Embed Size (px)
Citation preview

Life Sciences 89 (2011) 779–785
Contents lists available at SciVerse ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / l i fesc ie
(−)-Epigallocatechin-3-gallate blocks 3T3-L1 adipose conversion by inhibition of cellproliferation and suppression of adipose phenotype expression
Cheuk Ying Chan a,1, Lai Wei b,1, Federico Castro-Muñozledo c, Wing Leung Koo a,⁎a Department of Pharmacology & Pharmacy, The University of Hong Kong, Hong Kong SARb Department of Pathology, The University of Hong Kong, Hong Kong SARc Department of Cell Biology, CINVESTAV-IPN, Mexico
⁎ Corresponding author at: Department of PharmacMedicine, 21 Sassoon Road, Pokfulam, Hong Kongfax: +852 28170859.
E-mail address: [email protected] (W.L. Koo).1 Contributed equally to this work.
0024-3205/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.lfs.2011.09.006
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 22 March 2011Accepted 27 August 2011Keywords:(−)-Epigallocatechin-3-gallate (EGCG)3T3-L1AdipogenesisObesityC/EBPαPPARγCatechins
Aims: A major objective in the treatment of obese individuals is the development of agents that reduce bodyfat and improve metabolic homeostasis. Among a variety of biological effects, green tea exerts an anti-obesityeffect; however, the mechanisms behind its activity on adipose tissue are uncertain. Tea contains high levelsof (−)-epigallocatechin-3-gallate (EGCG), which is one of itsmain bioactive substances. Therefore, we studiedthe effects of EGCG on mouse embryonic fibroblast-adipose like cell line (3T3-L1) preadipocyte proliferationand differentiation.Main methods: 3T3-L1 cells were incubated with physiologically attainable (0.1–1 μM) and pharmacological(5 μM, 10 μM) concentrations of EGCG for various lengths of time. Cell proliferation was assessed by cellcounting and cell cycle analysis. Adipose conversion was evaluated by lipid accumulation and expressionof CCAAT/enhancer binding protein alpha (C/EBPα) and peroxisome proliferator-activated receptor gamma(PPARγ) transcription factors.
Key findings: A dose dependent suppressive effect on preadipocyte proliferation was observed, with the highestreduction in cell number at 10 μM EGCG. On the other hand, adipose conversion was fully inhibited with 10 μMEGCG. Flow-cytometric analysis showed that 3T3-L1 cells treated with EGCG underwent an arrest of cell cycle atG2/M. The inhibition of the expression of C/EBPα and PPARγwas accompanied by the inhibitory effect of EGCG.Microscopic observation showed that 3T3-L1 cells treated with EGCG maintained the fibroblastic shape andfailed to accumulate cytoplasmic fat droplets even after the induction of differentiation.Significance:Our results suggest that EGCG reduces adipogenesis through an arrest of cell cycle and a blockage ofadipose phenotype expression. These results also suggest that the anti-obesity activity of green teamaybe partiallyattributed to its suppressive effects in adipogenesis.© 2011 Elsevier Inc. All rights reserved.
Introduction
The health-promoting effects of green tea are mainly attributedto its polyphenol content. (−)-Epigallocatechin-3-gallate (EGCG) isthe major polyphenol catechin found in green tea and seems to beresponsible for the beneficial effects of green tea (Graham, 1992).EGCG has been considered as a preventive agent for cancer, obesity,diabetes and cardiovascular diseases (Ahmad and Mukhtar, 1999;Crespy and Williamson, 2004; Kao et al., 2000; Lin et al., 1999;Mitscher et al., 1997). It is believed that regular consumption of tea,that is rich in EGCG, may decrease the incidence of obesity, reducebodyweight and fat depot size (Nagao et al., 2005). EGCG is a powerful
ology & Pharmacy, Faculty ofSAR. Tel.: +852 28199256;
rights reserved.
inhibitor of lipid absorption (Koo and Noh, 2007; Raederstorff et al.,2003), probably through its ability to form complexes with lipidsand lipolytic enzyme (Koo and Noh, 2007; Raederstorff et al., 2003).The extent of catechin effects on health depends on the amountconsumed and on their bioavailability (Khan and Mukhtar, 2007;Manach et al., 2005).
Several pharmacological effects of catechins were reported afterin-vitro studies in which EGCG was used at high concentrations(10–100 μM). However, in many animal and human studies theusual plasma concentrations of EGCG were found to be around1 μM. Since EGCG has a half-life of about 5 to 6 h (Yang et al., 1998),and it is slowly removed from the body, therefore higher plasmaconcentrations of EGCG could be achievedwhen it is regularly consumed(Manach et al., 2004).
Obesity is a major health issue in developed countries, and itsfrequency is increasing both in developing and undeveloped countries.Obesity has now become a worldwide health problem, including China(Cheng, 2004a, 2004b; Mokdad et al., 2003). The latest WHO globalprojection of peoplewith obesity is 400 million, with another 1.6 billion

Fig. 1. Schematic representation of the experimental design. 3T3-L1 cells were seededas indicated (see Methods). (A) One day after plating cell cultures were supplementedwith medium containing EGCG at the indicated concentrations, and cells were grownto post-confluent (Day −5 to Day 0). (B) Two days after confluent (Day 0), cellswere changed to medium without EGCG and cultured until the eighth-day (Day 0 toDay 8). (C) Other cultures were induced to differentiation for 2 days as indicated(see Materials and methods), in the presence of EGCG (Day −5 to Day 8). (D) Finally,other cultures were supplemented with EGCG from Day −5, to the end of experiment(Day −5 to Day 13).
780 C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
overweight. Development of obesity is characterized by hyperplasiaand hypertrophy of adipose tissue.
Evidence suggested that EGCG induces lipolysis (Lee et al., 2009a,2009b; Mochizuki and Hasegawa, 2004), modulates the expression ofadipocyte-related genes (Hsu and Yen, 2008; Lee et al., 2009a, 2009b;Moon et al., 2007) and down-regulates lipogenic enzyme activities(Kao et al., 2006; Wolfram et al., 2005). A high concentration ofEGCG was found in green tea (Lin et al., 2003; Yang et al., 2001).Thus, a better understanding of the mechanism through which EGCGmodulates adipogenesis would be of great value to obese patients.
When mouse embryonic fibroblast - adipose like cell line (3T3-L1)cells undergo differentiation, they show striking cell shape changesand massive lipid accumulation. These changes are controlled andregulated by the expression and activity of transcription factors(Rosen et al., 2000). In the present study, the effects of EGCG on thedifferentiation of adipocytes, usingmouse 3T3-L1 cells as an experimentalmodel, were analyzed. In order to simulate real life situation, the effectsof physiologically attainable concentrations of EGCG on protein expres-sion and metabolic signaling pathways during adipogenesis wereinvestigated.
