Embed Size (px)
Citation preview

Parasitol Res (1993) 79:409-415
Parasitology Research �9 Springer-Verlag 1993
Electron immunogold labeling of regulatory peptide immunoreactivity in the nervous system of Moniez& expansa (Cestoda: Cyclophyllidea) G.P. Brennan 1, D.W. Halton 1, A.G. Maule 2, C. Shaw 2
Comparative Neuroendocrinology Research Group, 1 School of Biology and Biochemistry, The Queen's University of Belfast, Belfast BT7 INN, Northern Ireland, UK 2 School of Clinical Medicine, The Queen's University of Belfast, Belfast BT7 INN, Northern Ireland, UK
Received: 12 February 1993 /Accepted: 15 March 1993
Abstract. An electron immunogold-labeling technique was used in conjunction with a post-embedding proce- dure to demonstrate for the first time the ultrastructural distribution of the parasitic platyhelminth neuropeptide, neuropeptide F (NPF), in the nervous system of the ces- tode Moniezia expansa. Two axon types, distinguished by their populations of different-sized electron-dense ve- sicles, were identified. Immunogold labeling demon- strated an apparent homogeneity of PP, FMRFamide and NPF (M. expansa) antigenic sites throughout the larger dense-cored vesicles within the central nervous system. Triple labeling clearly demonstrated the co-local- isation of immunoreactivities (1R) for NPF, PP and FMRFamide within the same dense-cored vesicles. The presence of NPF-IR within the vesicles occupying the perikaryon of the neuronal cell body indicated that the peptides had undergone post-translational C-terminal amidation prior to entering the axon. Antigen pre-ab- sorption experiments using NPF prevented labeling with either PP or FMRFamide antisera, and the failure of these antisera to block NPF-IR supports the view that some, if not all, of the PP/FMRFamide-IR is due to NPF-like peptides.
Immunocytochemistry studies have shown that regulato- ry peptides occur as major components of the nervous system of parasitic platyhelminths (for reviews see Hal- ton et al. 1990; Fairweather and Halton 1991; Gustafs- son 1992; Halton et al. 1992). Studies on a number of cestode and trematode species have demonstrated im- munoreactivites in both the central and peripheral ner- vous systems to some 30 different vertebrate and inverte- brate peptides using well-characterised peptide antisera. However, although there are several light microscopic accounts of peptide immunoreactivities in tapeworms, only four species have been examined in any detail: Di- phyllobothrium dendriticum (Gustafsson etal. 1985,
Correspondence to: G.P. Brennan
1986; Gustafsson and Wikgren 1989), Trilocularia acan- thiaevulgaris (Fairweather et al. 1990), Hymenolepis di- minuta (Fairweather et al. 1988; McKay et al. 1991) and Moniezia expansa (Maule et al. 1993). Furthermore, rela- tively few studies have been carried out on flatworm neuropeptides at the ultrastructural level. Thus far, Hal- ton et al. (1991) have examined the sub-cellular distribu- tion of pancreatic polypeptide (PP) immunoreactivity in the egg-forming apparatus of the monogenean Diclido- phora merIangi, and Brennan et al. (1992) have demon- strated peptide co-localisation in neuronal vesicles in the central nervous system of D. merlangi using antisera to the C-terminal hexapeptide amide of PP and to the in- vertebrate peptide FMRFamide.
Since the discovery of the first native parasite regula- tory peptide, neuropeptide F (NPF, M. expansa) by Maule et al. (1991), recent studies have indicated that some, if not all, of the PP/FMRFamide immunoreactivi- ty that has been demonstrated previously in the nerves of flatworm parasites may be due to NPF-related pep- tides. Thus, work on D. merIangi by Maule et al. (1992a) using immunocytochemistry and studies by Brennan et al. 1993) using an immunogold-labeling procedure at the electron microscopic level have shown that the im- munostaining pattern observed for NPF immunoreactiv- ity paralleled that documented previously for the verte- brate pancreatic polypeptide (PP) family of peptides and for FMRFamide. These data suggest that an NPF-like peptide, and not PP or FMRFamide, is the predominant native neuropeptide in this monogenean parasite.
