Embed Size (px)
Citation preview

Enriched Odor Exposure Increases the Number of NewbornNeurons in the Adult Olfactory Bulb and Improves Odor Memory
Christelle Rochefort,1* Gilles Gheusi,1,2* Jean-Didier Vincent,1 and Pierre-Marie Lledo1
1Perception and Memory Laboratory, Centre National de la Recherche Scientifique Unite de Recherche Associee 2182,Institut Pasteur, 75 724 Paris Cedex 15, France, and 2Laboratory of Ethology, Centre National de la RechercheScientifique Unite de Recherche Associee 7025, Universite Paris XIII, 93430 Villetaneuse, France
In the mammalian forebrain, most neurons originate from pro-liferating cells in the ventricular zone lining the lateral ventricles,including a discrete area of the subventricular zone (SVZ). Inthis region, neurogenesis continues into adulthood. Most of thecells generated in the SVZ are neuronal precursors with prog-eny that migrate rostrally along a pathway known as the rostralmigratory stream before they reach the main olfactory bulb(MOB) where they differentiate into local interneurons. The ol-factory system thus provides an attractive model to investigateneuronal production and survival, processes involving interplaybetween genetic and epigenetic influences. The present studywas conducted to investigate whether exposure to an odor-enriched environment affects neurogenesis and learning inadult mice. Animals housed in either a standard or an odor-enriched environment for 40 d were injected intraperitoneallywith bromodeoxyuridine (BrdU) to detect proliferation among
progenitor cells and to follow their survival in the MOB. Thenumber of BrdU-labeled neurons was not altered 4 hr after asingle BrdU injection. In contrast, the number of survivingprogenitors 3 weeks after BrdU injection was markedly in-creased in animals housed in an enriched environment. Thiseffect was specific because enriched odor exposure did notinfluence hippocampal neurogenesis. Finally, we showed thatadult mice housed in odor-enriched cages display improvedolfactory memory without a change in spatial learning perfor-mance. By maintaining a constitutive turnover of granule cellssubjected to modulation by environmental cues, ongoing bul-bar neurogenesis could be associated with improved olfactorymemory.
Key words: behavior; cell survival; interneurons; hippocam-pus; olfaction; progenitor cells; neurogenesis
Although most cells in the nervous system are born during theembryonic and early postnatal period, newborn neurons continueto be generated within at least two areas of the adult mammalianbrain (Altman and Das, 1965). The first region concerns thesubgranular zone of the dentate gyrus in the hippocampus for-mation that gives rise to granule cells (for review, see Gage, 2000;Gross, 2000). The second is the subventricular zone (SVZ), whichis located in the wall of the lateral ventricle that harbors steadilydividing stem cells and progenitors that produce new neurons (forreview, see Peretto et al., 1999; Temple and Alvarez-Buylla,1999). These neuronal progenitors then migrate by way of therostral migratory stream (RMS) to populate the main olfactorybulb where they differentiate into local inhibitory interneuronsand establish connections with their neuronal targets (Luskin,1993; Lois and Alvarez-Buylla, 1994).
Bulbar neurogenesis in adulthood implies three distinct pro-cesses: (1) cell proliferation, (2) neuroblast migration from theSVZ to the olfactory bulb, and (3) neuronal differentiation and
survival. For example, Ephrin molecules were shown to controlspecifically cell proliferation of neuroblasts in the SVZ (Conoveret al., 2000). With respect to migration, the polysialylated form ofneural cell-adhesion molecules (PSA-NCAM) has been identifiedas a key factor in assuring efficient migration of neuroblasts in theRMS (Bonfanti and Theodosis, 1994; Rousselot et al., 1995;Doetsch and Alvarez-Buylla, 1996; Lois et al., 1996; Chazal et al.,2000). Two members from the Slit family of soluble proteins arealso good candidates for guiding neuroblasts toward the olfactorybulb (Hu, 1999; Wu et al., 1999).
It has been proposed that target structures are the majorsources for providing attractive and survival factors (Kennedyand Tessier-Lavigne, 1995; Svendsen and Sofroniew, 1996). Thematuration and survival of SVZ-generated neurons are partlyunder the control of brain-derived neurotrophic factor (BDNF)(Kirschenbaum and Goldman, 1995). Although the sources ofthese factors in vivo are not precisely known, olfactory bulb-derived factors are likely to influence the proliferation and sur-vival of SVZ neuroblasts in the adult brain. In mice, neuralactivity may be important in regulating neuroblast proliferationand survival because closure of one nostril affects the dynamics ofneuronal birth and death in the corresponding olfactory bulb(Murray and Calof, 1999). However, it was demonstrated recentlythat SVZ cells continue to divide and migrate after transection ofthe olfactory peduncle (Jankovski et al., 1998) or after olfactorybulb removal (Kirschenbaum et al., 1999). This suggests thatactivity within the olfactory bulb is not essential for proliferationor the directional migration of newly generated interneurons.Thus, the role of the bulbar activity for the survival of newborn
Received Aug. 27, 2001; revised Nov. 29, 2001; accepted Jan. 23, 2002.This work was funded in part by the Centre National de la Recherche Scienti-
fique, the Ministere de l’Education Nationale, de la Recherche et de la Technologie(ACI Biologie du Developpement et Physiologie Integrative, 2000), and the InstitutUniversitaire de France. We thank J. Pastre and P. Chevalier for their help with theimmunohistochemistry of the hippocampus and water maze experiments. We arealso very grateful to J. Morante Oria, A. Saghatelyan, A. Jankovski, and H. McLeanfor their helpful comments on this manuscript.
*C.R. and G.G. contributed equally to the work.Correspondence should be addressed to Pierre-Marie Lledo, Institut Pasteur,
Perception and Memory Laboratory, 25 Rue Dr. Roux, 75 724 Paris Cedex 15,France. E-mail: [email protected] © 2002 Society for Neuroscience 0270-6474/02/222679-11$15.00/0
The Journal of Neuroscience, April 1, 2002, 22(7):2679–2689

neurons arriving in the main olfactory bulb remains to be clearlyestablished.
It is widely accepted that the adult brain can respond toenvironmental and internal challenges inducing significant func-tional and anatomical modifications, collectively termed “neuralplasticity.” This has been well documented for the perinatalperiod called “critical period,” in which sensory-driven activitypatterns are able to induce long-term changes in specific neuronalcircuits lasting throughout adulthood (Berardi et al., 2000). Inter-estingly, recent analyses of this plasticity indicate that a develop-mental mismatch between inhibition and excitation could providea time frame during which the reorganization of cortical circuitrycan be particularly influenced by sensory experience (Huang etal., 1999; Fagiolini and Hensch, 2000). The self-renewing capacityof the olfactory bulb inhibitory neuronal network leaves open thepossibility that the critical period may never end in the olfactorysystem. In fact, odor experiences have been reported to modulateadult olfactory bulb functions (Rabin, 1988; Rosselli-Austin andWilliams, 1990; Woo and Leon, 1995). If activity-dependent re-cruitment of neurons is related to odor stimulation, the question,then, is whether a change in the number of newborn interneuronsmight be related to changes in olfactory behavioral function.Using a combination of immunohistological and behavioral ap-proaches, we show that an odor-enriched environment enhancesthe bulbar interneuron population and improves olfactory mem-ory without upregulating hippocampal neurogenesis.
MATERIALS AND METHODSHousing conditions and animalsMale C57BL/6J mice (2 months old) used for this study were obtainedfrom Janvier (Le Genest St. Isle, France). On their arrival in thelaboratory, mice were held in standard laboratory cages (43 � 15 � 28cm) in groups of 10, with wood shavings for bedding. They were ran-domly assigned to two experimental groups. The enriched group (n �31) consisted of animals housed in an odor-exposure environment for 20or 40 d. Common odoriferous items were used to enrich the olfactory
environment of animals (Table 1). Odor-enriched mice were exposeddaily for 24 hr to different aromatic fragrances that were placed in a teaball hanging from the acrylic filtering cover of standard breeding cages.Standard mice (n � 34) were reared under the same conditions exceptthat the tea ball was left empty. Three days before the behavioralexperiments, mice were housed singly in polycarbonate cages (32 � 14 �20 cm) and familiarized with the test procedure (see below). All behav-ioral experiments took place in the home cage of the test animals duringthe dark phase of the day–night cycle under a red light.
