Embed Size (px)
Citation preview

Date: Tue, 30 May 2000 15:31:08 -0400From: OAO <[email protected]>To: [email protected]: WWW ILL Request
[email protected] (OAO) sent the following ILL request:----------------------------------------------------------------Author: Distel, D. L.Book Title: BioscienceJournal Article: "Evolution of chemoautotrophic endosymbioses in bivalves -Bivalve-bacteria chemosymbioses are phylogenetically diverse butmorphologically similar."Year: 1998Volume: 48(4):Pages: 277-286.6Location:Rush: onNeeded By:User Name: OAOE-Mail address: [email protected] #: 30-05-2000-_------------------____________________------------------------
Server Protocol: HTTP/1.1Remote Host: (200.10.149.11)

Evolution of ChemoautotrophicEndosymbioses in Bivalves
Bivalve-bacteria chemosymbioses are phylogenetically diversebut morphologically similar
H ydrothermal vents conjurethe very images that are as-sociated with primal life on
a newly formed Earth: massive stonechimneys rising from a dark sea-floor, plumes of sulfurous smoke bil-lowing from their peaks, and, allaround, strange creatures feeding si-lently in the shimmering geothermalheat. It seems only fitting that thehydrothermal vent environment isposited to be a refuge for speciesderived from primitive and ancientforms (Newman 1985, Tunnicliffe1992). However, one of the mostunusual biological phenomena dis-covered at the vents now appears tohave a long history spent in less ex-otic environments far removed fromhydrothermal vents.
This phenomenon is chemoau-totrophic endosymbiosis, a nutri-tional strategy found in an increas-ing number of marine invertebrates.With the aid of bacterial endosym-bionts, these “chemosymbiotic” ani-mals subsist on nutrients and energysources, including hydrogen sulfideand carbon dioxide, that are nor-mally unavailable to animals. Simi-lar strategies have been observed inat least five animal phyla, includingbivalve and gastropod molluscs andvestimentiferan, pogonophoran, an-nelid, and nematode worms. It isamong the bivalves, however, that
Daniel L. Distel (e-mail: [email protected]) is an assistant professor in the Depart-ment of Biochemistry, Microbiology andMolecular Biology at the University ofMaine, Orono, ME 04469-5735. 0 1998American Institute of Biological Sciences.
April 1998
Daniel L. Distel
The diversity and
antiquity of chemo-symbiotic bivalve and
bacterial lineages suggest
that chemosymbiosis mayhave played an important
role in the early diver-sification of the Bivalvia
this strategy is most widespread, bothphylogenetically and geographically.In this article, I examine where, when,and how these symbioses becameestablished in this important anddiverse invertebrate group.
In 1977, near the Galapagos Is-lands, geologists got their firstglimpse of the deep-sea hot springsknown as hydrothermal vents. Theexistence of these volcanic featureswas anticipated by theories of platetectonics and sea-floor spreading;however, the discovery of lush bio-logical communities surrounding thevents (Lonsdale 1977) was com-pletely unexpected. These diverse anddensely populated communities wereremarkable for a number of reasons.First, they stand in sharp contrast tothe long-held perception of the deep-sea floor as a uniformly spare anddesertlike environment. Second, theyare populated largely by species andhigher taxa that are completely newto science (Newman 1985, Tunni-
cliffe 1992). And third, the basis ofprimary productivity in the vent com-munities is not photosynthesis, as isthe case for all previously knownbiological communities, but bacte-rial chemosynthesis-rather thansunlight, these communities use geo-thermal energy stored in the form ofthermogenic hydrogen sulfide andother reduced inorganic compounds.Even more remarkable, however, wasthe discovery that many of the bacte-ria responsible for this primary pro-duction exist as chemoautotrophicendosymbionts that live intracellu-larly within specialized organs inmany vent-associated invertebrates(Cavanaugh et al. 1981, Felbeck etal. 1981).
Although fundamentally new toscience, chemoautotrophic endosym-biosis is a nutritional strategy thatloosely parallels photosynthesis inhigher plants. In plants, photosyn-thesis occurs in chloroplasts, or-ganelles that are now believed tohave evolved from endosymbioticphotosynthetic bacteria (cyano-bacteria). Both chemoautotrophicendosymbionts and photoauto-trophic chloroplasts use ATP as anenergy source to drive fixation ofcarbon dioxide into carbohydratesvia the Calvin-Benson Cycle. Thesecarbohydrates subsequently serve asthe primary nutritional carbon sourcefor their respective hosts. The two“symbioses” differ, however, in theway in which ATP is generated: Chlo-roplasts use light energy to synthe-size ATP, whereas chemoautotrophicendosymbionts harness chemical en-ergy stored in reduced sulfur com-
277

pounds. Chemosymbiotic inverte-brates are, thus, unique among highereukaryotes; their ability to use hy-drogen sulfide and carbon dioxide asprimary nutrients is as different from“typical” animal heterotrophy as isa plant’s ability to use carbon diox-ide and light.
The unusual nutritional strategyused by these chemosymbiotic ani-mals is reflected in their unusualappearance and morphology. Ventanimals that associate with chemoau-
totrophic symbionts include giantvestimentiferan tube worms (e.g.,Riftia pachyptila), which may growto more than two meters in length,and large clams (e.g., Calyptogenamagnifica) and mussels (e.g., Bathy-modiolus thermophilm), which mayreach nearly one-third of a meter.However, it is not their unusual sizethat is the most conspicuous differ-ence between these animals and theirmore typical heterotrophic counter-parts, but rather the relative sizes of
Figure 1. Sequential views of theanatomy of the symbiont-containinggills of Lucinoma aequizomta (Luci-nidae). I. L. aequizonatu with the leftshell removed to reveal the leftdemibranch of the paired gills (g). ILA plug of tissue removed from theboxed region in I is shown rotated90” about its horizontal axis, expos-ing the transparent filaments (f) andsymbiont-containing subfilamentartissue(s). Note the color differencecaused by white bacterial symbiontsin the subfilamentar tissue. III. A lightmicrograph of a stained thick seaionof tissue from the boxed region in II,showing individual filaments andsubfilamenrar tissue. IV. An electronmicrograph of the boxed region in IIIshows bacterial symbionts (b) withinbacteriocyres in the subfilamentartissue. V. Detail from boxed regionin IV showing two bacterial sym-bionts. VI. Schematic diagram ofsulfur-oxidizingchemoautotrophy inL. aequizonata symbionts, which oc-curs within the symbiont cells shownin the boxed region in V. Oxidationof reduced sulfur compounds (S,)provides energy for production ofATF’, which in turn drives fixation ofCOz into carbohydrate ( CH20), viathe Calvin-Benson cycle (CB). Thiscarbohydrate serves as the primarycarbon and energy source for thehost’s nutrition.
their symbiont-bearing organsand their digestive systems.