The present paper describes the first ultrastructura[ study of the nervous system of M. expansa using electron immunogold-labeling procedures to demonstrate the oc- currence, distribution and co-localisation of regulatory peptides within neurones.
Materials and methods
Specimens of Moniezia expansa were recovered from the intestines of freshly killed sheep at Bangor abattoir, County Down, and were maintained in warm (37 ~ C) saline solution [0.85% (w/v) NaC1]

410
containing 0.1% (w/v) glucose prior to their use. For electron mi- croscopy, transverse slices (1 mm in thickness) of worms containing approximately four proglottids were prepared for electron immun- ocytochemistry using a post-embedding technique. The slices of worm were fixed for 45 min in 1% double-distilled glutaraldehyde (DDG; Agar Scientific, Cambridge, UK) in 0.1 M sodium cacody- late buffer (pH 7.2) containing 3% sucrose at 4 ~ C. The tissues were buffer-washed, subdivided into smaller pieces, partially dehy- drated in ethanol (70%) and then transferred to 70% ethanol at - 2 0 ~ C to complete the dehydration prior to infiltration overnight in Lowicryl K4M resin (TAAB, Watford, England) at - 2 0 ~ C. Specimens were transferred to resin-filled gelatine capsules and were polymerised under UV light (360-nm longwave) for 30 h at - 2 0 ~ C and for a further 72 h at room temperature. Thin sections of approximately 70-80 nm in thickness were cut on a Reichert Ultracut E ultramicrotome and mounted on bare 200-mesh nickel grids.
After drying, the sections were rinsed with 20 mM TRIS/HC1 buffer (pH 8.2) containing 0.1% bovine serum albumin (BSA) and Tween 20 (1 : 40 dilution) and were incubated in normal goat serum (NGS) for 30 min at room temperature and then transferred to one of three primary antisera: pancreatic polypeptide (PP221), FMRFamide 494(2) and neuropeptide F 792(1) (M. expansa). The PP antiserum was raised to the C-terminal hexapeptide amide of bovine (b) PP, the FMRFamide was raised to the whole molecule and the NPF antiserum was raised to synthetic NPF (M. expansa, residues 1-39). The bPP and FMRFamide antisera were diluted 1:5000 with BSA/TRIS/HC1 buffer, whereas the NPF antiserum was diluted 1:30000; each was applied separately to the tissue sections for 18 h at room temperature. After being thoroughly washed in the buffer, the grids were transferred to a 25- 121 droplet of 15-nm-size gold-conjugated goat anti-rabbit IgG (BioCell, Car- diff, UK) and were incubated for 1 h at room temperature. The sections were then buffer-washed, lightly fixed with 1% DDG (2 rain) and finally washed with buffer and rinsed in distilled water. The sections were double-stained with uranyl acetate (20 rain) and aqueous lead citrate (12 min) and then viewed in a JEOL 100CX transmission electron microscope at 100 kV.
Sections for double immunogold labeling were stained sequen- tially with a combination of antisera using either PP/FMRFamide, PP/NPF or FMRFamide/NPF. Following incubation in the prima- ry antiserum as described above, the sections were washed in BSA/ TRIS/HCI buffer and incubated in a 25-121 droplet of 10-nm gold- conjugated goat anti-rabbit IgG for I h at room temperature, buffer-washed and then transferred to a second antiserum for a further 18-h incubation. The sections were again buffer-washed and gold-labeled, this time in a 25-121 droplet of 15-nm gold-conju- gated goat anti-rabbit antiserum, for 1 h and then processed as described above. Sections for multiple gold labeling were incubated sequentially with antisera, firstly directed to NPF, secondly to FMRFamide, and thirdly to PP antisera. After separate incuba- tions in primary and secondary antisera followed by gold labeling with two different-sized gold probes (10 and 15 nm), the sections were incubated in the third primary antiserum for a further 18 h at room temperature, thoroughly washed in buffer, and then trans- ferred to a 25-121 droplet of 5-nm gold-conjugated goat anti-rabbit IgG and incubated for 1 h at room temperature. The sections were finally washed in buffer, rinsed in double-distilled water and dou- ble-stained in uranyl acetate and lead citrate, and viewed in the electron microscope. The sequence of gold-probe sizes used in the multiple labeling procedure were: FMRFamide, 15nm; NPF, 10 nm; and PP, 5 nm. Controls consisted of (a) using a non-im- mune serum to replace the primary antiserum, (b) omission of the primary antiserum and (c) pre-absorption of the primary antise- rum with the appropriate antigen.