Materials and methods
Cell culture
3T3-L1Apreadipocytes (a gift fromM.Daniel Lane, The JohnsHopkinsUniversity School of Medicine, Maryland, USA) were maintained andpropagated in Dulbecco's modified Eagle medium (DMEM) (Invitrogen)containing 10% (v/v) calf serum (Invitrogen). Two-days post confluentcells were induced to differentiate into adipocytes with the addition ofdifferentiation mixture (designated as Day 0) with DMEM containing10% fetal bovine serum (FBS) (Invitrogen Life Technologies), 1 μg/mlinsulin (I) (Roche), 0.5 mM 3-isobutyl-1-methylxanthine (M), and1 μM dexamethasone (D). Two days later, culture medium waschanged to DMEM supplemented with 10% FBS and 1 μg/ml insulin for2 days. The medium was then replenished every other day with DMEMand 10% FBS during different periods (Fig. 1A–D). To study the effectsof EGCG, cell cultures were supplemented with EGCG at variousconcentrations, during the growth phase (from Day −5 to Day 0), fromthe growth phase to the differentiation phase (Day −5 to Day 8), thetime of stimulation of adipose conversion with the differentiationmixture (Day 0 to Day 8) and from the growth phase to matureadipocytes (Day −5 to Day 13) (see Fig. 1A–D). In all cases EGCG wasreplenished twice a day.
Cell counting
Cells were seeded into 24-well plates and treated with EGCG. Onthe day of counting, cells were rinsed twice with 37 °C sterile PBSand trypsinized, followed by centrifugation at 1000 rpm for 5 min.The supernatant was discarded and the cells were harvested in 3 mlsterile PBS. Cells were then counted with a Coulter Counter (CoulterZ™Series, Beckman Coulter, Inc, USA) at the indicated periods.
Cell cycle analysis by flow cytometry
The cell cycle distribution of 3T3-L1 cells grown on six-well plateswas determined by flow cytometry. The cells were harvested at theindicated time points. Cells were trypsinized, washed twice withcold PBS and resuspended in 300 μl PBS, followed by fixing with70% ice-cooled ethanol overnight at 4 °C. Cells were centrifuged andthe pellets were resuspended in 100 μl PBS containing 100 μg/mlDNase-free RNase A. Afterwards, cells were incubated at roomtemperature for 5 min. Then, 500 μl of 50 μg/ml propidium iodidewas added to cell suspension and incubated in the dark for further20 min at room temperature. Cell cycle was determined by using
flow cytometry (BD, FACSCalibur) and analyzed with MolFit softwareto determine the relative amount of cells in G1, S, and G2/M phases. Atleast 10,000 events were acquired per sample.
Oil-Red O staining and absorbance reading
Oil Red O staining and quantitation of triglycerides content wereperformed as previously described (Ramirez-Zacarias et al., 1992)with some modifications. Briefly, on Day 8 or Day 13, cells in 24-wellplate were rinsed by PBS twice. The cells were fixed with cold10% formaldehyde in isotonic solution for 1 hour. A 0.5% Oil RedO isopropanol solution was diluted with 1.5 volume of water, filtered,and added to the fixed adipocyte monolayer for 1 h. Cells were thenwashed with water. The stained triglyceride droplets were extractedwith 1 ml isopropanol and the absorbance was read at 510 nm. Thetriglyceride content is standardized by triolein (Ramirez-Zacarias etal., 1992).
Cells were grown on plastic coverslips in 24-well culture platesand pretreated with different concentrations of EGCG. Cells were fixedand stained with Oil Red O and later counterstained with Hematoxylin.They were then visualized by light microscopy and photographed.
Immunoblotting
At various time points, cell monolayers (6-cm dishes) werewashed once with cold PBS, pH 7.4, and then scraped into lysis buffer.Lysateswere heated at 95 °C for 10 min and clarified by centrifugation,and equal amounts of protein were separated by SDS/PAGE. Proteinswere transferred to PVDF membranes and immunoblotted withantibodies to CCAAT/ enhancer binding protein alpha (C/EBPα) andperoxisome proliferator-activated receptor gamma (PPARγ) (CellSignaling Technology, Inc., Danvers, MA).
RNA isolation and RT-PCR
RNA was extracted at various time points using TRIzol reagent(Life Technologies, Inc., Grand Island, NY, USA). For the RT reaction,
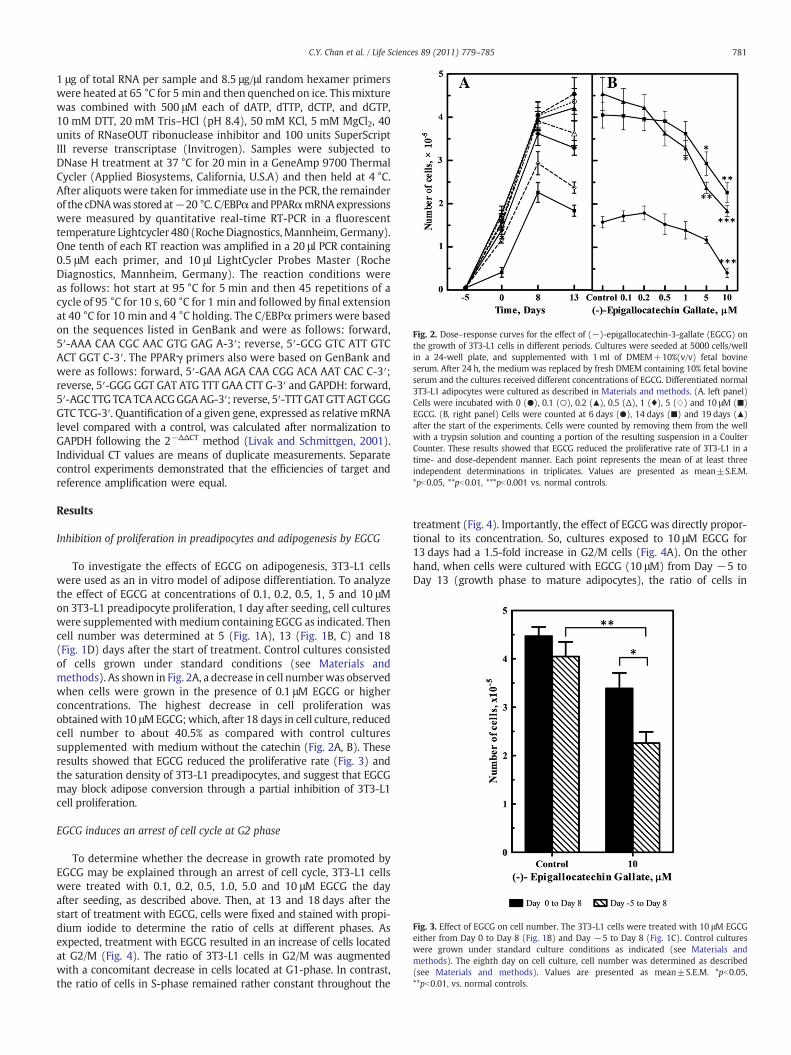
Fig. 2. Dose–response curves for the effect of (−)-epigallocatechin-3-gallate (EGCG) onthe growth of 3T3-L1 cells in different periods. Cultures were seeded at 5000 cells/wellin a 24-well plate, and supplemented with 1 ml of DMEM+10%(v/v) fetal bovineserum. After 24 h, the medium was replaced by fresh DMEM containing 10% fetal bovineserum and the cultures received different concentrations of EGCG. Differentiated normal3T3-L1 adipocytes were cultured as described in Materials and methods. (A. left panel)Cells were incubated with 0 (●), 0.1 (○), 0.2 (▲), 0.5 (Δ), 1 (♦), 5 (⋄) and 10 μM (■)EGCG. (B, right panel) Cells were counted at 6 days (●), 14 days (■) and 19 days (▲)after the start of the experiments. Cells were counted by removing them from the wellwith a trypsin solution and counting a portion of the resulting suspension in a CoulterCounter. These results showed that EGCG reduced the proliferative rate of 3T3-L1 in atime- and dose-dependent manner. Each point represents the mean of at least threeindependent determinations in triplicates. Values are presented as mean±S.E.M.*pb0.05, **pb0.01, ***pb0.001 vs. normal controls.