The symbiont-bearing organsare typically the largest and mostconspicuous organs in theseanimals, often accounting formore than one - th i rd o f theanimal’s total soft-tissue weight.In Riftia and other vestimenti-ferans, this organ (called thetrophosome) fills most of thebody cavity. Bivalves lack atrophosome; instead, they housetheir symbionts in specializedcells within the subfilamentar
tissue of their gills (Figure 1). Exten-sive development of this subfila-mentar tissue in chemosymbioticbivalves gives these gills a character-istic thickness and opacity that con-trasts sharply with the thin, delicate,translucent gills of most bivalves.
Whereas the symbiont-bearingorgans of chemosymbiotic animalsare enlarged, their adult digestivesystems are either completely absentor are considerably reduced in rela-tive size and complexity by compari-
278 Bioscience Vol. 48 ‘No. 4

son to those of symbiont-free taxa.This observation alone suggests astrong nutritional dependence on thesymbionts. In fact, this nutritionalrelationship has been confirmed formany taxa by extensive physiologi-cal, biochemical, ecological, morpho-logical, isotopic, enzymatic, micro-scopic, and molecular evidence (forreviews, see Felbeck and Somero1982,Felbeck et al. 1983,1985, Cava-naugh 1985, 1994, E. Southward1987, A. Southward 1989, Fisher1990, Felbeck and Distel 1992).
Although first discovered at hy-drothermal vents, chemoautotrophicendosymbioses are not limited tothese highly specialized environ-ments. Similar symbiotic mechanismshave been identified in at least fivecommon and widely distributed bi-valve families that have long beenknown to science: the Mytilidae,Solemyidae, Vesicomyidae, Thyasiri-dae, and Lucinidae (Figure 2). Untilrecently, however, the nutritionalstrategies of these symbiont-contain-ing taxa were either unknown ormisunderstood. For example, thegutless solemyids were proposed todigest food particles externally, bysecreting digestive enzymes into themantle cavity (Owen 1961), or toabsorb dissolved nutrients directlyfrom sea water (Reid 1980, Reid andBernard 1980), and the lucinids werethought to be adapted for suspen-sion feeding on large particles(Bretsky 1976). These adaptationswere proposed to allow lucinids andsolemyids to survive in marginal en-vironments unsuitable for most othermacrofaunal species (Bretsky 1976).
The environments that these taxainhabit are also sites of hydrogensulfide accumulation, even thoughthey are not associated with vents.The sulfides in vent communitiescome from fluids transported to thesea floor by geothermal flow (hydro-thermal vents) or by other hydro-logical processes (cold seeps); how-ever, in many marine environments,sulfide is produced in situ by biologi-cal processes. This biogenic sulfide,produced primarily by bacterial sul-fate reduction in anaerobic sedimentsthat are rich in organic material, canalso support chemosymbiotic com-munities. Such communities havebeen found in mangrove swamps, insea-grass beds, in anoxic marine ba-
April 1998
Figure 2. Chemosvmbiotic bivalves. Shells from 17 species demonstrate the diversity ofchemosymbiotic Bivalves. Shaded boxes in the key indicate members of the famihesVesicomyidae (l), Mytilidae (2), Solemyidae (3), and the superfamily Lucinacea (4).
sins, in sewage outfalls (Felbeck andDistel 1992), in rotting whale car-casses (Smith et al. 1989), and evenin the decaying cargo of a sunkencoffee freighter (Dando et al. 1992).The “marginal” nature (i.e., the lowmacrofaunal diversity) of these envi-ronments is likely explained by thetoxicity of hydrogen sulfide (a po-tent inhibitor of electron transport)and the low oxygen concentrationsassociated with sulfidic sediments(hydrogen sulfide oxidizes sponta-neously, so it accumulates only insediments in which oxygen concen-trations are low). Thus, chemoau-totrophic symbioses allow their hostsnot only to use nutrients normallyunavailable to other eukaryotes, butalso to inhabit environments that,due to the presence of these samecompounds, are inhospitable to mostother animals.
Antiquity and interdependence
Because chemosymbiosis was asso-ciated with hydrothermal vents, itinitially appeared to be an ancientnutritional strategy. Vent communi-ties have been proposed as refugiafor descendants of extremely ancient
taxa (Newman 1985, Tunnicliffe1992) because of their purportedsimilarity to primitive Earth envi-ronments, their extreme endemism,and their relative isolation from cata-clysmic events affecting Earth’s sur-face. Several lines of evidence nowprovide additional support for thisearly impression.
One indication of the antiquity ofthese symbioses is the degree to whichthe hosts and symbionts have be-come adapted to accommodate theirsymbiotic association. As previouslydiscussed, chemosymbiotic hosts dis-play dramatic modifications in theirphysiology and body plans that arespecifically related to their symbio-ses. Whereas individual morphologi-cal or physiological changes mayevolve quickly, integrated and coor-dinated changes in multiple featuresof both hosts and symbionts likelyreflect long-term selection and “finetuning” of traits favoring symbiosisspecifically.
Obligacy-the loss of the capac-ity of the host or symbiont for inde-pendent existence-may also indi-cate an ancient association. In mostbivalve chemosymbioses, the sym-bionts are thought to provide a sig-
279

nificant proportion of the host’s car-bon nutrition (Southward 1987,1989, Cary et al. 1989, Conway etal. 1989, 1993, Fisher 1990). Plau-sible alternative feeding modes oftendo not exist, particularly in the caseof gutless species. Furthermore,aposymbiotic (symbiont-free) adultspecimens of chemosymbiotic spe-cies are not observed in nature. Theseobservations suggest that the sym-bioses are obligate for their hosts.
Whether the bacteria found inthese hosts are obligate symbionts is,however, uncertain. Although sym-bionts have neither been successfullycultivated in vitro nor observed free-living in the environment, it is pos-sible that these bacteria are capableof independent existence. Most bac-teria (particularly endosymbionts)either have not or cannot be grownin pure culture, and cultivation ef-forts typically underrepresent bacte-rial diversity in natural environments(Giovannoni et al. 1995).
rRNAgenes (Distelet al. 1988,1994,Distel and Wood 1992, Eisen et al.1992, Distel and Cavanaugh 1994,Kim et al. 1995, Durand et al. 1996,Krueger and Cavanaugh 1997). Eachbivalve host harbors just onechemoautotrophic phylotype that isfound in all adult specimens, regard-less of location or condition. In somecases, the same symbiont phylotypehas been observed in more than onehost species (Durand et al. 1996).However, this observation does notnecessarily indicate that the sym-bionts are identical or that they maybe freely exchanged between thesehost species. Small subunit rRNAgenes evolve at such low rates(Ochman and Wilson 1987, Moranet al. 1993) that organisms with iden-tical 16s rRNA genes may as easilyhave shared their most recent com-mon ancestor one year or one mil-lion years ago.