Results
The nervous system in Moniezia expansa comprises a well-developed central (CNS) array of paired nerve
cords and associated ganglia together with a peripheral (PNS) network of cell bodies and anastomosing sub- surface fibres. The paired cerebral ganglia, which are connected by a transverse ring commissure, give rise to five pairs of longitudinal nerve cords, each comprising a central core of nerve axons with a surface arrangement of associated somata. The longitudinal nerve cords ex- tend posteriorly from the cerebral ganglia in the scolex through each proglottis, in the posterior region of which they are interconnected by a ring-like transverse com- missure. The PNS consists of an extensive sub-tegumen- tal nerve network and also provides innervation to the musculature of the acetabula and reproductive ducts.
Sections of the lateral nerve cords revealed profiles of duct-like axons, each of which was delimited by a membrane and contained membrane-bound, electron- dense vesicles, microtubules and mitrochondria (Fig. 1). Two distinct types of vesicle were identified within the axons. The first vesicle type had an homogeneous matrix and varied in diameter from 100 to 125 nm; the second type was smaller, measuring approximately 60-80 nm in diameter, with a dense-cored matrix. Although popu- lations of these vesicles occurred in adjacent axons, they were never found together within the same axon. Pre- vious observations using confocal scanning-laser micros- copy demonstrated the presence of three well-developed components to the nervous system: a cholinergic, a pep- tidergic and a serotoninergic component, the latter being the best developed (Maule et al. 1993). Within the pepti- dergic system, immunostaining with PP, FMRFamide and NPF (M. expansa) antisera was the most prominent, and although all three antisera produced identical stain- ing patterns, the most intense fluorescence was obtained with the NPF antiserum. Similarly, immunogold label- ing revealed immunoreactivity (IR) for each of the anti- sera only in those axons containing the larger of the two dense vesicles (Figs. 2~4). This labeling was very specific and was associated almost exclusively with the larger type of vesicle that occupied axons in both the central and the peripheral nervous systems. The gold probes were concentrated over the matrix of the vesicles, with relatively little of the gold occurring in the cytosol.
Fig. 1. A transmission electron photomicrograph showing a portion of longitudinal nerve axon from the anterior central nervous system (CNS) containing both small, electron-lucent vesicles (small arrows) and larger, dense-cored vesicles (large arrows). M, Mitochondria; Mu, muscle). Scale bar, 0.2 p~m. Figs. 2-4. Immunogold labeling of dense-cored vesicles. Fig. 2. Dense-cored vesicles in a nerve axon of the longitudinal nerve cord immunolabeled with gold probes (15-nm size; small arrowheads), demonstrating PP immunoreactivi- ty. A separate population of non-reactive neurovesicles is also pres- ent (large arrowhead). Scale bar, 0.I 12m. Fig. 3. Immunogold label- ing to demonstrate the presence of FMRFamide immunoreactivity in the dense-cored vesicles (15-nm-size gold probe, arrowheads). Note the neutrotubules (Nt) and mitochondria (M). Scale bar, 0.i 12m. Fig. 4. A portion of a longitudinal nerve axon following immunogold labeling to demonstrate the presence of neuropeptide F (NPF - Moniezia expansa) immunoreactivity in the dense-cored vesicles (15-nm-size gold probe, arrowheads). Note the neurotu- bules (Aft) and mitochondrion (M). Scale bar, 0.1 12m

T.

