Embed Size (px)
Citation preview

410-419 Clinical rheumatology, 1985, 4, N ~
Enhanced thromboxane synthesis and vacuolization in human polymorphonuclear leucocytes induced by human lymphokine containing supernatants
P . C O N T I , *M. A L L E G R E T T A , * * T . W . W I L L I A M S , * * * M . G . C I F O N E , ***E. A L E S S E , M. R E A L E , ***E. * R . A . D E M P S E Y
B O I D I ,
Cattedra di Immunologia, Istituto di Medicina Sperimentale, FacoltA di Medicina, via dei Vestini, 66100 Chieti, Italy. *Department of Medicine, Division of Hematol- ogy/Oncology, Experimental Medicine, TUFTS University, School of Medicine, Boston MA 02111, USA. **Immunology Center, Georgetown University, Washing- ton D.C. USA. ***Cattedra di Patologia Generale, FacoltA di Medicina, Colle- maggio, 67100 L'Aquila, Italy.
SUMMARY P M N ' s were cultivated in vitro and treated with supernatants obtained f r o m mitogenic-induced lymphocytes o f human tonsil. Cytoplasmic vacuolization increased with time and there was a lower number o f neutrophilic clumps in the culture treated with a supernatant containing lymphotoxin. In addition, the P M N ' s released more thromboxane B e which was inhibited by indomethacin. We conclude that the action o f lymphotoxin on the target P M N ' s is not mediated by the production o f thromboxane B2.
Key words : Polymorphonuclear, Thromboxane, Lymphocyte, Lymphokine, Lymphotoxin, Vacuol.
INTRODUCTION
Lymphocytes can be stimulated in vitro by a variety of specific and non-specific agents, (i.e. phytohemoagglutinin [PHA]) which in turn induce the release of different sub- stances into the cultura media (1). These substances are both demonstrable in vitro (2), and are generically called lymphokines
Received 29 October 1984, Revision - accepted 6 March 1985 Correspondence to : DR. P10 CONTI, Cattedra di Irnmunologia, Istituto di Medicina Speri- rnentale, Facolt~t di Medicina e Chirurgia, Universith di Chieti, Via dei Vestini, 66100 Chieti, Italy.
(LK). LK have a wide variety of biological activities, which vary from cell activation i.e. Macrophage Activation Factor (MAF), to cell lysis i.e. Lymphotoxins (LTs)(3). Considerable information has accumulated, although the activation of these substances on target cells is not completely understood (4). LT consist of four distinct types (5,6). The cytotoxic activity of these substances (7) is mostly non-reversible and is exerted di- rectly on the cell membrane and DNA syn- thesis (8,9). This may, in part, involve the activation of phospholipase A, since guinea pig LT has been reported to activate phos- pholipase A in target cell membranes which is associated with the increase in intracellu-

Human lymphokine and polymorphonuclear leucocytes" 411
lar Ca2 + and subsequent cell lysis (10,11). Further, this activity appears to induce cell vacuolation (12).
A wide variety of cells are reported to be the target of LK such as macrophages mono- cytes, epithelial cells, etc. Recently neutrophils have been shown to interact with supernatant f rom mitogen-stimulated mono- nuclear cells which resulted in increases in both respiratory and phagocytic activity (13). The present study involves the determi- nation of the effect o f supernatants obtained from mitogen-induced lymphocytes of hu- man tonsil on human peripheral blood neu- trophils (PMN's) . The production of LT's was used as a marker for lymphokine pro- duction. These levels were correlated with morphological , functional and biochemical changes in the neutrophils which were mea- sured by cell vacuolation, cellular clumping and thromboxane synthesis. Indomethacin has been shown to inhibit cyclo-oxigenase products and in some pathological cases, can improve cellular immunity (14). We, therefore, evaluated the effect of this drug.