Fig. 3. Effect of EGCG on cell number. The 3T3-L1 cells were treated with 10 μM EGCGeither from Day 0 to Day 8 (Fig. 1B) and Day −5 to Day 8 (Fig. 1C). Control cultureswere grown under standard culture conditions as indicated (see Materials andmethods). The eighth day on cell culture, cell number was determined as described(see Materials and methods). Values are presented as mean±S.E.M. *pb0.05,**pb0.01, vs. normal controls.
781C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
1 μg of total RNA per sample and 8.5 μg/μl random hexamer primerswere heated at 65 °C for 5 min and then quenched on ice. Thismixturewas combined with 500 μM each of dATP, dTTP, dCTP, and dGTP,10 mM DTT, 20 mM Tris–HCl (pH 8.4), 50 mM KCl, 5 mM MgCl2, 40units of RNaseOUT ribonuclease inhibitor and 100 units SuperScriptIII reverse transcriptase (Invitrogen). Samples were subjected toDNase H treatment at 37 °C for 20 min in a GeneAmp 9700 ThermalCycler (Applied Biosystems, California, U.S.A) and then held at 4 °C.After aliquots were taken for immediate use in the PCR, the remainderof the cDNAwas stored at−20 °C. C/EBPα andPPARαmRNAexpressionswere measured by quantitative real-time RT-PCR in a fluorescenttemperature Lightcycler 480 (RocheDiagnostics,Mannheim, Germany).One tenth of each RT reaction was amplified in a 20 μl PCR containing0.5 μM each primer, and 10 μl LightCycler Probes Master (RocheDiagnostics, Mannheim, Germany). The reaction conditions wereas follows: hot start at 95 °C for 5 min and then 45 repetitions of acycle of 95 °C for 10 s, 60 °C for 1 min and followed by final extensionat 40 °C for 10 min and 4 °C holding. The C/EBPα primers were basedon the sequences listed in GenBank and were as follows: forward,5′-AAA CAA CGC AAC GTG GAG A-3′; reverse, 5′-GCG GTC ATT GTCACT GGT C-3′. The PPARγ primers also were based on GenBank andwere as follows: forward, 5′-GAA AGA CAA CGG ACA AAT CAC C-3′;reverse, 5′-GGG GGT GAT ATG TTT GAA CTT G-3′ and GAPDH: forward,5′-AGCTTG TCATCAACGGGAAG-3′; reverse, 5′-TTTGATGTTAGTGGGGTC TCG-3′. Quantification of a given gene, expressed as relative mRNAlevel compared with a control, was calculated after normalization toGAPDH following the 2−ΔΔCT method (Livak and Schmittgen, 2001).Individual CT values are means of duplicate measurements. Separatecontrol experiments demonstrated that the efficiencies of target andreference amplification were equal.
Results
Inhibition of proliferation in preadipocytes and adipogenesis by EGCG
To investigate the effects of EGCG on adipogenesis, 3T3-L1 cellswere used as an in vitro model of adipose differentiation. To analyzethe effect of EGCG at concentrations of 0.1, 0.2, 0.5, 1, 5 and 10 μMon 3T3-L1 preadipocyte proliferation, 1 day after seeding, cell cultureswere supplemented withmedium containing EGCG as indicated. Thencell number was determined at 5 (Fig. 1A), 13 (Fig. 1B, C) and 18(Fig. 1D) days after the start of treatment. Control cultures consistedof cells grown under standard conditions (see Materials andmethods). As shown in Fig. 2A, a decrease in cell numberwas observedwhen cells were grown in the presence of 0.1 μM EGCG or higherconcentrations. The highest decrease in cell proliferation wasobtainedwith 10 μMEGCG;which, after 18 days in cell culture, reducedcell number to about 40.5% as compared with control culturessupplemented with medium without the catechin (Fig. 2A, B). Theseresults showed that EGCG reduced the proliferative rate (Fig. 3) andthe saturation density of 3T3-L1 preadipocytes, and suggest that EGCGmay block adipose conversion through a partial inhibition of 3T3-L1cell proliferation.
