Embed Size (px)
Citation preview

Volume 12, number 3 MOLECULAR • CELLULAR BIOCHEMISTRY September 30, 1976
STUDIES ON THE MECHANISM OF METABOLIC STIMULATION IN POLYMORPHONUCLEAR LEUKOCYTES DURING PHAGOCYTOSIS. ACTIVATORS AND INHIBITORS OF THE GRANULE BOUND NADPH OXIDASE*
Pierluigi PATRIARCAt , Pietro DRl , 'Katsuko K A K I N U M A and Filippo ROSSI
Istituto di Patologia Generale, Via A. Valerio, 34, University of Trieste, Trieste, Italy
(Received Febuary 2, 1976)
Summary Introduction
The effects of several known inhibitors and activators of peroxidase-catalyzed reactions have been studied on the N A D P H oxidase activity of granules isolated from polymor- phonuclear leukocytes at rest or during phagocytosis. Redogenic substances, such as ascorbate or hydroquinone, and superoxide dis- mutase, which are known to inhibit peroxidase- catalyzed reactions, also inhibited the N A D P H oxidase activity of granules. Oxidogenic sub- stances, such as guaiacol or resorcinol, anff manganese, which are known to stimulate peroxidase-catalyzed reactions, also activated the N A D P H oxidase activity of granules. Cyanide, an inhibitor of peroxidase-catalyzed reactions, inhibited the N A D P H oxidase activity of granules isolated from resting leukocytes but only slightly affected that of granules isolated from phagocytosing cells, as previously re- ported. A list of the properties of the N A D P H oxidase activity of granules and of peroxidase oxidase activity is given. The arguments in favor of and those against a possible identity of the two activities are discussed.
* This paper is publication 9 of a series entitled: "En- zymatic basis of the metabolic stimulation in phagocytosing leukocytes". The other publications of the series are those quoted in the Bibliography section as numbers 6, 8, 9, 11, 14, 16, 17, 36. t To whom correspondence should be addressed
An increased oxidation of reduced nicotinamide coenzymes is unanimously 1-11 regarded as the biochemical event that triggers the so called "metabolic burst" of phagocytosing polymorphonuclear leukocytes (PMN), with in- creased oxygen uptake, increased oxidation of glucose through the hexosemonophosphate shunt (HMS) and increased production of H202 as compared with PMN at rest.
It has still to be settled, (1) whether reduced nicotinamide adenine dinucleotide (NADH) 1-4"12 or reduced nicotinamide adenine dinucleotide phosphate (NADPH) 5 11 is preferentially ox- idized in phagocytosing PMN, and (2) the nature of the enzyme(s) that perform(s) the oxidation.
It was suggested by ROBERTS and QUASTEL in 196413 that myeloperoxidase (MPO) may be responsible for the oxidation of the nicotinamide coenzymes in leukocytes. O u r previous studies on the characterization of the N A D P H oxidizing activity of PMN granules have shown that this activity is localized in azurophilic granules 14, which also contain MPO 15. These studies have led to the proposal of a mechanism for the oxidation of N A D P H by granules 16. However attempts to separate the N A D P H oxidizing activity from the oxidase activity of MPO were unsuccessful due to inhibition of the former activity during solubili- zation procedures 17.
Dr. W. Junk b.v. Publishers - The Hague, The Netherlands 137

In this paper a different approach to discrimi- nate between the two activities has been fol- lowed: the biochemical requirements of the N A D P H oxidizing activity have been investi- gated in order to compare them with the established requirements of peroxidase oxidase reactions.
Materials and Methods
Preparation of leukocytes Leukocytes were obtained from guinea pig acute peritoneal exudates elicited by a 50 ml injection of a 1.2% sterile solution of sodium caseinate in 0.9% NaC1. The exudate was collected 14 hours later. After centrifugation, the sedimented cells underwent brief hypotonic t reatment (30-60 seconds) to remove con- taminating erythrocytes and finally were sus- pended in calcium-free Krebs Ringer Phosphate buffer (KRP) pH 7.4. The preparations contain- ing less than 90% polymorphonuclear leuko- cytes were discarded.
Phagocytosis Cells were incubated with heat-killed Bacillus subtilis (100 bacteria/leukocyte) or with latex spherules (200 particles/leukocyte) for 5 rain at 37 °C. At the end of the incubation the suspension was diluted in ice-cold Krebs Ringer Phosphate buffer and leukocytes were collected by centrifugation at 400 x g for 7 minutes.