412
Figs. 5, 6. Dense-cored vesicles in a longitudinal nerve axon follow- ing double immunogold labeling, demonstrating co-localisation of PP (15-nm-size gold label, large arrows) and NPF (10-nm-size gold label, small arrows) as well as FMRFamide (15-nm-size gold label, large arrows) and NPF immunoreactivity within the same vesicle. Fig. 5. Incubation in NPF antiserum followed by PP antiserum. Scale bar, 0.1 ~tm. Fig. 6. Incubation in NPF antiserum followed
by FMRFamide antiserum. Scale bar, 0.1 gm. Fig. 7. Section of a longitudinal axon following triple immunogold labeling, demon- strating immunoreactivity in the one dense-cored vesicle to NPF (10-rim gold probe, medium-sized arrow), PP (5-nm gold probe, small arrow) and FMRFamide (15-nm gold probe, large arrow). Scale bar, 0.1 gm
A number of the vesicles remained untagged, and no labeling was observed outside the axon. Nerve axons containing the smaller dense-cored vesicles were at all times negative.
The double immunogold-labeling technique demon- strated the occurrence of PP /NPF (M. expansa) and FMRFamide /NPF (M. expansa) within the same dense- cored secretory vesicles (Figs. 5, 6). The triple-labeling sequence illustrated the presence of PP-, FMRFamide- and N P F - I R within the same dense-cored vesicles (Fig. 7). Immunoreactivity was quenched following the prein-
cubation of sections with synthetic NPF (residues 1-39), and sections from incubations in which the primary anti- serum was omitted or replaced with non-immune serum were at all times negative.
In sections containing neuronal cell bodies, immuno- staining was always confined to the dense-cored vesicles distributed through the cytoplasm (Fig. 8). Probe counts showed a significant increase in the number of gold par- ticles per vesicle in the neck region of the neurone as distinct from those in the perikaryon area. Thus, the number of gold probes counted in the vesicles occupying
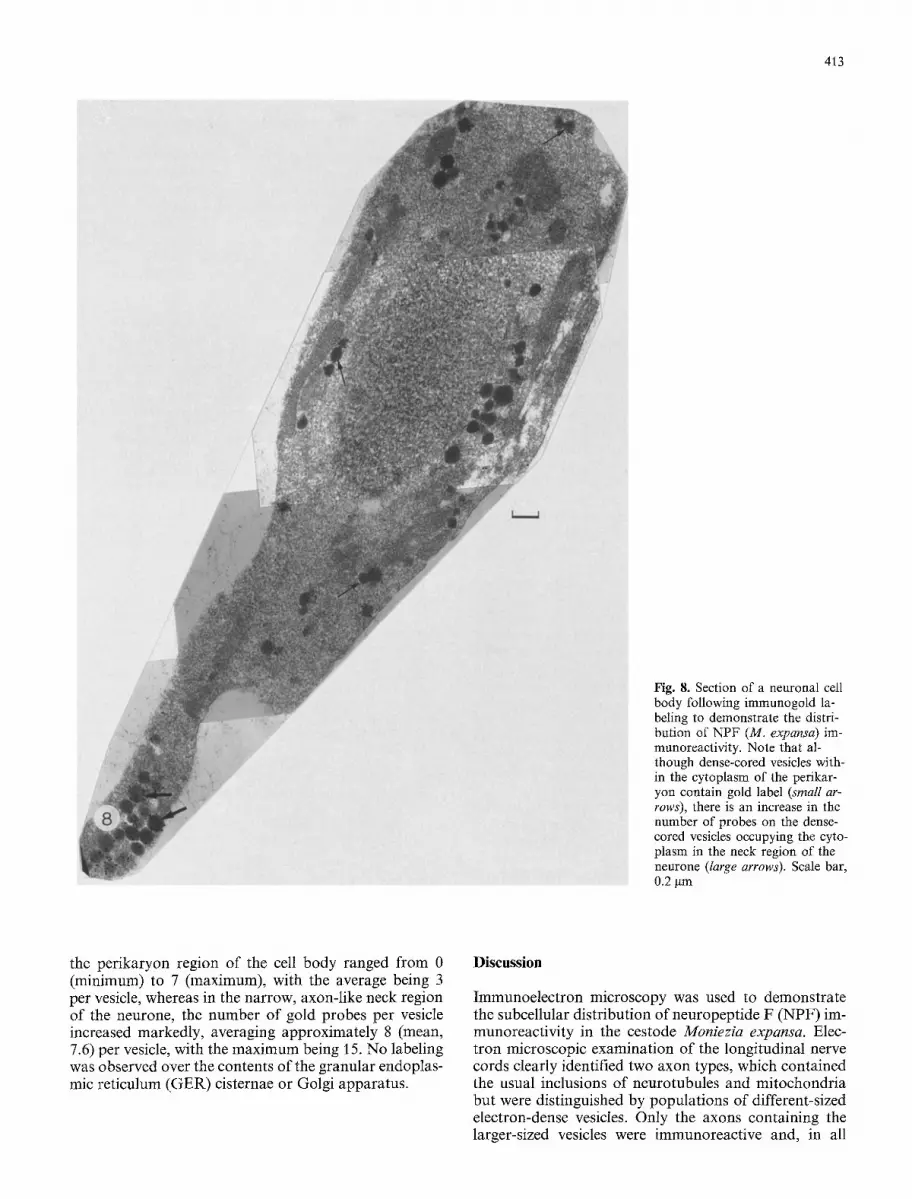
413
Fig. 8. Section of a neuronal cell body following immunogold la- beling to demonstrate the distri- bution of NPF (M. expansa) im- munoreactivity. Note that al- though dense-cored vesicles with- in the cytoplasm of the perikar- yon contain gold label (small ar- rows), there is an increase in the number of probes on the dense- cored vesicles occupying the cyto- plasm in the neck region of the neurone (large arrows). Scale bar, 0.2 gm