MATERIALS AND M E T H O D S
Preparation of human lymphocytes : Human lymphocytes were obtained f rom tonsil tissue immediately after surgical removal f rom healthy young children. The tissue was rinsed three times with RPMI 1640 medium supplemented with 10070 heat-inactivated fe- tal calf serum, 100ttg/ml streptomycin, 100U/ml penicillin, and 25/zg/ml fungi- zone. This medium was used throughout the study. The tissue was minced into frag- ments. Suspension of 9907o viable small lym- phocytes were obtained after the following differential centrifugation technique : 10ml of cell suspension was placed in 25 x 145 'mm screw capped tubes and centrifuged for 1 min at 75 x g in an International PR-2 re- frigerated centrifuge at 4~
The supernatant was transferred to an- other tube and spun for 1 min at 135 z g. The cells remaining in this supernatant were
pelleted by final centrifugation at 400 x g for 3 min. The cell pellet was resuspended in 1.0-5.0 ml medium and viable cell count de- termined by the cell's ability to exclude 0.1070 eosin Y. Lymphotoxin containing su- pernatant is prepared as follows: lympho- cytes prepared as above were cultured in 30ml volumes of 1640 medium at a concen- tration of 5 x 10 6 cells/ml in 250 ml plastic tissue culture flasks. The cells were activated by the addition of 20/zg/ml PHA-p (Difco). Control medium received no PHA. The cells were incubated at 37~ in a humidified at- mosphere of 9507o air-5070 CO2 for 120 hrs, after which the medium was cleared of cells and debris by centrifugation at 800 g for 30 min. P H A to 20#g/ml was then added to the control medium, and all supernatants were stored at - 2 0 ~ until use.
Lymphotoxin assay: LT activity was assay- ed by establishing 1 x 104 mouse L-929 cells in 200#1 1640 medium plus 0 .5#g /ml mito- mycin-C in the wells of a microtiter plate. After 24 hr incubation at 37~ the medium was aspirated off, the cells washed with 100#l of Hanks Balanced Salt Solution (HBSS), and 200td of various dilutions of LT and control medium were added. After 18 hrs incubation at 37~ the cells were ob- served microscopically and the pulse labelled for their ability to incorporate 3H-amino acid into cellular protein in the following manner : the medium was aspirated of f the monolayers, washed once with 200#1 warm HBSS and replaced with 100/zl warm (37~ labelling medium which consisted of HBSS supplemented with 2~ dialyzed fetal calf se- rum and 5Ci/ml 3H-amino acid mixture.
The cultures were incubated at 37~ for 30 min, after which the wells were harvested on fiberglass filters on a Bellico Micro- harvester using distilled water. The filters were dried in air and placed in 2 ml toluene scintillation fluid containing 4 g / 1 0 m n i - fluor, and counted in a Beckman scintil- lation counter. Killing percent of target L cells is calculated as follows : 070killing=

412 P. Conti , M. Allegretta, T.W. Williams, et al.
100 • (1 - c p m ( + P H A / c p m ( - P H A ) ) .
LT ac t iv i ty in u n i t s / m l is d e f i n e d as the reci-
p roca l o f the h ighes t d i l u t i on wh ich causes
50~ cell ki l l ing. T h e p resen t s t udy u t i l i zed
an L T p r e p a r a t i o n c o n t a i n i n g 50 U / m l .
Collection of blood and isolation of poly- morphonuclear leucocyte ( P M N ' s ) : B l o o d
was o b t a i n e d f r o m 4 hea l t hy adu l t d o n o r s
and co l l ec t ed in p o l y p r o p y l e n e tubes
c o n t a i n i n g su f f i c i en t h e p a r i n to o b t a i n a fi-
na l c o n c e n t r a t i o n o f 20 U / m l . Tes ts were
p e r f o r m e d i m m e d i a t e l y a f t e r co l l ec t ion .