Preparation of granules Packed cells were suspended in ice-cold 0.34 M sucrose (30-40 x 107/ml) and homogenized with a Potter- type homogenizer until about 90% of the cells were disrupted, as judged by light micro- scopic examination. The homogenate was diluted about ten times with 0.34 u sucrose and centrifuged at 250 x g for 5 min to sediment unbroken cells, nuclei and cell debris. The resulting supernatant was centrifuged at 20,000 x g for 15 min and the pellet, containing the granules, was suspended in 0.34 M sucrose.
Enzymatic assay N A D P H oxidase activity was measured as ox- ygen uptake with a Clark oxygen electrode 6 or by following the oxidation of N A D P H
138
spectrophotometrically at 340 nm 16 as previ- ously described.
Materials The sources of materials used were as follows: N A D P H (enzymatically reduced) from Boehringer, Mannheim, West Germany; L- ascorbic acid and hydroquinone from B D H Chemicals Ltd., Poole, England; resoricinol from Mallinckrodt Chemical Works, St. Louis, USA; Guaiacol from Sigma Chemical Co., St. Louis, USA; superoxide dismutase (bovine erythrocyte) from Truet t Laboratories, Dallas, USA.
Experimental
The scheme proposed by YAMAZAKI and cowor- kers 18-2~ for the peroxidase oxidase reaction can be summarized with the following set of reactions:
peroxidase 2AH2 + H202 ~ 2AH" + 2H20
AH" + 0 2 ) A + O2"+ H +
02-" + H + + AH2 AH" + H202
(1)
(2)
(3)
The overall reaction is composed of an enzyma- tic formation of free radicals (reaction 1) and a chain reaction (reactions 2 and 3 ). The scheme is completed by two other reactions that ac- count for chain termination.
202" + 2H + sup . . . . ide di .... tase ~ H202 + 02 (4)
AH" +AH" > A H 2 + A (5)
The properties of the peroxidase oxidase reaction are not always the same for each hydrogen donor and are greatly influenced by environmental conditions (for example pH of the reaction) and by a number of chemical and enzymatic agents.
Table 1 and Table 2 show the effect of selected agents, that are known to affect perox- idase oxidase activity, on the N A D P H oxidase activity of isolated PMN granules measured at acid pH and at alkaline pH, respectively. A detailed comment on the effect of each agent is given in the following paragraphs.

Table 1 - ACTIVATORS AND INHIBITORS OF THE OXIDATION OF NADPH BY PMN GRANULES AT PH 5.5
Additions n a t o m s o x y g e n / m i n / g r a n u l e s f r o m 108 c e l l s
G r a n u l e s f r o m G r a n u l e s f r o m PMN Pb~ at rest under phagocytosis
n o n e 26 129
+ 1 mM KCN 0 115
+ 0 . 5 mM b~C12 1 , 6 0 0 6 , 2 0 0
+ 0 . 5 mM ~ C 1 2 + 1 mM KCN 30 5 , 6 4 2
+ 2 m M R e s o r c i n o l 2 , 8 0 0 2 ,545
+ 0.5 mM MnCI 2 + 2 mM Resorcinol 52,000 54,300
+ 0.5 mM ~CI 2 + O.I mM Hydroquinone 800 930
+ 0.5 mM HnCI 2 + 1 .24 Hydroquinone 510 700
+ 0.5 mM ~CI 2 + O.05 mM Ascorbate 0 O
Assay medium: 75 ~,I phosphate buffer pll 5.5, 170~M sucrose. Final volume 2 ml.
The reaction was started by adding 1 mbl NADPH. Temperature 37 ° C.