EGCG induces an arrest of cell cycle at G2 phase
To determine whether the decrease in growth rate promoted byEGCG may be explained through an arrest of cell cycle, 3T3-L1 cellswere treated with 0.1, 0.2, 0.5, 1.0, 5.0 and 10 μM EGCG the dayafter seeding, as described above. Then, at 13 and 18 days after thestart of treatment with EGCG, cells were fixed and stained with propi-dium iodide to determine the ratio of cells at different phases. Asexpected, treatment with EGCG resulted in an increase of cells locatedat G2/M (Fig. 4). The ratio of 3T3-L1 cells in G2/M was augmentedwith a concomitant decrease in cells located at G1-phase. In contrast,the ratio of cells in S-phase remained rather constant throughout the
treatment (Fig. 4). Importantly, the effect of EGCG was directly propor-tional to its concentration. So, cultures exposed to 10 μM EGCG for13 days had a 1.5-fold increase in G2/M cells (Fig. 4A). On the otherhand, when cells were cultured with EGCG (10 μM) from Day −5 toDay 13 (growth phase to mature adipocytes), the ratio of cells in
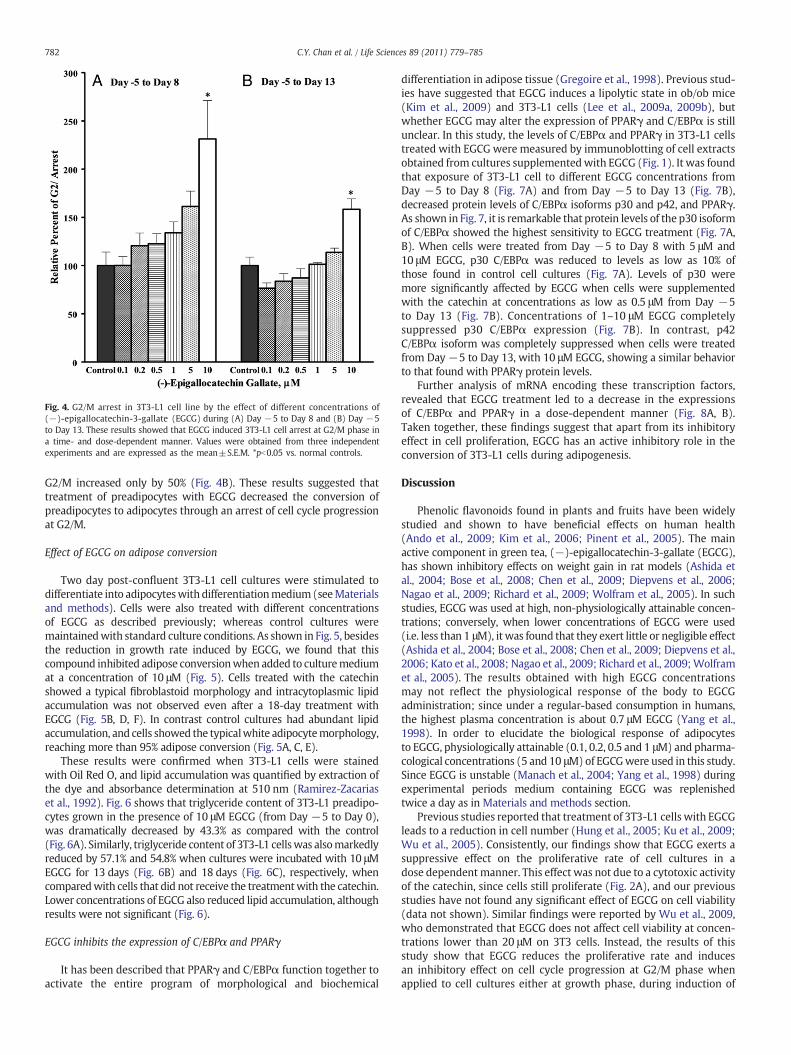
Fig. 4. G2/M arrest in 3T3-L1 cell line by the effect of different concentrations of(−)-epigallocatechin-3-gallate (EGCG) during (A) Day −5 to Day 8 and (B) Day −5to Day 13. These results showed that EGCG induced 3T3-L1 cell arrest at G2/M phase ina time- and dose-dependent manner. Values were obtained from three independentexperiments and are expressed as the mean±S.E.M. *pb0.05 vs. normal controls.
782 C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
G2/M increased only by 50% (Fig. 4B). These results suggested thattreatment of preadipocytes with EGCG decreased the conversion ofpreadipocytes to adipocytes through an arrest of cell cycle progressionat G2/M.
Effect of EGCG on adipose conversion
Two day post-confluent 3T3-L1 cell cultures were stimulated todifferentiate into adipocyteswith differentiationmedium (seeMaterialsand methods). Cells were also treated with different concentrationsof EGCG as described previously; whereas control cultures weremaintainedwith standard culture conditions. As shown in Fig. 5, besidesthe reduction in growth rate induced by EGCG, we found that thiscompound inhibited adipose conversionwhen added to culturemediumat a concentration of 10 μM (Fig. 5). Cells treated with the catechinshowed a typical fibroblastoid morphology and intracytoplasmic lipidaccumulation was not observed even after a 18-day treatment withEGCG (Fig. 5B, D, F). In contrast control cultures had abundant lipidaccumulation, and cells showed the typicalwhite adipocytemorphology,reaching more than 95% adipose conversion (Fig. 5A, C, E).
These results were confirmed when 3T3-L1 cells were stainedwith Oil Red O, and lipid accumulation was quantified by extraction ofthe dye and absorbance determination at 510 nm (Ramirez-Zacariaset al., 1992). Fig. 6 shows that triglyceride content of 3T3-L1 preadipo-cytes grown in the presence of 10 μM EGCG (from Day −5 to Day 0),was dramatically decreased by 43.3% as compared with the control(Fig. 6A). Similarly, triglyceride content of 3T3-L1 cellswas alsomarkedlyreduced by 57.1% and 54.8% when cultures were incubated with 10 μMEGCG for 13 days (Fig. 6B) and 18 days (Fig. 6C), respectively, whencomparedwith cells that did not receive the treatmentwith the catechin.Lower concentrations of EGCG also reduced lipid accumulation, althoughresults were not significant (Fig. 6).
EGCG inhibits the expression of C/EBPα and PPARγ
It has been described that PPARγ and C/EBPα function together toactivate the entire program of morphological and biochemical
differentiation in adipose tissue (Gregoire et al., 1998). Previous stud-ies have suggested that EGCG induces a lipolytic state in ob/ob mice(Kim et al., 2009) and 3T3-L1 cells (Lee et al., 2009a, 2009b), butwhether EGCG may alter the expression of PPARγ and C/EBPα is stillunclear. In this study, the levels of C/EBPα and PPARγ in 3T3-L1 cellstreated with EGCG were measured by immunoblotting of cell extractsobtained from cultures supplementedwith EGCG (Fig. 1). It was foundthat exposure of 3T3-L1 cell to different EGCG concentrations fromDay −5 to Day 8 (Fig. 7A) and from Day −5 to Day 13 (Fig. 7B),decreased protein levels of C/EBPα isoforms p30 and p42, and PPARγ.As shown in Fig. 7, it is remarkable that protein levels of the p30 isoformof C/EBPα showed the highest sensitivity to EGCG treatment (Fig. 7A,B). When cells were treated from Day −5 to Day 8 with 5 μM and10 μM EGCG, p30 C/EBPα was reduced to levels as low as 10% ofthose found in control cell cultures (Fig. 7A). Levels of p30 weremore significantly affected by EGCG when cells were supplementedwith the catechin at concentrations as low as 0.5 μM from Day −5to Day 13 (Fig. 7B). Concentrations of 1–10 μM EGCG completelysuppressed p30 C/EBPα expression (Fig. 7B). In contrast, p42C/EBPα isoform was completely suppressed when cells were treatedfrom Day −5 to Day 13, with 10 μM EGCG, showing a similar behaviorto that found with PPARγ protein levels.
Further analysis of mRNA encoding these transcription factors,revealed that EGCG treatment led to a decrease in the expressionsof C/EBPα and PPARγ in a dose-dependent manner (Fig. 8A, B).Taken together, these findings suggest that apart from its inhibitoryeffect in cell proliferation, EGCG has an active inhibitory role in theconversion of 3T3-L1 cells during adipogenesis.
