calculated as a modi-fication of the Symbula and Day (1988) method,which was originally designed to estimate below-ground production:
ðai � ai�1Þ þ ðai�1ktiÞ if ai � ai�1 > 0;
ðai�1 � aiÞ þ ðai�1ktiÞ if ai � ai�1 < 0;
where ai)1 corresponds to dead material quantityat the start of the interval, ai is the total amount ofdead material remaining at the end of the inter-val ti and k is the decomposition rate. We used therate of 0.17% D.W. dead stand crop d)1 as thedetritus decomposition rate of T. domingensis(Goncalves-Junior et al., 2004) to calculate the
62

dead stand crop exported to the water columnduring the sampling period.
Thus, the nutrient release dynamics was esti-mated during the sampling period based on carbon(C), nitrogen (N) and phosphorous (P) contents inT. domingensis recorded by Furtado (1998) inImboassica lagoon (C = 41.0% D.W.; N = 0.8%D.W.; P = 0.5% D.W.). Total N and P in thewater were measured monthly in a central point inthe lagoon by Mackereth et al. (1978) andGolterman et al. (1978), respectively (data fromBrazilian Long Term Ecological ResearchProgram – LTER Site 5).
The macrophyte net primary production (NPP)was estimated according to Junk and Piedade(1993). This method considers biomass losses of 10and 25% between sampling periods due todecomposition, mortality and herbivory. In thepresent research we adopted the decompositionrate of 10% once the 25% decomposition rate mayoverestimates the net annual primary productionin about 80% (Santos, 2004). The NPP calcula-tions follow as:
NPP=NPP
Z to
0
dBþ p:B dt if dB > 0;
NPP=NPP
Z to
0
p:B dt if dB < 0;
where NPP is the net primary productivity increase(t); p, the decomposition rate (10%); B(t), thebiomass as a function of time.
Statistical analysis
The ANOVA test was performed to comparebiomass, dead stand crop, leaves density, shootsdensity and shoot length between time and sam-pling sites. In case of normality, homocedasticityand independency assumptions violation, theStandard tests routine and the Monte-CarloANOVA tests were used to test the followinghypothesis: the mean variation in each group ishigher than the casualty (Gotelli & Entsminger,2001). This routine randomizes data betweengroups and estimates the expected variation be-tween the different data groups means. The meansare similar under the null hypothesis. These anal-yses were performed using the software ECOSIM,version 7.22.
Finally, Spearman correlation was used toevaluate water lever fluctuation on biomass, deadstand crop, leaves density, shoots density andlength and dead stand crop production.Correlations were performed using the softwareSTATISTICA v5.5 for Windows StatSoft, Inc.(2000).
Results
The sampling sites were completely dried after thesand bar breaching. After 1 year, the water col-umn was reestablished at Imboassica lagoon,reaching 0.9 and 1.0 m in the middle and in theborder of the stand, respectively (Fig. 1a). Thedepth in each sampling site was considered statis-tically different (F(Monte-Carlo) = 14.08; p<0.001).
Biometric parameters
Even considering the abrupt variation caused bythe sand bar breaching (considering the two sam-pling sites together), the water level depth was posi-tively correlated to leaf density (r-Spearman = 0.67;p<0.001), shoots density (r-Spearman = 0.48;p<0.05), and NPP (r-Spearman = 0.85;p<0.001), and negatively correlated to the deadstand crop (r-Spearman = )0.60; p<0.01).
The shoots length decreased gradually dueto the sand bar breaching in both sites, but thedecrease was more evident in the middle of thestand (Fig. 1b). During the reestablishing of watercolumn, the shoots grew up reaching 1.3 and 1.4 m,respectively in the border and in the middle ofthe stand, at the end of the sampling period. Theshoots length were considered similar between thesampling sites (F(Monte-Carlo) = 1.18; p = 0.28).
