Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Karyotype, chromosomal characteristics of multiple rDNA clusters and intragenomicvariability of ribosomal ITS2 in Caryophyllaeides fennica (Cestoda)☆
Martina Orosová a,b,⁎, Králová-Hromadová Ivica a, Bazsalovicsová Eva a, Špakulová Marta a
a Parasitological Institute, Slovak Academy of Sciences, Hlinkova 3, 040 01 Košice, Slovakiab Institute of Parasitology, Biology Centre ASCR, Branišovská 31, 370 05 České Budějovice, Czech Republic
a b s t r a c ta r t i c l e i n f o
Article history:Received 1 March 2010Received in revised form 27 April 2010Accepted 28 April 2010Available online 11 May 2010
Keywords:Tapeworm cytogeneticKaryotypeHeterochromatinFluorescent in situ hybridizationNucleolar organizer regionDivergent ITS copies
Karyotype and chromosomal characteristics, i.e. number and location of ribosomal DNA (rDNA) clusters, andsequence variation of the ribosomal internal transcribed spacer 2 (ITS2) were studied in a monozoic(unsegmented) tapeworm, Caryophyllaeides fennica (Caryophyllidea), using conventional and Ag-staining,fluorescent in situ hybridization (FISH) with 18S rDNA probe, and PCR amplification, cloning and sequencingof the complete ribosomal ITS2 spacer. The karyotype of this species was composed of ten pairs ofmetacentric (m) chromosomes (2n=20). All chromosomes except the pair No. 2 displayed DAPI-positiveheterochromatin in centromeric regions. In addition, two distinct interstitial DAPI-positive bands wereidentified on chromosome pair No. 7. FISH with 18S rDNA probe revealed four clusters of major ribosomalgenes situated in the pericentromeric region of the short arms in two pairs of metacentric chromosomes Nos.8 and 9. Hybridization signals were stronger in the pair No. 8, indicating a higher amount of rDNA repeats atthis nucleolar organizer region (NOR). Analysis of 15 ITS2 rDNA sequences (five recombinant clones fromeach of three individuals) showed 13 structurally different ribotypes, distinguished by 26 nucleotidesubstitutions and variable numbers and combinations of short repetitive motifs that allowed sorting thesequences into four ITS2 variants. These results contribute to recently published evidence for theintraindividual ribosomal ITS sequence variability in basal tapeworms with multiple rDNA loci and implythat both phenomena may be mutually linked.
© 2010 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Caryophyllidea (Cestoda) are intestinal parasites of freshwatercypriniform and siluriform fishes that utilize aquatic annelids asintermediate hosts and have almost global distribution [1]. Morpho-logically, they exhibit unique features among tapeworms in havingneither internal proglottidization nor external segmentation, andpossessing a single set of reproductive organs. The group apparentlyplayed a key role in the evolution of cestodes [2]. Caryophyllideansrepresent basal or nearly basal tapeworms, however, their evolution-ary interrelationships with the most relative groups of tapeworms(Diphyllobothriidea, Bothriocephallidea, and Spathebothriidea) stillremain unsolved. Similarly, phylogenetic relationships within theorder Caryophyllidea and systematic position of some caryophylli-dean tapeworms are an ongoing theme [3–5].
Within the order, specific molecular and genetic features havebeen observed. Triploidy was detected in Atractolytocestus huronensis
[6,7], and parallel diploid/triploid populations are known forGlaridacris catostomi, Glaridacris laruei, Isoglaridacris bulbocirrus, andCaryophyllaeus laticeps [8–10]. Besides, intragenomic ITS1 and ITS2rDNA variants were revealed in A. huronensis [7].
Caryophyllidean tapeworms, with their special morphological,molecular and genetic features seem to be a suitable model group forthe study of molecular mechanisms of gene conversion and concertedevolution [7]. In addition to recently used molecular approachesincluding sequential gene analyses, advanced cytogenetic investiga-tions of chromosome structure and localization of genes couldsignificantly contribute to understanding these mechanisms.
Molecular-cytogenetic methods, namely various modifications offluorescent in situ hybridization (FISH), visualize spatial distributionof particular DNA sequences or even selected genes, which can bethen used as species-specific markers. For example, FISH with probesfor ribosomal RNA genes, the so-called rDNA FISH, is used todetermine the number and chromosomal location of major rDNAs,i.e. the nucleolar organizer regions (NORs). However, this method hasonly rarely been applied to parasitic worms. With rDNA-FISH,Bombarová et al. [11] revealed interspecific karyotype differencesbetween two sibling species of the acanthocephalan genus Pomphor-hynchus. Recently, Králová-Hromadová et al. [7] and Orosová et al.[12] determined the number and location of NORs for two
Parasitology International 59 (2010) 351–357
☆ Nucleotide sequence data reported in this paper are available in the GenBankTM,EMBL and DDBJ databases under the accession numbers GU325692–GU325706.⁎ Corresponding author. Parasitological Institute, Slovak Academy of Sciences,
Hlinkova 3, 040 01 Košice, Slovakia. Tel.: +421 55 6334455; fax: +421 55 6331414.E-mail address: [email protected] (M. Orosová).
1383-5769/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.parint.2010.04.007
Contents lists available at ScienceDirect
Parasitology International
j ourna l homepage: www.e lsev ie r.com/ locate /par in t

Author's personal copy
caryophyllidean tapeworms, A. huronensis and Khawia saurogobii,respectively.
The internal transcribed spacers (ITS) of the rDNA clustersrepresent frequently used species-specific molecular markers usedalso in phylogenetic studies of lower taxonomic groups. Lowintraspecific variation, but consistent interspecific differences aregeneral features of ribosomal spacers [13,14], that corresponds withthe traditional theories of concerted evolution [15]. Contrary to this,divergent intragenomic ITS copies have recently been observed invarious groups of organisms, including parasitic flatworms [7,16–20].Ribosomal spacers are often applied in taxonomic, population andphylogenetic studies of cestodes [21], and, therefore, adequateknowledge of ribosomal intra- and interspecific variability needs tobe understood.
The monozoic tapeworm Caryophyllaeides fennica (Schneider,1902) (Lytocestidae) was originally described from the ruddScardinius erythrophthalmus (L.) captured in south coast of Finland(Schneider 1902). C. fennica is a frequent and widely distributedparasite throughout Europe and Russia, and it has been reported frommany cyprinid fish hosts [22,23]. The aim of this study was to extendthe knowledge of molecular and genetic characteristics of thecaryophyllidean C. fennica. We examined karyotype, i.e. number,size, morphology, DAPI-positive bands, and the number and locationof NORs. We also amplified, cloned, and sequenced the ribosomalinternal transcribed spacer 2 (ITS2). Cytogenetic data, multiple rDNAloci and divergent intragenomic ITS copies are discussed as possibletools for evolutionary studies in basal tapeworms.
