Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
Contents lists available at SciVerse ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Environmental and climatic control on seasonal stable isotope variation of freshwatermolluscan bivalves in the Turkana Basin (Kenya)
Hubert B. Vonhof a,⁎, Josephine C.A. Joordens a,b, Marlijn L. Noback c, Jeroen H.J.L. van der Lubbe d,Craig S. Feibel e,f, Dick Kroon g
a Faculty of Earth and Life Sciences of the VU-University Amsterdam, De Boelelaan 1085, 1081HV Amsterdam, The Netherlandsb Faculty of Archaeology, Leiden University, P.O. Box 9515, 2300 RA Leiden, The Netherlandsc Paleoanthropology, Department of Early Prehistory and Quaternary Ecology, Senckenberg Center for Human Evolution and Paleoecology,Eberhard Karls University of Tübingen, Rümelinstrasse 23, D-72070 Tübingen, Germanyd Institute of Geosciences, University of Kiel, Ludewig-Meyn-Str. 10, D-24118 Kiel, Germanye Dept. of Geology, Rutgers University, 610 Taylor Road, Piscataway, NJ 08854-8066, USAf Dept. of Anthropology, Rutgers University, 610 Taylor Road, Piscataway, NJ 08854-8066, USAg School of Geosciences, The University of Edinburgh, West Mains Road, Edinburgh EH9 3JW, UK
⁎ Corresponding author. Tel.: +31 205987366.E-mail address: [email protected] (H.B. Vonhof).
0031-0182/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.palaeo.2013.04.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 October 2012Received in revised form 13 April 2013Accepted 18 April 2013Available online 30 April 2013
Keywords:Lake TurkanaMolluscsStable isotopesPalaeoclimateWater chemistry
We present growth incremental stable isotope records (δ18O and δ13C) of modern and ~2 Ma fossil bivalveshells from the semi-arid Turkana Basin (N. Kenya, S. Ethiopia). These data suggest that seasonal cyclicityin δ18O and δ13C recorded by modern and fossil shells is driven by wet–dry seasonal changes in host waterchemistry, forced by monsoonal rainfall over the Ethiopian Highlands.Fully lacustrine shells show lower amplitude, or even absent seasonal cyclicity in comparison with deltaic orriverine shells because of the buffering effect of the large water volume in the lake setting. Riverine shellsarguably have the clearest relation to seasonally variable climate parameters. Riverine molluscs thus providea potentially valuable proxy for varying rainfall δ18O values in the Turkana Basin catchment.Cross plots of molluscan δ18O and δ13C data reveal dominant environmental control on molluscan isotopevalues with remarkably large isotopic differences between lacustrine, deltaic and riverine environments.We interpret this isotope pattern to directly result from the different mixing proportions of Omo River sourcewater with evaporated lake water in these environments. The interpretation of fossil molluscan δ18O andδ13C data in a palaeoclimatological context is not straightforward, since the potential influence of temporalchanges in lake water temperature, surface evaporation or river discharge on the δ18O budget of the lake issmaller than the isotopic shifts caused by shifting facies patterns in the sedimentary record.Even though it is clear from the rich molluscan faunas that the ~2 Ma palaeo-Lake Lorenyang must have beensignificantly less alkaline than modern Lake Turkana and likely provided good drinking water and abundantavailability of food for the different species of hominins inhabiting the region at that time, stable isotopevalues of molluscan bivalves are not suitable to record the difference in alkalinity between these two settings.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Fluctuations in climatic and environmental conditions have beeninvoked as major drivers for hominin evolution and migration inPleistocene East Africa (deMenocal, 2004; Kingston, 2007; Potts,2007; Trauth et al., 2007; Maslin and Trauth, 2009). Especially,climate change influencing rainfall seasonality may have providedkey selective forces in the evolution of the human lineage in Africa.The Turkana Basin (Kenya, Ethiopia) in the East African Rift Systemis well known for its rich hominin fossil record. Of particular interestin the Turkana Basin is the time period at ~2 million years ago (Ma),when palaeo-Lake Lorenyang occupied the Turkana Basin. In that
rights reserved.
climatically wet period, a hominin fauna including two early Homospecies and Paranthropus robustus was present in the area (Feibelet al., 1989) (Fig. 1). To better understand the climatic and environ-mental context of hominin occurrence in this area, accurate palaeo-climatic and palaeoenvironmental reconstructions are required. Ideally,such reconstructions should be based on proxy records derived fromlocal/regional climate archives.
Lake Lorenyang existed from ~2–1.7 Ma. The general hydrology ofLake Lorenyang was much like that of modern Lake Turkana, with the(palaeo) Omo River from the N as its main (~90%) water source(Yuretich, 1979; Joordens et al., 2011). In contrast with modern LakeTurkana, Lake Lorenyang had a rich molluscan fauna (Williamson,1982; Van Bocxlaer and van Damme, 2009).
Seasonality in equatorial Africa is primarily expressed asmonsoonalrainfall variation (Nicholson, 1999; Aronson et al., 2008), which is

50 km
Fig. 1. Location map of Lake Turkana with collection sites of modern and fossil molluscs indicated. Grey zones mark the general areas around the lake with outcropping Pleistocene–Holocene sedimentary sequences.
17H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
reflected in strong seasonal fluctuation of Omo River discharge intothe lake. Seasonal water balance changes affect oxygen isotope varia-tion of Turkana Basin lake waters (δ18Ow), with lowest δ18Ow valuescorresponding to peak Omo River discharge (typically occurring May–November; (Yuretich and Cerling, 1983). Seasonal variation in lake-and river-water oxygen and carbon isotope ratios is captured at high
temporal resolution in growth incremental records of aragonitic mol-luscan shells (δ18Oar and δ13Car).
Generally, the conversion of (molluscan) carbonate stable isotoperecords into climatic or environmental parameters like temperature, rain-fall amount, or lake level is a challenging task, because all these climateparameters influence molluscan stable isotope records simultaneously,

18 H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
and the relative contribution of each is difficult to deconvolve from thestable isotope data alone (Abell, 1982; Ricketts and Johnson, 1996;Ricketts and Anderson, 1997; Versteegh et al., 2010a). In a recent study(Garcin et al., 2012), a relatively precise lake level history wasreconstructed for Holocene Lake Turkana by mapping and C14 dating ofshore line deposits. In that study it was shown that stable isotope dataof molluscs have a first-order relationship with lake level variation. How-ever, if one would reconstruct lake levels based on oxygen isotope dataalone, large uncertainties in the lake level model become evident, oncemore emphasising that molluscan stable isotope records are not entirelygoverned by lake level variation.
In the present study, we aim to unravel in more detail whatclimatic and environmental parameters influence the δ18Oar andδ13Car values of molluscan shells in the Turkana Basin. To do so, weanalysed growth incremental (sclerochronological) series of stableisotope values of modern molluscan shells from the Omo River andSanderson's Gulf, which can directly be compared with hydrologicaland climatic data at seasonal resolution. These data are then com-pared with a similar dataset of ~2 million year old fossil molluscanshells from palaeo-Lake Lorenyang that occupied the Turkana Basinat that time.