The leaves density was significantly higher(F(Monte-Carlo) = 53.2; p<0.001) in the border ofthe stand when compared to the middle (Fig. 1c).Meanwhile, the greater density in the middle wasrecorded before the sand bar breaching. The samepattern was observed to the shoots density beingdifferent between sites (F(Monte-Carlo) = 13.02;p<0.001) (Fig. 1d).
The sand bar breaching resulted in a decreaseof T. domingensis biomass in both samplings sites(Fig. 1e). The highest biomass was recorded in themiddle of the stand (1,457.9 g D.W. m)2) before
63
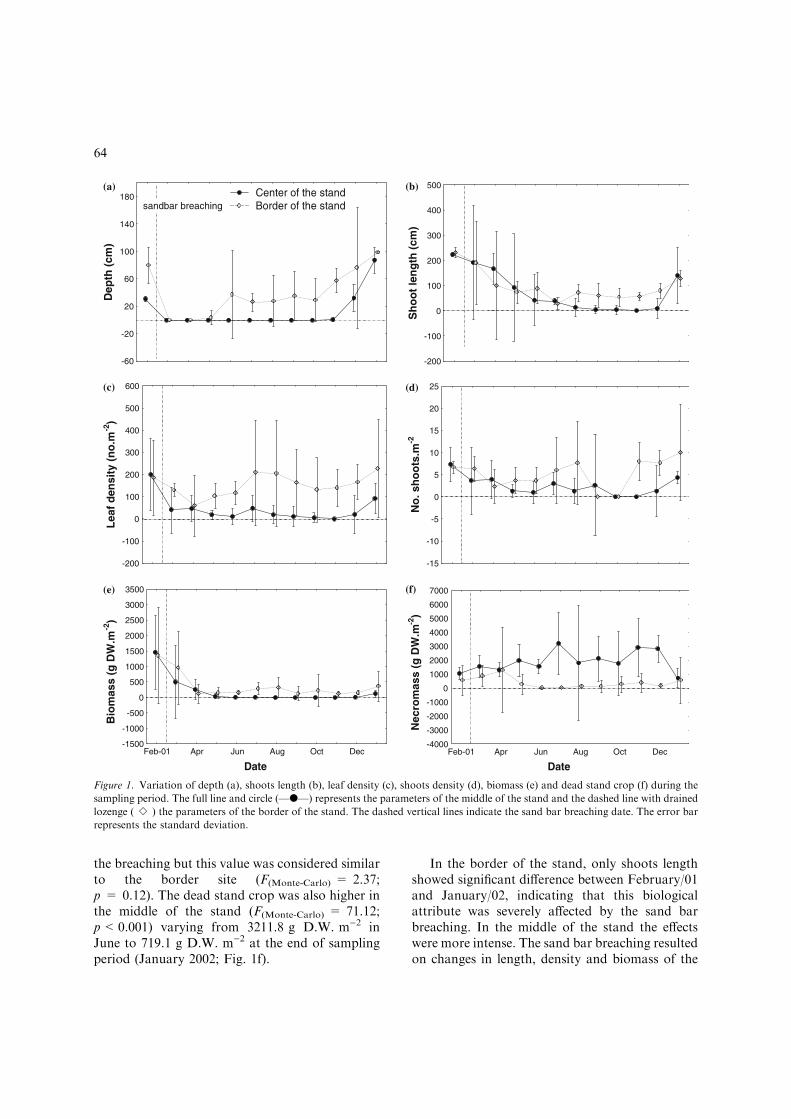
the breaching but this value was considered similarto the border site (F(Monte-Carlo) = 2.37;p = 0.12). The dead stand crop was also higher inthe middle of the stand (F(Monte-Carlo) = 71.12;p<0.001) varying from 3211.8 g D.W. m)2 inJune to 719.1 g D.W. m)2 at the end of samplingperiod (January 2002; Fig. 1f).