P M N ' s were i so l a t ed by the f o l l o w i n g m e t h -
o d : b l o o d was c e n t r i f u g e d o v e r F ico l l -
H y p a q u e in 2070 d e x t r a n fo r 30 m i n u t e s at
r o o m t e m p e r a t u r e (15). A f t e r w a s h i n g the
cells t w o t imes wi th b u f f e r e d sa l ine , t he r ed
b l o o d cells were lysed by dis t i l led wa te r , t he
cells were w a s h e d and r e s u s p e n d e d in G e y ' s
b u f f e r a n d v iab i l i ty c h e c k e d by the t rypan~
b leu e x c l u s i o n dye m e t h o d . T h e cells were
i n c u b a t e d at a dens i ty o f 106 pe r ml at 3 7 ~
in t r ip l ica tes . T h e cells cons i s t ed o f m o r e
t h a n 95~ P M N ' s . S o m e samples were p re in -
c u b a t e d fo r 10 m i n wi th i n d o m e t h a c i n , in a
f inal c o n c e n t r a t i o n o f 50 n g / m l pe r 106 ceils
in a 3 7 ~ w a t e r ba th . S a m p l e a n d c o n t r o l s
were t r e a t ed wi th 200#1 o f s u p e r n a t a n t f r o m
l y m p h o c y t e cu l tu re s o r f resh m e d i u m a n d
i n c u b a t e d at 3 7 ~ fo r d i f f e r e n t p e r i o d s (see
T a b l e I). A l i q u o t s o f 50/zl o f cell s u p e r n a -
Table 1 Treatment o f PMNs from stimulated and non-stimulated lymphocytes
Time of incubation Control Control Control Sample Sample Sample Sample
(min) 1 2 3 1 2 3 4
0 0
30 0
60
120
180
240
0 0 0 0 0 0
0.5 6.0 13.5 11.3 11.0 4.0 _ +_ _+ • __. _
0.4 2.5 4.5 2.1 4.3 1.0
0.5 10 7.8 31.5 12.3 18.3 16.5 ___ + • ___ ___ • •
0.3 7.1 7.1 11.3 4.5 8.0 11.5
8.4 22 23 57 36.5 35 38 __. _+ • +_ __. _ •
5.0 9.0 8.9 18.1 13.5 15.5 12
9.3 41 30.6 77.4 45.6 56.3 71.5 _+ __. • + _+ • ___
8.9 11 18.1 16.2 23.5 13.6 22.5
21.5 47 48.8 98 84.3 74.3 97 • _-+ __. + • • __.
11.6 13 18.4 12.5 10.2 4.1 11
The mean and the standard error in percentage values of the cells (PMN's) showing vacuoles after incubation for different times in four experiments (counted on slides after cytocentrifugation). Control 1 = Supematants from unstimulated lymphocytes Control 2 = Fresh medium plus PHA Control 3 = Supernatants from unstimulated lymphocytes plus PHA Sample 1 = Supernatants from stimulated lymphocytes with PHA Sample 2 = Supernatants from stimulated lymphocytes with PHA diluted 1 § 1 with supernatants control 1 Sample 3 = Supernatants from stimulated lymphocytes with PHA diluted 1 § 2 with supernatants control 1 Sample 4= Supernatants from stimulated lymphocytes with PHA plus indomethacin.

Human lymphokine and polymorphonuclear leucocytes 413
t a n t were t a k e n at v a r i o u s t i m e s a n d s m e a r s
w e r e p r e p a r e d by c y t o c e n t r i f u g a t i o n .
S i m u l t a n e o u s l y , s a m p l e s o f 25/~1 were
t a k e n a n d a d h e r e n c e to g lass was e v a l u a t e d
a c c o r d i n g to a p r e v i o u s l y d e s c r i b e d p r o c e -
d u r e (16). Al l t he s m e a r s were s t a i n e d wi th
G i e m s a . T h e p e r c e n t o f v a c u o l a t e d cells was
e v a l u a t e d by counting 200 P M N ' s . T h e
n u m b e r o f smal l (15 to 100) or l a rge ( m o r e
t h a n 100) P M N c l u m p s were a l so r e c o r d e d .