2. Material and methods
2.1. Mollusc samples
Due to its high alkalinity of ~20–23 meq/l (Cerling, 1979), modernLake Turkana has a rather impoverished molluscan fauna consistingof stunted and thin-shelled specimens (Cohen, 1986; Van Dammeand Van Bocxlaer, 2009), lacking large bivalves suitable forsclerochronological isotope analysis. As a result biogenic carbonatesin modern Lake Turkana are primarily composed of ostracods andsmall molluscs. Larger modern shells can be found in areas withlower alkalinity, for example in the Omo River and occasionally(when it is flooded) in Sanderson's Gulf, located at the fringes of theOmo River Delta in the NW part of the lake. For this study, a modernOmo River shell (river oyster, Etheria elliptica) was collected in 2003by F.H. Brown in Ethiopia (coordinates: N 05°.41′ E 35°.91′). Anotherlarge shell (Chambardia wahlbergi) was collected in Sanderson's Gulfin 1968, and has been made available to us by D. van Damme.
Extant small gastropods, as analysed in this study, were collectedat the eastern shores of Lake Turkana near Koobi Fora in 2006.
Five large, well-preserved fossil bivalve shells from the TurkanaBasin were collected in 2004 and 2006 from several stratigraphiclevels in the upper Burgi Member of the Koobi Fora Formation, eastof present-day Lake Turkana (Areas 100, 102 and 123; Fig. 1,Table 1). Magneto- and cyclostratigraphic data provide stratigraphiccontrol on the molluscan shells studied (Joordens et al., 2011). All
Table 1Summarised isotope ranges data of molluscs and ostracods presented in this study. Indicatebivalves and for the complete set for the modern gastropods and ostracods. The ostracod d
Identifier Species Sed facies Strat Locati
FB2003 Omo Etheria elliptica River Modern Omo RDD68 Sangulf Chambardia wahlbergi Delta front Modern Sandecoquina Koobi Fora Melanoides tuberculata Lake Modern KoobiOstracods Various species Lake Modern From
et al. (KJ04-28 Iridina omoensis Delta Early Pleistocene Area 1KJ04-53.1 Pleiodon bentoni Lacustrine Early Pleistocene Area 1KJ04-38.2 Iridina omoensis Lacustrine Early Pleistocene Area 1KJ04-44.1 Coelatura sp. Lacustrine Early Pleistocene Area 1KJ06-21 Etheria elliptica Undetermined Early Pleistocene Area 1
fossil shells derive from sediments of the early Pleistocene (~2.0–1.7 Ma) Lorenyang lake phase.
2.2. Water samples
Water samples were collected from Lake Turkana (SE shore, Eshore, Ferguson's Gulf at the W shore), the Omo River Delta andthe Turkwel River in the SW Turkana Basin. Samples were filteredand stored in 30 ml glass bottles prior to stable isotope analysis(Table 2).
2.3. Micromilling and stable isotope analysis
Shells selected for growth incremental sampling (Table 1) wereembedded in epoxy resin. Sections of ~0.3 mm thickness were cutalong the longest growth axis of the bivalve (Fig. 2). Growth incre-mental series in the nacreous layer of the shells were sampled witha Merchantek MicroMill microsampler (Versteegh et al., 2010a).Depending on the width of the shell section, approximately 40 to100 samples (sample spacing ~100–150 μm) per bivalve weretaken. Each micromilled sample powder was collected separately ina small glass container. Stable isotope (δ18O and δ13C) measurementson shell aragonite powders were performed at the FALW-VUA stableisotope laboratory in Amsterdam, either on a Thermo Finnigan MAT252 mass spectrometer with an automated carbonate extraction line(Kiel-II device), or on a Thermo Finnigan Delta+ mass spectrometerwith a Gasbench-II preparation device. On both systems thelong-term standard deviation of a routinely analysed in-houseCaCO3 standard was b0.1‰ (1σ) for both carbon and oxygen isotoperatios. This CaCO3 standard is calibrated to NBS18, NBS19 and NBS20.Both δ18O and δ13C of shell aragonite are reported relative toVienna-Peedee belemnite (VPDB).
Most water sample δ18O analyses were performed on a ThermoFinnigan Delta+ mass spectrometer equipped with a Gasbench-II.The standard CO2 equilibration technique yields a long-term standarddeviation of a routinely analysed in-house water standard at b0.1‰for δ18O (1σ).
Water samples for which we have coupled δ2H and δ18O data wereanalysed on a Thermo Finnigan Delta XP mass spectrometer equippedwith a TC-EA pyrolysis furnace. For that instrument, injection of~0.2 μl of water sample in a He-purged TC-EA reactor at 1400°Celsiusresults in quantitative conversion to H2 and O. In the reactor, freeoxygen is converted to CO gas prior to isotope analysis. GC separationof both gases is sufficient to analyse the H2 as well as the CO from asingle injection, applying a magnet jump routine to rapidly switchbetween H2 and CO settings. Calculated averages of triplicate analysesof each sample result in a reproducibility of better than 1.5 per mille(1SD) for δ2H and 0.2 per mille (1SD) for δ18O.
All water sample isotope values are reported relative to V-SMOW.
d are minimum and maximum values as well as the median value per specimen for theataset is taken from Ng'ang'a et al. (1998). Molluscan data are from the present study.
on δ13C(‰VPDB)
δ18O(‰VPDB)
δ13C(‰VPDB)
δ13C(‰VPDB)
δ18O(‰VPDB)
δ18O(‰VPDB)
Median Median Minimum Maximum Minimum Maximum
iver −13.9 −1.9 −16.3 −7.3 −2.5 0.4rson's Gulf 0.1 −2.1 −4.6 5.0 −3.5 2.8Fora −3.7 3.4 −5.0 −3.3 2.8 3.9Ng'ang'a1998)
−0.1 3.9 −4.2 1.3 3.0 4.7
02 −1.6 −2.7 −6.4 3.3 −4.0 −1.602 2.1 3.6 1.2 4.3 2.9 4.400 −1.5 3.7 −2.8 −0.5 3.4 4.100 0.0 4.0 −3.9 2.0 4.3 4.402 n.a. n.a. n.a. n.a. n.a. n.a.

Table 2Isotope data of water samples analysed for the present study, completed with previously published water isotope data.