In the border of the stand, only shoots lengthshowed significant difference between February/01and January/02, indicating that this biologicalattribute was severely affected by the sand barbreaching. In the middle of the stand the effectswere more intense. The sand bar breaching resultedon changes in length, density and biomass of the
Dep
th (
cm)
-60
-20
20
60
100
140
180(a) (b)
(c) (d)
(e) (f)
Center of the standBorder of the standsandbar breaching
Sh
oo
t le
ng
th (
cm)
-200
-100
0
100
200
300
400
500
Lea
f d
ensi
ty (
no
.m-2
)
-200
-100
0
100
200
300
400
500
600
No
. sh
oo
ts.m
-2
-15
-10
-5
0
5
10
15
20
25
Date
Bio
mas
s (g
DW
.m-2
)
-1500
-1000
-500
0
500
1000
1500
2000
2500
3000
3500
Feb-01 Apr Jun Aug Oct Dec
Date
Nec
rom
ass
(g D
W.m
-2)
-4000
-3000
-2000
-1000
0
1000
2000
3000
4000
5000
6000
7000
Feb-01 Apr Jun Aug Oct Dec
Figure 1. Variation of depth (a), shoots length (b), leaf density (c), shoots density (d), biomass (e) and dead stand crop (f) during the
sampling period. The full line and circle (—d—) represents the parameters of the middle of the stand and the dashed line with drained
lozenge (ÆÆeÆÆ) the parameters of the border of the stand. The dashed vertical lines indicate the sand bar breaching date. The error bar
represents the standard deviation.
64

shoots. When considering the whole T. domingensisstand, both length and aerial biomass werereduced. The effects of the sand bar breaching onT. domingensis population after 1 year samplingare summarized in Table 1 (paired t-test).
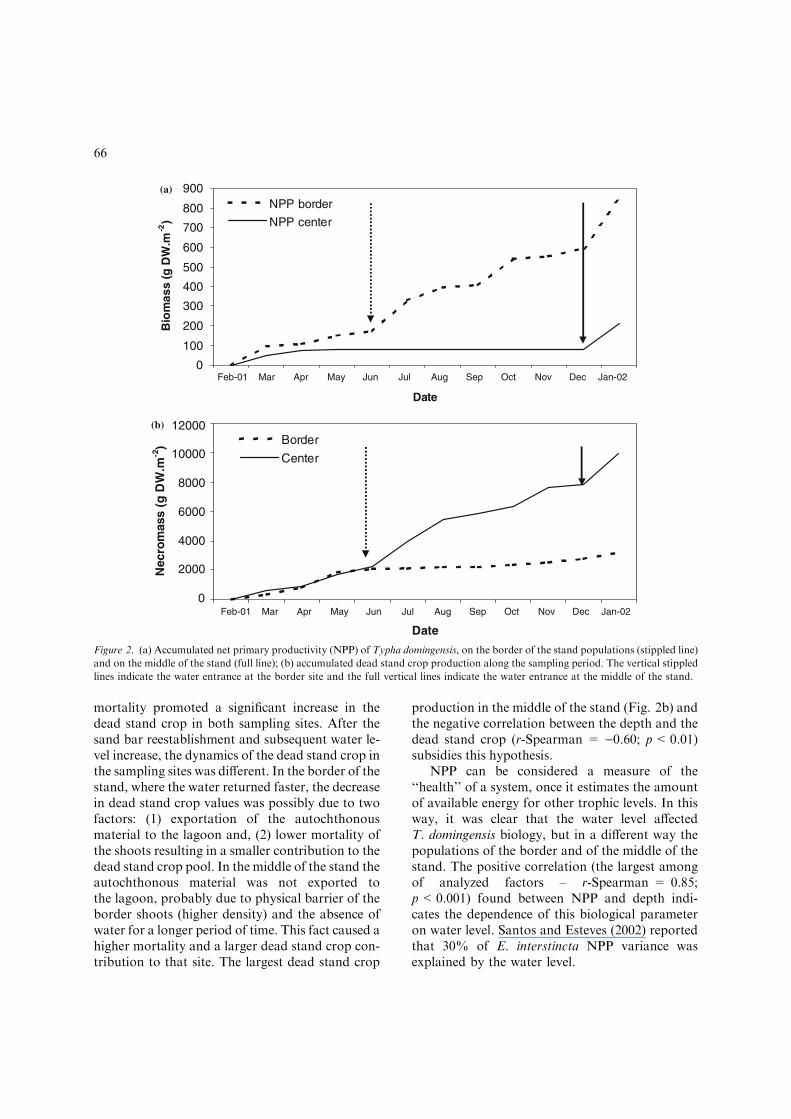
At the end of the study period, the accumulatedNPP was 844.1 g D.W. m)2 in the border of thestand and 213.0 g D.W. m)2 in the middle of thestand (Fig. 2a). The accumulated production ofdead stand crop (Fig. 2b) exhibited an inversepattern to the NPP, with higher values in themiddle of the stand (9977.6 g D.W. m)2) com-pared to the border (3200.0 g D.W. m)2). TheNPP was stimulated during the beginning of watercolumn reestablishment both in the border and inthe middle of the stand. This process was firstinitiated in the border in June, and in the middleonly in December. On the other hand, the watercolumn reestablishment reduced the dead standcrop production. In June 2001 the border popu-lation dead stand crop production was reduceddue to water return to the stand. The water ab-sence due to the sand bar breaching disturbed T.domingensis production and favored the deadstand crop production and accumulation.
Nutrient dynamics