the perikaryon region of the cell body ranged from 0 (minimum) to 7 (maximum), with the average being 3 per vesicle, whereas in the narrow, axon-like neck region of the neurone, the number of gold probes per vesicle increased markedly, averaging approximately 8 (mean, 7.6) per vesicle, with the maximum being 15. No labeling was observed over the contents of the granular endoplas- mic reticulum (GER) cisternae or Golgi apparatus.
Discussion
hnmunoelectron microscopy was used to demonstrate the subcellular distribution of neuropeptide F (NPF) im- munoreactivity in the cestode Moniezia expansa. Elec- tron microscopic examination of the longitudinal nerve cords clearly identified two axon types, which contained the usual inclusions of neurotubules and mitochondria but were distinguished by populations of different-sized electron-dense vesicles. Only the axons containing the larger-sized vesicles were immunoreactive and, in all

414
cases, labeling of peptide immunoreactivity was confined to the contents of the vesicle.
Previous immunocytochemistry studies of the ner- vous system of M. expansa by Maule et al. (1992 b, 1993) showed that, in common with most other parasitic platy- helminths examined, the most intense, immunoreactivity (IR) was obtained using antisera to members of the NPY superfamily, in particular an antiserum directed to the C-terminal hexapeptide amide of PP (Maule et al. 1989, 1990a, b). In the present study, immunogold labeling of the nervous system of M. expansa demonstrated clear- ly an apparent homogeneity of PP, FMRFamide and NPF (M. expansa) antigenic sites throughout the larger of the dense-cored vesicles within the central nervous system of the worm. The pre-absorption experiments us- ing NPF prevented immunolabeling with either PP or FMRFamide antisera, and this finding together with the failure of either PP or FMRFamide to block NPF-IR in M. expansa supports the view that some, if not all, of the PP/FMRFamide-IR demonstrable in parasitic platyhelminths (see Maule et al. 1992a; Brennan et al. 1993) is due to NPF-like peptides. Preincubation experi- ments clearly demonstrated that both the PP and the FMRFamide antisera employed cross-reacted with NPF (M. expansa), and this cross-reactivity would account for the co-localisation of immunoreactivities for NPF, PP and FMRFamide observed in the same dense-cored vesicles using the multiple gold-labeling procedure.