Some indication of the potentialability of certain symbionts to liveindependently comes from studies ofsymbiont transfer mechanisms. Al-though the precise mechanisms ofsymbiont transfer are not fully un-derstood in any bivalve system, twodistinct modes of symbiont transferhave been proposed. In members oftwo families (Vesicomyidae andSolemyidae), indirect evidence sug-gests that symbionts may be presentin reproductive tissues, indicatingthat symbionts may be transferredby a “vertical” mechanism-that is,directly from parent to offspringthrough the gametes (Cary andGiovannoni 1993, Cary 1994,Krueger et al. 1996). In this case,there would be no need for sym-bionts to exist apart from their hosts.However, experiments with onelucinid species suggest that its sym-bionts are acquired from the envi-ronment (Gros et al. 1996); if so,these particular symbionts must becapable of free-living existence.
In a few cases, two symbiontphylotypes have been detected withina single clam (Fisher et al. 1993,Distel and Cavanaugh 1994,1995).The second phylotype belongs to aphylogenetically and physiologicallydistinct group of bacteria known asmethanotrophs. These symbiontsoxidize methane as an energy sourceand use methane and probably othersingle-carbon compounds as theirprimary carbon source. These dualsymbioses are unique in that they arethe only animal-bacterial symbiosesknown in which two phylogeneti-tally distinct symbiont phylotypescoexist within a single host cell (Disteland Cavanaugh 1995). Methano-trophic symbioses in bivalves areobserved only in the family Mytilidaeand can exist exclusive of, or in con-junction with, chemoautotrophicsymbioses in different mytilid spe-cies (for reviews, see Fisher 1990,Cavanaugh 1993).
Specificity ofbivalve chemosymbioses
In addition to species specificity,chemosymbiotic associations inbivalves exhibit group specificity-that is, closely related bivalve spe-cies associate with svmbionts thatare also closely related (Distel et al.1994). The occurrence of closely re-lated symbionts in closely related
Regardless of the mode of transmis- hosts suggests that these particularsion, host-symbiont associations are hosts and symbionts occur togetherhighly species specific, as indicated because their most recent commonby comparative sequence analysis of ancestors occurred together in a com-the symbiont’s small subunit (16s) mon ancestral symbiosis-that is,
they are associated by descent(Brooks andMcLennan 1993). How-ever, other explanations for groupspecificy are possible. For example,if symbiotic “infections” are trans-mitted laterally by colonization ofnew host taxa, and if these infectionsare more easily transmitted betweenclosely related hosts than distantones, then the same pattern of host-symbiont relationships might arise.The distinction between these twoscenarios is an important one: If sym-bionts were acquired before the di-vergence of their hosts, symbiosismay have been an important evolu-tionary force driving those diver-gences, but if symbionts were ac-quired after host divergence, thesymbioses may simply represent simi-lar but independent solutions to a com-mon set of environmental challenges.
History by comparison
It may be possible to distinguish be-tween these alternatives by compar-ing host and symbiont phylogeny. Ifsymbionts exist primarily within theirhosts and are transmitted from gen-eration to generation with extremelyhigh specificity and fidelity, thenspecies divergences among symbiontsmight eventually be expected to mir-ror those observed among their hosts.In this case, phylogenetic trees inde-pendently inferred for hosts and theirrespective symbionts should showsimilar patterns of branch order andbranch length. When observed, suchcongruence is evidence of associa-tion by descent and can be used togauge the antiquity of the associa-tions using the fossil record of thehosts for chronological calibration(Moran and Telang 1998 ). However,any congruent phylogenetic signalscould easily be obscured by sortingevents, such as symbiont transfersbetween host species or de novo es-tablishment of symbiotic associa-tions, even if such events are exceed-ingly rare. Thus, although absolutecongruence between host and sym-biont trees has a simple interpreta-tion, partial congruence may not.
The reason for this difficulty ininterpretation becomes clear whenone considers the problems inherentin assessing the evolution of a sym-biosis as compared with that of asingle organismic lineage. In exam-
280 BioScience Vol. 48 No. 4
ining a symbiosis, at least three his-tories need to be considered: the his-tory of the hosts, the history of thesymbionts, and the history of theassociation itself. Knowing the his-tory of the symbionts, for example,does not necessarily reveal the his-tory of the symbiosis because hostsmay gain or lose symbionts and sym-bionts may switch hosts. Suchreassortment of partners could re-sult in closely related hosts with phy-logenetically, or even functionally,unrelated symbionts-that is, host,symbiont, and symbioses may havedistinctly different histories. Unlessthe phylogenetic relationships amonghosts and those among symbionts arevery nearly parallel, the history of thesymbiosis becomes difficult to resolve.
Comparison of host andsymbiont phylogenies
Although the phylogeny of chemoau-totrophic symbionts has been exten-sively explored using 16s rRNAanalyses, similarly comprehensivemolecular analyses of host genes arenot yet available. However, com-parison of the molecular phylogenyof the svmbionts and the classicalsystematics of the hosts provides in-sights into the origins and history ofthese symbioses (Distel et al. 1994).
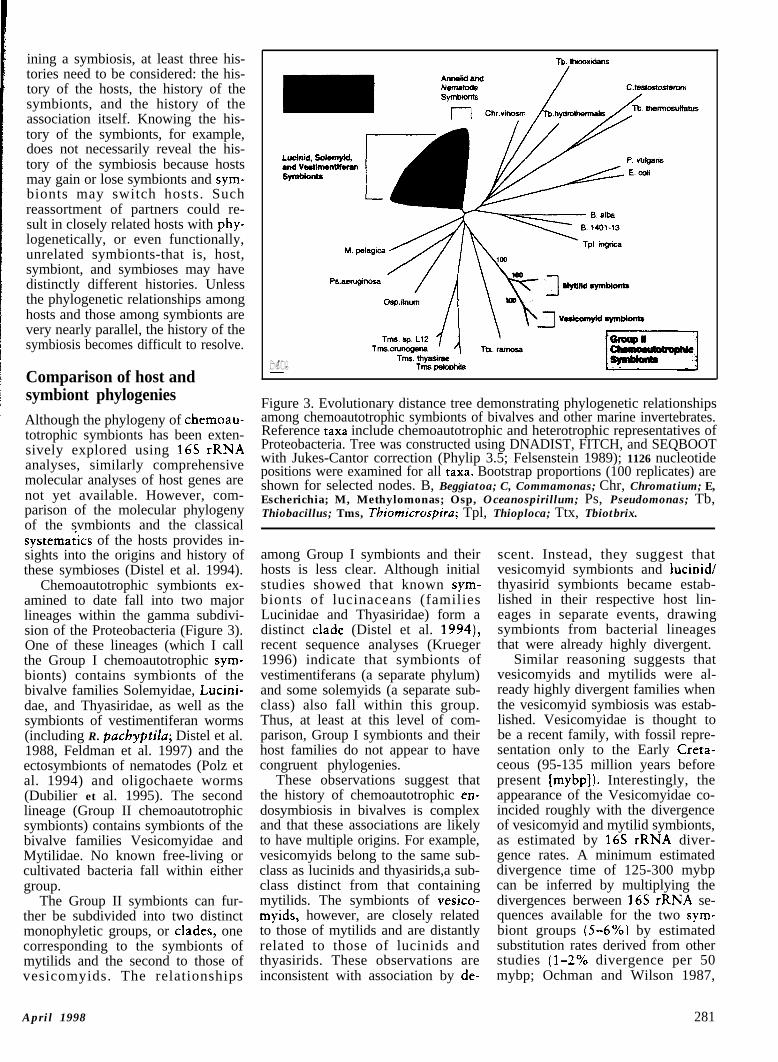
Chemoautotrophic symbionts ex-amined to date fall into two majorlineages within the gamma subdivi-sion of the Proteobacteria (Figure 3).One of these lineages (which I callthe Group I chemoautotrophic sym-bionts) contains symbionts of thebivalve families Solemyidae, Lucini-dae, and Thyasiridae, as well as thesymbionts of vestimentiferan worms(including R. pachyptila; Distel et al.1988, Feldman et al. 1997) and theectosymbionts of nematodes (Polz etal. 1994) and oligochaete worms(Dubilier et al. 1995). The secondlineage (Group II chemoautotrophicsymbionts) contains symbionts of thebivalve families Vesicomyidae andMytilidae. No known free-living orcultivated bacteria fall within eithergroup.