Animals were allowed access to food and tap water ad libitum and werekept on a reversed 12 hr light /dark cycle with lights off from 8 A.M. to 8P.M., with constant temperature. Throughout all experiments, observerswere blind to the rearing conditions. All the experiments were performedaccording to the principles of laboratory animal care published by theFrench Ethical Committee, and every attempt was made to minimize thenumber of killed animals.
BrdU labeling and detectionTo determine the quantity of newly generated cells, 5-bromo-2�-deoxyuridine (BrdU; at 15 mg/ml; Sigma, St. Louis, MO), a marker ofcell proliferation, was administered intraperitoneally (50 �g/g of bodyweight dissolved in 0.4N NaOH with 0.9% NaCl). Detection of thelabeled progeny cells was done immunohistologically after two differentsurvival times. After a 20 d period of enrichment, a single dose of BrdUwas given 4 hr before killing the animal (to assess proliferation, see Fig.4a), or four injections repeated every 2 hr were administrated to animalsbefore they were replaced in their respective cages for 20 more days (toassess neurogenesis, which includes proliferation, migration, and sur-vival; see Fig. 1a).
ImmunohistochemistryAfter odor enrichment, mice were given an overdose of sodium pento-barbital (100 mg/kg; Sanofi, France) and perfused transcardially with 50ml of saline (NaCl 0.9%) containing heparin (5 � 10 3 U/ml) at 37°Cfollowed by 200 ml of 4% paraformaldehyde in 0.1 M phosphate buffer,pH 7.3. The brain was excised and immersed overnight in the samefixative at 4°C. Coronal sections were serially cut using a vibratingmicrotome (Leica) and collected in PBS (0.1 M), pH 7.3. BrdU stainingwas performed on free-floating 40 �m sections that were pretreated byDNA denaturation (2N HCl for 30 min at room temperature). Theprimary antibodies used were rat monoclonal anti-BrdU antibody (1:200;Accurate Scientific, Harlan Sera-Lab, Loughborough, UK) and mousemonoclonal anti-NeuN antibody (1:200; Chemicon, Temecula, CA). Todetermine the number of BrdU-positive cells, staining for BrdU with theperoxidase method was used (ABC system) with biotinylated donkeyanti-rat IgG antibodies (1:200) and diaminobenzidine (0.05%) as chro-mogen (Vector Laboratories, Burlingame, CA). The double-labeled im-munofluorescence was performed with the following fluorescent second-ary antibodies: Alexa 568-labeled goat anti-rat IgG antibody and Alexa488-labeled goat anti-mouse IgG antibody (both at 1:500) from Molecu-lar Probes (Eugene, OR). Cell-specific markers were used to phenotypethe newly born neurons after longer survival times. For this purpose, �12BrdU-positive cells in each of eight randomly chosen sections of theolfactory bulbs were examined for each animal (n � 5 in standard andenriched groups), and counting is expressed in percentage. Sections wereanalyzed using either a standard microscope (Leica) for the peroxidasemethod or a Zeiss confocal microscope with its complementary softwarepackage for double labeling.
Image analysis and quantificationThe Spot camera and its accompanying software (Sterling Heights, MI)were used to determine the two- or three-dimensional numerical densityof BrdU-immunoreactive (IR) cells, either by selecting predeterminedareas to assess proliferation or by analyzing the entire structure to assessneurogenesis in the main olfactory bulb and in the hippocampus.
Olfactory bulb. Immunostained nuclei visualized through a 20� objec-tive (Leica) were numbered in a one-in-three section, 120 �m apart. Thegranule cell area includes the mitral cell layer, internal plexiform layer,and granule cell layer. The number of BrdU-labeled cells per section wasthen related to granule cell layer sectional volume to express a densityper millimeter cubed.
Lateral ventricle and RMS. As depicted in Figure 4c, determined areas(50 � 50 �m) were analyzed on every third section (40 �m thickness).All BrdU-positive nuclei in these selected areas, visualized through a
Table 1. List of different natural odors used to enrich the olfactoryenvironment and chemical molecules to challenge olfactoryperformances
Olfactory enrichment Behavioral test
Lavender M-XylolGarlic OctanolPaprika VanillaMarjoram (�)-CarvonCurry (�)-CarvonRosemary Isoamyl acetateNutmeg CaprinaldehydThyme PropionaldehydBasil leaves ButyraldehydCumin ValeraldehydCardamom CapronaldehydTarragon AnethWhole cloves MassaleChocolate PaprikaCelery MintAnise CoffeeGinger NutmegLemon CurryOrange RosemaryBanana Cinnamon
2680 J. Neurosci., April 1, 2002, 22(7):2679–2689 Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis

40� objective, were counted and presented as the number of cells permillimeter squared (see Fig. 4).
Dentate gyrus. To assess survival of newly generated cells in the dentategyrus of the hippocampus, BrdU-positive cells were numbered in aone-in-two series of sections (80 �m apart) through a 40� objective(Olympus) between the stereotaxic coordinates bregma �1.40 and �2.88mm. The area and the volume of the dentate gyrus were determined(20� magnification) in a one-in-two series of sections counterstainedwith cresyl violet. The number of BrdU-labeled cells was then related togranule cell layer sectional volume to have the density per millimetercubed.
Olfactory memory and specificityOdors were presented by placing 6 �l of the odor stimulus onto a 6 mmcircle of filter paper (Whatman #1). The filter paper was put into a glassPasteur pipette. This pipette was then introduced through the center ofone side of the mesh top, so that the filter paper was �8 cm from thefloor. This procedure allowed the observer to change the odor stimuluswithout disturbing the animal. All odors were dissolved in mineral oil(10 �3) and freshly prepared before each experiment.
Three days before tests were run, animals in their home cage wereexposed to odor stimuli different from those used in the test session tofamiliarize them with the procedure. A test session consisted of two 5min odor presentations of the same odor with a 30, 120, 180, 240, or 480min interval. These different intervals were tested randomly in separatesessions spaced at least 24 hr apart. Different odors were used in each testand counterbalanced across the different delay conditions (Table 1). Werecorded the time that the animals spent rearing and sniffing at thefilter paper. A significant decrease in investigation time during the secondpresentation indicates that mice were able to recognize an odor that hadbeen presented previously. To assess the specificity of odor recognition, thesame animals in a separate experiment were presented with a first odor,followed 30, 120, 180, or 240 min later by exposure to a different one.
Retroactive interferenceNewly stored information may affect retrieval of memories acquiredearlier in time. This principle was used in a second series of experimentsaimed to examine the consequences of an enriched olfactory environ-ment on the ability of mice to retain the memory of an odor stimulusimmediately followed by the presentation of a different odor. To assessthe possibility of impairing odor recognition, enriched and standard micewere presented with a second odor (odor 2) 5 min after having beenexposed to the first one (odor 1). This was followed by a second presen-tation of odor 1, 30 min after it first presentation. Each presentationlasted 5 min, and odors were presented as described previously. The totalamount of time investigating the source of odor was recorded duringeach presentation, as in the previous experiment. A significant decreasein investigation time during the second presentation of odor 1 indicatesthat test animals were able to recognize this odor despite the presenta-tion of odor 2.
Water mazeTwenty-one adult male mice in each group were used in this experiment.Animals were housed in groups of two to three per cage immediatelyafter the end of the 40 d period. The animals were tested in a circulartank (140 cm diameter) filled to a depth of 40 cm with white-coloredwater at 23 � 2°C. The escape platform (12 cm diameter) was submerged1 cm below the surface of the water. The pool was located in a room withmultiple cues on all sides. Each mouse was tested during four daily trialson 4 successive days. The position of the platform was kept constantduring training. During the four trials, each mouse was started once fromthree start positions and allowed to search for the platform. The order ofstart positions was randomized. The trial ended either when the animalclimbed onto the platform or when a maximum of 90 sec elapsed. After theanimal found the platform, it was allowed to rest on it for 30 sec. If themouse had not found the platform at the end of the trial, it was gently ledto it and allowed to rest for 30 sec. The time taken to escape onto thesubmerged platform (escape latency) was recorded to assess performance.
A probe test was administered after the training trials on day 5. Duringthis test the platform was removed from the pool. Each animal wasstarted in a position opposite the location of the training platformposition and allowed to swim for 90 sec. The time spent by the animal inthe different quadrants was recorded.