T h r o m b o x a n e - B 2 (TXB2) was d e t e r m i n e d
u n d e r c o n d i t i o n s d e s c r i b e d p r e v i o u s l y by
Table 1I Thromboxane-B e production by PMN's
Treatment TXB 2 ng/ml • 106 cells in two separate experiments
Experiment No 1 _+ SEM No 2 +_ SEM S.L. S.L.
CONTROL 1 Supernatant from non-stimu- lated lymphocytes
CONTROL 2 Fresh medium plus fresh PHA
CONTROL 3 Supernatants from non-stimu- lated lymphocytes plus fresh PHA
SAMPLE 1 Supernatants from stimulated lymphocytes with PHA
SAMPLE 2 Supernatants from stimulated lymphocytes with PHA diluted 1 § 1 with supernatants Control 1
SAMPLE 3 Supernatants from stimulated lymphocytes with PHA diluted 1 + 2 with supernatants Control 1
SAMPLE 4 Indomethacin 50 ng/ml plus supernatants from stimulated lymphocytes
SAMPLE 5 Indomethacin 50 ng/ml plus fresh medium plus fresh PHA
0.384 -+ 0.008 - - 0.621 _+ 0.05
0.442 _+ 0.009 p < 0.01 0.599 +_ 0.005 N.S.
0.421 + 0.03 N.S. 0.578 _+ 0.009 N.S.
0.462 +_ 0.006 p < 0.01 0.745 +_ 0.01 p < 0.01
0.409 + 0.02 N.S. 0.658 + 0.05 N.S.
0.417 -+ 0.01 N.S. 0.647 _+ 0.02 N.S.
0.378 _+ 0.06 N.S. 0.415 _+ 0.009 p < 0.05
0.425 _+ 0.03 N.S. 0.428 _+ 0.02 p < 0.01
TXB 2 production by PMNs after 30 min incubation (in ng/ml • 106 cells) : median and standard error of vacuoles as measured in 2 experiments.
414 P, Conti, M. Allegretta, T.W. Williams, et al.
Jakschik (17). The incubationa were made in triplicate at a concentration of 10 6 cells per ml. Indomethacin was added to the tubes containing a final concentration of 50 ng/ml, and were incubated for 10 min at 37~ The incubation system was prepared as indicated in Table lI and further incubat- ed for 30 rain. Then, the concentration of indomethacin was increased to 200 #g/ml to stop the reaction, and the samples were cen- trifuged. TXB2 concentration was deter- mined by radioimmunoassay on the culture supernatants (18).
RESULTS
The mean values -!-_ SEM of the percent of PMN's presenting vacuolization when ex- posed to different supernatants of cultures and controls are presented in Table I. There was no P M N vacuole format ion at t ime zero. Af t e r 30 rain incubation at 37~ va- cuole format ion began, the highest percent (13.5+_4.5) was found in sample 1 group, corresponding to the supernatant of lym- phocyte culture stimulated with PHA. After