During the sampling period, the stand ofT. domingensis contributedwith 11.50 kgC, 0.22 kgN and 0.13 kg P m)2 to the water column. Assum-ing that the whole T. domingensis stand area canvary from 5 to 10% of the total area of the lagoon,total nutrient input to water column could be be-tween 1882.4 and 3764.7 ton C, 18.0 and 35.9 tonNand 9.0 and 17.9 ton P. Total N and P stock in thewater columnvariedbetween 4890.0 and 76.0 ton to4661.8 and 123.2 ton, respectively, after 1 year
sampling (LTER, Brazil – site 5). T. domingensisdecomposition could has contributed to 19.1–37.9%of the total P stock increase during this study.
Discussion
The water level affected negatively the stand ofT. domingensis, once shoots length and biomassvalues, registered before the sand bar opening,did not recover during the study period(Table 1). The effect of water level on the sur-vival of that species is clear when analyzing thetwo population phenotypes (border and middle)of the stand separately. The absence of water inthe border of the stand occurred only during2 months, while in the middle of the stand, itwas longer (8 months). The absence of the waterin the middle of the stand longer affected nega-tively that population, resulting in lower shootsdensity individuals and, consequently, lowerbiomass. The negative effect of the sand baropening was quite intense in the middle of thestand, once after 1 year sampling the populationwas not recovered yet. Santos and Esteves (2004)reported a 30 days biomass resilience time forrecovery of an Eleocharis interstincta stand inCabiunas coastal lagoon after an event of baropening. The atypically dry year postponed thewater column reestablishment and, consequently,the macrophytes recover in the present study.
The abrupt reduction of water level in coastallagoons, due to the opening of the sand bar, pro-motes the mass mortality of emerging aquaticmacrophytes and the exportation of autochthonousstand material (Palma-Silva et al., 2000; Santos &Esteves, 2002, 2004). This fact was also observed inthe present research, when at first, the shoots
Table 1. Mean value comparison of biological and population attributes before/after the sand bar breaching (first/last sampling
month) of Typha domingensis in the sampling sites (paired t-test)
Attributes Border Middle Total
Shoots length (cm) 231.7/128.4* 223.7/140.3* 227.7/134.4*
Leaf density (no m)2) 185.3/226.7 200.0/92.0 192.7/159.3
Shoots density (no m)2) 26.7/40.0 29.3/17.3* 28.0/28.7
Dead stand crop (g D.W. m)2) 575.0/561.2 1044.7/719.1 809.8/640.2
Biomass (g D.W. m)2) 1350.0/358.4 1458.0/122.2* 1404.0/240.3*
The asterisks mark the significant changes.