StatisticsAnatomical data were compared between the two groups using Student’st tests for each position of the sections. Behavioral data were analyzed byANOVA with repeated measures. Olfactory memory analysis included abetween group factor (standard versus odor enriched) and two withinfactors (intervals and exposures). For water maze learning and probe testanalyses, the within factor was “days” and “quadrants,” respectively.Levels of significance were set at 0.05.
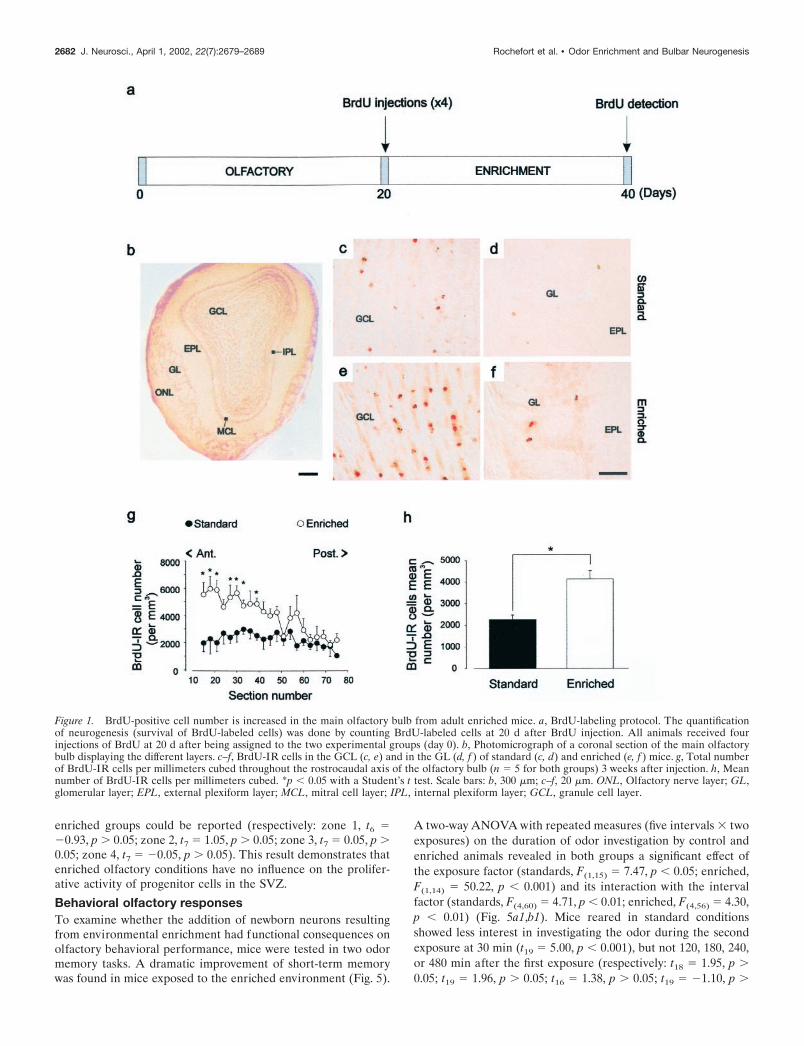
RESULTSEnriched odor exposure and survival ofBrdU-labeled cellsAfter a daily exposure to different odors and 3 weeks after BrdUinjection, most BrdU-labeled cells were found scattered through-out the granule cell layer of the olfactory bulb (Fig. 1c,e). A fewcells were also found in the external plexiform and the periglo-merular layers (Fig. 1d,f) and occasionally in the mitral cell layer(data not shown). To quantify this neurogenesis, an observerblind to the conditions counted the labeled profiles from bothgroups. The mean number of BrdU-labeled cells per millimeterscubed found in the main olfactory bulb was significantly higher inthe enriched group than in the standard one (t8 � 2.94; p � 0.05)(Fig. 1g,h). An analysis of simple comparisons confirmed thatdifferences between standard and enriched groups were highlysignificant in the anterior part of the rostrocaudal axis fromsection 15 to section 39 (respectively: t6 � 3.29, p � 0.05; t7 �2.55, p � 0.05; t7 � 3.89, p � 0.01; t7 �2.65, p � 0.05; t7 � 3.36,p � 0.01; t7 � 3.10, p � 0.05; t8 � 4.29, p � 0.01) (Fig. 1g).
Differentiation of newly generated cells was examined in themain olfactory bulb by double labeling for BrdU and a neuronalmarker, NeuN (Mullen et al., 1992). We used confocal microscopyto count the number of double- or single-labeled BrdU-positivecells present in enriched and standard olfactory bulbs (Fig. 2). Inall cases, cell counts revealed that BrdU-labeled cells were mostlyNeuN positive (Fig. 2a,b). Cells labeled with BrdU but not NeuN(Fig. 2c,d) are likely to be either glial cells or neurons that have notyet begun to express NeuN. Interestingly, the percentage of BrdU-labeled cells that expressed the neuronal-specific marker did notdiffer between enriched and standard groups (respectively: 85.8 �2.7% and 81.6 � 3.2%; t8 � �1.01; p 0.05) (Fig. 2e). From theseresults, we inferred that the enhanced number of BrdU-positivecells seen in Figure 1h resulted from an increase in the newbornneurons in the experimental bulbs. This finding may result fromincreased cell proliferation, survival, or both.
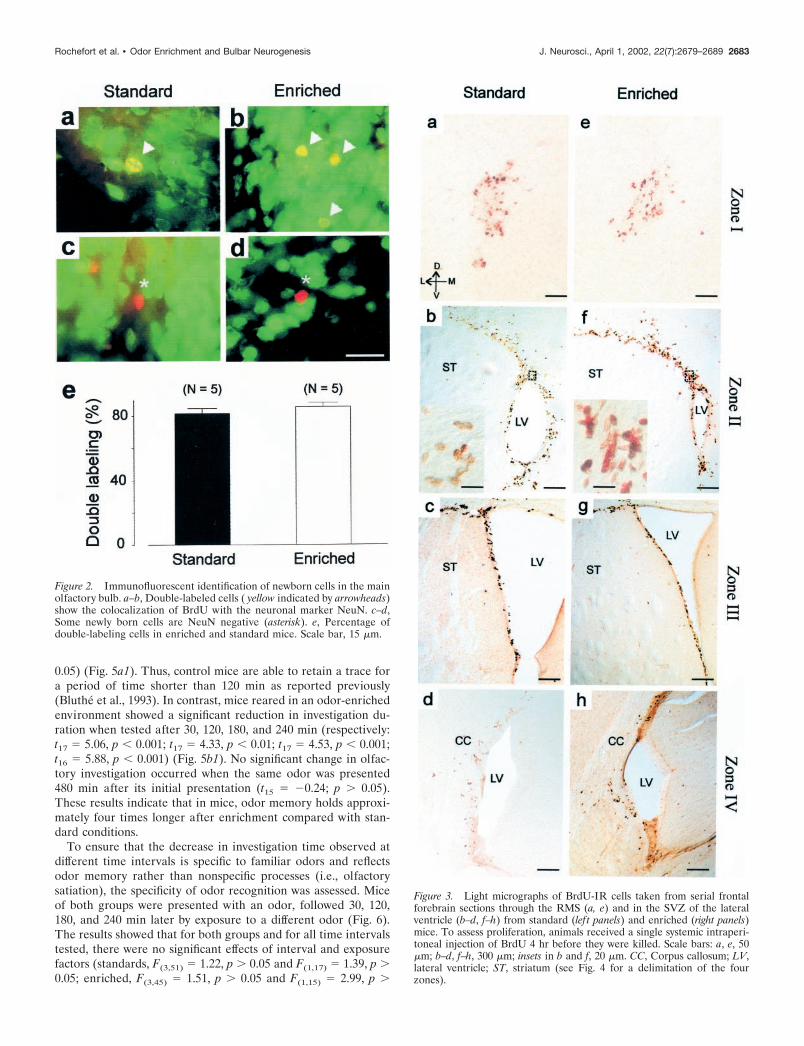
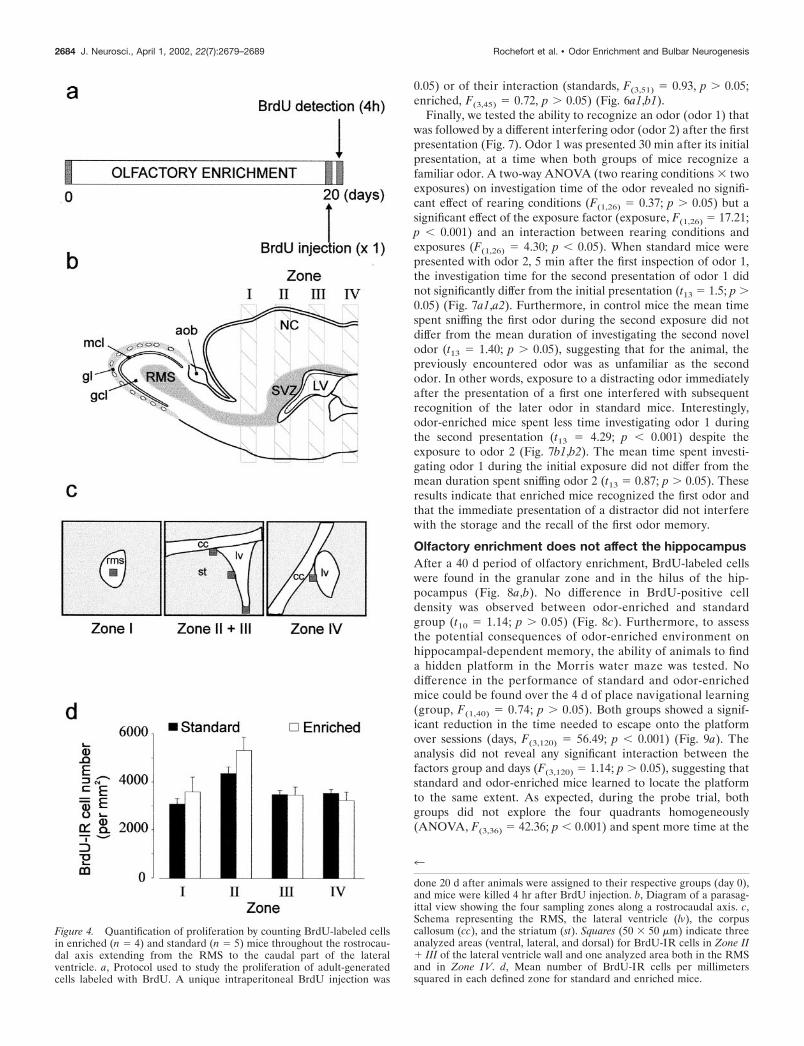