Number Location Date δ18O (‰VSMOW) δ2H (‰VSMOW) Data source
– Omo River – −0.9 Ng'ang'a et al. (1998)– Omo River – −1.2 to −0.1 Ricketts and Johnson (1996)– Kerio River – −0.7 Ng'ang'a et al. (1998)– Lake Turkana – 5.3 to 6.0 Ng'ang'a et al. (1998)– Lake Turkana – 5.1 to 5.7 Ricketts and Johnson (1996)– Lake Turkana – 5.6 to 6.1 37.0 to 40.0 Cerling et al. (1988)– Average Turkana source water – −1.7 Ricketts and Johnson (1996)KWJ06-1 SE shore lake Turkana 26-Jul-06 6.39 n.a. This studyKWJ06-2 Lake water at Koobi Fora spit 6-Aug-06 5.84 n.a. This studyKWJ06-3 Koobi Fora spit after local rainshower 7-Aug-06 5.57 n.a. This studyOmo-1 Omo River Delta peak flooding. N4o35.388′ E 36o02.247′ 12-Aug-04 −1.08 5.7 This studyOmo-2 Omo River Delta peak flooding. N4°28.855′ E 35°59.632' 12-Aug-04 −1.01 5 This studyTurkana-1 Lake water at Koobi Fora spit 10-Aug-04 4.44 36.9 This study09JJ-2 Turkwel River at TBI field station Jan-09 0.41 4.6 This study09JJ-3 Lake Turkana (Ferguson's Gulf) Jan-09 7.21 n.a. This study09JJ-4 Lake Turkana (Ferguson's Gulf) Jan-09 6.95 n.a. This study
19H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
2.4. Isotope equilibrium calculations
With few exceptions, freshwater bivalve shells are considered tobe in oxygen isotopic equilibrium with their host water (Dettman etal., 1999; Kaandorp et al., 2003; Versteegh et al., 2010a). Their shellsare aragonitic, requiring the application of an aragonite-based isotopeequilibrium equation for the calculation of the equilibrium oxygenisotope values for the shell (δ18Oar), and host water (δ18Ow) at specif-ic temperatures. We here apply a commonly accepted equation byGrossman and Ku (1986) in the slightly modified form suggested byDettman et al. (1999):
1000lnα ¼ 2:559 106T−2� �
þ 0:715 ð1Þ
where T is the water temperature in degrees Kelvin and α is the frac-tionation between water and aragonite described by:
α ¼ 1000þ δ18Oar VSMOWð Þ� �
= 1000þ δ18Ow VSMOWð Þ� �
: ð2Þ
While such isotope equilibrium equations for fossil shells typicallyhave two unknowns, being temperature and δ18Ow, one should gen-erally expect that temperature variation will play a subordinate rolefor Turkana molluscs. Due to the near equatorial setting of theTurkana Basin, seasonal surface water temperature variation inpresent-day Lake Turkana is very small, ranging between 27.5 and
Shell growth direction 1
Umbo
Shell growth direction 2
Umbo
Location ofsection with longest
Ventral margin
longest growthaxis
Fig. 2. Schematic representation of the micromilling technique for sclerochronologicalstable isotope analysis. Milling is done with a Merchantek Micromill, allowing sam-pling resolution of b100 μm along the growth axis of the molluscs. Most shells weremicrosampled along a section indicated as “shell growth direction 2” in this figure.
29.5 °C. Temperature variation in the deeper water is even smaller,between 25.5 and 26.5°°Celsius (Yuretich and Cerling, 1983; Cohen,1986). It is thus to be expected that observed molluscan δ18Oar varia-tion in such a tropical setting is controlled by monsoonal changes inδ18Ow rather than by variation in temperature (Kaandorp et al.,2003). We should note however, that in the very shallow waters ofSanderson's Gulf and other marginal lacustrine areas, temperaturevariation can be more extensive than the instrumental data for LakeTurkana suggests.
3. Results and discussion
3.1. Diagenetic screening
The modern Etheria elliptica shell from the Omo River, used as areference for well preserved molluscan aragonite here, shows thecharacteristic pattern of lamellar aragonite crystal growth with clearand sharp boundaries between crystals (Fig. 3A, B). Most fossil shellsalso consist of lamellar aragonite crystals, and lack evidence for re-crystallization (Fig. 3C–F). The structure of fossil aragonite occasion-ally displays evidence of minor dissolution along small cracks thatrun through the crystal (Fig. 3D), but no new formation of diageneticCaCO3 is observed. The aragonite crystals can further appear slightlymore rounded at the corners and boundaries between crystals incomparison with the modern shells (Fig. 3E–F).
Fossil shell KJ06-21 (Etheria elliptica; Fig. 3G–H) has an exceptionalgrowth structure. It exhibits a highly porous shell structure in whichlayers of thin lamellar aragonite enclose large voids in the shell. Thisstructure shows signs of dissolution as well as recrystallization in thevoids, indicating significant diagenetic alteration of this specimen.Based on these results, we reject shell KJ06-21from our dataset. Weconclude that all other fossil shells studied are chemically well pre-served and must be expected to have retained their original stableisotope composition.
3.1.1. Stable isotope ratios of modern bivalve shells gastropods andostracods
The sclerochronological isotope patterns of the modern Turkanashells span a relatively wide range in δ13Car as well as in δ18Oar values.The Omo River Etheria elliptica (Fig. 4) shows clear cyclic variation inδ18Oar values over a range of several ‰. δ13Car values show similarcyclicity, albeit in antiphase with δ18Oar cycles. The Sanderson's GulfChambardia wahlbergi (Fig. 5) shows even larger isotope variationthan the Omo shell, and is distinct in the fact that δ13Car and δ18Oar
vary in phase. Furthermore, the Sanderson's Gulf shell reaches muchhigher isotope values (both for δ13Car and δ18Oar) than the OmoRiver shell.

Fig. 3. Scanning electron microscopic (SEM) images of broken cross sections of the shells studied. A. Lamellar aragonite layers of FB2003Omo, modern Etheria elliptica from the OmoRiver. B: Detail of the aragonite crystals of FB2003Omo. C: Lamellar aragonite layers of KJ04-28, fossil Iridina omoensis from layer C3. D: Minor cracks in aragonite crystals ofKJ04-53.1, fossil Pleiodon bentoni from layer C2. E: Slightly rounded aragonite crystals with blurred borders, in KJ04-53.1. F: Detail of borders between aragonite crystals ofKJ04-53.1. G: Dissolution and recrystallization in KJ06-21, fossil E. elliptica from layer A1. H: Detail of dissolution and recrystallization in KJ06-21.
20 H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
The clearly cyclic sclerochronological isotope variation in both shellsis presumably driven by seasonally changing water supply and isotopecomposition of the Omo River. Thus, both the Sanderson's GulfChambardia wahlbergi and Omo River Etheria elliptica appear to haverecorded at least one year of environmental change in their growthincremental stable isotope values. Seasonal growth stops, which com-monly occur in freshwater bivalve shells at higher latitudes (Dettmanet al., 1999; Versteegh et al., 2010a, 2010b) are not apparent in thedataset, with the probable exception of one level in the Sanderson'sGulf mollusc where the isotope values jump several ‰ between twogrowth incremental samples. This is likely related to the specific ecologyof Chambardia, which is known to aestivate during unfavorable condi-tions (Van Bocxlaer and van Damme, 2009).