2. Materials and methods
2.1. Specimens
Specimens of C. fennicawere obtained from the intestines of whitebream Blicca bjoerkna collected in the Tisa and Latorica Rivers (easternSlovakia) in May 2008 and April 2009. Helminths were isolated fromthe intestine of freshly killed hosts, rinsed several times in salinesolution, and immediately used for chromosome preparations or fixedin 96% ethanol for DNA extraction.
2.2. Chromosome preparations
Whole living tapeworms were treated with hypotonic solution of75 mM KCl for 2 h at room temperature (RT). The material was thenfixed in freshly prepared fixative solution (methanol:acetic acid, 3:1)with two changes, 15 min each. Spread preparations of mitotic andmeiotic chromosomes were made as described by Frydrychová andMarec [24]. Small portions (medullary parenchyma with testes) offixed worms were transferred into a drop of 60% acetic acid on a slideand torn into fine pieces with a help of tungsten needles. Then theslide was placed on a heating plate at 45 °C, and the drop of cellsuspension was slowly drawn along the slide until it evaporated.Slides were dehydrated in an ethanol series (70%, 80%, and 100%,1 min each) and stored at −20 °C until use.
2.3. Karyological analysis
Slides were stained with 4% Giemsa solution (pH 6.8) for 30 min.Chromosome indexes (absolute and relative length and centromericindex) were calculated in 10 best mitotic spreads out of 58 analyzedcells chosen from three worms. The chromosomes were classifiedaccording to the nomenclature of Levan et al. [25].
2.4. Silver nitrate (AgNO3) staining
AgNO3 staining was performed as described by Howell and Black[26] with slight modifications. Six drops of 50% AgNO3 in water and
three drops of 2% gelatine in 1% formic acid weremixed on the surfaceof spread chromosome preparation and covered with a cover slip.Then the slide was incubated at RT until colour of the preparationchanged to light brown.
2.5. Fluorescence in situ hybridization with 18S rDNA probe
For FISH technique, unlabelled, about 2100 bp long 18S rDNAprobe was generated by PCR from the C. laticeps genomic DNA.Specimens of C. laticeps, used for DNA extraction, were obtained fromthe intestine of common bream Abramis brama collected from the TisaRiver, (eastern Slovakia). The genomic DNA was extracted using aDNeasy Tissue Kit (Qiagen, Hilden, Germany). PCR was carried outusing two primers, 18S-WormA forward (5′-GCGAATGGCTCAT-TAAATCAG-3′) and 18S-WormB reverse (5′-CTTGTTACGACTTT-TACTTCC-3′) as described by Littlewood and Olson [27]. The probewas labelled with biotin-14-dATP by nick translation using a BionickLabeling System (Invitrogen, Life Technologies Inc., San Diego, CA,USA).
For FISHwe used the procedure described by Sahara et al. [28] withslight modifications as given in Fuková et al. [29]. After removal fromthe freezer, chromosome preparations were passed through anethanol series and air dried. Then the preparations were treatedwith proteinase K (20 mg/ml) in 1× PBS for 5 min at 37 °C, washedtwice in 1× PBS for 5 min each, and then digested with 100 μg/mlRNase A in 2× SSC for 1 h at 37 °C and washed twice in 2× SSC for5 min each. The slides were incubated in 5× Denhard's solution for30 min at 37 °C. Denaturation of chromosomal DNAs was done in 70%formamide in 2× SSC for 3 min and 30 s at 68 °C. The probe cocktail forone slide (10 μl; 50% deionized formamide, 10% dextran sulfate in 2×SSC) contained ∼30 ng of biotinylated 18S rDNA probe and 25 μg ofsonicated salmon spermDNA (Sigma-Aldrich, St. Louis, MO, USA). Theprobe was denatured at 90 °C for 5 min. Hybridization at 37 °C for20 h was followed by stringent washes, which included 50%formamide in 2× SSC (3× 5 min, 46 °C) (Fluka, Buchs, Switzerland),2× SSC (5× 2 min, 46 °C), 0.1× SSC (3× 5 min, 62 °C), and 4× SSCcontaining 0.1% Tween 20 (3×3 min, 37 °C). Hybridization signalswere detected with Cy3-conjugated streptavidin (Jackson Immu-noRes. Labs. Inc., West Grove, PA, USA), amplified with one round ofbiotinylated anti-steptavidin (Vector Labs. Inc., Burlingame, CA, USA)and Cy3-conjugated streptavidin. The preparations were counter-stained with 0.5 μg/ml DAPI (4′,6-diamino-2-phenylidole; Sigma-Aldrich) and mounted in DABCO-based antifade (Sigma-Aldrich).
2.6. Microscopy and image processing
Preparations were observed in conventional fluorescence micro-scopes, Zeiss Axioplan 2 (Carl Zeiss Jena, Germany) equipped with F-View CCD camera (Soft Imaging System GmbH, Münster, Germany)and Olympus B51 equipped with DP70 CCD camera (Olympus Europa,GmbH, Hamburg, Germany). Black-and-white fluorescent imageswere pseudocoloured (light blue for DAPI, and red for Cy3) andprocessed with Adobe Photoshop, version 7.0.
2.7. PCR amplification, cloning and sequencing of ribosomal ITS2
Three C. fennica tapeworm individuals were used. Immediatelyafter dissection, they were washed several times in physiologicalsolution and stored in 96% ethanol. Genomic DNA was isolated usingthe QIAamp DNA Kit (Qiagen). For PCR amplification, a total volume ofamplification mixture was 20 μl and contained 10–20 ng of genomicDNA, 20 pmol of each of the forward and reverse primers, 0.2 mM ofeach of the deoxynucleotide triphosphate (Fermentas UAB, Vilnius,Lithuania), 0.5 U of Taq DNA polymerase (Invitrogen, CA, USA) withcorresponding reaction buffer and 1.5 mMMgCl2. For amplification ofthe complete ITS2 region, the universal forward primer 5.8S–2 (5′-
352 M. Orosová et al. / Parasitology International 59 (2010) 351–357
Author's personal copy
GTCGATGAAGAGCGCAGC-3′) [30] and reverse primer ITS-2 (5′-AGGAGGCGAATCACTAT-3′) [31] with annealing positions in the 5.8SrRNA and 28S rRNA, respectively, were applied.