65
mortality promoted a significant increase in thedead stand crop in both sampling sites. After thesand bar reestablishment and subsequent water le-vel increase, the dynamics of the dead stand crop inthe sampling sites was different. In the border of thestand, where the water returned faster, the decreasein dead stand crop values was possibly due to twofactors: (1) exportation of the autochthonousmaterial to the lagoon and, (2) lower mortality ofthe shoots resulting in a smaller contribution to thedead stand crop pool. In the middle of the stand theautochthonous material was not exported tothe lagoon, probably due to physical barrier of theborder shoots (higher density) and the absence ofwater for a longer period of time. This fact caused ahigher mortality and a larger dead stand crop con-tribution to that site. The largest dead stand crop
production in the middle of the stand (Fig. 2b) andthe negative correlation between the depth and thedead stand crop (r-Spearman = )0.60; p<0.01)subsidies this hypothesis.
NPP can be considered a measure of the‘‘health’’ of a system, once it estimates the amountof available energy for other trophic levels. In thisway, it was clear that the water level affectedT. domingensis biology, but in a different way thepopulations of the border and of the middle of thestand. The positive correlation (the largest amongof analyzed factors – r-Spearman = 0.85;p<0.001) found between NPP and depth indi-cates the dependence of this biological parameteron water level. Santos and Esteves (2002) reportedthat 30% of E. interstincta NPP variance wasexplained by the water level.
0
100
200
300
400
500
600
700
800
900
Date
Bio
mas
s (g
DW
.m-2
)
NPP border
NPP center
0
2000
4000
6000
8000
10000
12000
Feb-01 Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan-02
Feb-01 Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan-02
Date
Nec
rom
ass
(g D
W.m
-2) Border
Center
(a)
(b)
Figure 2. (a) Accumulated net primary productivity (NPP) of Typha domingensis, on the border of the stand populations (stippled line)
and on the middle of the stand (full line); (b) accumulated dead stand crop production along the sampling period. The vertical stippled
lines indicate the water entrance at the border site and the full vertical lines indicate the water entrance at the middle of the stand.
66

The initial decomposition stages in aquaticmacrophytes occur in the presence of waterthrough leaching (Mason & Bryant, 1975). Oncewe could not observe water column presence in themiddle of the stand for 8 months, a great part ofthe dead stand crop produced remained stockedabove the sediment. This fact impeded the growthof new shoots and leaves, reflecting in the lowNPP, length, density and biomass in the middle ofthe stand. After the water column reestablishmentthe decomposition of the dead material processwas accelerated (Fig. 2b). In the border of thestand, the resilience time was shorter (Fig. 2a)than in the middle, due to two main factors: thefirst one is that the water column reestablished firstand the second is that the macrophyte could pro-ject the new leaves and shoots in the direction ofthe lagoon colonizing exposed areas.
Previously, it was believed that the biomassdynamics in tropical areas was constant, balancedby production processes and equivalent decom-position, a ‘‘steady state’’. Later, some researchesdemonstrated that in these areas, the biomassproduction is influenced by depth (Santos &Esteves, 2002, 2004). Aquatic macrophytes canmaximize the eutrophication process in floodedareas releasing nutrients during decomposition(Planter, 1970; Schoroder, 1973). On the otherhand, they can act in the reduction of eutrophi-cation absorbing N and P of polluted waters(Toth, 1972). Besides, the aquatic macrophytes areable to regulate the nutrients balance in floodedareas and control phytoplanktonic productionthrough the production processes and biomassdecomposition (Mason & Bryant, 1975). In thisresearch, we observed a negative balance betweenNPP and dead stand crop production (Fig. 2a, b),both regulated by water level. This fact indicatesthat the sand bar breaching could promote aninternal fertilization process in Imboassica lagoon,contributing significantly to the nutrients increase.One year after the management, the total amountof N in water (in whole lagoon) showed a slightdecrease from 4890.0 to 4661.8 ton, probably dueto sediment sequestration (Enrich-Prast, pers.com.). However, we could observe a significantincrease in total amount of P, from 76 ton beforesand bar breaching to 123.2 ton in the water col-umn, at the end of the study period. From thesurplus 47 ton P in the water column recorded, we
hypothesized that 19.1–37.9% (considering 5 and10% of covering area, respectively), could beattributed to T. domingensis decomposition re-lease. Then, the potential nutrient removal,through the NPP, of aquatic macrophytes wassupplanted by massive nutrients release, due toshoots death and consequent decomposition. Wecould so recognize that besides exporting nutrientsfrom eutrophic coastal lagoons, the sand barbreaching management may lead to a negativenutrient feedback response due to massivedecomposition process in aquatic macrophytesrich ecosystems, mainly in dry periods or years.