3.1.2. Oxygen isotope variation from Omo River to Lake TurkanaThe δ18Ow variation in coupled river and lake systems is relatively
well studied (see e.g. (Keith et al., 1964) for earlier work on the sub-ject). Comparatively low δ18Ow values occur in the river, which in thetropics usually display seasonal δ18Ow cyclicity due to seasonal rain-fall variation in the river catchment (Kaandorp et al., 2003). LakeTurkana is primarily (80–90%; Yuretich, 1979; Yuretich and Cerling,1983) fed by the Omo River that has measured δ18Ow values rangingfrom −1.2 to −0.1‰ (Table 2). Lake surface water in coupled river–lake systems generally exhibits higher δ18Ow values than those of therivers feeding the lake (e.g. Keith et al., 1964; Liu et al., 2009). Strongevaporation from the lake surface causes a significant δ18Ow contrastbetween the Omo River and Lake Turkana (Ricketts and Johnson,
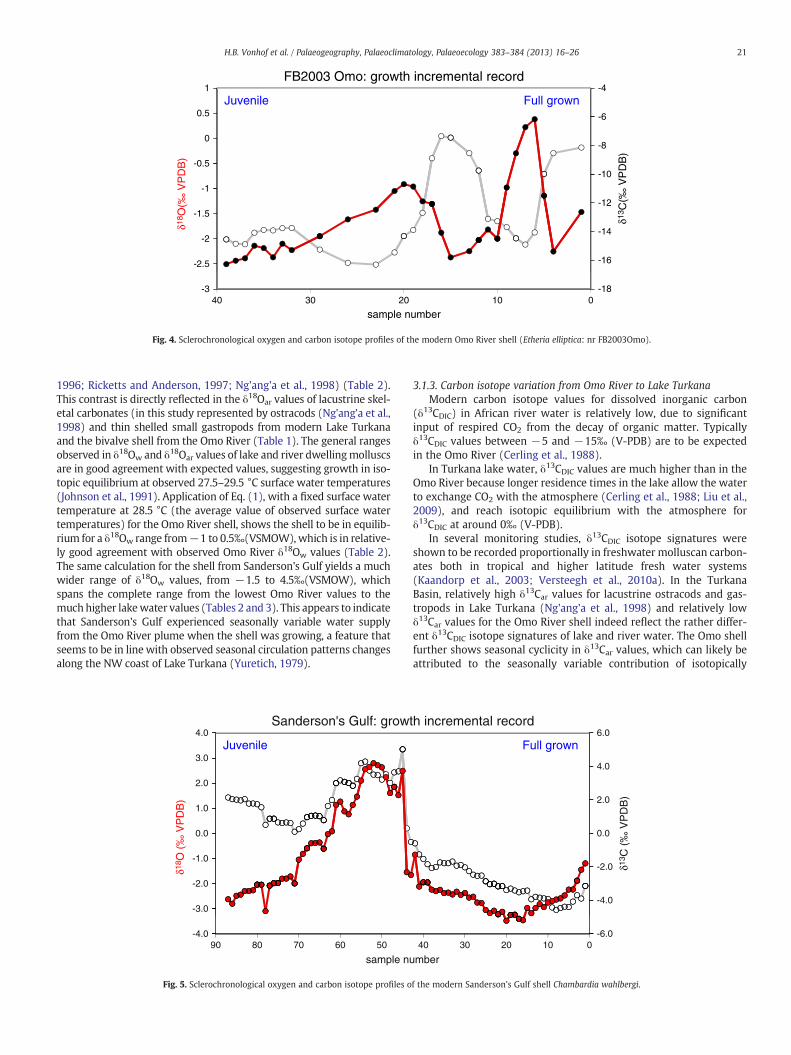
FB2003 Omo: growth incremental record1 -4
Juvenile Full grown
0
0.5 -6
-0.5
-8
-1
-12
-10δ1
8 O(‰
VP
DB
)
-1.5
-14
δ13 C
(‰ V
PD
B)
-2.5
-2
-16
-3 -18010203040
sample number
Fig. 4. Sclerochronological oxygen and carbon isotope profiles of the modern Omo River shell (Etheria elliptica: nr FB2003Omo).
21H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
1996; Ricketts and Anderson, 1997; Ng'ang'a et al., 1998) (Table 2).This contrast is directly reflected in the δ18Oar values of lacustrine skel-etal carbonates (in this study represented by ostracods (Ng'ang'a et al.,1998) and thin shelled small gastropods from modern Lake Turkanaand the bivalve shell from the Omo River (Table 1). The general rangesobserved in δ18Ow and δ18Oar values of lake and river dwellingmolluscsare in good agreement with expected values, suggesting growth in iso-topic equilibrium at observed 27.5–29.5 °C surface water temperatures(Johnson et al., 1991). Application of Eq. (1), with a fixed surface watertemperature at 28.5 °C (the average value of observed surface watertemperatures) for the Omo River shell, shows the shell to be in equilib-rium for a δ18Ow range from−1 to 0.5‰(VSMOW), which is in relative-ly good agreement with observed Omo River δ18Ow values (Table 2).The same calculation for the shell from Sanderson's Gulf yields a muchwider range of δ18Ow values, from −1.5 to 4.5‰(VSMOW), whichspans the complete range from the lowest Omo River values to themuchhigher lakewater values (Tables 2 and 3). This appears to indicatethat Sanderson's Gulf experienced seasonally variable water supplyfrom the Omo River plume when the shell was growing, a feature thatseems to be in line with observed seasonal circulation patterns changesalong the NW coast of Lake Turkana (Yuretich, 1979).
Sanderson's Gulf: growt4.0
Juvenile3.0
1.0
2.0
0.0
-1.0
δ18 O
(‰
VP
DB
)
-3.0
-2.0
-4.05060708090
sample nu
Fig. 5. Sclerochronological oxygen and carbon isotope profiles o
3.1.3. Carbon isotope variation from Omo River to Lake TurkanaModern carbon isotope values for dissolved inorganic carbon
(δ13CDIC) in African river water is relatively low, due to significantinput of respired CO2 from the decay of organic matter. Typicallyδ13CDIC values between −5 and −15‰ (V-PDB) are to be expectedin the Omo River (Cerling et al., 1988).
In Turkana lake water, δ13CDIC values are much higher than in theOmo River because longer residence times in the lake allow the waterto exchange CO2 with the atmosphere (Cerling et al., 1988; Liu et al.,2009), and reach isotopic equilibrium with the atmosphere forδ13CDIC at around 0‰ (V-PDB).
In several monitoring studies, δ13CDIC isotope signatures wereshown to be recorded proportionally in freshwater molluscan carbon-ates both in tropical and higher latitude fresh water systems(Kaandorp et al., 2003; Versteegh et al., 2010a). In the TurkanaBasin, relatively high δ13Car values for lacustrine ostracods and gas-tropods in Lake Turkana (Ng'ang'a et al., 1998) and relatively lowδ13Car values for the Omo River shell indeed reflect the rather differ-ent δ13CDIC isotope signatures of lake and river water. The Omo shellfurther shows seasonal cyclicity in δ13Car values, which can likely beattributed to the seasonally variable contribution of isotopically
h incremental record6.0
Full grown
4.0
2.0
0.0
-2.0 δ13 C
(‰
VP
DB
)
-4.0
-6.0010203040
mber
f the modern Sanderson's Gulf shell Chambardia wahlbergi.

Table 3Endmember values for river and lake water as applied in the binary mixing model ofFig. 6.
Data relevant to the δ13C binary mixing model in Fig. 6
δ13Car Lake Turkana (‰VPDB) 1δ13Car Omo River (‰VPDB) −12DIC Lake Turkana (millimol/L) 20DIC Omo River (millimol/L) 2
22 H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
light groundwater to the river (Versteegh et al., 2010a, 2010b). Dur-ing dry (wet) season, groundwater contribution to the Omo Rivercan be assumed to be relatively high (low) resulting in low (high)δ13CDIC and δ13Car values. As a result, cyclicity in δ13Car and δ18Oar isin antiphase in Omo River shells.