The PCR amplifications were run as follows: 5 min at 94 °C as aninitial step; then 30 cycles of 1 min at 94 °C, 1 min at 55 °C, and 2 minat 72 °C. The final step was 5 min at 72 °C. The PCR products wereloaded in the 1.5% agarose gel, stained with ethidium bromide, andpurified using the Wizard PCR purification Kit (Promega, Madison,Wisconsin, USA). Purified PCR products of ITS2 spacer regionamplified from three individuals were cloned into the pGEM-T Easy
vector (Promega) following the manufacturer's protocol. For eachstudied individual, DNA from five recombinant clones was purifiedwith the Plasmid miniprep kit (Genomed, Löhne, Germany) andsequenced with universal primers T7 and Sp6 using the AppliedBiosystems 3130xl Genetic Analyzer (Applied Biosystems, Foster City,CA, USA) and BigDye Terminator v3.1 Cycle sequencing kit (AppliedBiosystems).
The boundaries of both spacers were determined according to thesequences of phylogenetically related cestodes, bothriocephalideanBothriocephalus acheilognathi (GenBank accession no. EF417920), and
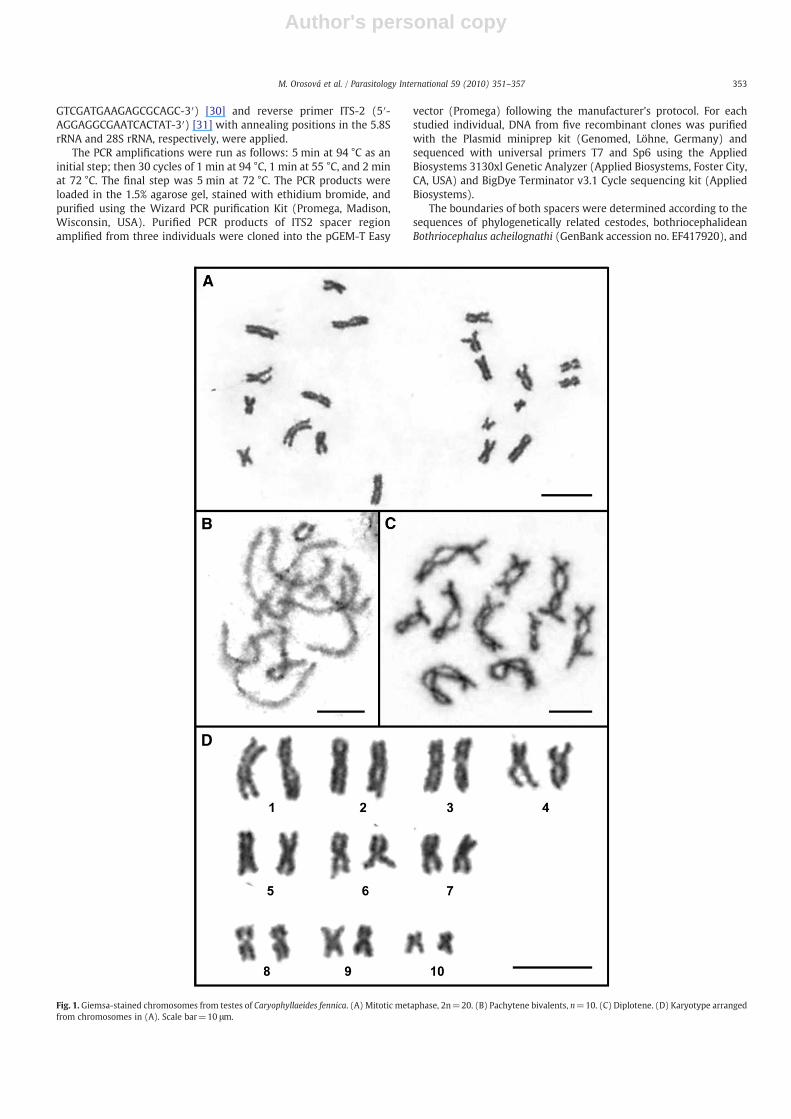
Fig. 1. Giemsa-stained chromosomes from testes of Caryophyllaeides fennica. (A) Mitotic metaphase, 2n=20. (B) Pachytene bivalents, n=10. (C) Diplotene. (D) Karyotype arrangedfrom chromosomes in (A). Scale bar=10 μm.
353M. Orosová et al. / Parasitology International 59 (2010) 351–357

Author's personal copy
diphyllobothriideans Diplogonoporus grandis, Ligula sp. [32], andDiphyllobothrium nihonkaiense (GenBank accession nos. AB298514,AF385766, and AB288368). The sequence alignments were performedusing ClustalW [33]. Sequences were deposited in the GenBankdatabases under the accession numbers GU325692–GU325706.
3. Results
3.1. Karyotype of C. fennica
Out of seven cytologically analyzed C. fennica specimens, dividingmitotic and meiotic cells were found in five worms. The diploidchromosome number was 2n=20 and FN=40 (Fig. 1A). Inpachytene and diplotene/diakinesis stages, a haploid set of 10 meioticbivalents was regularly observed (Fig 1B, C). This number ofchromosomes was found in 58 dividing mitotic cells (81.7%) from71 examined. The remaining spreads were incomplete having 14–19chromosomes that is most probably due to the preparation techniqueused. All chromosomes were metacentric (Fig. 1D; for chromosomemeasurements, see Table 1).
The chromosomes are relatively large; in 10metaphases used for themetric characterization, the largest chromosome measured 8.19±1.29 μm and the smallest was 2.8±0.46 μm long (Table 1). Themean total length of haploid genome (TLC) reached 58.65 μm. Nosecondary constrictionwas observed on Giemsa-stained chromosomes.
Fluorescent DAPI-staining revealed a distinct banding pattern onthe chromosomes. Chromosomes of nine pairs showed each a brightheterochromatin band in the centromeric region while chromosomesof pair No. 2 were without any DAPI-positive band in all analyzedmetaphases (see asterisks in Fig. 2A). Additionally, two distinctinterstitial heterochromatin bands were identified in chromosomes ofpair No. 7, one on the short arm and the other on the long arm (seearrows in Fig. 2C, D).