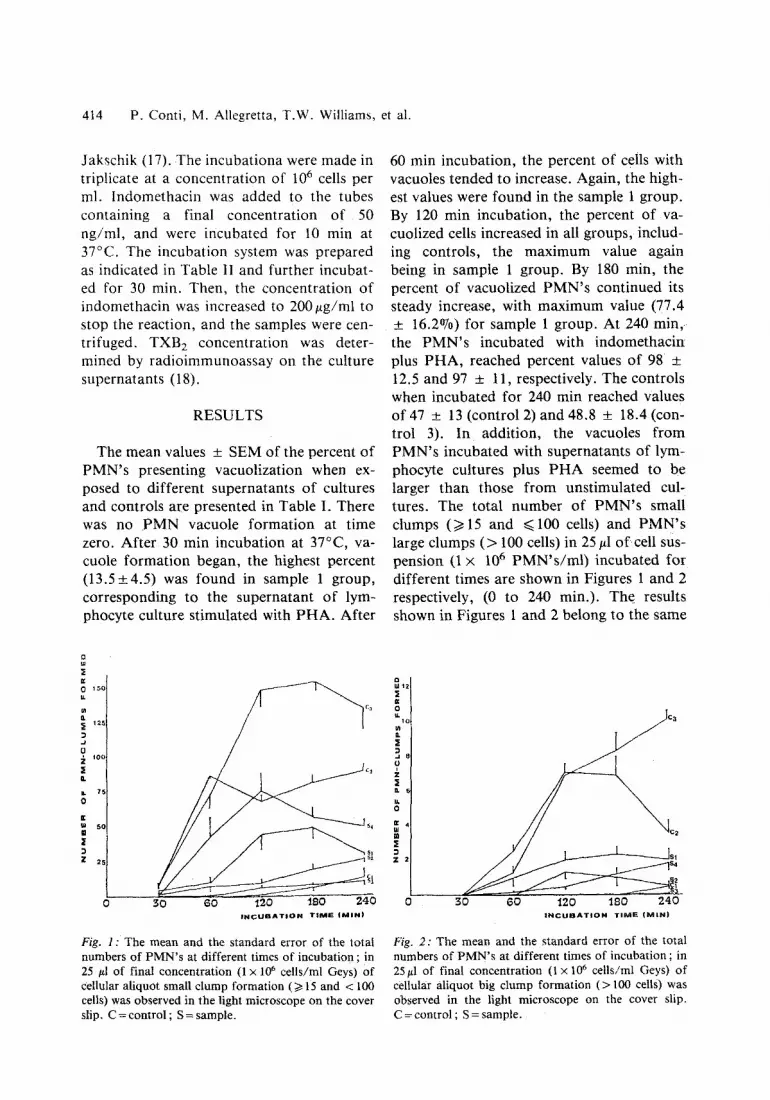
60 min incubation, the percent of cells with vacuoles tended to increase. Again, the high- est values were found in the sample 1 group. By 120 min incubation, the percent of va- cuolized cells increased in all groups, includ- ing controls, the maximum value again being in sample 1 group. By 180 min, the percent of vacuolized P M N ' s continued its steady increase, with maximum value (77.4 + 16.2%) for sample 1 group. At 240 m i n , the P M N ' s incubated with indomethacin plus PHA, reached percent values of 98 :t: 12.5 and 97 _+ 11, respectively. The controls when incubated for 240 min reached Values of 47 +_ 13 (control 2) and 48.8 + 18.4 (con- trol 3). I n addition, the vacuoles f rom PMN's incubated with supernatants of lym- phocyte cultures plus P H A seemed to be larger than those f rom unstimulated cul- tures. The total number of P M N ' s small clumps (>t15 and ~<100 cells) and P M N ' s large clumps ( > 100 cells) in 25 #1 o f cell sus- pension (1 x 106 P M N ' s / m l ) incubated for different times are shown in Figures 1 and 2 respectively, (0 to 240 min.). The results shown in Figures 1 and 2 belong to the same
150 r
,~ 12s
J 0 ;~ 1oo
o.
0
r s o
Z 25
't
30 60 120 ~B0 240 I N C U B A T I O N T I M E ( M I N )
Fig. 1 : The mean and the standard error of the total numbers of P M N ' s at different times of incubation ; in 25 #1 of final concentration (1 • 106 cells/ml Geys) of cellular aliquot small c lump formation (/> 15 and < 100 cells) was observed in the light microscope on the cover slip. C = control ; S = sample.
i 12
r 2
0
2 o.
m
Z
r
C2
$1 4
L 0 30 60 120 "180 240
I N C U B A T I O N T I M E (~R IN)
Fig. 2: The mean and the s tandard error of the total numbers of P M N ' s at different times of incubation ; in 25#1 of final concentration (1 x 106 cells/ml Geys) of cellular aliquot big clump formation ( > 100 cells) was observed in the light microscope on the cover slip. C = control ; S = sample.