3.1.4. The mixing zone: isotope signatures in the Omo delta environmentThe Chambardia wahlbergi mollusc from the delta-influenced
Sanderson's Gulf spans an exceptionally wide sclerochronologicalrange of δ13Car and δ18Oar values, that bridges the gap between con-trasting δ13Car and δ18Oar values of lacustrine and riverine shells asdescribed above.
At such contrasting isotope values of water endmembers in theOmo delta, the isotope composition of deltaic water is primarilygoverned by the mixing proportions of lake and river waters. As aresult, variation of isotope composition of shells living in the deltaenvironment can reflect this mixing process, rather than local climaticparameters. In the Chambardia wahlbergi mollusc from the delta-influenced Sanderson's Gulf, higher values for both δ13Car and δ18Oar
are interpreted to indicate increasing lake water influence. Themixing process thus accounts for observed in-phase cyclicity ofδ13Car and δ18Oar, in the C. wahlbergi mollusc (Fig. 5), as opposed toantiphase cyclicity in the Omo River Etheria elliptica mollusc.
Under these conditions, the δ13Car and δ18Oar values of delta-dwelling molluscs and ostracods can potentially be used to quantita-tively reconstruct past lakewater/riverwater mixing. For δ18Oar datathis can simply be done by application of a linear mixing model,which only requires the assumed δ18Oar endmember values of lake-and river-dwelling molluscs as input. For δ13Car values the equationis slightly more complex because the DIC concentrations of LakeTurkana water is much higher than those of Omo River water. The
5
stable isotope va6
Omo River bivalve
4 modern ostracods
2
3 modern gastropods
Sanderson's Gulf
1bivalve
-1
0
δ18 O
ar (
VP
DB
)
-2
-4
-3
-20 -15 -10
δ13Car
Fig. 6. Cross plotted δ13Car and δ18Oar values of modern shells analysed, including small gastNg'ang'a et al., 1998). The blue shading indicates the modelled binary mixing pathway bendmember values and the dissolved inorganic carbon concentrations given in Table 3.
required equation in the case of δ13Car is that of binary mixing (Liuet al., 2009), in which not only the isotope values of the endmembersare input parameters, but also the DIC concentrations.
3.1.5. δ13Car and δ18Oar cross-plotsIn Fig. 6 we have cross-plotted δ13Car and δ18Oar values of modern
Lake Turkana molluscs and δ13Ccalcite and δ18Ocalcite values of ostra-cods used in this study. While molluscs are generally thought tohave negligible interspecific isotope effects (Versteegh et al., 2010a),interspecific offsets in ostracod δ18Ocalcite of up to 2‰ are document-ed (von Grafenstein et al., 1999). We note, however, that the overallisotope variation in the present study far exceeds the typical ampli-tude of such effects. In Fig. 6 we determined three fields reflectingthe typical isotope compositions of riverine, lacustrine and deltaic(mixing) environments. We further indicated the mixing hyperbolathat represents the combined variation of δ13Car and δ18Oar spanningthe complete mixing range between the selected riverine and lacus-trine endmembers. At typical modern endmember values (Table 3),the mixing hyperbola lines up reasonably well with the point cloudof observed values for the delta-influenced Chambardia wahlbergimollusc. We thus interpret this pattern to support the control ofriver–lake water mixing on the isotope composition of the C.wahlbergi specimen. Since mixing proportions are facies specific, wesuggest the combined δ13Car and δ18Oar data of molluscan bivalvesto be a robust and easy to analyse indicator of the environment(lake, river or delta) in which the molluscs lived.
3.2. Stable isotope ratios of fossil bivalve shells
The Pleistocene shells that passed diagenetic screening wereanalysed using the same techniques applied to the modern shells.The total range of δ13Car values for the fossil shells is significantlysmaller than for the modern shells, because the lower δ13Car valuesare not present in the fossil shells. The δ18Oar range of the Pleistoceneshells compares well with the range observed in the modern dataset.In Fig. 7 we have plotted the sclerochronological stable isotope pro-files of the Pleistocene shells analysed. Most show only limited iso-tope variation, particularly those specimens that exhibit relativelyhigh δ13Car and δ18Oar values. Clearest cyclicity in both δ13Car andδ18Oar occurs in shell KJ04-28. This shell is interpreted to haverecorded ~4 years of growth, at ~2‰ seasonal amplitude in δ18Oar.
0 5 10
lues modern taxa
-5
(VPDB)
ropod samples from Koobi Fora (this study) and a set of modern ostracods (taken frometween riverine and lacustrine endmember based on the oxygen and carbon isotope

110
100
90
80
70
60
Dep
th (
m)
50
40
30
20 Gastropods
KJ04-44.1
KJ04-38.2
KJ04-53.1
KJ04-28
Legend
MolluscsDiatomaceousRhizolithsAbundant fish remainsGypsum
D
C2r
Old
uvai
sub
chro
n (C
2m)
M
C1
C2 MM
Lorenyang Tuff
C3 M
KBS Tuff
< 50% sand
10
0
Fig. 7. Left panel: Sclerochronological oxygen and carbon isotope profiles of the Pleistocene molluscs analysed. Carbon isotope data in grey and oxygen isotope data in red. Rightpanel: simplified stratigraphic log of the sequences from which the Early Pleistocene fossils were derived (Joordens, 2011).
23H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
δ13Car values in this shell span ~8‰ (VPDB). Combined δ18Oar andδ13Car values for shell KJ04-28 compare remarkably well with theSanderson's Gulf shell from the modern data set, which we take tosuggest that shell KJ04-28 derives from a deltaic setting. We notethat the phase relationship between δ13Car and δ18Oar is weak whencompared with the Sanderson's Gulf shell which seems to suggestthat CO2 exchange with the atmosphere was not the only processcontrolling the isotope composition of DIC at that time.
Interpreting the isotope pattern of shell KJ04-28 to derive from adelta environment is supported by independent sedimentologicalfield evidence, placing shell KJ04-28 in a sequence of deltaic strataof the upper Burgi Formation, while all other fossil specimens studiedare from sediments interpreted to be fully lacustrine. The unique iso-tope composition of shell KJ04-28 is summarised in the δ13Car andδ18Oar cross plot shown in Fig. 8. In that plot, the fields of modernriver, delta and lake composition are indicated as derived fromFig. 6. From Fig. 8 one may conclude that most of the Pleistoceneshells indeed plot in the lacustrine field. Such an interpretation is in
good agreement with the generally low seasonal amplitude in bothδ13Car and δ18Oar of these shells, because the lacustrine seasonal var-iation of δ13CDIC and δ18Ow usually is considerably dampened com-pared with that of river water feeding the lake (Kaandorp et al.,2006). The typical modern seasonal δ18Ow and δ13CDIC range of lakesurface water near Koobi Fora spans 2‰ and 1‰ respectively(T. Cerling pers comm., Johnson et al., 1991). In contrast, typical sea-sonal δ18Ow and δ13CDIC variation for the Omo River may be as high as4 and 8‰ respectively (as recorded in the Modern Omo shell δ18Oar
and δ13Car isotope range).In the Pleistocene isotope dataset as we have it now, it appears
that no fully riverine molluscs are present.