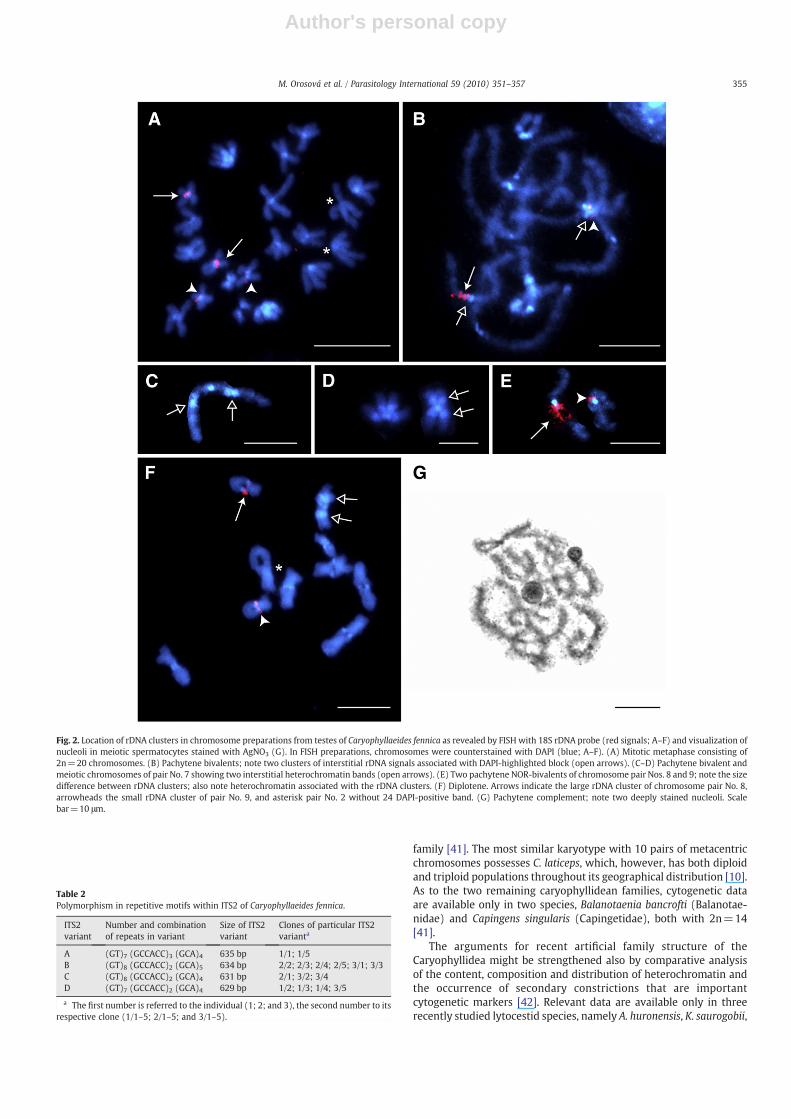
3.2. Localization of rDNA
In mitotically dividing spermatogonia, FISH with 18S rDNA proberevealed four positive signals situated in the pericentromeric region ofshort arms in two pairs of 10 metacentric chromosomes Nos. 8 and 9(Fig. 2A). Hybridization signals were much stronger in both chromo-somes of the pair No. 8 (Fig. 2A, arrows), indicating a markedly higheramount of rDNA at this site in comparison with the NORs of bothchromosomes of the pair No. 9, which showed very small signals(Fig. 2A, arrowheads).
In meiotically dividing spermatocytes, pachytene nuclei showedclumps of 10 bivalents. In each pachytene complement, rDNA-FISHidentified two rDNA clusters located interstitially, close to thecentromeres of both NOR-bearing bivalents (Fig. 2B) as indicated bytheir association with centromeric heterochromatin highlighted byDAPI (see open arrows in Fig. 2B). The size difference in rDNA clusters
between the chromosome pairs No. 8 and 9 was evident also inpachytene complements (Fig. 2E). In the diplotene/diakinesis stage,two bivalents showed clear, interstitially located rDNA signals(Fig. 2F).
The results of rDNA-FISH in mitotic and meiotic chromosomesclearly showed the presence of two NOR-bearing chromosome pairsin the karyotype of C. fennica. To find out whether the rDNA sitesrepresent active NORs, we applied Ag-staining which depicts NORassociated proteins denoting the activity of rRNA genes in theprevious interphase [26,46]. In most interphase nuclei, one largecompact nucleolus was observed (not shown). In meiotic spermato-cytes, two nucleoli, one larger than the other, were regularly observedfrom early prophase I nuclei until pachytene (Fig. 2G). The nucleoliwere associated eachwith a different bivalent and disappeared duringlate prophase I before reaching meiotic metaphase I. Thus, the resultsof Ag-staining proved that the two rDNA sites are active NORs.
3.3. ITS2 rDNA
Considerable intraindividual differences occurred in the ITS2ribosomal spacers of three individuals of C. fennica. A total of 15recombinant clones yielded 13 different sequence types (ribotypes).The differences between themwere partly due to polymorphism in 26nucleotides (transition/transversion ratio was 19:7) and partly due tovariable numbers of short repetitive elements that resulted indifferent lengths of the ITS variants (Table 2). Three differentrepetitive motifs, (GT)n, (GCCACC)n, and (GCA)n, were present inthe repeats, in different numbers and in different combinations,allowing assortment of the ITS2 sequences into four ITS2 variants(Table 2). Each variant was detected in multiple clones per individualand/or in more than one individual, suggesting that the differencesobserved among different variants are not PCR artefacts.
4. Discussion
The order Caryophyllidea, consisting of four families (Balanotae-nidae, Capingetidae, Caryophyllaeidae, and Lytocestidae), is presentlysubjected to extensive and complex studies using morphological,molecular, and also cytogenetic approaches [3,7,34–40]. Recentcladistic data suggest that phylogenetic relationships within theorder are only partially compatible with the traditional classificationand that probably all caryophyllidean families are paraphyletic [3,4].Chromosomal characteristics, such as the number and structure ofchromosomes, distribution of heterochromatin and chromosomallocation of particular genes or DNA sequences, provide helpfulcytogenetic markers that can help us clarify phylogenetic relation-ships and consequently taxonomic classification.
The unresolved systematics and taxonomy of the Caryophyllidea isalso illustrated by existing cytogenetic data. Except for C. fennica, eightspecies of the family Lytocestidae have been studied karyologically todate [for a review see 12,41]. Five species have the diploid number of2n=16 (Khawia iowensis, K. rossittensis, K. sinensis, K. saurogobii, andLytocestus indicus); Caryoaustralus sprenti has 2n=6 and Notolyto-cestus minor 2n=12. The last lytocestid examined is A. huronensis,that is triploid with 3n=24 [6,7]. Chromosome morphology wasexamined only in three species, A. huronensis, K. saurogobii, and K.sinensis. Their karyotypes are dominated by mono-armed chromo-somes. On the contrary, the complement of C. fennica is composedexclusively of bi-armed chromosomes (2n=20 m). It is thereforeapparent that C. fennica differs considerably from all so far studiedrepresentatives of Lytocestidae.