Enriched olfactory environment does not affectcell proliferationFour hours after a single BrdU injection, animals were killed toassess the proliferation of the progenitors (see Fig. 4a). Cellcounts were performed along the entire rostrocaudal axis of thelateral ventricles including the RMS (Fig. 3a,e), the anterior (Figs.3b,c,f,g), and the posterior parts of the lateral ventricles (Figs.3d,h). The rates of cell proliferation were quantified by countingthe profile number of BrdU-labeled nuclei within the four rostro-caudal regions of the SVZ (see delimitation of zones in Fig. 4b).No significant difference was found between the odor-enriched(n � 4) and standard groups (n � 5) in all of the regions thatimmediately surround the lateral ventricles when expressed asdensity (respectively: zone 1, t7 � �0.76, p 0.05; zone 2, t7 �1.58, p 0.05; zone 3, t7 � 0.06, p 0.05; zone 4, t7 � 0.78, p 0.05) (Fig. 4d). Because a difference could appear in others regionof the SVZ, cells counts were also performed on the entiresection. Similarly, no difference between standard and odor-
Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis J. Neurosci., April 1, 2002, 22(7):2679–2689 2681
enriched groups could be reported (respectively: zone 1, t6 ��0.93, p 0.05; zone 2, t7 � 1.05, p 0.05; zone 3, t7 � 0.05, p 0.05; zone 4, t7 � �0.05, p 0.05). This result demonstrates thatenriched olfactory conditions have no influence on the prolifer-ative activity of progenitor cells in the SVZ.
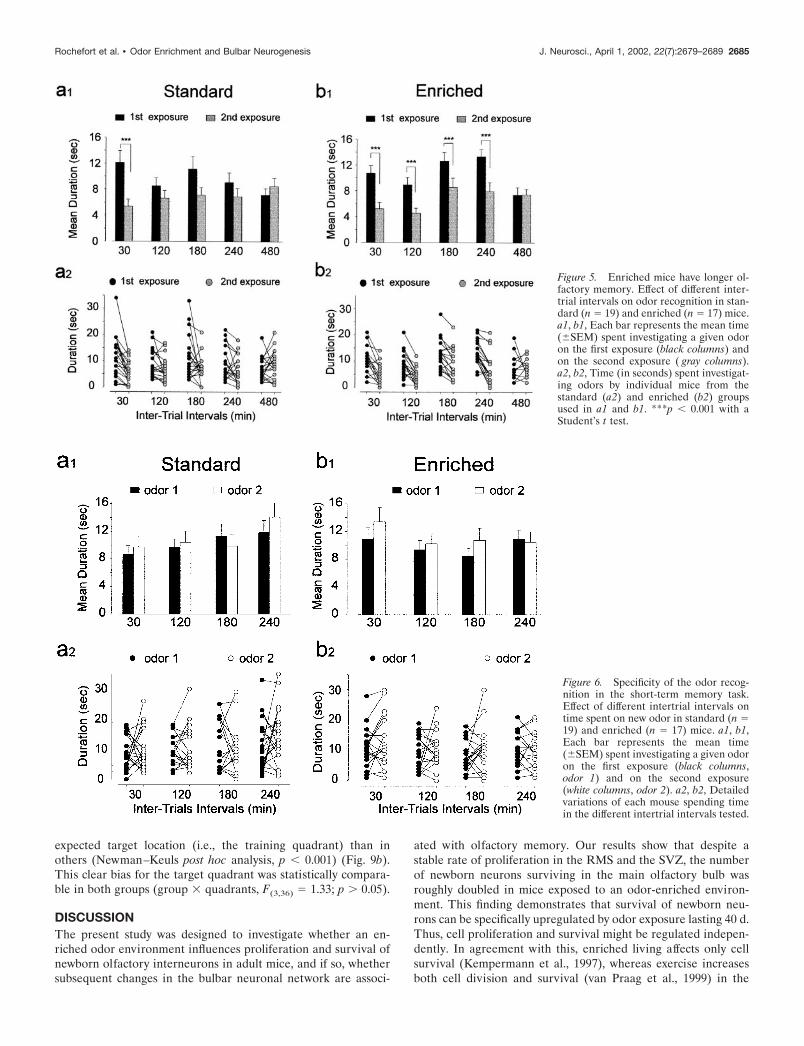
Behavioral olfactory responsesTo examine whether the addition of newborn neurons resultingfrom environmental enrichment had functional consequences onolfactory behavioral performance, mice were tested in two odormemory tasks. A dramatic improvement of short-term memorywas found in mice exposed to the enriched environment (Fig. 5).
A two-way ANOVA with repeated measures (five intervals � twoexposures) on the duration of odor investigation by control andenriched animals revealed in both groups a significant effect ofthe exposure factor (standards, F(1,15) � 7.47, p � 0.05; enriched,F(1,14) � 50.22, p � 0.001) and its interaction with the intervalfactor (standards, F(4,60) � 4.71, p � 0.01; enriched, F(4,56) � 4.30,p � 0.01) (Fig. 5a1,b1). Mice reared in standard conditionsshowed less interest in investigating the odor during the secondexposure at 30 min (t19 � 5.00, p � 0.001), but not 120, 180, 240,or 480 min after the first exposure (respectively: t18 � 1.95, p 0.05; t19 � 1.96, p 0.05; t16 � 1.38, p 0.05; t19 � �1.10, p
Figure 1. BrdU-positive cell number is increased in the main olfactory bulb from adult enriched mice. a, BrdU-labeling protocol. The quantificationof neurogenesis (survival of BrdU-labeled cells) was done by counting BrdU-labeled cells at 20 d after BrdU injection. All animals received fourinjections of BrdU at 20 d after being assigned to the two experimental groups (day 0). b, Photomicrograph of a coronal section of the main olfactorybulb displaying the different layers. c–f, BrdU-IR cells in the GCL (c, e) and in the GL (d, f ) of standard (c, d) and enriched (e, f ) mice. g, Total numberof BrdU-IR cells per millimeters cubed throughout the rostrocaudal axis of the olfactory bulb (n � 5 for both groups) 3 weeks after injection. h, Meannumber of BrdU-IR cells per millimeters cubed. *p � 0.05 with a Student’s t test. Scale bars: b, 300 �m; c–f, 20 �m. ONL, Olfactory nerve layer; GL,glomerular layer; EPL, external plexiform layer; MCL, mitral cell layer; IPL, internal plexiform layer; GCL, granule cell layer.