Discussion
Phenolic flavonoids found in plants and fruits have been widelystudied and shown to have beneficial effects on human health(Ando et al., 2009; Kim et al., 2006; Pinent et al., 2005). The mainactive component in green tea, (−)-epigallocatechin-3-gallate (EGCG),has shown inhibitory effects on weight gain in rat models (Ashida etal., 2004; Bose et al., 2008; Chen et al., 2009; Diepvens et al., 2006;Nagao et al., 2009; Richard et al., 2009; Wolfram et al., 2005). In suchstudies, EGCG was used at high, non-physiologically attainable concen-trations; conversely, when lower concentrations of EGCG were used(i.e. less than 1 μM), it was found that they exert little or negligible effect(Ashida et al., 2004; Bose et al., 2008; Chen et al., 2009; Diepvens et al.,2006; Kato et al., 2008; Nagao et al., 2009; Richard et al., 2009;Wolframet al., 2005). The results obtained with high EGCG concentrationsmay not reflect the physiological response of the body to EGCGadministration; since under a regular-based consumption in humans,the highest plasma concentration is about 0.7 μM EGCG (Yang et al.,1998). In order to elucidate the biological response of adipocytesto EGCG, physiologically attainable (0.1, 0.2, 0.5 and 1 μM) and pharma-cological concentrations (5 and 10 μM) of EGCGwere used in this study.Since EGCG is unstable (Manach et al., 2004; Yang et al., 1998) duringexperimental periods medium containing EGCG was replenishedtwice a day as in Materials and methods section.
Previous studies reported that treatment of 3T3-L1 cells with EGCGleads to a reduction in cell number (Hung et al., 2005; Ku et al., 2009;Wu et al., 2005). Consistently, our findings show that EGCG exerts asuppressive effect on the proliferative rate of cell cultures in adose dependent manner. This effect was not due to a cytotoxic activityof the catechin, since cells still proliferate (Fig. 2A), and our previousstudies have not found any significant effect of EGCG on cell viability(data not shown). Similar findings were reported by Wu et al., 2009,who demonstrated that EGCG does not affect cell viability at concen-trations lower than 20 μM on 3T3 cells. Instead, the results of thisstudy show that EGCG reduces the proliferative rate and inducesan inhibitory effect on cell cycle progression at G2/M phase whenapplied to cell cultures either at growth phase, during induction of
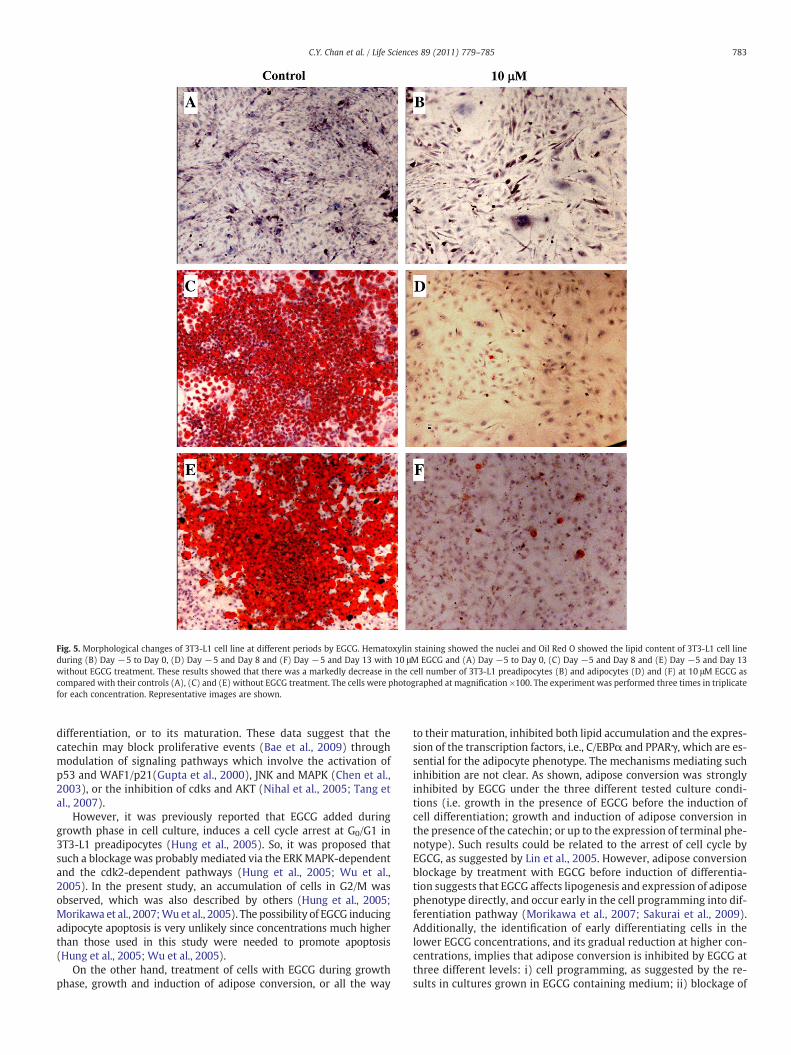
Fig. 5. Morphological changes of 3T3-L1 cell line at different periods by EGCG. Hematoxylin staining showed the nuclei and Oil Red O showed the lipid content of 3T3-L1 cell lineduring (B) Day −5 to Day 0, (D) Day −5 and Day 8 and (F) Day −5 and Day 13 with 10 μM EGCG and (A) Day −5 to Day 0, (C) Day −5 and Day 8 and (E) Day −5 and Day 13without EGCG treatment. These results showed that there was a markedly decrease in the cell number of 3T3-L1 preadipocytes (B) and adipocytes (D) and (F) at 10 μM EGCG ascompared with their controls (A), (C) and (E) without EGCG treatment. The cells were photographed at magnification ×100. The experiment was performed three times in triplicatefor each concentration. Representative images are shown.
783C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
differentiation, or to its maturation. These data suggest that thecatechin may block proliferative events (Bae et al., 2009) throughmodulation of signaling pathways which involve the activation ofp53 and WAF1/p21(Gupta et al., 2000), JNK and MAPK (Chen et al.,2003), or the inhibition of cdks and AKT (Nihal et al., 2005; Tang etal., 2007).
However, it was previously reported that EGCG added duringgrowth phase in cell culture, induces a cell cycle arrest at G0/G1 in3T3-L1 preadipocytes (Hung et al., 2005). So, it was proposed thatsuch a blockage was probably mediated via the ERKMAPK-dependentand the cdk2-dependent pathways (Hung et al., 2005; Wu et al.,2005). In the present study, an accumulation of cells in G2/M wasobserved, which was also described by others (Hung et al., 2005;Morikawa et al., 2007;Wu et al., 2005). The possibility of EGCG inducingadipocyte apoptosis is very unlikely since concentrations much higherthan those used in this study were needed to promote apoptosis(Hung et al., 2005; Wu et al., 2005).