Human lymphokine and polymorphonuclear leucocytes 415
experimental group; at time zero, no va- cuoles were seen either in sample groups or in controls. After 30 min incubation, only control 1 had no clumps while in all the other groups the counts were not higher than 10 small clumps, without any large clumps. After one hour incubation, only PMN incu- bated with supernatants plus fresh PHA were significantly clumped, forming either small or large clumps (Figures 1 and 2). Samples containing lymphotoxin showed only a slight ability to form PMN clumps ; this ability decreased with the dilution of the sample. The sample containing indometha- cin ($4) had intermediate values. After 120 min incubation, control C 2 did not increase the number of small clumps at 60 rain, while the large clump count was increased (Fig. 2). Clump formation induced by S~ was always lower than those for S 2 and $3, either for small or large clumps. The number of clumps did not vary in any of the sample or control group after further incubation (240 min, total incubation time). Production of
TXB 2 by PMN's in culture was measured and the results are shown in Table II. Super- natants of lymphocyte cultures stimulated with P H A induce a significant release of TXB 2 by PMN's . This ability was abolished by preincubation for 10 rain with indome- thacin. No significant increases were found in TXB 2 concentration with respect to con- trol 1 when supernatants of lymphocyte cul- tures were diluted, or when controls 2 and 3 were tested.
DISCUSSION
Supernatants from cultures of stimulated lymphocytes contain a variety of active sub- stances, i.e. LK. This study reports that su- pernatants of lymphocyte cultures stimulat- ed with P H A induce PMN's vacuole for- mation, and that this vacuolation is dose and time dependent. There are at least two possible mechanisms which may be involved in this vacuolation. The first mechanism may be related to PHA since it is known that
Fig. 3: Control l : PMNs show no vacuol and no clumping formation after incubation for 2 hr at 37~ (light microscope 10 x).

416 P. Cont i , M. Al legret ta , T . W . Wi l l i ams , et al.
Fig. 4." Sample 1 : a) PMNs showing vacuole-formation after treatment with supernatant from lymphocytes stimu- lated with P H A for 2 hr at 37~ (light microscope 40 x ) .
] i i i i i ~ ] ] i i ] i i ] i i i i ] ] ~ ] i i ] i ] ] ] ] i ] ] ] ] ] i i ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] ] � 8 4 ~ ~ ~ ~ ~ ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~ : ~
i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 ~ i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i i i i �84184 i i i i i i i i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84 i �84 i �84184184184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84184184 i i i i i i i i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84184184 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 ~ i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 ~ i �84 i �84 i �84 i: i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i: i �84 i �84 i �84 i �84 i �84 i �84 i �84 i �84 i: i �84 ~: i �84 i �84 i �84 ~: i: ~: i �84 i: i: i: ~: i: i: i: i: i: i: i: ~: ~: i: i: i: i: i: i: i: i: i: i: i: i: i: i: i: i �84 i: i �84 i �84 i �84 i: ~ i: i: ~ i �84 ~ i �84 i
~!~i~i~ii~i~ii~iJ~i~!iJ~!~!~!iJ~!~i~i~i~i~i~i~!~iii~i~!~i~i~ii~i~ii~i~i~i~i!~i~i!~i~]~!~i~i~!!~!~i~i~ii~i~i~i~i~iiiiiiii!~i~!~! �84 b) One PMN in the center of the picture shows great vacuoles (after same treatment as a), light microscope, 100 x ).

Human lymphokine and polymorphonuclear leucocytes 417
r ~
o
, e ~ ~
Fig. 5: Sample 1: PM N' s treated with supernatants from lymphocytes stimulated with P H A for 2 hr at 37~ small c lump formation was found (>t 15 and ~< 100 cells) in light microscope (10 x ) .
Fig. 6." Control 3 : PMNs treated with supernatants from non-stimulated lymphocytes with addition of fresh P H A for 2 hr at 37~ : great clumps were found in light microscope (10 x ) .