Effect of Mn ++ Manganese is known to stimulate the oxidation by peroxidase of several hydrogen donors, in- cluding reduced nicotinamide coenzymes 22'23. The magnitude of the effect of Mn ++ greatly depends on the experimental conditions. Thus no appreciable effect of Mn ++ alone was found, at neutral pH, on the oxidation of N A D P H or N A D H by horse radish peroxidase (HRP) 22"24 or myeloperoxidase 24 or uterine peroxidase 25,
whereas at acid pH Mn ++ alone markedly enhanced the oxidation of both nucleotides by horse radish peroxidase 13. However at either neutral pH or acid pH Mn +÷ strongly stimulated the oxidation of nicotinamide coenzymes in the presence of phenols 22'25"26 or phenol-like com- pounds 24 by different types of peroxidase. It has been proposed by YAMAZAKI and PIErrE that the mechanism of activating effect of Mn ++, at acid pH, is its oxidation to Mn +÷+ by O~" produced
Table 2 - ACTIVATORS AND INHIBITORS OF THE OXIDATION OF NADPH BY Pb~ GRANULES AT pH 7.2
Additions n a t o m s o x y g e n / m i n / g r a n u i e s f r o m 108 c e l l s
G r a n u l e s f r o m G r a n u l e s f r o m Pb~ PMN a t r e s t u n d e r p h a g o e y t o s i s
n o n e 14 52
+ 1 nibl KCN 1 49
+ 0 . 5 mbl b~C12 14 52
+ 2 mN IvlnC12 15 51
+ 2 mM R e s o r c i n o l 2 , 8 0 0 2 , 7 2 0
+ 2 mHResorcinol + 0.5 mMMnCl 2 25,200 3 4 , 3 0 0
+ 2 n~l A s c o r b a t e - 53
+ 2 mM H y d r o q u i n o n e - 20
A s s a y medium: 50 mM p h o s p h a t e b u f f e r pH 7 . 2 and 170 mM s u c r o s e . T h e r e a c t i o n w a s s t a r t e d by a d d i n g 2 mM NADPH. F i n a l v o l u m e 2 m l . T e m p e r a t u r e 37 ° C.
139
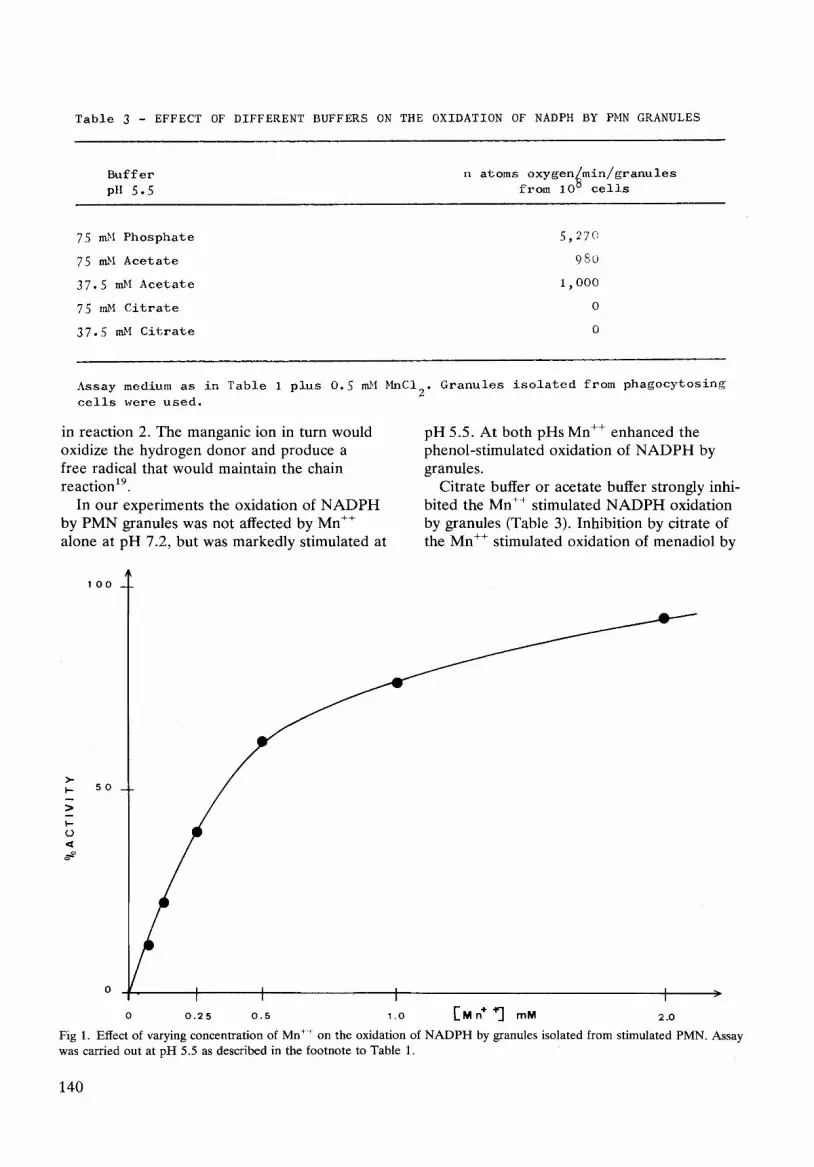
Table 3 - EFFECT OF DIFFERENT BUFFERS ON THE OXIDATION OF NADPH BY PMN GRANULES
B u f f e r n atoms oxygen~min/granules pH 5 . 5 f r o m 10 u c e l l s
75 mbl Phosphate 5 , 2 7 0
75 mM Acetate 9 8 0
3 7 . 5 nLM Acetate 1 , 0 0 0
75 mM Citrate 0
37.5 mM Citrate 0
Assay medium as in Table 1 plus 0.5 n~l MnCI 2. Granules isolated from phagocytosing
cells w e r e used.