The Group II symbionts can fur-ther be subdivided into two distinctmonophyletic groups, or clades, onecorresponding to the symbionts ofmytilids and the second to those ofvesicomyids. The relationships
April 1998
l-B. thmoxdans
Ann&ii andN.9fll.WZ&
Lucbtid. Sobmybd,
1.0% Tmspebphila-
Figure 3. Evolutionary distance tree demonstrating phylogenetic relationshipsamong chemoautotrophic symbionts of bivalves and other marine invertebrates.Reference taxa include chemoautotrophic and heterotrophic representatives ofProteobacteria. Tree was constructed using DNADIST, FITCH, and SEQBOOTwith Jukes-Cantor correction (Phylip 3.5; Felsenstein 1989); 1126 nucleotidepositions were examined for all taxa. Bootstrap proportions (100 replicates) areshown for selected nodes. B, Beggiatoa; C, Commamonas; Chr, Chromatium; E,Escherichia; M, Methylomonas; Osp, Oceanospirillum; Ps, Pseudomonas; Tb,Thiobacillus; Tms, Tbiomicrospiru; Tpl, Thioploca; Ttx, Tbiotbrix.
among Group I symbionts and theirhosts is less clear. Although initialstudies showed that known sym-bionts of lucinaceans (familiesLucinidae and Thyasiridae) form adistinct clade (Distel et al. 1994),recent sequence analyses (Krueger1996) indicate that symbionts ofvestimentiferans (a separate phylum)and some solemyids (a separate sub-class) also fall within this group.Thus, at least at this level of com-parison, Group I symbionts and theirhost families do not appear to havecongruent phylogenies.
These observations suggest thatthe history of chemoautotrophic en-dosymbiosis in bivalves is complexand that these associations are likelyto have multiple origins. For example,vesicomyids belong to the same sub-class as lucinids and thyasirids,a sub-class distinct from that containingmytilids. The symbionts of vesico-myids, however, are closely relatedto those of mytilids and are distantlyrelated to those of lucinids andthyasirids. These observations areinconsistent with association by de-
scent. Instead, they suggest thatvesicomyid symbionts and lucinid/thyasirid symbionts became estab-lished in their respective host lin-eages in separate events, drawingsymbionts from bacterial lineagesthat were already highly divergent.
Similar reasoning suggests thatvesicomyids and mytilids were al-ready highly divergent families whenthe vesicomyid symbiosis was estab-lished. Vesicomyidae is thought tobe a recent family, with fossil repre-sentation only to the Early Creta-ceous (95-135 million years beforepresent [mybp]). Interestingly, theappearance of the Vesicomyidae co-incided roughly with the divergenceof vesicomyid and mytilid symbionts,as estimated by 16s rRNA diver-gence rates. A minimum estimateddivergence time of 125-300 mybpcan be inferred by multiplying thedivergences berween 16s rRNA se-quences available for the two sym-biont groups (5-6%) by estimatedsubstitution rates derived from otherstudies (l-2% divergence per 50mybp; Ochman and Wilson 1987,
281

LMoran et al. 1993). Could the diver-gence of vesicomyid and mytilid sym-bionts reflect the establishment ofseparate and exclusive associationswith Mytilidae and the newly emer-gent Vesicomyidae? If so, did sym-biosis arise independently in thesetwo families? Or was symbiosis firstestablished in one family (perhaps inthe more ancient modioloid mytilids)and then transferred to the other?The resolution of these and otherquestions awaits further phylogeneticanalyses.
Diversity of chemoauto-trophic symbionts
Why so many diverse hosts are asso-ciated with just a few closely relatedlineages of chemoautotrophic bacte-ria is not clear. Sulfur-based chemo-autotrophy is found in an extraordi-narily diverse variety of bacteriallineages, both within the Proteo-bacteria and in other prokaryoticphyla. There is no a priori reason toassume that the physiology of theseother bacterial lineages is fundamen-tally incompatible with chemoau-totrophic endosymbiosis. Thus, theparticular success of Groups I and IIin forming endosymbiotic associa-tions remains unexplained.
Although the reason for their suc-cess as chemosymbionts is unknown,the fact that members of Groups Iand II have become established asendosymbionts in so many diversehosts indicates that specialization forassociation with marine invertebrateshas been important to their diver-gence from other bacteria1 taxa, muchas the specialization for associationwith a broad range of vertebratehosts has been an important featurein the evolution of the enteric bacte-ria. The observation that all knownmembers of Groups I and II are sym-biotic suggests that this specializa-tion became established early in theirdiversification. Like the enterics,Groups I and II may yet be found tocontain species that are not symbi-otic. Nevertheless, the importanceof symbiosis in their evolutionaryhistory is clear.
If symbiotic association is ances-tral in Group I chemoautotrophicsymbionts, then the diversity of thisgroup indicates that this specializa-tion is extremely ancient-a finding
that agrees well with paleontologi-cal data (discussed later). Group Isymbionts are highly diverse and areassociated with ancient host taxa,whereas the symbionts of Group IIare less diverse and are associatedwith correspondingly more recenthost lineages (Distel et al. 1994).This finding suggests that symbiosesinvolving Group II arose separatelyand perhaps more recently than as-sociations involving Group I.
Diversity ofchemosymbiotic hosts
The phylogenetic diversity of chemo-symbiotic bivalves also sheds lighton the antiquity and origins’ ofchemoautotrophic symbioses. Al-though relatively few bivalve fami-lies host such symbioses, these fami-lies are phylogenetically diverse andare distributed among the most dis-tantly related bivalve taxa. Chemo-symbiotic lineages have been foundin three of the six subclasses ofbivalvia (Newell 1969) and are rep-resented among protobranch, fili-branch, and eulamellibranch gilltypes. Paleontological evidence sug-gests that these three subclasses di-verged early in the evolution of theBivalvia.