2682 J. Neurosci., April 1, 2002, 22(7):2679–2689 Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis
0.05) (Fig. 5a1). Thus, control mice are able to retain a trace fora period of time shorter than 120 min as reported previously(Bluthe et al., 1993). In contrast, mice reared in an odor-enrichedenvironment showed a significant reduction in investigation du-ration when tested after 30, 120, 180, and 240 min (respectively:t17 � 5.06, p � 0.001; t17 � 4.33, p � 0.01; t17 � 4.53, p � 0.001;t16 � 5.88, p � 0.001) (Fig. 5b1). No significant change in olfac-tory investigation occurred when the same odor was presented480 min after its initial presentation (t15 � �0.24; p 0.05).These results indicate that in mice, odor memory holds approxi-mately four times longer after enrichment compared with stan-dard conditions.
To ensure that the decrease in investigation time observed atdifferent time intervals is specific to familiar odors and reflectsodor memory rather than nonspecific processes (i.e., olfactorysatiation), the specificity of odor recognition was assessed. Miceof both groups were presented with an odor, followed 30, 120,180, and 240 min later by exposure to a different odor (Fig. 6).The results showed that for both groups and for all time intervalstested, there were no significant effects of interval and exposurefactors (standards, F(3,51) � 1.22, p 0.05 and F(1,17) � 1.39, p 0.05; enriched, F(3,45) � 1.51, p 0.05 and F(1,15) � 2.99, p
Figure 3. Light micrographs of BrdU-IR cells taken from serial frontalforebrain sections through the RMS (a, e) and in the SVZ of the lateralventricle (b–d, f–h) from standard (lef t panels) and enriched (right panels)mice. To assess proliferation, animals received a single systemic intraperi-toneal injection of BrdU 4 hr before they were killed. Scale bars: a, e, 50�m; b–d, f–h, 300 �m; insets in b and f, 20 �m. CC, Corpus callosum; LV,lateral ventricle; ST, striatum (see Fig. 4 for a delimitation of the fourzones).
Figure 2. Immunofluorescent identification of newborn cells in the mainolfactory bulb. a–b, Double-labeled cells ( yellow indicated by arrowheads)show the colocalization of BrdU with the neuronal marker NeuN. c–d,Some newly born cells are NeuN negative (asterisk). e, Percentage ofdouble-labeling cells in enriched and standard mice. Scale bar, 15 �m.
Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis J. Neurosci., April 1, 2002, 22(7):2679–2689 2683
0.05) or of their interaction (standards, F(3,51) � 0.93, p 0.05;enriched, F(3,45) � 0.72, p 0.05) (Fig. 6a1,b1).
Finally, we tested the ability to recognize an odor (odor 1) thatwas followed by a different interfering odor (odor 2) after the firstpresentation (Fig. 7). Odor 1 was presented 30 min after its initialpresentation, at a time when both groups of mice recognize afamiliar odor. A two-way ANOVA (two rearing conditions � twoexposures) on investigation time of the odor revealed no signifi-cant effect of rearing conditions (F(1,26) � 0.37; p 0.05) but asignificant effect of the exposure factor (exposure, F(1,26) � 17.21;p � 0.001) and an interaction between rearing conditions andexposures (F(1,26) � 4.30; p � 0.05). When standard mice werepresented with odor 2, 5 min after the first inspection of odor 1,the investigation time for the second presentation of odor 1 didnot significantly differ from the initial presentation (t13 � 1.5; p 0.05) (Fig. 7a1,a2). Furthermore, in control mice the mean timespent sniffing the first odor during the second exposure did notdiffer from the mean duration of investigating the second novelodor (t13 � 1.40; p 0.05), suggesting that for the animal, thepreviously encountered odor was as unfamiliar as the secondodor. In other words, exposure to a distracting odor immediatelyafter the presentation of a first one interfered with subsequentrecognition of the later odor in standard mice. Interestingly,odor-enriched mice spent less time investigating odor 1 duringthe second presentation (t13 � 4.29; p � 0.001) despite theexposure to odor 2 (Fig. 7b1,b2). The mean time spent investi-gating odor 1 during the initial exposure did not differ from themean duration spent sniffing odor 2 (t13 � 0.87; p 0.05). Theseresults indicate that enriched mice recognized the first odor andthat the immediate presentation of a distractor did not interferewith the storage and the recall of the first odor memory.
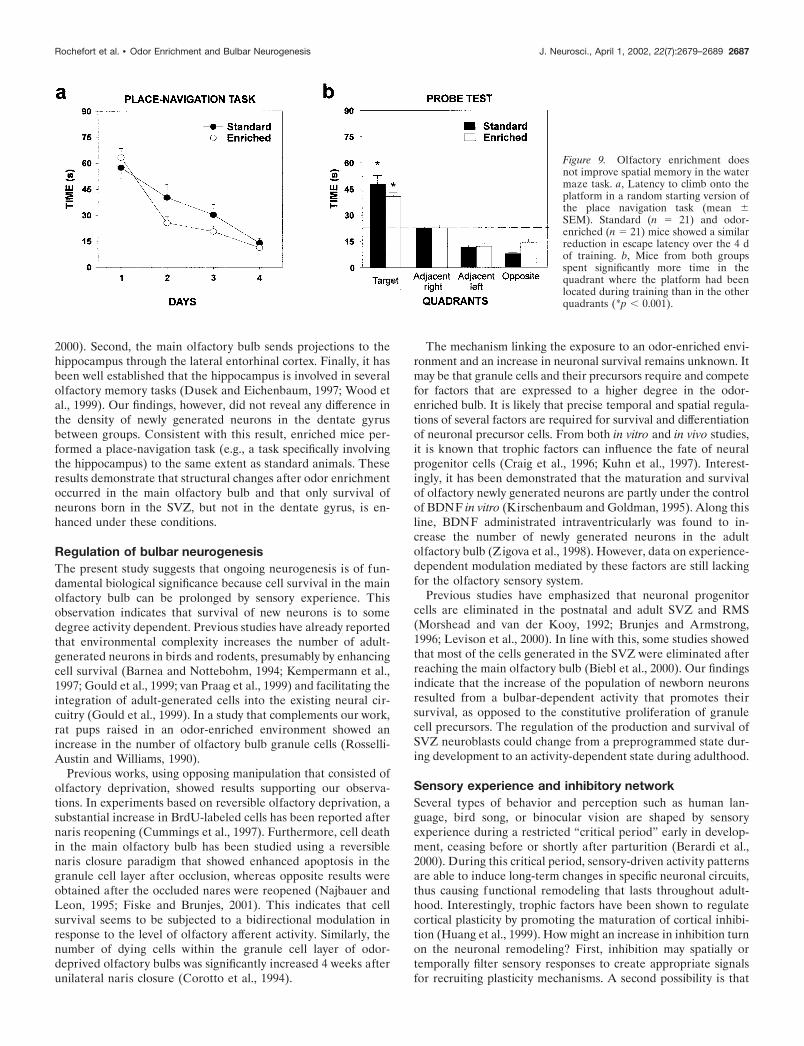
Olfactory enrichment does not affect the hippocampusAfter a 40 d period of olfactory enrichment, BrdU-labeled cellswere found in the granular zone and in the hilus of the hip-pocampus (Fig. 8a,b). No difference in BrdU-positive celldensity was observed between odor-enriched and standardgroup (t10 � 1.14; p 0.05) (Fig. 8c). Furthermore, to assessthe potential consequences of odor-enriched environment onhippocampal-dependent memory, the ability of animals to finda hidden platform in the Morris water maze was tested. Nodifference in the performance of standard and odor-enrichedmice could be found over the 4 d of place navigational learning(group, F(1,40) � 0.74; p 0.05). Both groups showed a signif-icant reduction in the time needed to escape onto the platformover sessions (days, F(3,120) � 56.49; p � 0.001) (Fig. 9a). Theanalysis did not reveal any significant interaction between thefactors group and days (F(3,120) � 1.14; p 0.05), suggesting thatstandard and odor-enriched mice learned to locate the platformto the same extent. As expected, during the probe trial, bothgroups did not explore the four quadrants homogeneously(ANOVA, F(3,36) � 42.36; p � 0.001) and spent more time at the
4
done 20 d after animals were assigned to their respective groups (day 0),and mice were killed 4 hr after BrdU injection. b, Diagram of a parasag-ittal view showing the four sampling zones along a rostrocaudal axis. c,Schema representing the RMS, the lateral ventricle (lv), the corpuscallosum (cc), and the striatum (st). Squares (50 � 50 �m) indicate threeanalyzed areas (ventral, lateral, and dorsal) for BrdU-IR cells in Zone II� III of the lateral ventricle wall and one analyzed area both in the RMSand in Zone IV. d, Mean number of BrdU-IR cells per millimeterssquared in each defined zone for standard and enriched mice.