in reaction 2. The manganic ion in turn would oxidize the hydrogen donor and produce a free radical that would maintain the chain reaction 19.
In our experiments the oxidation of NADPH by PMN granules was not affected by Mn ÷÷ alone at pH 7.2, but was markedly stimulated at
pH 5.5. At both pHs Mn ++ enhanced the phenol-stimulated oxidation of NADPH by granules.
Citrate buffer or acetate buffer strongly inhi- bited the Mn ÷÷ stimulated NADPH oxidation by granules (Table 3). Inhibition by citrate of the Mn +÷ stimulated oxidation of menadiol by
>- p-
t - O
1 0 0
5 0
t I I I 0 0 . 2 5 0 .5 1.0 EM n + ~ mM 2.0
Fig 1. Effect of varying concentrat ion of Mn ÷+ on the oxidation of N A D P H by granules isolated from st imulated PMN. Assay was carried out at pH 5.5 as described in the footnote to Table 1.
140

horse radish peroxidase has been reported by KLAPPER and H A C K E T 1 "27.
Manganese could not be replaced, in our experiments, by other divalent cations such as Mg ++, Ca ++, Zn ++, Fe ++, A1 ++, as has been reported for peroxidase oxidase activity 22'25 Manganese exerted its stimulatory effect at low concentration as shown in Figure 1.
Thus the oxidation of the reduced nicotinamide coenzymes by PMN granules has identical prop- erties to the peroxidase oxidase reaction as far as the response to Mn ÷+ is concerned.
Effect of oxidogenic substrates YAMAZAKI has proposed a tentative classification of the substrates of peroxidase oxidase reactions into oxidogenic and redogenic substrances, ac- cording to the oxido-reductive characteristics of the intermediate formed during peroxidase reac-
tion 18. Redogenic substrates form free radical intermediates with strong reducing activity whereas oxidogenic substrates form very unstable free radicals which are strong one-electron oxidizing agents. Among the latter series of substrates are resorcinol and other phenols that are not readily oxidized to the corresponding quinone. Oxidogenic substances all activate the peroxidase oxidase reaction at both neutral and acid pH 19'21"22'25'28"29. The phenol is effective in
catalytic amounts and is not consumed during the reaction. It has been proposed that the phenol undergoes a cycle of oxidation and reduction with formation of a free radical, which, in turn, oxidizes the hydrogen donor (for example NAD(P)H) 18'2°'22.
Oxidation of N A D P H by PMN granules is also stimulated by resorcinol at both neutral pH and acid pH, and the stimulation is enhanced by
i
,d-
n
F: 3 10 r-
E r- ~J
X 0 u) E
1 0 - - c
b.-
I.--
o I i i 5 ,x,5 0 5 50 2 5 0
J R e s o r c i n o l j H M
Fig. 2. Effect of varying concentration of resorcinol on the oxidation of NADPH by granules isolated from either resting ([]--[]) or phagocytosing (O--O) PMN. Assay at pH 5,5 in the presence of 0.5 mM Mn ++. For other experimental details see Table 1.