On the other hand, treatment of cells with EGCG during growthphase, growth and induction of adipose conversion, or all the way
to their maturation, inhibited both lipid accumulation and the expres-sion of the transcription factors, i.e., C/EBPα and PPARγ, which are es-sential for the adipocyte phenotype. The mechanisms mediating suchinhibition are not clear. As shown, adipose conversion was stronglyinhibited by EGCG under the three different tested culture condi-tions (i.e. growth in the presence of EGCG before the induction ofcell differentiation; growth and induction of adipose conversion inthe presence of the catechin; or up to the expression of terminal phe-notype). Such results could be related to the arrest of cell cycle byEGCG, as suggested by Lin et al., 2005. However, adipose conversionblockage by treatment with EGCG before induction of differentia-tion suggests that EGCG affects lipogenesis and expression of adiposephenotype directly, and occur early in the cell programming into dif-ferentiation pathway (Morikawa et al., 2007; Sakurai et al., 2009).Additionally, the identification of early differentiating cells in thelower EGCG concentrations, and its gradual reduction at higher con-centrations, implies that adipose conversion is inhibited by EGCG atthree different levels: i) cell programming, as suggested by the re-sults in cultures grown in EGCG containing medium; ii) blockage of
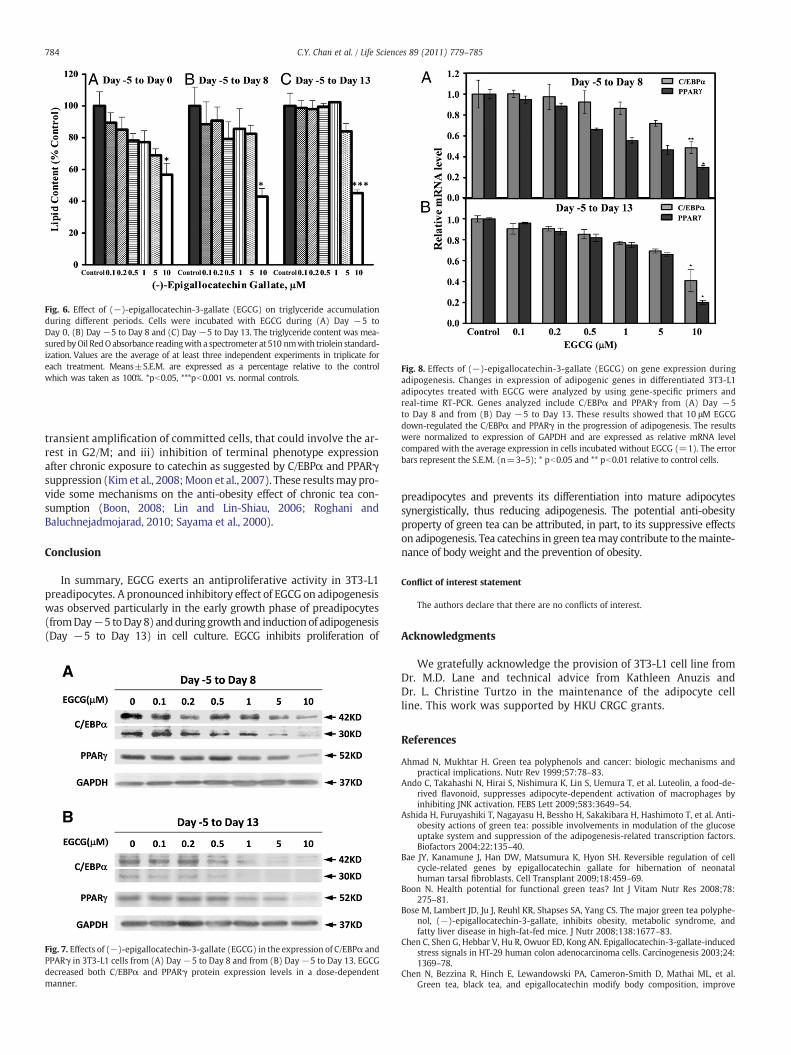
Fig. 6. Effect of (−)-epigallocatechin-3-gallate (EGCG) on triglyceride accumulationduring different periods. Cells were incubated with EGCG during (A) Day −5 toDay 0, (B) Day −5 to Day 8 and (C) Day −5 to Day 13. The triglyceride content was mea-sured byOil RedOabsorbance readingwith a spectrometer at 510 nmwith triolein standard-ization. Values are the average of at least three independent experiments in triplicate foreach treatment. Means±S.E.M. are expressed as a percentage relative to the controlwhich was taken as 100%. *pb0.05, ***pb0.001 vs. normal controls.
Fig. 8. Effects of (−)-epigallocatechin-3-gallate (EGCG) on gene expression duringadipogenesis. Changes in expression of adipogenic genes in differentiated 3T3-L1adipocytes treated with EGCG were analyzed by using gene-specific primers andreal-time RT-PCR. Genes analyzed include C/EBPα and PPARγ from (A) Day −5to Day 8 and from (B) Day −5 to Day 13. These results showed that 10 μM EGCGdown-regulated the C/EBPα and PPARγ in the progression of adipogenesis. The resultswere normalized to expression of GAPDH and are expressed as relative mRNA levelcompared with the average expression in cells incubated without EGCG (=1). The errorbars represent the S.E.M. (n=3–5); * pb0.05 and ** pb0.01 relative to control cells.
784 C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
transient amplification of committed cells, that could involve the ar-rest in G2/M; and iii) inhibition of terminal phenotype expressionafter chronic exposure to catechin as suggested by C/EBPα and PPARγsuppression (Kim et al., 2008;Moon et al., 2007). These resultsmaypro-vide some mechanisms on the anti-obesity effect of chronic tea con-sumption (Boon, 2008; Lin and Lin-Shiau, 2006; Roghani andBaluchnejadmojarad, 2010; Sayama et al., 2000).
Conclusion
In summary, EGCG exerts an antiproliferative activity in 3T3-L1preadipocytes. A pronounced inhibitory effect of EGCG on adipogenesiswas observed particularly in the early growth phase of preadipocytes(fromDay−5 toDay 8) andduring growth and induction of adipogenesis(Day −5 to Day 13) in cell culture. EGCG inhibits proliferation of
Fig. 7. Effects of (−)-epigallocatechin-3-gallate (EGCG) in the expression of C/EBPα andPPARγ in 3T3-L1 cells from (A) Day−5 to Day 8 and from (B) Day−5 to Day 13. EGCGdecreased both C/EBPα and PPARγ protein expression levels in a dose-dependentmanner.
preadipocytes and prevents its differentiation into mature adipocytessynergistically, thus reducing adipogenesis. The potential anti-obesityproperty of green tea can be attributed, in part, to its suppressive effectson adipogenesis. Tea catechins in green teamay contribute to themainte-nance of body weight and the prevention of obesity.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgments
We gratefully acknowledge the provision of 3T3-L1 cell line fromDr. M.D. Lane and technical advice from Kathleen Anuzis andDr. L. Christine Turtzo in the maintenance of the adipocyte cellline. This work was supported by HKU CRGC grants.
