Embed Size (px)
Citation preview

Molecular Phylogenetics and Evolution 52 (2009) 84–102
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/locate /ympev
Escape from the Ponto-Caspian: Evolution and biogeography of an endemicgoby species flock (Benthophilinae: Gobiidae: Teleostei)
Matthew E. Neilson, Carol A. Stepien *
Great Lakes Genetics Laboratory, Lake Erie Center and Department of Environmental Sciences, University of Toledo, 6200 Bayshore Rd., Toledo, OH 43618, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 29 September 2008Revised 22 December 2008Accepted 30 December 2008Available online 14 January 2009
Keywords:ApolloniaBabkaGobiidaeMesogobiusMitochondrial gene phylogenyNuclear gene phylogenyNeogobiusPonticolaPonto-CaspianProterorhinus
1055-7903/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.ympev.2008.12.023
* Corresponding author. Fax: +1 419 530 8399.E-mail addresses: [email protected]
[email protected] (C.A. Stepien).
Endemic Ponto-Caspian gobies include a flock of �24 ‘‘neogobiin” species (containing the nominal generaand subgenera Apollonia, Babka, Neogobius, Mesogobius, Ponticola, and Proterorhinus; Teleostei: Gobiidae),of which a large proportion (5 species; �21%) recently escaped to invade other freshwater Eurasian sys-tems and the North American Great Lakes. We provide its first comprehensive phylogenetic and biogeo-graphic analysis based on 4709 bp sequences from two mitochondrial and two nuclear genes withmaximum parsimony, likelihood, and Bayesian approaches. We additionally compare its relationshipswith the tadpole gobies (Benthophilus and Caspiosoma), which comprise a related endemic Ponto-Caspiangobiid group; along with a variety of postulated relatives and outgroups. Results of all phylogeneticapproaches are highly congruent and provide very strong support for recognizing the subfamily Bentho-philinae; which encompasses both the ‘‘neogobiins” and tadpole gobies, and genetically diverges fromother Gobiidae subfamilies—including (non-monophyletic) Gobiinae and Gobinellinae. Benthophilinaecontains three tribes: Neogobiini (Neogobius, which is synonymized here with Apollonia; containing thetype species N. fluviatilis, along with N. melanostomus and N. caspius), Ponticolini (containing the generaMesogobius, Proterorhinus, Babka, and Ponticola—elevating the latter two from subgenera and removingthem from the formerly paraphyletic Neogobius), and Benthophilini (tadpole gobies). Within Ponticolini,Proterorhinus and Mesogobius comprise the sister clade of the Ponticola and Babka clade. Further work isneeded to clarify the interrelationships of the tadpole gobies. Invasiveness is widespread in freshwaterand euryhaline taxa of Neogobius, Proterorhinus, Babka, and Ponticola; but not in marine species, Mesogo-bius, or tadpole gobies.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
Exotic species pose one of the most serious threats to nativeecosystems worldwide (Simberloff and Von Holle, 1999; Sax andGaines, 2008) and often present analytical and conceptual chal-lenges—including resolving their taxonomic identity and system-atic relationships. As species introductions increase (Cohen andCarlton, 1998; Lockwood et al., 2006), more nonindigenous taxawill originate from poorly-known groups lacking identificationkeys and analysis with modern phylogenetic methodology. Theseproblems preclude our understanding of fundamental ecologicalrequirements of introduced taxa, including how they adapt to no-vel habitats and alter the evolutionary trajectory of native ecosys-tems (Mooney and Cleland, 2001), thereby impeding effectivemanagement or control. Phylogenetic and biogeographic analysesof DNA sequence data, as accomplished here, thus provide us withthe means to identify invasive taxa, elucidate cryptic species, ana-
ll rights reserved.
(M.E. Neilson), carol.stepie-
lyze whether congeners and relatives invade in concert, and pre-dict potential new invaders.