141

Effect of redogenic substrates Redogenic substrates such as ascorbic acid, hydroquinone or benzoquinone inhibit peroxidase oxidase reaction 19'21. Ascorbate and hydroquinone strongly inhibit the oxidation of NADPH by PMN granules at acid pH. At
LU
0 z ,,~ co
0
co
neutral pH hydroquinone is still inhibitory but ascorbic acid has no effect. When ascorbic acid is tested at much lower concentrations than those shown in Table 1 it causes a transient inhibition of NADPH oxidation which then resumes after a lag time. Addition of H202 reduces or abolishes the lag time (Fig. 3). This suggests that H202 may be the initiator of the reaction and that ascorbate may act by consum- ing it. The lag time is not directly proportional to the concentration of ascorbic acid. Such an irregular relationship is also found between the lag time and the amount of granules added (Fig. 4).
Similar results have been obtained by Yamazaki and Yamazaki while studying the effect of ascorbic acid on the oxidation of indole-3-acetic acid (IAA) by purified horse radish peroxidase 3°. Those authors have interpreted the non linear dependence of the lag time upon the concentration of ascorbate as suggestive of an involvement of a kind of chain reaction in
1 . 8 -
1 . 6 -
1 . 4 -
1 . 2 -
I ~ lag t i m e - - ~
I I i \ \1
NA
Mn ÷+. Figure 2 shows the effect of varying resorcinol concentration on the Mn ÷+- stimulated oxidation of NADPH by granules isolated from cells at rest or under phagocytosis. It can be easily calculated from the figure that the oxygen consumed during oxidation of NADPH by granules, in the presence of resor- cinol, greatly exceeds the amount of oxygen that would be required for complete oxidation of the phenol present in the reation mixture. This indicates that the phenol acts catalytically and suggests that the mechanism proposed for the stimulation of the peroxidase oxidase reactions by phenols may apply to the NADPH oxidase activity of PMN granules.
T o ! I I I I I "
0 2 4 6 8 10
T I M E ( r a i n ) Fig. 3. Effect of hydrogen peroxide on the ascorbate-induced lag time of the oxidation of NADPH by granules isolated from phagocytosing PMN. Spectrophotometric recordings of the disappearance of NADPH at 340 mp, A: control. B: A + 2 IXM ascorbate + 4 IXM H20> C: A+ 2 IXM ascorbate. The assay medium contained 75 mM phosphate buffer pH 5.5, 170 mR sucrose, 0.5 mM MnC12 and granules corresponding to 2 x 107 cells. The reaction was started by adding NADPH (0.3 mM final). Volume 3 ml. Temperature 37 °C.
142

16
12
e- .B g
8 LU =E
<~ 4 -
A
0 1 2 3 4
lAscorbatel /aM
3 2 .
• ~- 2 4 E
"LU 5;
I-- 16
.-I
0 I I 1 0 5 0 100 2 0 0
Granules iJI
Fig. 4. Effect of varying concentration of ascorbate (A) and amount of granules (B) on the ascorbate-induced lag time of NADPH oxidation by granules isolated from phagocytosing PMN. Assay as in Figure 3. The suspension of granules used contained granules corresponding to 2 x 10 ~ cells per mI. In A 100 txl of granules were used. In B 4 tXM ascorbate was used.
the initiation of I A A oxidation. A chain reac- t ion would also account for several propert ies of the oxidat ion of N A D P H by granules 16.
Effect of superoxide dismutase Superoxide dismutase (SOD) has been repor ted t o inhibit the aerobic oxidat ion of N A D H by M P O and the format ion of c o m p o u n d I I I in the presence of 2,6-dicloropheno131. This result is cons ide red evidence that bo th O~" and com- p o u n d I I I are impor tan t intermediates in the peroxidase oxidase reaction. Figure 5 shows that S O D markedly inhibits the oxidat ion of N A D P H by granules when added ei ther before the start of the react ion or half way during the react ion. A more detailed account of the effect of S O D on the oxidat ion of N A D P H by P M N granules is given in another paper 16.
Effect of cyanide Cyanide strongly inhibits the oxidat ion of N A D P H by granules isolated f rom cells at rest bu t affects only slightly the oxidat ion of N A D P H by granules isolated f rom phagocytos- ing cells, in keeping with previous results 6.
1000 f
900~-
=; 6oo
600
0 - ~ t I I I I 0 60 120 180 240 300 TIME (seconds)
Fig. 5. Effect of superoxide dismutase (SOD) on the oxidation of NADPH by granules isolated from phagocytos- ing PMN. Recording of the oxygen uptake at pH 5.5 without Mn ++. Granules corresponding to 108 cells were used. a: control (no SOD added), b: SOD (30 p,g) added 90 seconds after the addition of NADPH, c: SOD (30/xg) added before the addition of NADPH. For other experi- mental details see Table 1.