Figure 4. Quantification of proliferation by counting BrdU-labeled cellsin enriched (n � 4) and standard (n � 5) mice throughout the rostrocau-dal axis extending from the RMS to the caudal part of the lateralventricle. a, Protocol used to study the proliferation of adult-generatedcells labeled with BrdU. A unique intraperitoneal BrdU injection was
2684 J. Neurosci., April 1, 2002, 22(7):2679–2689 Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis
expected target location (i.e., the training quadrant) than inothers (Newman–Keuls post hoc analysis, p � 0.001) (Fig. 9b).This clear bias for the target quadrant was statistically compara-ble in both groups (group � quadrants, F(3,36) � 1.33; p 0.05).
DISCUSSIONThe present study was designed to investigate whether an en-riched odor environment influences proliferation and survival ofnewborn olfactory interneurons in adult mice, and if so, whethersubsequent changes in the bulbar neuronal network are associ-
ated with olfactory memory. Our results show that despite astable rate of proliferation in the RMS and the SVZ, the numberof newborn neurons surviving in the main olfactory bulb wasroughly doubled in mice exposed to an odor-enriched environ-ment. This finding demonstrates that survival of newborn neu-rons can be specifically upregulated by odor exposure lasting 40 d.Thus, cell proliferation and survival might be regulated indepen-dently. In agreement with this, enriched living affects only cellsurvival (Kempermann et al., 1997), whereas exercise increasesboth cell division and survival (van Praag et al., 1999) in the
Figure 5. Enriched mice have longer ol-factory memory. Effect of different inter-trial intervals on odor recognition in stan-dard (n � 19) and enriched (n � 17) mice.a1, b1, Each bar represents the mean time(�SEM) spent investigating a given odoron the first exposure (black columns) andon the second exposure ( gray columns).a2, b2, Time (in seconds) spent investigat-ing odors by individual mice from thestandard (a2) and enriched (b2) groupsused in a1 and b1. ***p � 0.001 with aStudent’s t test.
Figure 6. Specificity of the odor recog-nition in the short-term memory task.Effect of different intertrial intervals ontime spent on new odor in standard (n �19) and enriched (n � 17) mice. a1, b1,Each bar represents the mean time(�SEM) spent investigating a given odoron the first exposure (black columns,odor 1) and on the second exposure(white columns, odor 2). a2, b2, Detailedvariations of each mouse spending timein the different intertrial intervals tested.
Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis J. Neurosci., April 1, 2002, 22(7):2679–2689 2685
hippocampus. In the main olfactory bulb, the increase in the totalnumber of newborn granule cells is accompanied by a longer andstronger (e.g., resistance to retroactive interference) olfactorymemory. Although this result does not demonstrate a direct linkbetween neurogenesis and behavioral performance, it is likelythat a combination of an increase in the number of olfactory bulbinterneurons and structural or functional changes at synaptic and
dendritic levels, as well as the release of factors mediated bybulbar activity, may participate to improve olfactory memory.
In the present study, we were also interested in investigating theeffects of odor enrichment on hippocampal neurogenesis forthree main reasons. First, similar relationships between morpho-logical and behavioral changes have been reported recently in thehippocampus after environmental enrichment (Rampon et al.,
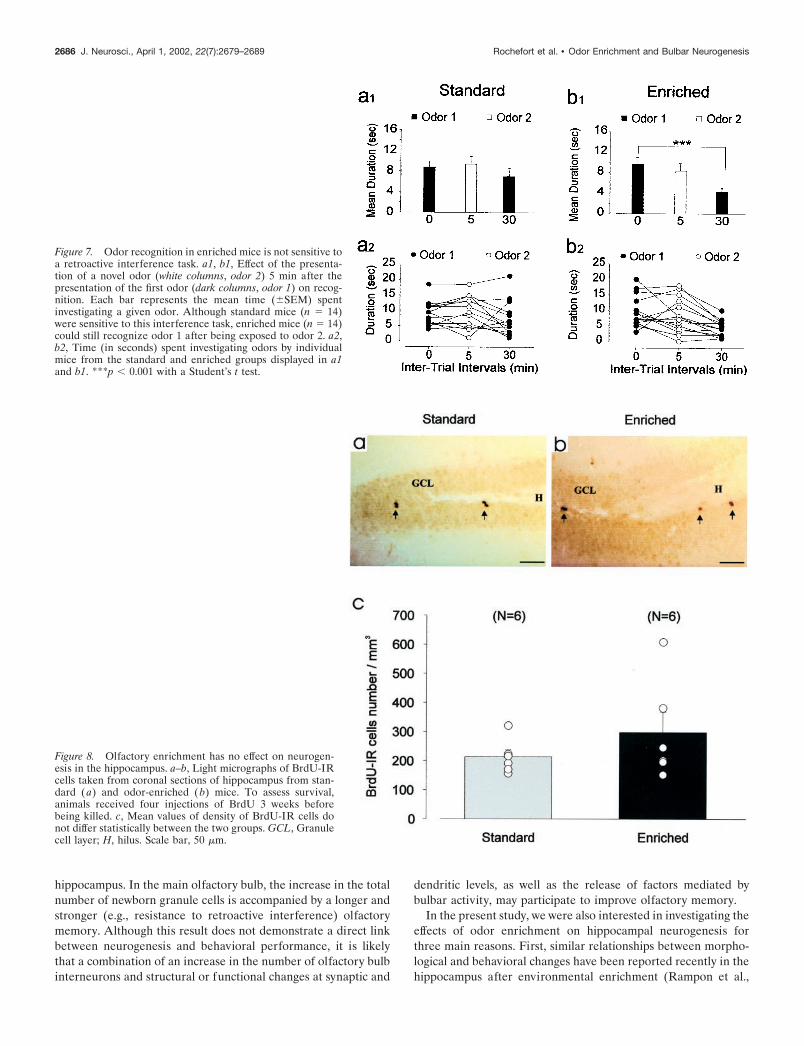
Figure 7. Odor recognition in enriched mice is not sensitive toa retroactive interference task. a1, b1, Effect of the presenta-tion of a novel odor (white columns, odor 2) 5 min after thepresentation of the first odor (dark columns, odor 1) on recog-nition. Each bar represents the mean time (�SEM) spentinvestigating a given odor. Although standard mice (n � 14)were sensitive to this interference task, enriched mice (n � 14)could still recognize odor 1 after being exposed to odor 2. a2,b2, Time (in seconds) spent investigating odors by individualmice from the standard and enriched groups displayed in a1and b1. ***p � 0.001 with a Student’s t test.
Figure 8. Olfactory enrichment has no effect on neurogen-esis in the hippocampus. a–b, Light micrographs of BrdU-IRcells taken from coronal sections of hippocampus from stan-dard (a) and odor-enriched (b) mice. To assess survival,animals received four injections of BrdU 3 weeks beforebeing killed. c, Mean values of density of BrdU-IR cells donot differ statistically between the two groups. GCL, Granulecell layer; H, hilus. Scale bar, 50 �m.