Conclusions
We conclude that the water level plays a funda-mental role on the biology of emergent aquaticmacrophytes in coastal lagoons, regulating bio-mass, NPP, nutrients among other biologicalattributes (Palma-Silva et al., 2000; Santos &Esteves, 2002; Amado et al., 2005).
We can also conclude that the water level fastreestablishment is essential to the macrophytebiological attributes recover once it accelerates thedead stand crop decomposition and exportation.The NPP increase in the presence of water alsocorroborates this hypothesis.
The high dead stand crop production due tomortality of the shoots may consists on a newdisturbance to Imboassica lagoon, which is theinternal fertilization. The decomposition processcould release great amounts of nutrients to thewater column while the macrophytes resproutabsorbing nutrients from the sediment. Thisproblem becomes worse due to the dilution factor,once the nutrients stocked in the dead stand cropare diluted in a very smaller volume compared tothe normal water lagoon volume. The main aim of‘‘cleaning’’ the lagoon was not successfullyaccomplished. Thus, this kind of management,without any ecological support, may amplify theeutrophication process.
Acknowledgements
The authors specially thanks by sampling help bycolleagues from Lab. Limnologia of UFRJ. Forthe financial support by PETROBRAS and by the
67

Scholarships from CNPq, CAPES and FAPERJ.To Adriana M. Rocha, Dr Alex Enrich-Prast andValeria M. Amado for invaluable contributions tothis manuscript and language help.
References
Amado, A. M., F. A. Esteves, M. P. Figueiredo-Barros & A. M.
Santos, 2005. Short-term and seasonal effects of water level
variation chance on Eleocharis interstincta (VAHL) nutrient
content in a tropical coastal lagoon. Acta Limnologica
Brasiliensia 17: 91–99.
Branco, C. W. C., B. K. Suzuki & J. L. Attayde, 1998. Zoo-
plankton community of a coastal lagoon subjected to an-
tropogenic influences (Lagoa Imboacica, Macae, RJ, Brazil).
Archiv fur Hydrobiologie 81: 69–84.
Esteves, F. A., 1998. Lagoas Costeiras: origem, funcionamento
e possibilidades de manejo. In Esteves, F. A. (ed.) Ecologia
das Lagoas Costeiras do Parque Nacional da Restinga de
Jurubatiba e do Municıpio de Macae (RJ) Rio de Janeiro.
NUPEM/UFRJ, in Portuguese: 63–87.
Fiderj, 1977. Estudos para o planejamento ambiental. Fun-
dacao Instituto de Desenvolvimento Economico e Social do
Rio de Janeiro. Rio de Janeiro 67 (in Portuguese).
Furtado, A. L. S., 1998. Ash free dry weight, organic carbon,
nitrogen and phosphorous content of Typha domingensis
PERS. (Typhaceae), an aquatic macrophyte. Verhandlungen
Internationale Vereinigung fur theoretische und angewandte
Limnologie 26: 1842–1845.
Golterman, H. L., R. S. Clymo & M. A. M. Ohnstad, 1978.
Methods of physical and chemical analysis of fresh water.
Blackwell Scientific Publishers, Oxford 214 pp.
Goncalves, J. F. Jr., A. M. Santos & F. A. Esteves, 2004. The
influence of the chemical composition of Typha domingensis
and Nymphaea ampla detritus on invertebrate colonization
during decomposition in a Brazilian coastal lagoon. Hydro-
biologia 527: 125–137.