3.3. Palaeoclimatic interpretation of oxygen isotope variation
δ18Oar values of fossil skeletal or authigenic carbonate have beenused to reconstruct palaeoclimatic variation in relation to lake levelchange in the Turkana Basin, particularly for the Holocene (Halfman

stable isotope values Lorenyang shells
5
6
4
2
3
1
-1
0δ18 O
(V
PD
B)
-2
-4
-3
0-20 -15 -10 -5 5 10
δ13C (VPDB)
44.1
38.2
28C3-2
53.1
Fig. 8. Cross plotted δ13Car and δ18Oar values of Pleistocene shells analysed, as compared with the typical fields of modern molluscan river, delta and lake dwellers determined inFig. 6.
24 H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
et al., 1989; Johnson et al., 1991; Mohammed et al., 1996; Ricketts andJohnson, 1996; Ricketts and Anderson, 1998), and also for the Pleisto-cene (Abell, 1982). These reconstructions are based on measuring inthese carbonates the balance between isotopically-light input ofriver water in the lake on the one hand, and evaporativeenrichment of 18O in lake water on the other. For the Holocene,δ18Oar profiles derived from outcrops and cores collected in LakeTurkana have therefore been interpreted to reflect climate-drivenlake level variation. A recent study by Garcin et al. (2012), combinesmolluscan δ18Oar profiles with precisely measured and C14-datedshore line deposits. This study not only demonstrates the general fitbetween lake level and δ18Oar data, but also shows that lake levelreconstructions on the basis of δ18Oar data alone have large uncer-tainties. For the Pleistocene we lack reliable independent measure-ments of lake level in the Turkana Basin, due to ongoing tectonicmovements in the area, and due to the scarcity of preserved shoreline deposits of earlier Pleistocene age. Thus it is even more difficultto couple δ18Oar data to lake level variation for the Pleistocene thanit is for the Holocene.
Still, there is valuable information to be retrieved from δ18Oar dataof Pleistocene molluscs from the Turkana Basin. The fully lacustrinePleistocene molluscs as presented in this study have δ18Oar valuesthat largely overlap with modern molluscan stable isotope values.However, from a faunal perspective it is clear that Pleistocene LakeLorenyang was much less alkaline than modern Lake Turkana. Thriv-ing Pleistocene molluscan faunas are very unlikely to have sustainedalkalinities of more than 16 meq/L (Cerling, 1979; Abell, 1982),while modern Lake Turkana has values of ~22 meq/L (hence theimpoverished modern molluscan fauna). It thus appears that a signif-icant contrast in evaporation-driven lacustrine alkalinity betweenmodern and Pleistocene times does not translate into significantδ18Oar differences. This apparent insensitivity of δ18Oar to increasingalkalinity can be understood by considering the fractionation effectsof the input of water to the lake by riverine discharge, and waterloss through evaporation. It has been estimated that evaporationremoves a ~2.3 m watercolumn from the surface of the lake on anannual basis (Yuretich and Cerling, 1983), which is balanced byOmo River discharge, as long as lake levels are more or less stable.Thus, ongoing evaporative enrichment of lacustrine δ18Ow values iscounteracted by continuous input of isotopically light δ18Oar valuesfrom the Omo River. This input/output balance causes the evaporativeδ18Ow enrichment of lake water as compared with the Omo River
source water not to exceed the typical fractionation between surfacewater and evaporated moisture. In the Turkana Basin, with surfacewater temperatures close to 30°°Celsius this fractionation is in theorder of 8‰ (Majoube, 1971; Ricketts and Johnson, 1996). Whenevaporation has increased lake surface water δ18Ow by ~8‰, theisotopic composition of evaporating moisture approaches that of theriverine input, resulting in stabilisation of lacustrine δ18Ow values.
In a previous study of the δ18Ow evolution of Holocene LakeTurkana, Ricketts and Johnson (1996) presented model experimentswhich show that in response to changes in the input and/or outputrates of the system, it takes the lake no more than a few decades toreach such stabilised lacustrine δ18Ow values, independent of lakelevel. Such isotopic stabilisation of lake water is much more rapidthan the increase of alkalinity in terminal lake systems. For Turkana,mass balance calculations based on the modern chloride budget ofthe lake and the contributing rivers show that Lake Turkana salinityand alkalinity have been increasing steadily for the past 3000–6000 years (Yuretich and Cerling, 1983; Cerling, 1986; Barton et al.,1987). Lacustrine δ18Ow evolution over that same time interval isbelieved to be controlled by slowly changing δ18Ow value of riverineinput to Lake Turkana (Ricketts and Johnson, 1996) which has nodirect relation to lacustrine alkalinity. In our view, it is this decouplingbetween lacustrine δ18Ow and alkalinity that explains why shells fromthe supposedly much less alkaline lake waters of the ~2.0–1.7 MaLorenyang lake phase can have δ18Oar values that are no differentfrom those in modern Lake Turkana (Fig. 9).
In comparison with modern and early Pleistocene shells reportedhere, remarkably low δ18Oar values around 0‰ (VPDB) of gastropodsand bivalves have been reported from early Holocene Lake Turkana(e.g. Garcin et al., 2012). It has been suggested that these low valuesmust have been caused by longer-lasting changes in the oxygen iso-tope composition of the inflow to the lake (i.e. changes of the isotopecomposition of Omo water (Ricketts and Johnson, 1996). Nowadays,the δ18Ow value of Omo River water is relatively high. This is believedto be due to contribution of Atlantic moisture from the Congo Basin tothe Omo catchment, controlled by the position of the “Congo AirBoundary”, which separates Atlantic from Indian Ocean moisturesources (Levin et al., 2009). A different configuration of the CongoAir Boundary has been suggested for the early Holocene (Tierney etal., 2011), potentially causing increased Indian Ocean-derived mois-ture in the Omo catchment. This would have resulted in significantlylower δ18Ow values of early Holocene Omo River water as compared

Fig. 9. Stratigraphic plot of oxygen and carbon isotope ranges of Pleistocene and modern molluscs studied here. Isotope data are plotted as “box and whisker” diagrams (showingmedian, upper and lower quartile, and minimum and maximum isotope values of each specimen studied. Regardless of their stratigraphic age, specimens from lacustrine faciesgenerally have overlapping isotope values. Specimens from what we interpret to be deltaic and riverine facies have distinctively lower isotope values, most clearly so in theδ18Oar data. Horizontal dashed line separates modern specimens (above) from early Pleistocene specimens (below).
25H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
with the present, as reflected in observed low δ18Oar values of earlyand middle Holocene shell aragonite from Lake Turkana (Rickettsand Johnson, 1996; Levin et al., 2009; Garcin et al., 2012). In this con-text it is noteworthy that the higher oxygen isotope values that weobserve for early Pleistocene Lake Lorenyang molluscs then suggestan atmospheric configuration which resembles modern conditionsmore than early Holocene conditions.