For example, the ecology of the North American Great Lakes re-cently has been restructured by waves of invaders accidentallyintroduced from ships’ ballast water, primarily from the EurasianPonto-Caspian region (including the Aral, Azov, Black, and CaspianSeas and associated drainages; Mills et al., 1993; Ricciardi andMacIssac, 2000). Notable for their ecological effects are the dreisse-nid zebra and quagga mussels, Dreissena polymorpha and D. bugen-sis, which first appeared in the Great Lakes in the mid-1980s viaballast water introduction. Two Ponto-Caspian gobies then enteredthe Great Lakes in 1990 (Crossman et al., 1992)—the round gobyNeogobius melanostomus (Pallas 1814) (= Apollonia melanostomaper Stepien and Tumeo, 2006) and the freshwater tubenose gobyProterorhinus semilunaris (Heckel 1837) (formerly grouped as a sin-gle species with the marine P. marmoratus [Pallas 1814]; Stepienand Tumeo, 2006). Like the zebra mussel, the round goby spreadrapidly throughout all five Great Lakes (USGS, 2003) and is nowone of the most abundant benthic fish species (Jude and DeBoe,1996; Johnson et al., 2005). Its invasion success likely was aidedby the prevalence of its native dreissenid mussel prey (Ray and

M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 85
Corkum, 1997). Such facilitative interactions among co-evolvedinvaders may significantly augment the success of invasive com-munities (Simberloff and Von Holle, 1999; Ricciardi and MacIssac,2000), with widespread ecological consequences—as has occurredwith the growing dominance of the dreissenid mussel/round gobybenthic community (Vanderploeg et al., 2002).
The round and tubenose gobies are members of an enigmaticgroup native to the Ponto-Caspian region containing �24 speciesarranged (prior to the present study) in four genera (Apollonia,Mesogobius, Neogobius, and Proterorhinus; Miller, 2003b; Stepienand Tumeo, 2006; see Table 1), which have been variously termed‘‘neogobiins”. Several taxa also contain putative subspecies dividedbetween the Black and Caspian Sea basins. This group meets thedefinition of a species flock sensu Greenwood (1984)—a geograph-ically circumscribed, monophyletic taxon characterized by markedradiation. The historic endemism and taxonomic diversity of thePonto-Caspian ‘‘neogobiins” are remarkable, and knowledge oftheir evolutionary history may yield insight on the evolution ofspecies flocks (Johns and Avise, 1998), factors leading to their rapidevolutionary diversification, as well as invasive success in newhabitats.
Despite their remarkable radiation, the systematic relationshipsand placement of Ponto-Caspian ‘‘neogobiin” gobies have been dis-puted and unclear. This phylogenetic confusion is highlighted bythe fact that a large number of the group are invasive [5 species;�21%; including the round goby N. melanostomus, monkey gobyN. fluviatilis (= A. fluviatilis) (Pallas 1814), racer goby N. gymnotra-chelus (Kessler 1857), bighead goby N. kessleri (Gunther 1861),and freshwater tubenose goby P. semilunaris] in freshwater sys-tems of Eastern/Central Europe and/or North America. For exam-ple, N. melanostomus is invasive in both Europe and NorthAmerica, and in the latter has undergone rapid range expansionsince its 1990 introduction (Charlebois et al., 1997) and is impli-cated in the decline of native Great Lakes fishes (Jude et al.,1995; Corkum et al., 2004).
Detailed investigations of morphology, osteology, and system-atics of the Ponto Caspian ‘‘neogobiin” gobies have only recentlybegun; and relationships of the genus Neogobius sensu lato withother taxa have been disputed. For example, members of the gen-era Neogobius sensu lato and Proterorhinus were regarded as sub-genera of Gobius by Vasil’eva (1989, 1991, 1999) based on cranialosteology; which is a historic perspective predating Berg (1949)that was not accepted by the research community (see Miller,2003). Birdsong et al. (1988), in a study of vertebral column andmedian fin osteology, failed to place Neogobius (the sole Ponto-Cas-pian representative in their study) in any of their hypothesizedgenus-groups, whereas Pezold (1993) proposed that this genusmay belong to the subfamily Gobionellinae based on its patterningof infraorbital pores in the cephalic lateral line system (although hedid not directly examine any ‘‘neogobiin” material). Simonovic(1999) described divergence for Neogobius sensu lato and Protero-rhinus of ‘‘neogobiins” from other taxa in the subfamily Gobiinae,hypothesizing close relationship but distinctiveness between‘‘gobiins” and ‘‘neogobiins”, based on external morphometrics,osteology, and karyology, but did not examine Mesogobius. Ahneltand Duchkowitsch’s (2004) study of postcranial osteology of Prote-rorhinus placed it along with Neogobius sensu lato in the Gobiinae.