2686 J. Neurosci., April 1, 2002, 22(7):2679–2689 Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis
2000). Second, the main olfactory bulb sends projections to thehippocampus through the lateral entorhinal cortex. Finally, it hasbeen well established that the hippocampus is involved in severalolfactory memory tasks (Dusek and Eichenbaum, 1997; Wood etal., 1999). Our findings, however, did not reveal any difference inthe density of newly generated neurons in the dentate gyrusbetween groups. Consistent with this result, enriched mice per-formed a place-navigation task (e.g., a task specifically involvingthe hippocampus) to the same extent as standard animals. Theseresults demonstrate that structural changes after odor enrichmentoccurred in the main olfactory bulb and that only survival ofneurons born in the SVZ, but not in the dentate gyrus, is en-hanced under these conditions.
Regulation of bulbar neurogenesisThe present study suggests that ongoing neurogenesis is of fun-damental biological significance because cell survival in the mainolfactory bulb can be prolonged by sensory experience. Thisobservation indicates that survival of new neurons is to somedegree activity dependent. Previous studies have already reportedthat environmental complexity increases the number of adult-generated neurons in birds and rodents, presumably by enhancingcell survival (Barnea and Nottebohm, 1994; Kempermann et al.,1997; Gould et al., 1999; van Praag et al., 1999) and facilitating theintegration of adult-generated cells into the existing neural cir-cuitry (Gould et al., 1999). In a study that complements our work,rat pups raised in an odor-enriched environment showed anincrease in the number of olfactory bulb granule cells (Rosselli-Austin and Williams, 1990).
Previous works, using opposing manipulation that consisted ofolfactory deprivation, showed results supporting our observa-tions. In experiments based on reversible olfactory deprivation, asubstantial increase in BrdU-labeled cells has been reported afternaris reopening (Cummings et al., 1997). Furthermore, cell deathin the main olfactory bulb has been studied using a reversiblenaris closure paradigm that showed enhanced apoptosis in thegranule cell layer after occlusion, whereas opposite results wereobtained after the occluded nares were reopened (Najbauer andLeon, 1995; Fiske and Brunjes, 2001). This indicates that cellsurvival seems to be subjected to a bidirectional modulation inresponse to the level of olfactory afferent activity. Similarly, thenumber of dying cells within the granule cell layer of odor-deprived olfactory bulbs was significantly increased 4 weeks afterunilateral naris closure (Corotto et al., 1994).
The mechanism linking the exposure to an odor-enriched envi-ronment and an increase in neuronal survival remains unknown. Itmay be that granule cells and their precursors require and competefor factors that are expressed to a higher degree in the odor-enriched bulb. It is likely that precise temporal and spatial regula-tions of several factors are required for survival and differentiationof neuronal precursor cells. From both in vitro and in vivo studies,it is known that trophic factors can influence the fate of neuralprogenitor cells (Craig et al., 1996; Kuhn et al., 1997). Interest-ingly, it has been demonstrated that the maturation and survivalof olfactory newly generated neurons are partly under the controlof BDNF in vitro (Kirschenbaum and Goldman, 1995). Along thisline, BDNF administrated intraventricularly was found to in-crease the number of newly generated neurons in the adultolfactory bulb (Zigova et al., 1998). However, data on experience-dependent modulation mediated by these factors are still lackingfor the olfactory sensory system.
Previous studies have emphasized that neuronal progenitorcells are eliminated in the postnatal and adult SVZ and RMS(Morshead and van der Kooy, 1992; Brunjes and Armstrong,1996; Levison et al., 2000). In line with this, some studies showedthat most of the cells generated in the SVZ were eliminated afterreaching the main olfactory bulb (Biebl et al., 2000). Our findingsindicate that the increase of the population of newborn neuronsresulted from a bulbar-dependent activity that promotes theirsurvival, as opposed to the constitutive proliferation of granulecell precursors. The regulation of the production and survival ofSVZ neuroblasts could change from a preprogrammed state dur-ing development to an activity-dependent state during adulthood.
Sensory experience and inhibitory networkSeveral types of behavior and perception such as human lan-guage, bird song, or binocular vision are shaped by sensoryexperience during a restricted “critical period” early in develop-ment, ceasing before or shortly after parturition (Berardi et al.,2000). During this critical period, sensory-driven activity patternsare able to induce long-term changes in specific neuronal circuits,thus causing functional remodeling that lasts throughout adult-hood. Interestingly, trophic factors have been shown to regulatecortical plasticity by promoting the maturation of cortical inhibi-tion (Huang et al., 1999). How might an increase in inhibition turnon the neuronal remodeling? First, inhibition may spatially ortemporally filter sensory responses to create appropriate signalsfor recruiting plasticity mechanisms. A second possibility is that
Figure 9. Olfactory enrichment doesnot improve spatial memory in the watermaze task. a, Latency to climb onto theplatform in a random starting version ofthe place navigation task (mean �SEM). Standard (n � 21) and odor-enriched (n � 21) mice showed a similarreduction in escape latency over the 4 dof training. b, Mice from both groupsspent significantly more time in thequadrant where the platform had beenlocated during training than in the otherquadrants (*p � 0.001).
Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis J. Neurosci., April 1, 2002, 22(7):2679–2689 2687

experience-dependent induced changes involve plasticity of in-hibitory circuits. These possibilities suggest that high levels ofinhibition may not be responsible for keeping neuronal plasticityclosed, but instead the major role of inhibition in vivo may be topromote the onset of neuronal remodeling.
The large-scale production of immature inhibitory interneu-rons in the main olfactory bulb provides a substrate by whichneural regulatory factors and olfactory bulb activity in the adultanimal could direct the organization of developing neuronalinhibitory connections. Although the overall effects of enrichmentseem to be beneficial, it is not clear how long these effects maylast, nor what could happen when enrichment is discontinued.Previous studies showed that morphological and chemicalchanges within the brain are reversed when animals are removedfrom the enriched environment (Bennett et al., 1974; Green andGreenough, 1986). In contrast, we observed that mice that hadlived in an enriched environment for 40 d and were then returnedto standard housing still differed from controls in olfactory mem-ory tests 2 months later (our unpublished data).
The main olfactory bulb is an important brain structure be-cause it plays a central role in odor encoding and discrimination(for review, see Mori et al., 1999). Studies have provided evidencethat, in addition to acquisition, the olfactory bulb has a transientrole in memory and demonstrates a high degree of structuralplasticity (Lynch and Granger, 1991). The relationships betweenbulbar neurogenesis and olfactory performance suggest a func-tion for these new neurons in certain types of memory. In fact, theimmature status of adult-generated bulbar interneurons makesthem uniquely qualified to participate in the requisition andtransient storage of information. Future experiments will be nec-essary to determine the mechanisms by which olfactory experi-ence mediates greater survival of newly generated neurons in themain olfactory bulb. However, it is clear from the present studythat changes in the environment of an adult animal and the waysthat an individual animal reacts to those changes can have pro-found and robust effects on the brain and the behavior of thatanimal.
REFERENCESAltman J, Das GD (1965) Autoradiographic and histological evidence of
postnatal neurogenesis in rats. J Comp Neurol 124:319–335.Barnea A, Nottebohm F (1994) Seasonal recruitment of hippocampal
neurons in adult free-ranging black-capped chickadees. Proc Natl AcadSci USA 91:11217–11221.
Bennett EL, Rosenzweig MR, Diamond MC, Morimoto H, Hebert M(1974) Effects of successive environments on brain measures. PhysiolBehav 12:621–631.
Berardi N, Pizzorusso T, Maffei L (2000) Critical periods during sensorydevelopment. Curr Opin Neurobiol 10:138–145.
Biebl M, Cooper CM, Winkler J, Kuhn HG (2000) Analysis of neuro-genesis and programmed cell death reveals a self-renewing capacity inthe adult rat brain. Neurosci Lett 291:17–20.
Bluthe RM, Gheusi G, Dantzer R (1993) Gonadal steroids influence theinvolvement of arginine vasopressin in social recognition in mice.Psychoneuroendocrinology 18:323–351.
Bonfanti L, Theodosis DT (1994) Expression of polysialylated neuralcell adhesion molecule by proliferating cells in the subependymal layerof the adult rat, in its rostral extension and in the olfactory bulb.Neuroscience 62:291–305.
Brunjes PC, Armstrong AM (1996) Apoptosis in the rostral migratorystream of the developing rat. Brain Res Dev 92:219–222.