For part of the early Pleistocene, riverine outflow from the TurkanaBasin has been suggested, for example based on biogeographical evi-dence (Feibel, 1993), or motivated by geochemical arguments(Joordens et al., 2011). For reasons outlined above, riverine outflowfrom Lake Lorenyang cannot be provenwith the presently available sta-ble isotope data of shells from this time interval. However, an outflowwould be in line with lower lacustrine salinity and alkalinity, asinterpreted from thriving early Pleistocene molluscan faunas (Cerling,1979).
3.4. Use of sclerochronological isotope data for palaeoanthropologicalresearch in the Turkana Basin
In recent studies, the impact of Milankovitch-forced ariditychanges on hominin evolution and migration in East Africa has beenthe focus of attention (Trauth et al., 2007; Maslin and Trauth, 2009;Joordens et al., 2011). Much less investigated until now, is the poten-tially significant impact of changes in seasonal aridity on earlyhominin faunas. In this study we have demonstrated that aquaticbivalves are faithful recorders of seasonal variation of water chemis-try of their host water. However, when bivalve shells live in a lake,they will record the seasonal variation in lake water δ18Ow andδ13CDIC, which tends to be dampened considerably as comparedwith the seasonal variation present in rainwater, and in the riversfeeding the lake. Thus the amplitude of sclerocheronological δ13Cand δ18O cyclicity in freshwater bivalves is controlled more by theshell's habitat than by actual climate seasonality. Arguably the bestapproximation of the strength of seasonality can be retrieved fromriverine shells, because river water records the seasonally changing
δ18Ow values of rainwater in its catchment much more directly thanlake water.
In this study we have further shown how combined δ18O and δ13Canalyses of bivalve shells typify the environment in which they lived,with riverine shells exclusively showing low δ18O and δ13C values, gen-erally with antiphased sclerochronological δ18Oar and δ13Car cyclicity.This means that we can geochemically identify riverine shells in fossilcollections with little effort, after which full sclerochronological stableisotope analysis can provide insight in the seasonal variation of riverwater chemistry, and thus in seasonality in that area through time.Thus, (Milankovitch-forced) seasonality variation should be capturedin the amplitude of sclerochronological δ18Oar and δ13Car records of fos-sil riverine shells from the Turkana Basin, which potentially provides uswith the possibility to make proxy-based seasonality reconstructionsfor time intervals in the Quaternary in which key evolutionary eventstook place in the hominin record.
4. Conclusions
Modern molluscan bivalves from Lake Turkana and the Omo Riverrecord a wide range of oxygen and carbon isotope values. Deposition-al environment exerts the primary control on molluscan stable iso-tope variation, with river, delta and lake dwelling shells occupyingdistinctly different zones in a δ13Car–δ18Oar cross plot. Mixing be-tween isotopically contrasting lake and river water is the main mech-anism underlying the environmental control on molluscan stableisotope variation.
Fossil shells from the ~2 Ma Lorenyang lake phase in the TurkanaBasin have stable isotope ratios that compare well with the modernspecimens. The typically low seasonal stable isotope amplitude infully lacustrine shells from Lake Lorenyang is not necessarily theresult of decreased monsoonal strength, but rather the effect of thedampened response of lake chemistry to seasonal environmental var-iation. Riverine shells must be expected to provide the most accuratestable isotope records of climate (rainfall) seasonality in the area.
δ18Oar values of fully lacustrine molluscs from Lake Lorenyang andmodern Holocene Lake Turkana are comparable, while alkalinity of

26 H.B. Vonhof et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 383–384 (2013) 16–26
Lake Lorenyang must have been much lower than that of the modernlake.We argue that this reflects the non-linear response of δ18Ow valuesto ongoing evaporation from already evaporated surface water. Theimplication is that molluscan δ18Oar values of evaporative basins likeTurkana are a relatively poor proxy to quantify climate-controlled evap-oration from lake water.
Acknowledgements
This study was supported by the Netherlands Organisation forScientific Research (NWO). We thank Kamoya Kimeu, NdoloMuthoka, Muthoka Kivingo, Halewijn Scheuerman († 9–12-2009),the Koobi Fora Research Project and the Turkana Basin Institute forlogistical support in the field. We are grateful to Frank Brown andDirk van Damme for providing key shell specimens, and to LouiseLeakey and Ikal Angelei for water sampling at the west side of LakeTurkana. Furthermore, we owe thanks to Wynanda Koot and SuzanVerdegaal for analytical assistance, and to Saskia Kars for makingSEM photographs. We gratefully acknowledge the National Museumsof Kenya and the Government of Kenya for facilitating our research inthe Koobi Fora region.
References
Abell, P.I., 1982. Paleoclimates at Lake Turkana, Kenya, from oxygen isotope ratios ofgastropod shells. Nature 297 (5864), 321–323.
Aronson, J.L., Hailemichael, M., Savin, S.M., 2008. Hominid environments at Hadar frompaleosol studies in a framework of Ethiopian climate change. Journal of HumanEvolution 55 (4), 532–550.
Barton, C.E., Solomon, D.K., Bowmann, J.R., Cerling, T.E., 1987. Chloride budgets in tran-sient; Lakes Baringo, Naivasha, and Turkana. Limnology and Oceanography 23,745–751.
Cerling, T.E., 1979. Paleochemistry of Plio-Pleistocene lake Turkana, Kenya.Palaeogeography, Palaeoclimatology, Palaeoecology 27, 247–285.
Cerling, T.E., 1986. A mass-balance approach to basin sedimentation — constraintson the recent history of the Turkana Basin. Palaeogeography, Palaeoclimatology,Palaeoecology 54 (1–4), 63–86.
Cerling, T.E., Bowman, J.R., Oneil, J.R., 1988. An isotopic study of a fluvial–lacustrinesequence — the Plio-Pleistocene Koobi Fora sequence, East Africa. Palaeogeography,Palaeoclimatology, Palaeoecology 63 (4), 335–356.
Cohen, A.S., 1986. Distribution and faunal associations of benthic invertebrates at LakeTurkana, Kenya. Hydrobiologia 141 (3), 179–197.
deMenocal, P.B., 2004. African climate change and faunal evolution during thePliocene-Pleistocene. Earth and Planetary Science Letters 220 (1–2), 3–24.
Dettman, D.L., Reische, A.K., Lohmann, K.C., 1999. Controls on the stable isotope com-position of seasonal growth bands in aragonitic fresh-water bivalves (Unionidae).Geochimica et Cosmochimica Acta 63 (7–8), 1049–1057.
Feibel, C.S., 1993. Freshwater stingrays from the Plio-Pleistocene of the Turkana Basin,Kenya and Ethiopia. Lethaia 26 (4), 359–366.
Feibel, C.S., Brown, F.H., McDougall, I., 1989. Stratigraphic context of fossil hominidsfrom the Omo group deposits — northern Turkana Basin, Kenya and Ethiopia.American Journal of Physical Anthropology 78 (4), 595–622.
Garcin, Y., Melnick, D., Strecker, M.R., Olago, D., Tiercelin, J.-J., 2012. East African mid-Holocene wet-dry transition recorded in palaeo-shorelines of Lake Turkana, northernKenya Rift. Earth and Planetary Science Letters 331, 322–334.