418 P. Conti , M. Allegretta, T.W. Williams, et al.
lectins can induce cell vacuolation (19,20). The second may involve LT since this has been shown to induce vacuolation in target cells (12). The present study suggests that both mechanisms may be involved since there is an increase in PMN vacuolation in the presence of supernatants f rom stimulat- ed lymphocytes. This is in contrast to what is seen when PMN's are incubated with con- trol supernatants f rom non-stimulated lym- phocytes and fresh PHA. These results sug- gest a distinct enhancement of cell vacuol- lation which may be attributable to LT. Fur- ther, this vacuolation process is not related to prostaglandin synthesis since PMN's preincubated with indomethacin, a cycloxy- genase inhibitor, show similar vacuolation. The phenomenon of cell vacuolation is not totally unders tood. Some authors have cor- related cytoplasm vacuole formation with PMN's activation (21), while others have suggested that it is an indicator of cellular damage (22). Another relevant effect indu- ced by the lymphocyte supernatant was PMN's clumping. Our results show that a maximum degree of PMN' s clumping was obtained when fresh P H A was added to the unstimulated lymphocyte supernatants (23) compared to supernatants from lymphocyte cultures stimulated with P H A which induced fewer and smaller clumps. This phenomenon may have at least two possible explanations : a) P H A alone induces cellular clumping since mitogens are known to act directly on cell membrane to produce cell agglutination (24). The decrease in the number of clumps compared to control supernatants with fresh PHA, seen in this report, may result f rom a
lower concentration of P H A in the superna- tant f rom stimulated lymphocytes since some of it would have been bound during stimulation, b) Conversely, factor(s) may be released by the stimulated lymphocytes which interact with the neutrophil cell membranes blocking the action of PHA. Further, as seen in the vacuolation studies, cycloxygenase products appear to play a role since preincubation of the PMN's with in- domethacin has no effect on the clump for- mation. Finally, in the present study, PMN's were found to increase the release of TXB2 compared to normal control levels. Further, this increase was blocked by the preincubation of P M N ' s with indomethacin. To our knowledge, this is the first time PMN's have been shown to produce throm- boxane in response to lymphocyte-derived factors.
Nevertheless it has been shown recently that lymphocytes can induce the production of PGE l in tumor cells (25). The production or increase in production of PGs may be in- duced by lymphotoxins since they have re- cently been reported to activate phospholy- pase A (10) which may result in the release of the th romboxane precursor, namely ara- chidonic acid. The results of these studies show that supernatants f rom P H A stimulat- ed lymphocytes induce marked vacuolation, and increase in TXB2 production with a pos- sible reduction in PHA-induced cellular clumping in peripheral blood PMN's . These effects may in part be induced by LTs and play and important role in modulating PMN's function during an inf lammatory response.
REFERENCES
1. Bellanti, J.A. Immunology I1. Ed. Asian, W.B. Saunders Co. 1978, 236.
2. Granger, G.A., Shimizu, I., Andreson, J. et al. Detection of lymphotoxins in vivo. Clin Immunol Immunopathol 1978, 10, 104-115.
3. Hiserodt, J.C., Prieru, A.M., Granger, G.A. In vitro lymphocyte cytotoxicity. 1. Evidence of mul- tiple cytotoxic molecules secreted by mitogen acti-
vated human lymphoid cells in vitro. Cell lmmu- nol, 1976, 24, 2777.
4. Granger, G.A., Kolb, W.P. Lymphocyte in vitro cytotoxicity. Mechanisms of immune and non- immune, small, lymphocyte-mediated target L cell destruction. J, lmmunol 1968, 101, 111.
5. Granger, G.A., Yamamoto, R.S., Fair, D.S., et al. The human LT system. 1: Physical heteroge-

Human lymphokine and polymorphonuclear leucocytes 419
neity of LT molecules released by mitogen-activat- ed human lymphoeytes in vitro. Cell Immunol 1978, 38, 403.
6. Harris, P.C., Yamamoto, R.S., Crane, J., et al. The human LT serum. X. The initial form releas- ed by T-enriched lymphocytes is 150,000 m.w. as- sociated with small non-lyric components, and can dissociate into the smaller, B, and m.w. classes. J Immunol 1981, 126, 2165.