Although chemosymbiosis occursin just a few bivalve families, itspervasiveness within these familiessuggests that it is evolutionarily im-portant. In four of the five bivalvefamilies that contain chemosymbioticmembers, symbionts are present inmost, if not all, known species. In thefifth family (the Mytilidae), only ahandful of the more than 2.50 namedspecies associate with chemoau-totrophic symbionts (Turner 1985).These symbiont-containing speciescomprise a single lineage, which wasdesignated as a new subfamily(Bathymodiolinae; Kenk and Wilson1985). Analyses of 18s rRNA se-quences (Daniel L. Distel; Ellen Rice-Kenchington, Department of Fisher-ies and Oceans, Canada; Eli Chuang,and Colleen M. Cavanaugh, Depart-ment of Organismic and Evolution-ary Biology, Harvard University; un-published data) support subfamilystatus for the Bathymodiolinae, alineage that, to date, has been foundto contain no symbiont-free species.These observations strongly support
the contention that the establishmentof symbiosis may have been highlyinfluential in the emergence and sub-sequent diversification of higher bi-valve taxa (Reid and Brand 1986).
Similarities amongbivalve chemosymbioses
Despite their considerable phyloge-netic diversity, bivalve chemosym-bioses exibit remarkable similarities.In all cases, symbionts are foundwithin the gills, in tissues that areessentially identical in anatomicallocation and developmental origin.The symbionts of protobranchs,filibranchs, and eulamellibranchs arecontained in the intralamellar junc-tions (Figure 1). In addition, eulamel-libranchs also contain symbionts inthe interfilamentar junctions. Thesesubfilamentar tissues are far moreextensively developed than the ho-mologous tissues in symbiont-freebivalves. In fact, the symbiont-con-taining subfilamentar tissue typicallycomprises the bulk of the mass andvolume of the gill.
Many similarities are also seen inthe cellular architecture of the sym-biont-bearing tissues. The symbiont-containing tissues are typically com-posed of two sheets of simpleepithelium surrounding a very nar-row central blood sinus. Each epi-thelium is primarily composed oftwo cell types in approximately equalproportion. One is the symbiont-containing bacteriocytes, which areroughly cuboidal and which containbasal nuclei and apical symbiont-containing vesicles. The other is thetrumpet-shaped, symbiont-free inter-calary cells, which expand from anarrow base to a broad, often sheet-like apex that may extend to covermuch of the external surface of theneighboring bacteriocytes. Both celltypes may contain numerous mi-crovilli but lack cilia and flagella.Large, conspicuous bodies contain-ing numerous concentric membranewhorls are nearly universally ob-served within bacteriocytes, often inclose association with the nuclei.These structures, which are thoughtto be lysosomal residual bodies, fre-quently contain objects resemblingpartially lysed symbiotic bacteria.
The remarkable contrast of mor-phological similarity and phyloge-
2x2 Bioscience Vol. 48 No. 1

netic diversity in bivalve-bacteriachemosymbioses raises an interest-ing evolutionary question: Are thesimilarities due to convergence? Ordo the similarities indicate that thepresence of gill-borne symbionts isan ancestral condition retained injust a few modern lineages? It can beargued that subfilamentar gill tissueis particularly well suited for provid-ing symbionts with exposure to boththe host’s blood supply and to dis-solved nutrients in seawater, so it isa natural site for symbioses to formconvergently. However, other tissues,such as the mantle epithelium or thelabial palps, might serve this func-tion equally well. Indeed, endosym-bionts have recently been observedin the mantle and foot of one mytilidspecies (Streams et al. 1997). Even inthis species, however, the gills re-main the primary symbionr-bearingorgan. If, on the other hand, housingbacterial endosymbionts is an ances-
5 tral function of the subfilamentartissues of these diverse chemo-symbiotic bivalves, the common sym-biont-bearing ancestor would haveto be extraordinarily ancient. To pre-date the divergence of protobranchs,eulamellibranchs, and filibranchs,this hypothetical ancestral hosr couldhave arisen no later than the Ordovi-cian and would be counted amongthe oldest bivalve ancestors.
The fossil recordSome chemosymbiotic bivalve lin-eages are, in fact, extremely ancient(Figure 4). Lucinaceans are proposedto be the direct descendants of theBabinkacea (McAlester 1966), anextinct superfamily that dates to theLower Ordovician and contains someof the oldest undisputed bivalve fos-sils (Cox 1969). Families withchemosymbiotic descendants are alsodisproportionately well representedamong early bivalve fossils. Six re-cent bivalve families are representedby fossils in Silurian deposits (Kriz1984); two of these (Lucinidae andSolemyidae) host chemoautotrophicsymbionts in all extant species. Al-though Thyasiridae does not appearuntil the Cretaceous, this family isthought to have arisen directly fromwithin the Lucinidae (McAlester 1966,Kauffman 1969). Mytilidae appearsin the Devonian, but mytilid taxa
April 1998
Figure 4. First appear-ances in the fossilrecord of bivalve taxawith extant chemosym-biotic descendants.
System/Series
unequivocally asso-ciated with chemo-symbiotic commu-nities can be tracedonly to the Late Ju-rassic (Campbelland Bottjer 1993).Finally, Vesicomyi-dae is the most re-cent family, makingits first known ap-pearance in Creta-ceous rocks (Kanieet al. 1993). Thus,t h e m o s t r e c e n tfamilies with exclu-sively chemosym-biotic descendantsappeared more than65 million years ago,and the most ancientmay approach half abillion years in age!
Paleocene
Jurassic
DevonianUnfortunately,
Ordovician
whether the ances-tors of these mod-ern symbiont-bear-ing bivalve lineagesthemselves hostedchemoautotrophicsymbioses is notknown. Bacterial en-dosymbionts rarely,if ever, leave a traceof their existence inthe fossil record. Ina few cases, how-ever, isotopic data, based on deple-tion of the heavy isotope (13C) inchemoautotrophically fixed carbon,has been used to suggest thatchemosymbiosis was in place in somefossil bivalves. In these experiments,examination of carbon isotope ra-tios in the protein matrix preservedin fossilized shells provided evidencethat chemosymbioses existed inlucinid bivalves up to approximately120,000 years old (CoBabe 1991).However, poor preservation of theorganic matrix has, so far, preventedthe successful use of this techniquewith older fossils.
known members of the familiesLucinidae, Solemyidae, and Vesico-myidae has often been interpreted asevidence that symbioses are ances-tral in these families. However, it isalso possible that symbioses becameestablished later in the evolution ofthese families and spread laterallywithin, and perhaps among, them.Such pervasive transmission, if itwere combined with the extinctionof nonsymbiotic taxa, could explainthe universal occurrence of symbio-sis in these families.
Despite the lack of fossil data, theobservation that chemoautotrophicsymbionts have been found in all
Even though symbionts themselvesleave no fossil record, paleontologi-cal evidence does support the antiq-uity of these symbioses. Extantchemosymbiotic organisms and those
First Appearance
+ Vesicomyidae
Chemosymbiotic
a =mytilids?Bivalvesat vents
* Mytilidae
+ Solemyidae
q.- Lucinidae
+- Babinkacea
+ First bivalves?