On the other hand, chromosome characteristics of C. fennica seemto be similar to those in members of the family Caryophyllaeidae. Fourspecies (C. laticeps, G. catostomi, G. vogei, and Monobothrium hunteri)have also the diploid number of 2n=20 and bi-armed chromosomesapparently prevail in the complements of representatives of the
Table 1Measurements (means±SD) and classification of chromosomes of Caryophyllaeides fennica.
Chromosomenumber
Absolute length(mean±SD)(μm)
Relative length(mean±SD)(%)
Centromericindex(mean±SD)
Classificationa
1 8.19±1.29 13.97±2.20 42.37±3.03 m2 7.75±1.21 13.22±2.06 43.79±1.99 m3 7.11±0.92 11.98±1.64 46.25±1.80 m4 6.51±0.88 11.04±1.54 44.38±1.92 m5 6.12±0.87 10.45±1.48 46.13±2.62 m6 5.80±1.00 9.89±1.72 44.98±2.79 m7 5.39±0.66 9.18±1.12 47.10±2.54 m8 4.76±0.69 8.13±1.17 46.02±2.67 m9 4.22±0.55 7.20±0.94 45.63±1.89 m10 2.80±0.46 4.47±1.39 45.36±2.80 m
a m — metacentric chromosome.
354 M. Orosová et al. / Parasitology International 59 (2010) 351–357
Author's personal copy
family [41]. The most similar karyotype with 10 pairs of metacentricchromosomes possesses C. laticeps, which, however, has both diploidand triploid populations throughout its geographical distribution [10].As to the two remaining caryophyllidean families, cytogenetic dataare available only in two species, Balanotaenia bancrofti (Balanotae-nidae) and Capingens singularis (Capingetidae), both with 2n=14[41].
The arguments for recent artificial family structure of theCaryophyllidea might be strengthened also by comparative analysisof the content, composition and distribution of heterochromatin andthe occurrence of secondary constrictions that are importantcytogenetic markers [42]. Relevant data are available only in threerecently studied lytocestid species, namely A. huronensis, K. saurogobii,
Fig. 2. Location of rDNA clusters in chromosome preparations from testes of Caryophyllaeides fennica as revealed by FISH with 18S rDNA probe (red signals; A–F) and visualization ofnucleoli in meiotic spermatocytes stained with AgNO3 (G). In FISH preparations, chromosomes were counterstained with DAPI (blue; A–F). (A) Mitotic metaphase consisting of2n=20 chromosomes. (B) Pachytene bivalents; note two clusters of interstitial rDNA signals associated with DAPI-highlighted block (open arrows). (C–D) Pachytene bivalent andmeiotic chromosomes of pair No. 7 showing two interstitial heterochromatin bands (open arrows). (E) Two pachytene NOR-bivalents of chromosome pair Nos. 8 and 9; note the sizedifference between rDNA clusters; also note heterochromatin associated with the rDNA clusters. (F) Diplotene. Arrows indicate the large rDNA cluster of chromosome pair No. 8,arrowheads the small rDNA cluster of pair No. 9, and asterisk pair No. 2 without 24 DAPI-positive band. (G) Pachytene complement; note two deeply stained nucleoli. Scalebar=10 μm.
Table 2Polymorphism in repetitive motifs within ITS2 of Caryophyllaeides fennica.
ITS2variant
Number and combinationof repeats in variant
Size of ITS2variant
Clones of particular ITS2varianta
A (GT)7 (GCCACC)3 (GCA)4 635 bp 1/1; 1/5B (GT)8 (GCCACC)2 (GCA)5 634 bp 2/2; 2/3; 2/4; 2/5; 3/1; 3/3C (GT)8 (GCCACC)2 (GCA)4 631 bp 2/1; 3/2; 3/4D (GT)7 (GCCACC)2 (GCA)4 629 bp 1/2; 1/3; 1/4; 3/5
a The first number is referred to the individual (1; 2; and 3), the second number to itsrespective clone (1/1–5; 2/1–5; and 3/1–5).
355M. Orosová et al. / Parasitology International 59 (2010) 351–357

Author's personal copy
and C. fennica [7,12, this paper]. In spite of their placement in onefamily, the pattern of heterochromatin differs markedly in thesespecies. In K. saurogobii, pericentromeric bands are present in allchromosomes except the chromosome pair No. 1 [12]. Similarly, onepair of chromosomes (No. 2) lacks heterochromatin in C. fennica.However, chromosomes No. 7 have two interstitial heterochromatinbands, each on a different arm (present data). In triploid A. huronensis,pericentromeric heterochromatin bands were found only in tripletsNos 2, 4, 6, and 7 [7]. In contrast to heterochromatin bands,identification of secondary constrictions as NOR-sites (i.e., clustersof rRNA genes), was found to be problematic by means of classicalstaining methods. These constrictions were observed in A. huronensisand K. saurogobii, but not in C. fennica.
The location of rDNA clusters, evidenced by rDNA-FISH, differsmarkedly in the three lytocestid species examined. Two distinctinterstitial rDNA clusters per each of three homologues of the tripletNo. 2 were identified in triploid parthenogenetic species A. huronensis[7]. Whereas a single pair of NORs, located in the pericentromericregion of the long arm of metacentric chromosomes No. 7, was foundin K. saurogobii; in addition, a size dimorphism of homologous FISHsignals was reported in this species [12]. The present study revealedthat C. fennica has two different rDNA clusters located near thecentromeres of two metacentric pairs Nos. 8 and 9. Interestingly, sizevariation of the non-homologous rDNA clusters was apparent as theFISH signals on the pair No. 8 were always stronger than those on thepair No. 9. Regardless, both rDNA clusters were found to betranscriptionally active, being positive after Ag-staining [43].
Variation in FISH signals is not a new phenomenon in animals andit is commonly observed between homologous chromosomes or evenbetween two chromatids of the same chromosome [44–48]. As rDNAconsists of highly repeated sequences arranged in tandem arrays, themass and intensity of the FISH signal could reflect the variation incopy number of rRNA genes arrayed at that site [49].
It is known that NOR regions are capable of spreading throughoutthe genome creating new rDNA loci [50,51] and that the loss orduplication of rDNA can be caused by unequal crossing over and bynonhomologous recombination [52]. Chromosomal rearrangements,i.e. translocations or inversions involving NOR-bearing chromosomes,may move rDNA repeats between or within chromosomes [53,54].Consequently, minor rDNA loci may enhance their copy number viaunequal crossing over or retrotransposition and finally becomegenome specific and detectable with rDNA-FISH as chromosomemarkers [54].