Chazal G, Durbec P, Jankovski A, Rougon G, Cremer H (2000) Con-sequences of neural cell adhesion molecule deficiency on cell migrationin the rostral migratory stream of the mouse. J Neurosci 20:1446–1457.
Conover JC, Doetsch F, Garcia-Verdugo JM, Gale NW, YancopoulosGD, Alvarez-Buylla A (2000) Disruption of Eph/ephrin signaling af-fects migration and proliferation in the adult subventricular zone. NatNeurosci 3:1091–1097.
Corotto FS, Henegar JR, Maruniak JA (1994) Odor deprivation leads to
reduced neurogenesis and reduced neuronal survival in the olfactorybulb of the adult mouse. Neuroscience 61:739–744.
Craig CG, Tropepe V, Morshead CM, Reynolds BA, Weiss S, van derKooy D (1996) In vivo growth factor expansion of endogenous sub-ependymal neural precursor cell populations in the adult mouse brain.J Neurosci 16:2649–2658.
Cummings DM, Henning HE, Brunjes PC (1997) Olfactory bulb recov-ery after early sensory deprivation. J Neurosci 17:7433–7440.
Doetsch F, Alvarez-Buylla A (1996) Network of tangential pathways forneuronal migration in adult mammalian brain. Proc Natl Acad Sci USA93:14895–14900.
Dusek JA, Eichenbaum H (1997) The hippocampus and memory fororderly stimulus relations. Proc Natl Acad Sci USA 94:7109–7114.
Fagiolini M, Hensch TK (2000) Inhibitory threshold for critical-periodactivation in primary visual cortex. Nature 404:183–186.
Fiske BK, Brunjes PC (2001) Cell death in the developing and sensory-deprived rat olfactory bulb. J Comp Neurol 431:311–319.
Gage FH (2000) Mammalian neural stem cells. Science 287:1433–1438.Gould E, Beylin A, Tanapat P, Reeves AJ, Shors TJ (1999) Learning
enhances adult neurogenesis in the hippocampal formation. Nat Neu-rosci 2:260–265.
Green EJ, Greenough WT (1986) Altered synaptic transmission in den-tate gyrus of rats reared in complex environments: evidence fromhippocampal slices maintained in vitro. J Neurophysiol 55:739–750.
Gross CG (2000) Neurogenesis in the adult brain: death of a dogma. NatRev Neurosci 1:67–73.
Hu H (1999) Chemorepulsion of neuronal migration by Slit2 in thedeveloping mammalian forebrain. Neuron 23:703–711.
Huang ZJ, Kirkwood A, Pizzorusso T, Porciatti V, Morales B, Bear MF,Maffei L, Tonegawa S (1999) BDNF regulates the maturation of in-hibition and the critical period of plasticity in mouse visual cortex. Cell98:739–755.
Jankovski A, Garcia C, Soriano E, Sotelo C (1998) Proliferation, migra-tion and differentiation of neuronal progenitor cells in the adult mousesubventricular zone surgically separated from its olfactory bulb. EurJ Neurosci 10:3853–3868.
Kempermann G, Kuhn GH, Gage FH (1997) More hippocampal neu-rons in adult mice living in an enriched environment. Nature386:493–495.
Kennedy TE, Tessier-Lavigne M (1995) Guidance and induction ofbranch formation in developing axons by target-derived diffusible fac-tors. Curr Opin Neurobiol 5:83–90.
Kirschenbaum B, Goldman SA (1995) Brain-derived neurotrophic fac-tor promotes the survival of neurons arising from the adult rat fore-brain subependymal zone. Proc Natl Acad Sci USA 92:210–214.
Kirschenbaum B, Doetsch F, Lois C, Alvarez-Buylla A (1999) Adultsubventricular zone neuronal precursors continue to proliferate andmigrate in the absence of the olfactory bulb. J Neurosci 19:2171–2180.
Kuhn HG, Winkler J, Kempermann G, Thal LJ, Gage FH (1997) Epi-dermal growth factor and fibroblast growth factor-2 have differenteffects on neural progenitors in the adult rat brain. J Neurosci17:5820–5829.
Levison SW, Rothstein RP, Brazel CY, Young GM, Albrecht PJ (2000)Selective apoptosis within the rat subependymal zone: a plausiblemechanism for determining which lineages develop from neural cells.Dev Neurosci 22:106–115.
Lois C, Alvarez-Buylla A (1994) Long-distance neuronal migration inthe adult mammalian brain. Science 264:1145–1148.
Lois C, Garcia-Verdugo JM, Alvarez-Buylla A (1996) Chain migrationof neuronal precursors. Science 271:978–981.
Luskin MB (1993) Restricted proliferation and migration of postnatallygenerated neurons derived from the forebrain subventricular zone.Neuron 11:173–189.
Lynch G, Granger R (1991) Serial steps in memory processings: possibleclues from studies of plasticity in the olfactory-hippocampal circuit. In:Olfaction, a model system for computational neuroscience (Davis JL,Eichenbaum H, eds), pp 141–165. Cambridge, MA: MIT.
Mori K, Nagao H, Yoshihara Y (1999) The olfactory bulb: coding andprocessing of odor molecule information. Science 286:711–715.
Morshead CM, van der Kooy D (1992) Postmitotic death is the fate ofconstitutively proliferating cells in the subependymal layer of the adultmouse brain. J Neurosci 12:249–256.
Mullen RJ, Buck CR, Smith AM (1992) NeuN, a neuronal specific nu-clear protein in vertebrates. Development 116:201–211.
Murray RC, Calof AL (1999) Neuronal regeneration: lessons from theolfactory system. Semin Cell Dev Biol 10:421–431.
Najbauer J, Leon M (1995) Olfactory experience modulated apoptosisin the developing olfactory bulb. Brain Res 674:245–251.
Peretto P, Merighi A, Fasolo A, Bonfanti L (1999) The subependymallayer in rodents: a site of structural plasticity and cell migration in theadult mammalian brain. Brain Res Bull 49:221–243.
Rabin MD (1988) Experience facilitates olfactory quality discrimina-tion. Percept Psychophys 44:532–540.
Rampon C, Tang YP, Goodhouse J, Shimizu E, Kyin M, Tsien JZ (2000)Enrichment induces structural changes and recovery from nonspatial
2688 J. Neurosci., April 1, 2002, 22(7):2679–2689 Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis

memory deficits in CA1 NMDAR1-knockout mice. Nat Neurosci3:238–244.
Rosselli-Austin L, Williams J (1990) Enriched neonatal odor exposureleads to increased numbers of olfactory bulb mitral and granule cells.Dev Brain Res 51:135–137.
Rousselot P, Lois C, Alvarez-Buylla A (1995) Embryonic (PSA)N-CAM reveals chains of migrating neuroblasts between the lateralventricle and the olfactory bulb of adult mice. J Comp Neurol351:51–61.
Svendsen CN, Sofroniew MV (1996) Do central nervous system neuronsrequire target-derived neurotrophic support for survival throughoutadult life and aging? Perspect Dev Neurobiol 3:133–142.
Temple S, Alvarez-Buylla A (1999) Stem cells in the adult mammaliancentral nervous system. Curr Opin Neurobiol 9:135–141.
van Praag H, Kempermann G, Gage FH (1999) Running increases cellproliferation and neurogenesis in the adult mouse dentate gyrus. NatNeurosci 2:266–270.
Woo CC, Leon M (1995) Early olfactory enrichment and deprivationboth decrease beta-adrenergic receptor density in the main olfactorybulb of the rat. J Comp Neurol 360:634–642.
Wood ER, Dudchenko PA, Eichenbaum H (1999) The global record ofmemory in hippocampal neuronal activity. Nature 397:613–616.
Wu W, Wong K, Chen J, Jiang Z, Dupuis S, Wu JY, Rao Y (1999)Directional guidance of neuronal migration in the olfactory system bythe protein Slit. Nature 400:331–336.
Zigova T, Pencea V, Wiegand SJ, Luskin MB (1998) Intraventricularadministration of BDNF increases the number of newly generatedneurons in the adult olfactory bulb. Mol Cell Neurosci 11:234–245.
Rochefort et al. • Odor Enrichment and Bulbar Neurogenesis J. Neurosci., April 1, 2002, 22(7):2679–2689 2689





