143

Discussion
This paper shows that the reaction of NADPH oxidation by polymorphonuclear leukocyte granules shares several biochemical properties with the peroxidase oxidase reaction as judged from the effect on the NADPH oxidizing activ- ity of some reagents which are known to affect the peroxidase oxidase activity. Indeed all the activators and inhibitors of the peroxidase oxidase reaction, that have been tested, also activated or inhibited, respectively, the NADPH oxidizing activity of PMN granules, with the sole exception of cyanide. Cyanide in fact, which is known as a general inhibitor of peroxidase-catalyzed reactions, inhibited the oxidation of NADPH by granules isolated from resting PMN, but did not affect appreciably that by granules isolated from phagocytosing PMN.
The problem of discriminating between the peroxidase oxidase activity and a distinct, but still hypothetical, NADPH oxidase activity in PMN granules, has been a puzzle since the central role of the oxidation of nicotinamide coenzymes in the metabolic activation of PMN
has been appreciated. The difficulty arises from, (1) the failure to separate the t{vo activities, and (2) the lability of the NADPH oxidizing activity, which frustrates any attempts at its purification.
In order to make the discrimination possible many authors assay NADPH or NADH oxida- tion by granules in the presence of cyanide with the aim to inhibit myeloperoxidase. This proce- dure however does not seem to be entirely rationale for several reasons. (1) Such a dis- crimination is not possible in resting granules due to inhibition of both activities by cyanide. (2) The differential effect of cyanide on the two activities, in phagocytosing granules, is not evidence at all for two distinct enzymes or two distinct enzyme sites. It is true indeed that cyanide is a general inhibitor of peroxidase- catalyzed reactions, including myeloperoxidase from resting leucocytes, although some excep- tions have been reported in the literature 32-34, but no one knows what the effect of cyanide is on myeloperoxidase isolated from phagocytosing leucocytes. (3) During peroxidase reaction a series of intermediates of peroxidase at different oxidation states is formed which differ in their
144
TABLE 4 - A COMPARISON OF THE PROPERTIES OF PEROXIDATIC AND OXIDATIC ACTIVITI~$ OF PEROXIDASE FROM VARIOUS SOURCES WITH THE PROPERTIES OF NADpH-OXIDASE ACTIVITy OF POLYb~RPHONUCLEAR LEUCO- CYTE GRANULES.
EXPERIMENTAL APPROACH ~EROXIDASBS PROM VARIOUS SOURCES NADPH-OXIDASE ACTIVITy OXIDATIC OF PIll GRANULES PEROXIDATIC
ACTIVITY ~ ACTIVITY
INHIBITION (40) INHIBITION WITH FEW ~XCEPTIONS (19,33,34)
HYDRO~U~ONB INHIBITION OF NEU- INHIBITION ( 1 9 , 2 2 , ASCORBATE TROPHIL MPO OR PU- 27)
RIFIEDHRP ~unpu- bl l shed}
5-~IINO-I,3,4.TKIAZOLE INHIBITION (14) STIMULATION (42)
MANGANESE NO EFFECT ON NEU- STIMULATION (19,22, TROPHIL MP0 OR PU_ zS) RIFIED ~P ( u n p u - b l i s h e d )
RESORCXNOL INHIBITION ON NEU STIMULATION (19-22, 2,4-DICHLOROPBENOL TROPHIL MPO. NO E [ 25,31)
FECT ON PURIFIED HRp (unpttblished)
SUPBROXIDE DISMUTASE NO EFFECT ON NEU- INHIBITION (31) TROPHIL ~PO OR PU RIFIED I ~ P (unpuU bl i shed)