References
Ahmad N, Mukhtar H. Green tea polyphenols and cancer: biologic mechanisms andpractical implications. Nutr Rev 1999;57:78–83.
Ando C, Takahashi N, Hirai S, Nishimura K, Lin S, Uemura T, et al. Luteolin, a food-de-rived flavonoid, suppresses adipocyte-dependent activation of macrophages byinhibiting JNK activation. FEBS Lett 2009;583:3649–54.
Ashida H, Furuyashiki T, Nagayasu H, Bessho H, Sakakibara H, Hashimoto T, et al. Anti-obesity actions of green tea: possible involvements in modulation of the glucoseuptake system and suppression of the adipogenesis-related transcription factors.Biofactors 2004;22:135–40.
Bae JY, Kanamune J, Han DW, Matsumura K, Hyon SH. Reversible regulation of cellcycle-related genes by epigallocatechin gallate for hibernation of neonatalhuman tarsal fibroblasts. Cell Transplant 2009;18:459–69.
Boon N. Health potential for functional green teas? Int J Vitam Nutr Res 2008;78:275–81.
Bose M, Lambert JD, Ju J, Reuhl KR, Shapses SA, Yang CS. The major green tea polyphe-nol, (−)-epigallocatechin-3-gallate, inhibits obesity, metabolic syndrome, andfatty liver disease in high-fat-fed mice. J Nutr 2008;138:1677–83.
Chen C, Shen G, Hebbar V, Hu R, Owuor ED, Kong AN. Epigallocatechin-3-gallate-inducedstress signals in HT-29 human colon adenocarcinoma cells. Carcinogenesis 2003;24:1369–78.
Chen N, Bezzina R, Hinch E, Lewandowski PA, Cameron-Smith D, Mathai ML, et al.Green tea, black tea, and epigallocatechin modify body composition, improve

785C.Y. Chan et al. / Life Sciences 89 (2011) 779–785
glucose tolerance, and differentially alter metabolic gene expression in rats fed ahigh-fat diet. Nutr Res 2009;29:784–93.
Cheng TO. Obesity crisis comprised of danger and opportunity. J Am Diet Assoc2004a;104:1546.
Cheng TO. Childhood obesity among the Chinese [correction]. Circulation 2004b;110(12):e314. author reply e314.
Crespy V, Williamson G. A review of the health effects of green tea catechins in in vivoanimal models. J Nutr 2004;134:3431S–40S.
Diepvens K, Kovacs EM, Vogels N, Westerterp-Plantenga MS. Metabolic effects of greentea and of phases of weight loss. Physiol Behav 2006;87:185–91.
Graham HN. Green tea composition, consumption, and polyphenol chemistry. PrevMed 1992;21:334–50.
Gregoire FM, Smas CM, Sul HS. Understanding adipocyte differentiation. Physiol Rev1998;78:783–809.
Gupta S, Ahmad N, Nieminen AL, Mukhtar H. Growth inhibition, cell-cycle dysregula-tion, and induction of apoptosis by green tea constituent (−)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinomacells. Toxicol Appl Pharmacol 2000;164:82–90.
Hsu CL, Yen GC. Phenolic compounds: evidence for inhibitory effects against obesityand their underlying molecular signaling mechanisms. Mol Nutr Food Res2008;52:53–61.
Hung PF, Wu BT, Chen HC, Chen YH, Chen CL, Wu MH, et al. Antimitogenic effect ofgreen tea (−)-epigallocatechin gallate on 3T3-L1 preadipocytes depends on theERK and Cdk2 pathways. Am J Physiol Cell Physiol 2005;288:C1094–108.
Kao YH, Hiipakka RA, Liao S. Modulation of endocrine systems and food intake by greentea epigallocatechin gallate. Endocrinology 2000;141:980–7.
Kao YH, Chang HH, Lee MJ, Chen CL. Tea, obesity, and diabetes. Mol Nutr Food Res2006;50:188–210.
Kato K, Long NK, Makita H, Toida M, Yamashita T, Hatakeyama D, et al. Effects of greentea polyphenol on methylation status of RECK gene and cancer cell invasion in oralsquamous cell carcinoma cells. Br J Cancer 2008;99:647–54.
Khan N, Mukhtar H. Tea polyphenols for health promotion. Life Sci 2007;81:519–33.Kim HK, Nelson-Dooley C, Della-Fera MA, Yang JY, Zhang W, Duan J, et al. Genistein de-
creases food intake, body weight, and fat pad weight and causes adipose tissue ap-optosis in ovariectomized female mice. J Nutr 2006;136:409–14.
Kim NH, Choi SK, Kim SJ, Moon PD, Lim HS, Choi IY, et al. Green tea seed oil reducesweight gain in C57BL/6J mice and influences adipocyte differentiation by suppres-sing peroxisome proliferator-activated receptor-gamma. Pflugers Arch 2008;457:293–302.
Kim HJ, Jeon SM, Lee MK, Jung UJ, Shin SK, Choi MS. Antilipogenic effect of green teaextract in C57BL/6J-Lep ob/ob mice. Phytother Res 2009;23:467–71.
Koo SI, Noh SK. Green tea as inhibitor of the intestinal absorption of lipids: potentialmechanism for its lipid-lowering effect. J Nutr Biochem 2007;18:179–83.
Ku HC, Chang HH, Liu HC, Hsiao CH, Lee MJ, Hu YJ, et al. Green tea (−)-epigallocatechingallate inhibits insulin stimulation of 3T3-L1 preadipocyte mitogenesis via the 67-kDa laminin receptor pathway. Am J Physiol Cell Physiol 2009;297:C121–32.
Lee MS, Kim CT, Kim Y. Green tea (−)-epigallocatechin-3-gallate reduces body weightwith regulation of multiple genes expression in adipose tissue of diet-inducedobese mice. Ann Nutr Metab 2009a;54:151–7.
Lee MS, Kim CT, Kim IH, Kim Y. Inhibitory effects of green tea catechin on the lipid ac-cumulation in 3T3-L1 adipocytes. Phytother Res 2009b;23:1088–91.
Lin JK, Lin-Shiau SY. Mechanisms of hypolipidemic and anti-obesity effects of tea andtea polyphenols. Mol Nutr Food Res 2006;50:211–7.