Grossman, E.L., Ku, T.-L., 1986. Oxygen and carbon isotope fractionation in biogenicaragonite: temperature effects. Chemical Geology 59, 59–74.
Halfman, J.D., Johnson, T.C., Showers, W.J., Lister, G.S., 1989. Authigenic low-Mg calcitein Lake Turkana, Kenya. Journal of African Earth Sciences 8 (2–4), 533–540.
Johnson, T.C., Halfman, J.D., Showers, W.J., 1991. Paleoclimate of the past 4000 years atLake Turkana, Kenya, based on the isotopic composition of authigenic calcite.Palaeogeography, Palaeoclimatology, Palaeoecology 86, 189–198.
Joordens, J.C.A., 2011. The power of place: climate change as driver of hominin evolutionand dispersal over the past 5 million years. (PhD thesis) VU University Amsterdam,Amsterdam (119 pp.).
Joordens, J.C.A., et al., 2011. An astronomically-tuned climate framework for homininsin the Turkana Basin. Earth and Planetary Science Letters 307 (1–2), 1–8.
Kaandorp, R.J.G., et al., 2003. Seasonal stable isotope variations of the modern Amazonianfreshwater bivalve Anodontites trapesialis. Palaeogeography, Palaeoclimatology,Palaeoecology 194 (4), 339–354.
Kaandorp, R.J.G., Wesselingh, F.P., Vonhof, H.B., 2006. Ecological implications fromgeochemical records of Miocene Western Amazonian bivalves. Journal of SouthAmerican Earth Sciences 21 (1–2), 54–74.
Keith, M.L., Anderson, G.M., Eichler, R., 1964. Carbon and oxygen isotopic compositionof mollusk shells from marine and fresh-water environments. Geochimica etCosmochimica Acta 28, 1757–1786.
Kingston, J.D., 2007. Shifting adaptive landscapes: progress and challenges in reconstructingearly hominid environments. American Journal of Physical Anthropology 20–58.
Levin, N.E., Zipser, E.J., Cerling, T.E., 2009. Isotopic composition of waters from Ethiopiaand Kenya: insights into moisture sources for eastern Africa. Journal of GeophysicalResearch—Atmospheres 114.
Liu, W., Li, X., Zhang, L., An, Z., Xu, L., 2009. Evaluation of oxygen isotopes in carbonateas an indicator of lake evolution in arid areas: the modern Qinghai Lake, Qinghai–TibetPlateau. Chemical Geology 268 (1–2), 126–136.
Majoube, M., 1971. Oxygen-18 and deuterium fractionation between water and steam.Journal de Chimie Physique et de Physicochimie Biologique 68 (10), 1423-&.
Maslin, M.A., Trauth, M.H., 2009. Plio-Pleistocene East African pulsed climate variabilityand its influence on early human evolution. In: Grine, F.E., Fleagle, J.G., Leakey, R.E.(Eds.), The First Humans: Origin and Early Evolution of the Genus Homo. VertebratePaleobiology and Paleoanthropology. Springer, pp. 151–158.
Mohammed, M.U., Bonnefille, R., Johnson, T.C., 1996. Pollen and isotopic records in LateHolocene sediments from Lake Turkana, Kenya. Palaeogeography, Palaeoclimatology,Palaeoecology 119 (3–4), 371–383.
Ng'ang'a, P., Muchane, M.W., Johnson, T.C., Sturgeon, K., 1998. Comparison of isotopicrecords in abiogenic and biogenic calcite from Lake Turkana, Kenya. In: Lehman,J.T. (Ed.), Environmental Change and Response in East African Lakes. Springer.
Nicholson, S.E., 1999. Historical and modern fluctuations of Lakes Tanganyika andRukwa and their relationship to rainfall variability. Climatic Change 41 (1), 53–71.
Potts, R., 2007. Environmental hypotheses of Pliocene human evolution. In: Bobe, R.,Alemseged, Z., Behrensmeyer, A.K. (Eds.), Hominin Environments in the East AfricanPliocene: An Assessment of the Faunal Evidence. Springer, pp. 25–49.
Ricketts, R.D., Anderson, R.F., 1997. Stable Isotope Study of Lake Turkana, East Africa,Use of Isotope Techniques in Lake Dynamics Investigations. IAEA, Rehovot, Israel97–103.
Ricketts, R.D., Anderson, R.F., 1998. A direct comparison between the historical recordof lake level and the delta O-18 signal in carbonate sediments from Lake Turkana,Kenya. Limnology and Oceanography 43 (5), 811–822.
Ricketts, R.D., Johnson, T.C., 1996. Climate change in the Turkana basin as deduced froma 4000 year long delta O-18 record. Earth and Planetary Science Letters 142 (1–2),7–17.
Tierney, J.E., Lewis, S.C., Cook, B.I., LeGrande, A.N., Schmidt, G.A., 2011. Model, proxyand isotopic perspectives on the East African Humid Period. Earth and PlanetaryScience Letters 307 (1–2), 103–112.
Trauth, M.H., et al., 2007. High- and low-latitude forcing of Plio-Pleistocene East Africanclimate and human evolution. Journal of Human Evolution 53 (5), 475–486.
Van Bocxlaer, B., van Damme, D., 2009. Palaeobiology and evolution of the Late Cenozoicfreshwatermolluscs of the Turkana Basin: Iridinidae Swainson, 1840 and EtheriidaeDeshayes, 1830 (Bivalvia: Etherioidea). Journal of Systematic Palaeontology 7 (2),129–161.
Van Damme, D., Van Bocxlaer, B., 2009. Freshwater molluscs of the Nile, past and present.In: Dumont, H.J. (Ed.), The Nile: Origin, Environments, Limnology and Human Use.Springer, pp. 585–629.
Versteegh, E.A.A., Vonhof, H.B., Troelstra, S.R., Kaandorp, R.J.G., Kroon,D., 2010a. Seasonallyresolved growth of freshwater bivalves determined by oxygen and carbon isotopeshell chemistry. Geochemistry, Geophysics, Geosystems 11 (8).
Versteegh, E.A.A., Vonhof, H.B., Troelstra, S.R., Kroon, D., 2010b. Amolluscan perspectiveon hydrological cycle dynamics in northwestern Europe. Netherlands Journal ofGeosciences 89 (1), 49–58.
von Grafenstein, U., Erlernkeuser, H., Trimborn, P., 1999. Oxygen and carbon isotopesin modern fresh-water ostracod valves: assessing vital offsets and autecologicaleffects of interest for palaeoclimate studies. Palaeogeography, Palaeoclimatology,Palaeoecology 148 (1–3), 133–152.
Williamson, P.G., 1982. Molluscan biostratigraphy of the Koobi Fora hominid-bearingdeposits. Nature 295, 140–142.
Yuretich, R.F., 1979. Modern sediments and sedimentary processes in Lake Turkana(Lake Rudolf) eastern rift valley, Kenya. Sedimentology 26 (3), 313–331.
Yuretich, R.F., Cerling, T.E., 1983. Hydrochemistry of Lake Turkana, Kenya—mass balanceand mineral reactions in an alkaline lake. Geochimica et Cosmochimica Acta 47 (6),1099–1109.





