Since both the PP and the NPF antisera employed require C-terminal amidation and do not cross-react with PP or NPF free acids, it is possible to determine at what stage C-terminal amidation of the parasite pep- tide occurs. In this respect, the demonstration of NPF- IR within those vesicles occupying the cytoplasm of the neuronal cell body shows that the peptides have under- gone C-terminal amidation in the perikaryon prior to their entry into the axon. However, the absence of any gold labeling over the GER and Golgi apparatus indi- cates that this event in post-translational processing does not occur before the peptide is packaged into vesicles.
Since the discovery of NPF (M. expansa) by Maule et al. (1991), analogous peptides have been isolated and sequenced from the turbellarian Artioposthia triangulata (Curry et al. 1992), from the gastropod molluscs Helix aspersa (Leung et al. 1992) and Aplysia californica (Raj- para et al. 1992) and from the cephalopod mollusc Loli- go vulgaris (Smart et al. 1992). All of the NPF species thus far identified contain the sequence A r g - X - A r g - P h e - N H 2 (RXRFamide), where X is either Pro (P) or Thr (T). All of these NPF peptides cross-react with anti- sera directed to the C-terminal hexapeptide amide of PP and to the C-terminus of FMRFamide. This cross- reactivity probably accounts for the parallel immuno- staining observed for PP, FMRFamide and NPF. Addi- tionally, the inability of both PP and FMRFamide to block NPF-IR in M. expansa supports the interpretation that much, if not all, of the immunostaining for PP and FMRFamide that has previously been recorded in other parasitic platyhelminths is the result of a non-specific cross-reactivity of these antisera with an NPF-related peptide. Indeed, this would account for the present find-
ing of reactivity for NPF, PP and FMRFamide in the same dense-cored vesicles using the triple immunogold- labeling procedure.
In conclusion, the results presented herein represent the first ultrastructural description, using immunogold labeling, of regulatory peptide immunoreactivity in the nervous system of the cestode M. expansa. The study demonstrates the abundance of NPF-immunoreactive secretory vesicles within the CNS of the worm and indi- cates that C-terminal amidation of the NPF (M. expan- sa) peptide occurs within its carrier vesicles prior to its leaving the cell body.
References
Brennan GP, Halton DW, Maule AG, Shaw C, Johnston CF, Moore S, Fairweather I (1993) Immunoelectron microscopical studies of regulatory peptides in the nervous system of the monogenean parasite, Diclidophora merlangi. Parasitology 106 171-176
Curry J, Shaw C, Johnston CF, Thim L, Buchanan KD (1992) Neuropeptide F: primary structure from the turbellarian Artio- posthia triangulata. Comp Biochem Physiol [C] 101:269-274
Fairweather I, Halton DW (1991) Neuropeptides in platyhelminths. Parasitology 102: $77-$92
Fairweather I, McCartney GA, Johnston CF, Halton DW, Buchan- an KD (1988) Immunocytochemical demonstration of 5-hy- droxytryptamine (serotonin) and vertebrate neuropeptides in the nervous system of excysted cysticercoid larvae of the rat tapeworm (Hyrnenolepis diminuta (Cestoda, Cyclophyllidea). Parasitol Res 74:371-379
Fairweather I, Mahendrasingham S, Johnston CF, Halton DW, Shaw C (1990) Peptidergic nerve elements in three developmen- tal stages of the tetraphyllidean tapeworm Trilocularia acan- thiaevulgaris. An immunocytochemical study. Parasitol Res 76: 497-508
Gustafsson MKS (1992) The neuroanatomy of parasitic flatworms. Adv Neurol Immunol 2: 267-286
Gustafsson MKS, Wikgren MC (1989) Development of immunore- activity to the invertebrate neuropeptide small cardiac peptide B in the tapeworm Diphyllobothriurn dendriticum. Parasitol Res 75 : 396-400