Our study showed that C. fennica has two different rDNA clusters,both associated with pericentromeric heterochromatin, but thecluster on the chromosome pair No. 9 is considerably smaller thanthat on pair No. 8.We suppose that the smaller cluster could representan evolutionary new rDNA site. The new site might result either froma structural chromosome rearrangement, for example reciprocaltranslocation of chromosome arms between the chromosomes No.8 and 9, or from transposition. The heterochromatin association ofboth rDNA loci could facilitate the rearrangement between the twochromosomes. Previous cytogenetic studies, although very scanty,revealed a high karyotypic variability in caryophyllidean species. Thissuggests that structural chromosome rearrangements were frequentwithin this order and translocations might be involved in changes ofthe position of rDNA clusters between chromosomes. However, weprefer an alternative hypothesis that the increased number of rDNAclusters in C. fennica is a result of dispersion of few rRNA genes to anew chromosomal locus by transposition, where their amplificationvia unequal crossing-over gave rise to the new NOR. The small size ofrDNA cluster is consistent with this hypothesis.
Dispersal of rDNA loci on various chromosomes (in karyotypeswithmore than one NOR) is one of the possibilities, why locus-specificITS variants are generated and maintained [16,18]. Homogenizationprocesses that are involved in concerted evolution of ribosomal RNA
genes can be less efficient and not operating uniformly in organisms,in which rDNA is present at several different NORs [17]. Intraindivi-dual polymorphism of ITS rDNA was proven to be consistent not onlywith the existence of multiple rDNA sites, but also with severalother genetic phenomena, such as polyploidy and parthenogenesis[7,16,17,55,56].
The intragenomic ITS sequence variants havemost frequently beenobserved in plants [55] and insects [57], but also in free-livingflatworms [58]. In parasitic platyhelminths, this phenomenon wasfound in trematodes of the genera Paragonimus and Schistosoma, andin cyclophyllidean (Echinococcus, Taenia) and diphyllobothriidean(Ligula) tapeworms [16–20]. Even though karyological data on someof the above species are available [59–63], these helminths have neverbeen studied along with ITS sequences in a set of individuals withinthe same population.
The only platyhelminth species in which molecular and karyolog-ical data have been acquired for the same set of specimens is A.huronensis. The species represents a unique example of organism, inwhich polyploidy, parthenogenesis, multiple NORs and intraindivi-dual ITS variants were proven to be mutually linked [6,7]. Similarly,the current data provide parallel information on molecular andgenetic peculiarities in another caryophyllidean tapeworm, C. fennica.
Comparison of the morphology of chromosomes indicates thatkaryotype of this tapeworm, which is currently placed in theLytocestidae, is more closely related to representatives of theCaryophyllaeidae than to lytocestid species. However, much morekaryological information on other species, representing variousfamilies, is necessary to obtain to elucidate taxonomic problems andto reveal trends of chromosome evolution within the order Caryo-phyllidea. Although karyological data have not belonged to frequentlyusedmarkers in cestode systematics and phylogeny, the present studyindicates that detailed cytogenetic studies may provide valuableinformation and should be applied in future studies on cestodes.
Acknowledgements
We wish to thank František Marec (Institute of Entomology,Biology Centre ASCR, České Budějovice, Czech Republic) for criticalreading of the manuscript and to Tomáš Scholz (Institute ofParasitology, Biology Centre ASCR, České Budějovice, Czech Republic)for valuable comments. This work was supported by grants of theGrant Agency of the Slovak Republic (VEGA 2/0148/09 and 2/0014/10), the National Science Foundation, USA (PBI award Nos. 0818696and 0818823), and the Grant Agency of the Czech Republic (No. 524/08/0885), from the research projects of the Institute of Parasitology BCASCR (Z60220578, LC 522) and the project Centre of Excellence forParasitology (Code ITMS: 26220120022) based on the support of theOperational Programme “Research & Development” funded from theEuropean Fund of Regional Development (rate 0.1).
References
[1] Mackiewicz JS. Order Caryophyllidea van Beneden in Carus, 1863. In: Khalil LF,Jones A, Bray RA, editors. Keys to the cestode parasites of vertebrates. Wallingford,Oxon: CAB International; 1994. p. 21–43.
[2] Mackiewicz JS. Caryophyllidea (Cestoidea): molecules, morphology and evolution.Acta Parasitol 2003;48:143–54.
[3] Olson PD, Scholz T, Poddubnaya LG, Littlewood DTJ. On the derived position ofArchigetes and the early evolution of the tapeworms (Platyhelminthes: Cestoda).J Parasitol 2008;94:898–904.
[4] Oros M, Hanzelová V, Scholz T, Mackiewicz JS. Phylogenetic relationships of themonozoic tapeworms (Eucestoda: Caryophyllidea) inferred from morphologicalcharacters. Syst Parasitol 2008;70:1–14.
[5] Waeschenbach A, Webster BL, Bray RA, Littlewood DT. Added resolution amongordinal level relationships of tapeworms (Platyhelminthes: Cestoda) withcomplete small and large subunit nuclear ribosomal RNA genes. Mol PhylogenetEvol 2007;45:311–25.
[6] Jones AW, Mackiewicz JS. Naturally occurring triploidy and parthenogenesis inAtractolytocestus huronensis Anthony (Cestoidea: Caryophyllidea) from Cyprinuscarpio L. in North America. J Parasitol 1969;55:1105–18.
356 M. Orosová et al. / Parasitology International 59 (2010) 351–357

Author's personal copy
[7] Kráľová-Hromadová I, Štefka J, ŠpakulováM, OrosováM, BombarováM, HanzelováV, et al. Intraindividual ITS1 and ITS2 ribosomal sequence variation linked withmultiple rDNA loci: a case of triploid Atractolytocestus huronensis, the monozoiccestode of common carp. Int J Parasitol 2010;40:175–81.
[8] Grey AJ, Mackiewicz JS. Chromosomes of the caryophyllidean cestodes: diploidy,triploidy andparthenogenesis inGlaridacris catostomi. Int J Parasitol 1980;10:397–407.
[9] Mackiewicz JS. Caryophyllidea (Cestoidea): evolution and classification. AdvParasitol 1981;19:139–206.