7. Walker, S.M., Lee S.C., Lucas Z.J. Cytotoxic ac- tivity of lymphocytes. VI. Heterogeneity of cyto- toxic in supernatants of mitogen-activated lym- phocytes. J Immunol 1976, 116, 807.
8. Frienal, D.S., Rosenau, W. Target-cell membrane alterations induced by lymphotoxin: Ultrastruc- tural observation. Am J Pathol 1977, 86, 149.
9. Rosenau, W., Golgberg, M.L., Burke, G.C. Early biochemical alterations induced by lymphotoxin in target cells. J Immunol 1973, 111, 1128-1139.
10. Kobayashi, Y., Sanada, J.I., Osawa, T. Acti- vation of membrane phospholipase A by guinea pig lymphotoxin (GTL). J Immunol 1979, 122, 791.
11. Kobayashi, Y., Sanada, J., Osawa, T. Isolation and characterization of an inhibitory glycopeptide against guinea pig lymphotoxin from the surface of L cell. Immunochemistry 1978, 15, 61.
12. Williams, T.W., Granger, G.A. Lymphocyte in vitro cytotoxicity : Mechanism of human lympho- toxin-induced target cell destruction. Cell Immu- nol 1973, 6, 171-185.
13. Cohen, M.S., Metcalf, J.A., Root, R.K. Regu- lation of oxygen metabolism in human granulo- cytes: Relationship between stimulus binding and oxidative response using plant lectins as probes. Blood 1980, 55, 1003-1010.
14. Goodwin, J.S., Bankhurst, A.D., Murphy, S.A., et al. Partial reversal of the cellular immune defect in common immunodeficiency with indomethacin. J Clin Lab Immunol 1978, 1, 197-199.
15. Boyum, A. Isolation of lymphocytes, granulocytes and macrophages. Scand J Immunol 1976, 5, 9.
16. Boner, A., Zeligs, B.J., Bellanti, J.A. Chemiotat- tic responses of various differentiational stages of neutrophyls from human cordon and adult blood. Infections and Immunity 1982, 35, 921-928.
17. Jakschik, B.A., Falkenhein, S., Parker C.W. Precursor role of arachidonic acid in release of slow reacting substance from rat basophilic leukemia cells. Proc Natl Acad Sci USA 1977, 74, 4577-4581.
18. Morley J., Bray, M.A., Jones, R.W., et al. Pros- taglandin and thromboxane production by human and guinea pig macrophages and leukocytes. Pros- taglandin 1979, 17, 730-736.
19. Hampton, R.Y., Holz, R.W., Goldstein, l.J. Phospholipid glycolipid, and ion depencies of concanavalin A and Ricinus communis agglutinin induces agglutination of lipid vescicles. J Biol Chem 1980, 255, 6766-6771.
20. Sundler, R. Agglutination of glycolipid-phospho- lipid vescicles by concanavalin A. FEBS Letters, 1982, 141, 11-13.
21. Cross, A.S., Lowell, G.H. Stimulation of poly- morphonuclear leukocyte bactericidal activity by supenatants of activated human mononuclear cells. Infect Immun 1978, 22, 502-507.
22. Engel, A.G. Evolution and content of vacuoles in primary hypokalemic periodic paralysis. Mayo Clin Proc 1970, 45, 774-814.
23. Kirschbaum, B.B. Aggregation of renal brush border membranes by concanavalin A and heavy metals. Toxicology and Applied Pharmacol 1982, 64, 10-19.
24. Averdun, K.R., Gunther, T.H. Effect of concana- valin A on Ca 2§ binding, Ca 2§ uptake, and Ca 2+ ATPase of lymphocyte plasma membrane. Biochem Biophys Res Commun 1980, 97, 1146- 1153.
25. Owen, K., Gomolka, D., Droller, N.J. Lympho- cyte-induced production of prostaglandin E~ by tumor cell in vitro: Requirements for direct con- tact and lymphocyte viability. Cell Immunol 1980, 55, 428-435.



