Gustafsson MKS, Wikgren MC, Karhi TJ, Schot LPC (1985) Im- munocytochemical demonstration of neuropeptides and sero- tonin in the tapeworm Diphyllobothrium dendriticum. Cell Tis- sue Res 240: 255-260
Gustafsson MKS, Lehtonen MAI, Sundler F (1986) Immunocyto- chemical evidence for the presence of mammalian neurohor- monal peptides in neurons of the tapeworm Diphyllobothrium dendriticum. Cell Tissue Res 243:41-49
Halton DW, Fairweather I, Shaw C, Johnston CF (1990) Regulato- ry peptides in parasitic platyhelminths. Parasitol Today 6: 284- 290
Halton DW, Brennan GP, Maule AG, Shaw C, Johnston CF, Fair- weather I (1991) The ultrastructure and immunogold labelling of pancreatic polypeptide-immunoreactive ceils associated with the egg-forming apparatus of a monogenean parasite, Diclido- phora merlangi. Parasitology 102:429-436
Halton DW, Shaw C, Maule AG, Johnston CF, Fairweather I (1992) Peptidergic messengers: a new perspective of the nervous system of parasitic platyhelminths. J Parasitol 78:179-193
Leung PS, Shaw C, Maule AG, Thim L, Johnston CF, Irvine GB (1992) A molluscan neuropeptide F. Regul Peptides 35:242
Maule AG, Shaw C, Halton DW, Johnston CF, Fairweather I (1989) Localization, quantification and characterization of pan- creatic polypeptide immunoreactivity in the parasitic flatworm Diclidophora merlangi and its fish host (Merlangius merlangus). Gen Comp Endocrinol 74: 50-56

415
Maule AG, Halton DW, Johnston CF, Shaw C, Fairweather I (1990 a) The serotoninergic, cholinergic and peptidergic compo- nents of the nervous system in the monogenean parasite Diclido- phora merlangi: a cytochemical study. Parasitology 100:255 273
Maule AG, Halton DW, Johnston CF, Shaw C, Fairweather I (1990 b) A cytochemical study of the serotoninergic, cholinergic and peptidergic components of the reproductive sytstem in the monogenean parasite. Diclidophora merlangi. Parasitol Res 76: 409419
Maule AG, Shaw C, Halton DW, Thim L, Johnston CF, Fair- weather I, Buchanan KD (1991) Neuropeptide F: a novel para- sitic flatworm regulatory peptide from Moniezia expansa (Ces- toda: Cyclophyllidea). Parasitology 102:30%316
Maule AG, Brennan GP, Halton DW, Shaw C, Johnston CF, Moore S (1992a) Neuropeptide F-immunoreactivity in the monogenean parasite Diclidophora merlangi. Parasitol Res 78 : 655-660
Maule AG, Shaw C, Halton DW, Brennan GP, Johnston CF, Moore S (1992b) Neuropeptide F (Moniezia expansa): localiza-
tion and characterization using specific antisera. Parasitology 105:505-512
Maule AG, Halton DW, Shaw C, Johnston CF (1993) The choli- nergic, serotoninergic and peptidergic components of the ner- vous system of Moniezia expansa (Cestoda, Cyclophyllidea). Parasitology 106: 429-440
McKay DM, Fairweather I, Johnston CF, Shaw C, Halton DW (1991) Immunocytochemical and radioimmunometrical demon- stration of serotonin- and neuropeptide-immunoreactivities in the adult rat tapeworm, Hymenolepis diminuta (Cestoda, Cyclo- phyllidae). Parasitology 103:275-289
Rajpara SM, Garcia PD, Roberts R, Eliassen JC, Owens DF, Maltby D, Myers RM, Mayeri E (1992) Identification and mo- lecular cloning of a neuropeptide Y homolog that produces prolonged inhibition in Aplysia neurons. Neuron 9:505-513
Smart D, Shaw C, Johnston C, Thim L, Halton D, Buchanan K (1992) Peptide tyrosine phenylalanine: a novel neuropeptide F-related nonapeptide from the brain of the squid, Loligo vul- garis. Biochem Biophys Res Commun 186:1616-1623














![[Compared parasitic infection of Ligula intestinalis (Cestoda: Diphyllobothridae) in Cyprinidae species: Rutilus rubilio and Scardinius erythrophthalmus in two dam reserves in Tunisia]](https://img.pdfslide.net/doc/110x75/634ec2490d6acb880b02f6c2/compared-parasitic-infection-of-ligula-intestinalis-cestoda-diphyllobothridae.jpg)