283

found in fossil deposits show similar earliest appearance of these taxa inmorphological adaptations, life po- the fossil record.sitions, and characteristics of sur-rounding sediments. These similari-ties have been interpreted as evidence
Vents, seeps, and the origins of
that ancestral taxa were also adaptedbivalve chemosymbiosis
to chemosymbiosis (Seilacher 1990,Liljedahll991, Campbell and Bottjer1993). For example, solemyids,lucinids, and thyasirids form uniqueburrows whose features are specifi-cally related to their simultaneousneed to extract sulfide from deep inanaerobic sediments while obtain-ing oxygen from the overlying seawater. Studies of trace fossils and lifepositions indicate that these sameadaptations were present in ancientspecies (Bromley 1996).
Evidence supporting the antiquityof chemosymbiosis can also be foundin the fauna1 composition of manyfossil bivalve communities. Low in-fauna1 species diversity and the con-sistent co-occurrence and predomi-nance of lucinid, thyasirid, solemyid,mytilid, and vesicomyid clams arehighly characteristic of modernchemosymbiotic communities. Thesame features are also apparent infossil communities (Bretsky 1976,Hickman 1984, Campbell and Bottjer1993). For example, lucinids, thya-sirids, vesicomyids, and mytilidscharacteristically dominate modernchemosymbiotic communities asso-ciated with decaying whale carcasses.Similar species composition has beendemonstrated in association withfossil whale bones from the Oligocene(Goedert et al. 1995). Lucinids,solemyids, vesicomyids, and mytilidfossils dominate the Miocene “calcaria Lucina” deposits, which are scat-tered across the northern ItalianApennines; these sites are now inter-preted as chemosymbiotic cold-seepcommunities (Taviani 1994). LateEocene deposits in northern Wash-ington contain vesicomyids, mytilids,and thyasirids (Campbell and Bottjer1993). Lucinid, thyasirid, mytilid,and solemyid fossils co-occur in LateJurassic cold-seep communities inCalifornia (Campbell and Bottjer1993), and lucinids and solemyidsco-occur in Silurian deposits ofGotland (Liljedahl 1991). These ob-servations indicate that features ofcommunities and species interpretedas adaptations to chemosymbiosesin modern taxa can be traced to the
Finally, the question remains as tothe role of hydrothermal vents andcold seeps in the evolution ofchemosymbiosis in bivalves. The old-est vent-associated macrofossil as-semblage can be traced to the Sil-urian Yaman Kasy deposits in thesouthern Urals of Russia (Little et al.1997). Although bivalves and tube-worms dominate many modern ventcommunities, the most ancient fossilvent communities are dominated bybrachiopods, tubeworms, and mono-placophorans. In fact, in a survey of37 fossiliferous sites interpreted torepresent phanerozoic hydrothermalor cold vents, bivalves do not maketheir first appearance until the LateJurassic and do not become a pre-dominant component of the ventfauna until the Cretaceous (Campbelland Bottjer 1995). Therefore, themost ancient bivalve chemosym-bioses (those hosted by lucinids,thyasirids, and solemyids-that is,those involving Group I symbionts)may have been in existence for hun-dreds of millions of years before theappearance of bivalves at hydrother-mal vents and cold seeps.
The relationship of mytilid andvesicomyid chemosymbioses (i.e.,those involving Group II symbionts)to the vent environment is moreambiguous. These families have beenobserved primarily at vent and seepsites and have been associated withthese environments since their earli-est known fossil appearances. How-ever, both are also known in organi-cally enriched environments awayfrom vents and seeps, including de-caying whale carcasses (whale-falls)and sunken wood. Fossils associatedwith these environments includemytilids from fossil whale-fall com-munities and fossil wood, andvesicomyids from turbidity flow de-posits in the Oligocene Makah For-mation of western Washington(Goedert and Squires 1993, Goedertet al. 1995). Thus, it is not clearwhether chemosymbiosis in vesico-myids and mytilids evolved at ventsand seeps or whether these chemo-symbiotic taxa were simply highly
successful invaders of these environ-ments.
Conclusions
Much remains to be learned aboutthe history of chemosymbiosis inbivalves, but given the present stateof knowledge several things seemclear. First, it appears likely thatchemosymbioses in bivalves are ex-tremely ancient and extremely suc-cessful, allowing some bivalve fami-lies to span nearly half a billion yearswith apparently little change in lifehabits or appearance. Second, it ap-pears likely that chemoautotrophicendosymbiotic associations existingtoday have multiple origins, that atleast two similar but distinct bacte-rial lineages are involved, and thathost-switching events are likely tohave occurred throughout their his-tory. Third, although bivalve chemo-symbioses were first discovered athydrothermal vents and seeps, it nowappears unlikely that these symbio-ses orignated in these environments.Finally, the extraordinary diversityand proposed antiquity of chemo-symbiotic bivalve and bacterial lin-eages suggest that chemosymbiosismay have played an important role inthe early diversification of the Bivalvia.Continued molecular, physiological,and paleontological explorationspromise to reveal much about theremarkable history of this intriguingand important phenomenon.
Acknowledgments
I thank M. Polz, S. J. Roberts, R.Carnegie, B. Achorn, W. Morrill,and D. D. Pittman for helpful com-ments and discussion on earlierdrafts. Unpublished research cited inthis work was funded in part by NSFgrant no. DEB 9420051.
References citedBretsky SS. 1976. Evolution and classifica-
tion of the Lucinidae (Moilusca: Bivalvia).Palaeontographica Americana 8: 219-337.
Bromley RG. 1996. Bivalve chemosymbionts.Pages 75-78 in Trace Fossils. London:Chapman & Hall.
Brooks D, McLennan D. 1993. Macroevolu-rionary patterns of morphological diver-sificatton among pa ras i t i c f l a tworms(platyhelminthes: cercomeria). Evolution47: 495-509.
Campbell KA, Bottjer DJ. 1993. Fossil cold
284 Bidcience Vol. 48 No. 4

seeps. National Geographic Research andExploration 9: 326-343.
-. 1995. Brachiopods and chemosym-biotic bivalves in Phanerozoic hydrother-mal vent and cold seep environments.Geolog! 23: 321-324.
Cary SC. 1994. Vertical transmission of ac h e m o a u t o t r o p h i c s y m b i o n t i n t h ephotobranch bivalve, Solemya reidi. Mo-lecular Biology and Biotechnology 3: 12 l-130.
Cary SC, Giovannoni SJ. 1993. Transovarialinheritance of endosymbiotic bacteria inclams inhabiting deep-sea hydrothermalvents and cold seeps. Proceedings of theNational Academy of Sciences of theUnited States of America 90: 5695-5699.
Cary SC, Vetter RD, Felbeck H. 1989. Habi-tat characterization and nutritional strat-egies of the endosymbiont-bearing bivalveLucinoma aequizonata. Marine EcologyProgress Series 55: 31-45.
Cavanaugh CM. 1985. Symbiosis of chemo-lithotrophic bacteria and marine inverte-brates from hydrothermal vents and re-ducing sediments. Bulletin of the BiologicalSociety of Washington 6: 373-388.