-I~HIBITEDINGRANULBS FROM RESTING PHN ( 6 , t h l s papeF)
-U~AFFECTEV ~ G ~ U L B S FRO~ PHAGOC~TOSING FM~ (@, c~ubm paper)
INHIBITION ( t h i s papel')
STIMULATION (14)
STIMULATION (this paper)
STI~ULATION (this pa~e~)
INHIBITION (16)
TREATMENT OF GUINEA PIG SOLUBILIZATION AND .............. P~g GRANULES WITH CETyL- NO EFFECT ON THE A~ TRIMETHILAMMONrOM BROMI- TIVITY IN BOTH RE- DE, GUANIDINIUM CHLORIDE, STING AND PHAGOCY- KCI TOS~G GRANULES (17)
-RESTING GRA~UL£SIPARTIAL I~HIBITION AND SOLUBILI- ZATION [(~publlmh®d)
-~AGOCt~OSING C~"~NULESt HARKED INHIBITION {17) ~
SUBCELLULAR LOCALIZA- AZLq%OPHILIC GRANU . . . . . . . . . . . . . . . . . . AZ~IKOI~ILIC GRANULES ~q TION LES IN RABBIT P~ RABBIT P~ (14)
(14)
MPO-DEFICIBNT MAN ABSENT (36,38,39) ................. PRESENT (36,37)
* H e a r t e d by the ~ a i a c o l teB% ~* Due t o s t r o n g i n h i b i t i o n , i t i s not poss ib le to e s t a b l i s h the d~geee of s o ~ a b i i i z a t i o n ~ o ~ MyeiopeFoxidase N~trophll MPO: Myeloperoxidase activity of the total g~a~le populatlon of ~inea pig n~t~phlllc
l ~ c 6 c y t e s }~p: Horseradish peroxi~ase

affinity for cyanide , f e r rope rox idase , for example , be ing much less sensi t ive to cyanide than f e r r ipe rox idase 35.
A s u m m a r y of the p rope r t i e s of the N A D P H ox idase act ivi ty and of the m y e l o p e r o x i d a s e act ivi ty (oxidase and pe rox idase activit ies) is g iven in Tab le 4. A m o n g n u m e r o u s s imilar i t ies , some differences , bes ides the effect of cyanide tha t has jus t been discussed, can be o b s e r v e d b e t w e e n the two activi t ies , such as the r e sponse to some solubi l iz ing agents and the p re sence of N A D P H oxidase bu t no t of the classical p e r o x - idase act ivi ty in a M P O - d e f i c i e n t pa t i en t 36'37. H e r e again such di f ferences are not defini te p r o o f tha t two dis t inct enzymes exist.
The p resence of the N A D P H oxidase act iv- i ty 36'37 and of a r egu la r r e sp i r a to ry burs t 38'39 in
the M P O deficient pa t ien t s is, at first glance, the mos t ser ious o b j e c t i o n to the hypo thes i s of the iden t i ty of g ranule N A D P H oxidase and the ox idase act ivi ty of M P O . In fact M P O canno t be d e t e c t e d in neu t roph i l s and monocy te s of these subjec ts e i the r enzymat i ca l ly (classical pe rox ida se activity) 36'38, spec t roscop ica l ly or im- muno log ica l ly 41. This resul t does not necessar i ly m e a n tha t the enzyme is lacking in the cells. A n a l t e rna t ive exp l ana t i on might be tha t the en- zyme is a l t e red in its enzymat ic , h e m e - b i n d i n g and ant igenic sites. W h e t h e r such an a l t e red M P O may re ta in its ox idase act ivi ty has to be es tabl i shed .
Acknowledgements
Dr. K. KAKINUMA is a visi t ing inves t iga tor f rom the D e p a r t m e n t of B iochemis t ry , Kyushu Un i - vers i ty , F u k u o k a , Japan . This w o r k has been s u p o r t e d by g ran t No. 74 .00272 .04 f rom Na- t iona l R e s e a r c h Counc i l of I t a ly (CNR).