Lin JK, Liang YC, Lin-Shiau SY. Cancer chemoprevention by tea polyphenols throughmitotic signal transduction blockade. Biochem Pharmacol 1999;58:911–5.
Lin YS, Tsai YJ, Tsay JS, Lin JK. Factors affecting the levels of tea polyphenols and caffeinein tea leaves. J Agric Food Chem 2003;51:1864–73.
Lin J, Della-Fera MA, Baile CA. Green tea polyphenol epigallocatechin gallate inhibitsadipogenesis and induces apoptosis in 3T3-L1 adipocytes. Obes Res 2005;13:982–90.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-timequantitative PCR and the 2−ΔΔCT Method. Methods 2001;25:402–8.
Manach C, Scalbert A, Morand C, Remesy C, Jimenez L. Polyphenols: food sources andbioavailability. Am J Clin Nutr 2004;79:727–47.
Manach C, Williamson G, Morand C, Scalbert A, Remesy C. Bioavailability and bioeffi-cacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J ClinNutr 2005;81:230S–42S.
Mitscher LA, Jung M, Shankel D, Dou JH, Steele L, Pillai SP. Chemoprotection: a reviewof the potential therapeutic antioxidant properties of green tea (Camellia sinensis)and certain of its constituents. Med Res Rev 1997;17:327–65.
Mochizuki M, Hasegawa N. Effects of green tea catechin-induced lipolysis on cytosolglycerol content in differentiated 3T3-L1 cells. Phytother Res 2004;18:945–6.
Mokdad AH, Ford ES, Bowman BA, Dietz WH, Vinicor F, Bales VS, et al. Prevalence ofobesity, diabetes, and obesity-related health risk factors, 2001. JAMA 2003;289:76–9.
Moon HS, Chung CS, Lee HG, Kim TG, Choi YJ, Cho CS. Inhibitory effect of (−)-epigallo-catechin-3-gallate on lipid accumulation of 3T3-L1 cells. Obesity (Silver Spring)2007;15:2571–82.
Morikawa K, Ikeda C, Nonaka M, Pei S, Mochizuki M, Mori A, et al. Epigallocatechingallate-induced apoptosis does not affect adipocyte conversion of preadipocytes.Cell Biol Int 2007;31:1379–87.
Nagao T, Komine Y, Soga S, Meguro S, Hase T, Tanaka Y, et al. Ingestion of a tea rich incatechins leads to a reduction in body fat and malondialdehyde-modified LDL inmen. Am J Clin Nutr 2005;81:122–9.
Nagao T, Meguro S, Hase T, Otsuka K, Komikado M, Tokimitsu I, et al. A catechin-richbeverage improves obesity and blood glucose control in patients with type 2 dia-betes. Obesity (Silver Spring) 2009;17:310–7.
Nihal M, Ahmad N, Mukhtar H, Wood GS. Anti-proliferative and proapoptotic effects of(−)-epigallocatechin-3-gallate on human melanoma: possible implications for thechemoprevention of melanoma. Int J Cancer 2005;114:513–21.
Pinent M, Blade MC, Salvado MJ, Arola L, Hackl H, Quackenbush J, et al. Grape-seed de-rived procyanidins interfere with adipogenesis of 3T3-L1 cells at the onset of dif-ferentiation. Int J Obes (Lond) 2005;29:934–41.
Raederstorff DG, Schlachter MF, Elste V, Weber P. Effect of EGCG on lipid absorptionand plasma lipid levels in rats. J Nutr Biochem 2003;14:326–32.
Ramirez-Zacarias JL, Castro-Munozledo F, Kuri-Harcuch W. Quantitation of adiposeconversion and triglycerides by staining intracytoplasmic lipids with Oil red O. His-tochemistry 1992;97:493–7.
Richard D, Kefi K, Barbe U, Poli A, Bausero P, Visioli F. Weight and plasma lipid controlby decaffeinated green tea. Pharmacol Res 2009;59:351–4.
Roghani M, Baluchnejadmojarad T. Hypoglycemic and hypolipidemic effect and antiox-idant activity of chronic epigallocatechin-gallate in streptozotocin-diabetic rats.Pathophysiology 2010;17:55–9.
Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Transcriptional regulation of adipo-genesis. Genes Dev 2000;14:1293–307.
Sakurai N, Mochizuki K, Kameji H, Shimada M, Goda T. (−)-Epigallocatechin gallate en-hances the expression of genes related to insulin sensitivity and adipocyte differ-entiation in 3T3-L1 adipocytes at an early stage of differentiation. Nutrition2009;25:1047–56.
Sayama K, Lin S, Zheng G, Oguni I. Effects of green tea on growth, food utilization andlipid metabolism in mice. In Vivo 2000;14:481–4.
Tang Y, Zhao DY, Elliott S, Zhao W, Curiel TJ, Beckman BS, et al. Epigallocatechin-3 gal-late induces growth inhibition and apoptosis in human breast cancer cells throughsurvivin suppression. Int J Oncol 2007;31:705–11.
Wolfram S, Raederstorff D, Wang Y, Teixeira SR, Elste V, Weber P. TEAVIGO (epigallo-catechin gallate) supplementation prevents obesity in rodents by reducing adiposetissue mass. Ann Nutr Metab 2005;49:54–63.
Wu BT, Hung PF, Chen HC, Huang RN, Chang HH, Kao YH. The apoptotic effect of greentea (−)-epigallocatechin gallate on 3T3-L1 preadipocytes depends on the Cdk2pathway. J Agric Food Chem 2005;53:5695–701.
Wu D, Guo Z, Ren Z, Guo W, Meydani SN. Green tea EGCG suppresses T cell prolifera-tion through impairment of IL-2/IL-2 receptor signaling. Free Radic Biol Med2009;47:636–43.
Yang CS, Chen L, Lee MJ, Balentine D, Kuo MC, Schantz SP. Blood and urine levels of teacatechins after ingestion of different amounts of green tea by human volunteers.Cancer Epidemiol Biomarkers Prev 1998;7:351–4.
Yang M, Wang C, Chen H. Green, oolong and black tea extracts modulate lipid metab-olism in hyperlipidemia rats fed high-sucrose diet. J Nutr Biochem 2001;12:14–20.













![Synthesis of (−)-[4- 3 H ]Epigallocatechin Gallate and Its Metabolic Fate in Rats after Intravenous Administration](https://img.pdfslide.net/doc/110x75/635c9e05eab478482e016c9d/synthesis-of-4-3-h-epigallocatechin-gallate-and-its-metabolic-fate-in-rats.jpg)











![Chemoprevention of fibroid tumors by [−]-epigallocatechin-3-gallate in quail](https://img.pdfslide.net/doc/110x75/631aee3c2784ca2fc00514f1/chemoprevention-of-fibroid-tumors-by-epigallocatechin-3-gallate-in-quail.jpg)