-. 1993. Methanotroph-inver tebratesymhioses in the marine environment: UI-trastructural, biochemical, and molecularstudies. Pages 315-328 in Murrell JC,Kelly DP, eds. Microbial Growth on C,Compounds. Andover (UK): Intercept.
-. 1994. Microbial symbiosis: Patternsof diversity in the marine environment.American Zoologist 34: 79-89.
Cavanaugh CM, Gardiner SL, Jones ML,Jannasch HW, Waterbury JB. 1981. Pro-caryotic cells in the hydrothermal venttube worm. Science 2 13: 340-342.
CoBabe EA. 1991. Lucinid bivalve evolutionand the detection of chemosymbiosis inthe fossil record. Ph.D. dissertation. De-partment of Organismic and Evolution-ary Biology, Harvard University, Cam-bridge, MA.
Conway N, McDowell-Capuzzo J, Fry B.1989. The role of endosymbiotic bacteriain the nutrition of Solemya :wlum: Evi-dence from stable isotope analysis of en-dosymbionts and host. Limnology andOceanography 34: 249-255.
Conway NM, Kennicutt C, Van Dover CL.1993. Stable isotopes in the study of ma-rine chemosynthesis-based ecosystems.Paees 158-186 in Laitha 1. Michener R.ed: Stable Isotopes in’EcoIogy. New York:Blackwell.
Part N, Vol. 1, Mollusca 6, Bivalvia. PagesN2-N129 in Moore RC. ed. Treatise onInvertebrate Paleontology. Lawrence (KS):The Geological Society of America andthe University of Kansas Press.
Cox LR. 1969. General features of bivalvia:
Dando PR, Southward AJ, Southward EC.1992. Shipwrecked tube worms. Nature356: 667.
Distel DL, Cavanaugh CM. 1994. Indepen-dent phylogenetic origins of methano-trophic and chemoautotrophic bacterialendosymhioses in marine bivalves. Jour-nal of Bacterioloev 176: 1932-1938.
-. 1995. IntraYc’eIlular coexistence ofmethano- and thioautotrophic bacteria ina hydrothermal vent mussel. Proceedingsof the National Academy of Sciences of
the United States of America 92: 9598-9602.
Distel DL, Wood AP. 1992. Characterizationof the gill symbiont Tbyusira flexuosa(Thiasiridae: Bivalvia) by use of poly-merase chain reaction and 16s rRNA se-quence analysis. Journal of Bacteriology174: 6317-6320.
Distel DL, Lane DJ, Olsen GJ, GiovannoniSJ, Pace B, Pace NR, Stahl DA, Felbeck H.1988. Sulfur-oxidizing bacterial endosym-bionts: Analysis of phylogeny and speci-ficity by 16s rRNA sequences. Journal ofBacteriology 170: 2506-2510.
Distel DL, Felbeck H, Cavanaugh CM. 1994.Evidence for phylogenetic congruenceamong sulfur-oxidizing chemoautotrophicbacterial endosymbionts and their bivalvehosts. Journal of Molecular Evolution 38:533-542.
Dubilier N, Giere 0, Distel DL, CavanaughCM. 1995. Characterization of chemoau-totrophic bacterial symbionts in a gutlessmarine worm (Oligochaeta: Annelida) byphylogenetic 165 rRNA sequence analysisand in situ hybridization. Applied andEnvironmental Microbiology 61: 2346-2350.
Durand P, Gros 0 . Frenkiel L, Prieur D.1996. Phylogenetic characterization ofsulfur-oxidizing bacterial endosymbiontsin three tropical Lucinidae by 165 rDNAsequence analysis. Molecular Marine BI-ology and Biotechnology 5: 37-42.
Eisen JA, Smith SW, Cavanaugh CM. 1992.Phylogenetic relationships of chemoau-totrophic bacterial symbionts of Solemyrruelum Say (Mollusca: Bivalvia) determinedby 165 rRNA gene sequence analysis. Jour-nal of Bacteriology 174: 3416-3421.
Felbeck H, Distel DL. 1992. Prokaryotic sym-bionts in marine inver tebrates . Pages3 8 9 1 - 3 9 0 6 i n Ballows A , T r u p e r H ,Harder W. Schleifer KH. eds. The Prokarv-,otes. Berlin: Springer-Veriag.
FeIbeck H, Somero G. 1982. Primary produc-tion in deep-sea hydrothermal vent organ-isms: Roles of sulfide-oxidizing bacteria.Trends in Biochemical Sciences 7: 201-204.
Felbeck H, Childress JJ, Somero GN. 1981.Calvin-Benson cycle and sulfide oxida-tion enzymes in animals from sulfide-richhabitats. Nature 293: 291-293.
Felbeck H, Powell MA, Hand SC, SomeroGN. 1985. Metabolic adantations of hv-drothermal vent animals. Bulletin of TheBiological Society of Washington No. 0:261-272 .
. 1 9 8 3 . B i o c h e m i c a l i n t e r a c t i o n s b e -tween molluscs and their algal and bacte-rial symbionts. Pages 331-358 in Hocha-chak PW, ed. The Mollusca. New York:Academic Press.
Feldman RA, Black MB, Gary SC, Lutz RA,Vriienhoek RC. 1997. Molecular ohvIo-genetics of bacterial endosymbion& gridtheir vestimentiferan hosts. MolecularMarine Biology and Biotechnology 6: 268-_ - -L//.
Felsenstein J. 1989. Phylip-phylogeny infer-ence nackaee. Cladistics 5: 164-166.
F i she r CR. 1990. Chemoau to t roph ic andmethanotrophic symbioses in marine in-vertebrates. Reviews in Aquatic Sciences2: 399-436.
Fisher CR, Brooks JM, Vodenicher JS, ZandeJM, Childress JJ. Burke RA, Jr. 1993. Theco-occurrence of methanotrophic andchemoautotrophic sulfur-oxidizing bac-terial symbionts in a deep-sea mussel. P. S.Z. N. I. Marine Ecoloev 14: 277-289.
Giovannoni SJ, Mullins TE, Field KG. 1995.Microbial diversity in oceanic systems:rRNA a p p r o a c h e s t o t h e s t u d y o funculturable microbes. Pages 2 17-248 inJoint 1, ed. Molecular Ecology of AquaticMicrobes. New York: Sorineer-Verlae.
Goedert JL, Squires RL. 199’3. F&t Oligoc&erecords of Calyptogena (Bivalvia: Vesico-myidae). Veliger 36: 72-77.
Goedert JL, Squires RL, Barnes LG. 1995.Paleoecology of whale-fall habitats fromdeep-water Oligocene rocks, Olympic Pen-insu la , Wash ing ton s t a t e . Palaeogeo-graphy, Palaeoclimatology, Palaeoecology118: 151-158.
Gros 0, Darrasse A, Frenkiel L, Moeza M.1996. Environmental transmission of asulfur-oxidizing bacterial gill endosym-bion t in the t rop ica l luc in id b iva lveCod&a orbicularis. Applied and Envi-ronmental Microbiology 62: 2324-2330.