[10] Petkevičiūté R, Kuperman BI. Karyological investigation of Caryophyllaeus laticeps(Pallas, 1781) (Cestoda: Caryophyllidea). Folia Parasit 1992;39:115–21.
[11] Bombarová M, Marec F, Nguyen P, Špakulová M. Divergent location of ribosomalgenes in chromosomes of fish thorny-headed worms, Pomphorhynchus laevis andPomphorhynchus tereticollis (Acanthocephala). Genetica 2007;131:141–9.
[12] Orosová M, Marec F, Oros M, Xi BW, Scholz T. A chromosome study and 18S rDNAin situ hybridization of Khawia saurogobii (Cestoda: Caryophyllidea). Parasitol Res2010;106:587–93.
[13] Berzunza-Cruz M, Bricaire G, Suazo N, Perez-Montfort R, Becker I. PCR foridentification of species causing American cutaneous leishmaniasis. Parasitol Res2009;104:691–9.
[14] Gasser RB, Cantacessi C, Campbell BE. Improved molecular diagnostic tools forhuman hookworms. Expert Rev Mol Diagn 2009;9:17–21.
[15] Hillis DM, Dixon MT. Ribosomal DNA: molecular evolution and phylogeneticinference. Q Rev Biol 1991;66:411–53.
[16] van Herwerden L, Blair D, Agatsuma T. Intra- and inter-specific variation in nuclearribosomal internal transcribed spacer 1 of the Schistosoma japonicum speciescomplex. Parasitology 1998;116:311–7.
[17] van Herwerden L, Blair D, Agatsuma T. Intra- and interindividual variation in ITS1of Paragonimus westermani (Trematoda: Digenea) and related species: implica-tions for phylogenetic studies. Mol Phylogen Evol 1999;12:67–73.
[18] van Herwerden L, Gasser RB, Blair D. ITS-1 ribosomal DNA sequence variants aremaintained in different species and strains of Echinococcus. Int J Parasitol 2000;30:157–69.
[19] Hancock K, Broughel DE, Moura INS, Khan A, Pieniazek NJ, Gonzales AE, et al.Sequence variation in the cytochrome oxidase I, internal transcribed spacer 1, andTs14 diagnostic antigen sequences of Taenia solium isolates from South andCentral America, India and Asia. Int J Parasitol 2001;31:1601–7.
[20] Bouzid W, Štefka J, Hypša V, Lek S, Scholz T, Legal L, et al. Geography and hostspecificity: two forces behind the genetic structure of the freshwater fish parasiteLigula intestinalis (Cestoda: Diphyllobothriidae). Int J Parasitol 2008;38:1465–79.
[21] Nolan JM, Cribb TH. The use and implications of ribosomal DNA sequencing for thediscrimination of digenean species. Adv Parasitol 2005;60:101–63.
[22] Protasova EP, Kuperman BI, Roitman VA, Poddubnaya LG. Caryophyllid tapewormsof the fauna of USSR. Moscow: Nauka; 1990 (In Russian).
[23] Oros M, Hanzelová V. Re-establishment of the fish parasite fauna in the Tisa Riversystem (Slovakia) after a catastrophic pollution event. Parasitol Res 2009;104:1497–506.
[24] Frydrychová R, Marec F. Repeated losses of TTAGG telomere repeats in evolutionof beetles (Coleoptera). Genetica 2002;115:179–87.
[25] Levan A, Fredga K, Sandberg A. Nomenclature for centromere position onchromosomes. Heredita 1964;52:201–20.
[26] Howell WM, Black DA. Controlled silver staining of nucleolus organizer regionswith a protective colloidal developer: a 1-step method. Experientia 1980;36:1014–5.
[27] Littlewood DTJ, Olson PD. Small subunit rDNA and the Platyhelminthes: signal,noise, conflict and compromise. In: Littlewood DTJ, Bray RA, editors. Interrelation-ships of the Platyhelminthes. London: Taylor and Francis; 2001. p. 262–78.
[28] Sahara K, Marec F, Traut W. TTAGG telomeric repeats in chromosomes of someinsects and other arthropods. Chromosome Res 1999;76:449–60.
[29] Fuková I, Nguyen P, Marec F. Codling moth cytogenetics: karyotype, chromosomallocation of rDNA, and molecular differentiation of sex chromosomes. Genome2005;48:1083–92.
[30] Králová-Hromadová I, Scholz T, Shinn AP, Cunningham CO, Wootten R, HanzelováV, et al. A molecular study of Eubothrium rugosum (Batsch, 1786) (Cestoda:Pseudophyllidea) using ITS rDNA sequences, with notes on the distribution andintraspecific sequence variation of Eubothrium crassum (Bloch, 1779). ParasitolRes 2003;89:473–9.
[31] Cunningham CO. Species variation within the internal transcribed spacer (ITS)region of Gyrodactylus (Monogenea: Gyrodactylidae) ribosomal RNA genes.J Parasitol 1997;83:215–9.
[32] Olson PD, Littlewood DT, Griffiths D, Kennedy CR, Arme C. Evidence for the co-existence of separate strains or species of Ligula in Lough Neagh, Northern Ireland.J Helminthol 2002;76:171–4.
[33] Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity ofprogressive multiple sequence alignment throughout sequence weighting,position specific gap penalties and weight matrix choice. Nucl Acids Res1994;22:4673–80.
[34] Ibraheem MH, Mackiewicz JS. Scolex development, morphology and mode ofattachment of Wenyonia virilis Woodland, 1923 (Cestoidea, Caryophyllidea). ActaParasitol 2009;51:51–8.
[35] Oros M, Hanzelová V. The morphology and systematic status of Khawia rossittensis(Szidat, 1937) and K. parva (Zmeev, 1936) (Cestoda: Caryophyllidea), parasites ofcyprinid fishes. Syst Parasitol 2007;68:129–36.
[36] Oros M, Hanzelová V, Scholz T. Tapeworm Khawia sinensis: review of theintroduction and subsequent decline of a pathogen of carp, Cyprinus carpio. VetParasitol 2009;164:217–22.
[37] Bruňanská M, Drobníková P, Oros M. Vitellogenesis in the cestode Atractolytoces-tus huronensis Anthony, 1958 (Caryophyllidea: Lytocestidae). Parasitol Res2009;105:647–54.
[38] Świderski Z, Młocicki D, Mackiewicz JS, Miquel J, Ibraheem MH, Bruňanská M.Ultrastructure and cytochemistry of vitellogenesis in Wenyonia virilis Woodland,1923 (Cestoda, Caryophyllidea). Acta Parasitol 2009;54:131–42.