References
1. Baehner, R. L., Gilman, N. and Karnovsky, M. L., 1970. J. Clin. Invest. 49, 692-700.
2. Cagan, R. H. and Karnovshy, M. L., 1964, Nature (London) 204. 255-257.
3. Evans D. H., and Karnovsky, M. L., 1961. J. Biol. Chem. 236, Pc 30-32.
4. Evans, D. H. and Karnovsky, M. L., 1962. Biochemis- try 1,159-166.
5. Iyer, G. Y. N. and Quastel, J. H., 1963. Canad. J. Biochem. Physiol. 41,427-434.
6. Patriarca, P., Cramer, R., Moncalvo, S., Rossi, F. and Romeo, D., 1971. Arch. Biochem. Biophys. 145, 255-262.
7. Paul, B. B., Strauss, R. R., Jacobs, A. A. and Sbarra, A. J., 1972. Exp. Cell. Res. 73, 456-462.
8. Rossi, F. and Zatti, M., 1966. Biochim. Biophys. A'cta 121, 110-119.
9. Rossi, F. and Zatti, M., 1968. Biochim. Biophys. Acta 153, 296-299.
10. Stjernholm, R. L. and Manak, R. C., 1970. RES J. Reticuloendothel. Soc. 8, 550-560.
11. Zatti, M. and Rossi, F., 1966. Experientia 22, 758-759. 12. De Chatelet, R. L., McPhail, L. C., Mullikin, D. and
McCall, C., 1974. Infect. Immun. 10, 528-534. 13. Roberts, J. and Quastel, J. H., 1964. Nature 202,
85-87. 14. Patriarca, P., Cramer, R., Dri P., Fant, L., Basford, R.
E. and Rossi, F., 1973. Biochim. Biophys. Res. Com- mun. 53, 830-837.
15. Baggiolini, M., Hirsch, J. G. and de Duve C., 1969. J. Cell Biol. 40, 529-541.
16. Patriarca, P., Dri, P., Kakinuma, K., Tedesco, F. and Rossi, F., 1975. Biochim. Biophys. Acta 385, 380-386.
17. Patriarca, P., Basford, R. E., Cramer, R., Dri, P. and Rossi, F., 1974. Biochim. Biophys. Acta 362, 221-232.
18. Yamazaki, I., 1957. Proc. Intern. Symposium on En- zyme Chem. (Tokio-Kyoto) pp. 224-229.
19. Yamazaki. I. and Piette, L. H., 1963. Biochim. Biophys. Acta 77, 47-64.
20. Yamazaki, I. and Yokota, K., 1973. Molecular Cell Biochem. 2, 39-52.
21. Yokota, K. and Yamazaki, I., 1965. Biochim. Biophys. Acta 105, 301-312.
22. Akazawa, T. and Conn, E. E., 1958. J. Biol. Chem. 232, 403-415.
23. Chance, B., 1952. J. Biol. Chem. 197, 577-589. 24. Klebanoff, S. J., 1959. J. Biol. Chem. 234, 2480-2485. 25. Beard, J. and Hollander, V. P., 1962. Arch. Biochem.
Biophys. 96, 592-600. 26. Hollander, V. P. and Stephens, M. L., 1959. J. Biol.
Chem. 234, 1901-1906. 27. Klapper, M. H. and Hackett, D. D., 1963. J. Biol.
Chem. 238, 3736-3742. 28. Kenten, R. H. and Mann, P. J. G., 1949. Biochem. J.
45, 255-263. 29. Odajima, T. and Yamazaki, I., 1970. Biochim. Biophys.
Acta 206, 71-77. 30. Yamazaki, H. and Yamazaki, I., 1973. Arch. Biochem.
Biophys. 154, 147-159. 31. Odajima, T., 1971. Biochim. Biophys. Acta 235, 52-60. 32. King, C. M., Bedner, T. W. and Linsmaier-Bedner, E.
M., 1973. Chem. Biol. Interactions 7, 185-188. 33. Morita, Y. and Kameda, K., 1961. Mere. Res. Inst.
Food Sci. Kyoto Univ. 23, 1-14. 34. Nicholls, P., 1961. Fed. Proceed. 20, 50. 35. Phelps, C., Antonini, E. and Brunori, M., 1971.
Biochem. J. 122, 79-87. 36. Patriarca, P., Cramer, R., Tedesco, F. and Kakinuma,
K., 1975. Biochim. Biophys. Acta 385, 387-393. 37. Hohn, D. C. and Lehrer, R. I., 1975. J. Clin. invest. 55,
707-713.
145

38. Lehrer, R. I. and Cline, M., 1969. J. Clin. Invest. 48, 1478-1488.
39. Klebanott, S. J. and Pincus, S. H., 1971. J. Clin. Invest. 50, 2226-2229.
40. Agner, K., 1941. Acta Physiol. Scand. 2, suppl. 8, 4-62. 41. Salmon, S. E., Cline, M. J., Schultz, J. and Lehrer,
R. I., 1970. N. Eng. J. Med. 282, 250-253 42. Takanaka, K. and O'Brien, P. J., Biochem. Biophys.
Res. Commun. 62, 966-971.
146





