Hickman CS. 1984. Composition, structure,ecology, and evolution of six Cenozoicdeep-water mollusk communities. Jour-nal of Paleontology 58: 1215-1234.
Kanie Y, Yoshikawa Y, Sakai T, TakahashiT. 1993. The Cretaceous chemosyntheticcold water-dependent molluscan commu-nity discovered from Mikasa City, centralHokkaido. Scientific Reports of the Yoko-suka City Museum 41: 31-36.
Kauffman EG. 1969. Bivalvia-form, func-tion and evolution. Part N, Vol. 1, Mol-lusca 6. Bivalvia. Pages Nl29-N205 inMoore RC, ed. Treatise on InvertebratePaleontology. Lawrence (KS): The Geo-logical Society of America and the Univer-sity of Kansas Press.
Kenk VC, Wilson BR. 1985. A new mussel(Bivalvia: Mytilidae) from hydrothermalvents in the Galapagos Rift zone. Mala-cologia 26: 253-271.
Kim YW, Yasuda M, Yamagashi A, OshimaT, Ohta S. 1995. Characterization of theendosymbiont of the deep-sea bivalve,Calyptogena soyoae. Applied and Envi-ronmental Microbiology 61: 823-827.
Kriz J. 1984. Autecology and ecogeny ofSilurian Bivalvia. Special Papers in Pale-ontology 32: 183-195.
Krueger DM. 1996. Phylogenetic analysis ofthe genus Solemyn (Bivalvia: Protobranch-ia): Implicat ions for the evolut ion ofchemoautotropic symbioses. Ph.D. dis-sertation. Department of Organismic andEvolutionary Biology, Harvard University,Cambridge, MA.
Krueger DM, Cavanaugh CM. 1997. Phylo-genetic diversity of bacterial symbionts ofSolemya hosts based on comparativeanalysis of 16s rRNA genes. Applied andEnvironmental Microbiology 63: 91-98.
Krueger DM, Gustafson RG, Cavanaugh CM.1996. Vertical transmission of chemoau-totrophic symbionts in the bivalve Solemyauelum (Bivalvia: Protobranchia). Biologi-cal Bulletin 190: 195-202.
Liljedahl L. 1991. Contrasting feeding strar-egies in bivalves from the Silurian ofGotland. Paleontology 34: 219-235.
April 1998 285

Support America’scolleges. Because collegeis more than a placewhere young people arepreparing for their future.It’s where America is pre-paring for ifs future.
Giito
m
*wed.
0 .--.?+.a -mmn-- aa
8H6 Buy recyc eId .
It wouId meanthe world to them.
Recycling keeps working to protect
their fuUture wh b den you uy pro ucts
made from recycled materials. For
a free brochure, write Buy RecycIed,EntironmentaI Defense Fund, 257
Park Avenue South, New York, NY
10010 , or call I-800~CALL-EDF.
Newman WA. 1985. The abyssal hydrother-mal vent invertebrate fauna: A glimpse ofantiquity? Bulletin of the Biological Soci-ety of Washington 6: 231-242.
Ochman H, Wilson ,4C. 1987. Evolution inbacteria: Evidence for a universal substi-tution rate in cellular genomes. Journal of.Molecular Evolution 26: 74-86.
/Owen G. 1961. A note on the habits andnutrition of Solemya parkinsoni (Proto-branchia: Bivalvia). Quarterly Journal ofMicroscopical Science 102: 15-21.
Polz MF, Distel DL, Zarda B, Amann R,Felbeck H, Ott ]A, Cavanaugh CM. 1994.Phylogenetic analysis of a highly specificassociation between ectosymbiotic, sul-
Little CTS, Herrington RJ, Maslennikov W,.Morris NJ, Zaykov V. 1997. Silurian hy-drothermal vent community f rom thesouthern Urals, Russia. Science 385: 146-148.
Lonsdale PF. 1977. Clustering of suspension-feeding macrobenthos near abyssal hy-drothermal vents at oceanic spreadingcenters. Deep-Sea Research 24: 857-863.
McAlester AL. 1966. Systematics, affinitiesand life habits of Babinka, a transitionalOrdovician lucinoid bivalve. Paleontol-ogy 8: 231-246.
Moran NA, Telang A. 1998. Bacteriocyte-associated symbionts of insects. BioScience48: 295-304.
M o r a n N A , M u n s o n M A , B a u m a n n P ,Ishikawa H. 1993. A molecular clock inendosymbiotic bacteria is calibrated us-ing insect hosts. Proceedings of the RoyalSociety of London B Biological Sciences253: 167-171.
Newell ND. 1969. Classification of bivajyia.Part N, Volume 1, Mollusca 6, Bivalvia.Pages 205-224 in Moore RC, ed. Treatiseon Invertebrate Paleontology. Lawrence(KS): The Geological Society of Americaand the University of Kansas.
fur-oxidizing bacteria and a marine nema-tode. Applied and Environmental Micro-biology 60: 4461-4467.
Reid RGB. 1980. Aspects of the biology of agutless species of Solemya (Bivalvia:Protobranchia). Canadian Journal of Zo-ology 58: 386-393.
R e i d R G B , B e r n a r d F R . 1 9 8 0 . G u t l e s sbivalves. Science 208: 609-6 10.
Reid RGB, Brand DC. 1986. Sulfide-oxidiz-ing symbiosis in Lucinaceans: Implicationsfor bivalve evolution. Veliger 29: 3-24.
Seilacher A. 1990. Aberrations in bivalveevolution related to photo- and chemo-symbiosis. Historical Biology 3: 289-311.
Smith CR, Kukert H, Wheatcroft RA, JumarsPA, Deming JW. 1989. Vent fauna onwhale remains. Nature 341: 27-28.
Southward AJ. 1989. Animal communitiesfueled by chemosynthesis life at hydrother-mal vents cold seeps and in reducing sedi-ments. Journal of Zoology 217: 705-709.
Southward EC. 1987. Contribution of sym-biotic chemoautotrophs to the nutritionof benthic invertebrates. Pages 83-l 18 inSleigh MA, ed. Microbes in the Sea.Chichester (UK): Ellis Horwood.
Streams ME, Fisher CR, Fiaia-MCdioni A.1997. Methanotrophic symbiont locationand fa te of carbon incorporated f rommethane in a hydrocarbon seep mussel.Marme Biology 129: 465476.
Taviani IM. 1994. The “calcari a Lucina”macrofauna reconsidered: Deep-sea fau-nal oases from Miocene-age cold vents inthe Romagna Appennine, Italy. Geo-Ma-rme Letters 14: 185-191.
Tunnicliffe V. 1992. The nature and origin ofthe modern hydrothermal vent fauna.Palalos 7: 338-350.
Turner R. 1985. Notes on molluscs of deep-sea vents and reducing sediments. Ameri-can LMalacological Bulletm (special edi-tion) 1: 23-34.
286 Bioscience Vol. 48 No. 4





