[39] Xi BW, Oros M, Wang GT, Wu SG, Gao D, Nie P. Khawia saurogobii n. sp. (Cestoda:Caryophyllidea) from freshwater fish Saurogobio spp. (Cyprinidae) in China.J Parasitol 2009;95:1516–9.
[40] Oros M, Scholz T, Hanzelová V, Mackiewicz JS. Scolex morphology of monozoiccestodes (Caryophyllidea) from the Palaearctic Region: a useful tool for speciesidentification. Folia Parasit 2010;57.
[41] Petkevičiūté R. Knowledge of the chromosome set of cestodes. In: Galkin AK,Dubinina HV, editors. The problems of cestodology II. St. Petersburg: ZoologicalInstitute, Russian Academy of Sciences; 2002. p. 206–20. In Russian.
[42] Zuckerkandl E, Hennig W. Tracking heterochromatin. Chromosoma 1995;104:75–83.
[43] Hubbel HR. Silver staining as an indicator of active ribosomal genes. Stain Technol1985;60:285–94.
[44] Libertini A, Colomba MS, Vitturi R. Cytogenetics of the amphipod Jassa marmorata(Corophioidea: Ischyroceridae): karyotype morphology, chromosome banding,fluorescent in situ hybridization, and nuclear DNA content. J Crustacean Biol2000;20:350–6.
[45] Wang Y, Ximing G. Chromosomal mapping of major ribosomal rRNA genes in thehard clam (Mercenaria mercenaria) using fluorescence in situ hybridization. MarBiol 2007;150:1183–9.
[46] Amaro-Ghilardi RC, de Jesus Silva MJ, Trefaut Rodrigues MC, Yonenaga-Yassuda Y.Chromosomal studies in four species of genus Chaunus (Bufonidae, Anura):localization of telomeric and ribosomal sequences after fluorescence in situhybridization (FISH). Genetica 2007;134:159–68.
[47] Garcia C, Filho OM. Localization of ribosomal genes in three Pimelodus species(Siluriformes, Pimelodidae) of the São Francisco River: 5S genes as speciesmarkers and conservation of the 18S rDNA sites. Genet Mol Biol 2008;31:261–4.
[48] Citrini A, Simonazzi G, Cassanelli S, Ferrari M, Bizzaro D, Manicardi GC.Distribution of heterochromatin and rDNA on the holocentric chromosomes ofthe aphids Dysaphis plantaginea andMelanaphis pyraria (Hemiptera: Aphilididae).Eur J Entomol 2009;106:153–7.
[49] Cerbah M, Souza-Chies T, Jubier MF, Lejeune B, Siljak-Yakovlev S. Molecularphylogeny of the genus Hypochaeris using internal transcribed spacers of nuclearrDNA: inference for chromosomal evolution. Mol Biol Evol 1998;15:345–54.
[50] Castro J, Rodríguez S, Pardo BG, Sánchez L, Martínez P. Population analysis of anunusual NOR-site polymorphism in brown trout (Salmo trutta L.). Heredity2001;86:291–302.
[51] Dubcovsky J, Dvořák J. Ribosomal RNA multigene loci: nomads of the Triticeaegenomes. Genetics 1995;140:1367–77.
[52] Guillén AK, Hirai Y, Tanoue T, Hirai H. Transcriptional repression mechanisms ofnucleolus organizer region (NORs) in humans and chimpanzees. Chromosom Res2004;12:225–37.
[53] Hanson RE, Islam-Faridi MN, Percival EA, Crane ChF, Ji Y, McKnight TD, et al.Distribution of 5S and 18S–28S rDNA loci in a tetraploid cotton (Gossypiumhirsutum L.) and its putative diploid ancestors. Chromosoma 1996;105:55–61.
[54] Chung MCh, Lee YI, Cheng YY, Chou YJ, Lu ChF. Chromosomal polymorphism ofribosomal genes in the genus Oryza. Theor Appl Genet 2008;116:745–53.
[55] Rosselló JA, Lázaro A, Cosín R, Molins A. A phylogenetic split in Buxus balearica(Buxaceae) as evidenced by nuclear ribosomal markers: when ITS paralogues arewelcome. J Mol Evol 2007;64:143–57.
[56] D'Alelio D, Amato A, KooistraWHCF, Procaccini G, Casotti R, Montresor M. Internaltranscribed spacer polymorphism in Pseudo-nitzschia multistriata (Bacillariophy-ceae) in the Gulf of Naples: recent divergence or intraspecific hybridization?Protist 2009;160:9–20.
[57] Fairley TL, Kilpatrick CW, Conn JE. Intragenomic heterogeneity of internaltranscribed spacer rDNA in Neotropical malaria vector Anopheles aquasalis(Diptera: Culicidae). J Med Entomol 2005;42:795–800.
[58] Carranza S, Baguna J, Riutort M. Origin and evolution of paralogous rRNA geneclusters within the flatworm Dugesiidae (Platyhelminthes, Tricladida). J Mol Evol1999;49:250–9.
[59] Hirai H. Paragonimus ohirai: identification of Nucleolar Organizer Regions (NORs)and silver nitrate staining pattern in spermatogenesis. Exp Parasitol 1988;67:281–6.
[60] Hirai H, LoVerde PT. Triploid cells found in intramolluscan stages of Schistosomamansoni. J Parasitol 1989;75:800–2.
[61] Hirai H, Spotila LD, LoVerde PT. Schistosoma mansoni: chromosomal localization ofDNA repeat elements by in situ hybridization using biotinylated DNA probes. ExpParasitol 1989;69:175–88.
[62] Petkevičiuté R. Comparative cytogenetics of Diphyllobothrium ditremum (Creplin,1925) and Ligula intestinalis (Linnaeus, 1758) (Cestoda: Pseudophyllidea). SystParasitol 1992;23:167–73.
[63] Štefka J, Hypša V, Scholz T. Interplay of host specificity and biogeography inpopulation structure of a cosmopolitan endoparasite: microsatellite study ofLigula intestinalis (Cestoda). Mol Ecol 2009;18:1187–206.
357M. Orosová et al. / Parasitology International 59 (2010) 351–357



![[Compared parasitic infection of Ligula intestinalis (Cestoda: Diphyllobothridae) in Cyprinidae species: Rutilus rubilio and Scardinius erythrophthalmus in two dam reserves in Tunisia]](https://img.pdfslide.net/doc/110x75/634ec2490d6acb880b02f6c2/compared-parasitic-infection-of-ligula-intestinalis-cestoda-diphyllobothridae.jpg)

