Gotelli, N. J. & G. L. Entsminger, 2001. EcoSim: Null models
software for ecology. Version 7.22. Acquired Intelligence
Inc. & Kesey-Bear.
Junk, W. J. & M. T. F. Piedade, 1993. Biomass and primary-
production of herbaceous plant communities in the Amazon
floodplain. Hydrobiologia 263: 155–162.
Knoopers, B., 1994. Aquatic primary production in costal la-
goons. In Kjerfve, B. (ed.), Costal Lagoon Process, Elsevier
Science, Elsevier Series 60: 243–286.
Mason, C. F. & R. J. Bryant, 1975. Production, nutrient content
and decomposition of Phrgmites communis Trin. and Typha
angustifolia L. Australian Journal of Ecology 63: 71–95.
Mackeret, F. J. H., J. Heron & J. F. Talling, 1978. Water
analysis: some revised methods for limnologists. Freshwater
Biological Association, Science Publ No. 36, Cumbria and
Dorset.
Palma-Silva, C., E. F. Albertoni & F. A. Esteves, 2002. Clear
water associated with biomass and nutrient variation during
the growth of a Charophyte stand after a drawdown, in a
tropical coastal lagoon. Hydrobiologia 482: 79–87.
Palma-Silva, C., E. F. Albertoni & F. A. Esteves, 2000. Eleo-
charis mutata (L.) Roem. et Schult. subject to drawdowns in
a tropical coastal lagoon, State of Rio de Janeiro, Brazil.
Plant Ecology 148: 157–164.
Panosso, R. & F. A. Esteves, 2000. Effects of disturbance on
phosphorus turnover and plankton dynamics. Journal of
Plankton Research 22: 589–595.
Planter, L., 1970. Elution of mineral components out of dead
reed Phragmites communis Trin. Polskie Archwum Hydro-
biologii 17: 357–362.
Santos, A. M. & F. A. Esteves, 2002. Primary production and
mortality of Eleocharis interstincta in response to water level
fluctuations. Aquatic Botany 74: 189–199.
Santos, A. M., 2004. Macrofitas Aquaticas da Planıcie de
Inundacao do Alto Rio Parana: Produtividade Primaria,
Decomposicao, Ciclagem de Nutrientes e Diversidade.
Parana: Universidade Estadual de Maringa (UEM): 105 pp.
(Ph.D. Thesis).
Santos, A. M. & F. A. Esteves, 2004. Influence of water level
fluctuation on the mortality and aboveground biomass of the
aquatic macrophyte Eleocharis interstincta (VAHL) Roemer
et Schults. Brazilian Archives of Biology and Technology 47:
281–290.
Schroder, R., 1973. Die freisetzung von pflanzennahrstoffen in
schilfgebiet and ihr transport in das freiwasse am beispiel des
bodensee-untersees. Archiv fur Hydrobiologie 71: 145–158.
StatSoft, Inc., 2000. STATISTICA for Windows v5.5 [Com-
puter program manual] Tulsa, OK: StatSoft, Inc., 2300 East
14th Street, Tulsa, OK 74104.
Suzuki, M. S., 1995. Structural changes in the phytoplankton
community of Imboacica lagoon due to seawater inflow.
Abstracts of the XXVI Congress of International Associa-
tion of Theoretical and Applied Limnology, 349 pp.
Symbula, M. & F. P. Day Jr., 1988. Evaluation of two methods
for estimating belowground production in a freshwater
swamp forest. The American Midland Naturalist 120(2):
405–415.
Toth, L., 1972. Reeds control eutrophication of Balaton Lake.
Water Research 6: 1533–1539.
Vitousek, P. M. & H. A. Mooney, 1997. Estimates of coastal
populations – Response. Science 278: 1212–1212.
Wetzel, R. G., 1969. Excretion of dissolved organic compounds
by aquatic macrophytes. Bioscience 19: 539–540.
68


