Composition of the genus Neogobius sensu lato and its interrela-tionships also have been controversial. Miller and Vasil’eva (2003)summarized information, listing the genus as comprising 14 spe-cies separated into five subgenera—N. Apollonia (containing onlyN. melanostomus), N. Neogobius (restricted to N. fluviatilis), N. Eich-waldiella (containing only N. caspius), N. Babka (= N. gymnotrache-lus), and N. Ponticola (including N. cephalargoides, kessleri, ratan,syrman, etc.). Miller (2003b) elevated the subgenus N. Chasar togeneric status (containing a single taxon—C. bathybius) based on
increased modal number of dorsal fin rays and differences in thepattern of cephalic sensory papillae; however, this distinction isquestionable and the taxon is regarded as belonging in N. Ponticola(E. Vasil’eva, personal communication). Stepien and Tumeo (2006)elevated Apollonia (including N. melanostomus and N. fluviatilis) togeneric status due to its paraphyletic position relative to the othersubgenera of Neogobius sensu lato (i.e. N. Babka and N. Ponticola),based on mitochondrial DNA cytochrome b gene sequences. More-over, the clade containing Proterorhinus, Mesogobius, Apollonia sen-su Stepien and Tumeo (2006), and Neogobius sensu lato appearedseparated from the Gobiinae tested (Gobius and Zosterissessor;Stepien and Tumeo, 2006).
Miller and Vasil’eva (2003) noted that the systematic relation-ships of the Ponto-Caspian gobies are poorly understood, providedno hypothesis for their relationships, and expressed the need for adetailed cladistic revision. Although some prior studies examinedselected morphological aspects of their relationships and system-atics, none investigated relationships of the native Ponto-Caspiangobies as a whole and only those by our laboratory and one otherused a molecular approach (i.e., partial group analyses by Dougher-ty et al., 1996; Dillon and Stepien, 2001; Stepien and Tumeo, 2006;Neilson and Stepien, 2009).
Our central goal is to analyze the systematic relationshipsamong Ponto-Caspian ‘‘neogobiin” gobies, and to illuminate someof the factors (biogeographic, evolutionary, or phylogenetic) lead-ing to their diversification that also may augment their successas invasive species. In particular, we investigate the followingquestions: (1) are the currently recognized species of ‘‘neogobiin”gobies valid (i.e., reciprocally monophyletic) taxa? (2) are the cur-rent genera valid?, and (3) how was their speciation and diversifi-cation shaped by the geologic history of the Ponto-Caspian region?We analyze the phylogenetic relationships among ‘‘neogobiin” go-bies, in comparison with gobiin relatives and outgroup taxa, usingDNA sequence data from four gene regions: the mitochondrial (mt)cytochrome (cyt) b and cytochrome oxidase c subunit I (COI) genes,and the nuclear recombination activating gene 1 (RAG1) and S7ribosomal protein intron 1 (S7). We include the 19 most prevalentmembers of �24 nominal species (Miller, 2003b; Freyhof and Nas-eka, 2007; Kovacic and Engin, 2008; Neilson and Stepien, 2009; seeTable 1) in the most complete phylogenetic study of the group.
2. Methods
2.1. Taxon sampling
Taxa analyzed in this study, collection locations, and corre-sponding GenBank accession numbers (http://www.ncbi.nlm.nih.-gov) are listed in Table 1. Specimens were collected by us andcolleagues throughout the range of the ‘‘neogobiins” within thePonto-Caspian region (Fig. 1) via small seines, beam/otter trawls,or by hook and line, and include all widely distributed and com-mon taxa. We analyze all proposed subgenera of Neogobius sensulato (= N. Babka, N. Eichwaldiella, and N. Ponticola; Miller and Vasi-l’eva, 2003) and the genera Apollonia sensu Stepien and Tumeo(2006), Proterorhinus, and Mesogobius; absent taxa either are veryrare (e.g., Mesogobius nonultimus), confined to deeper water (N.bathybius), were only recently described (N. rizensis and N. turani;Kovacic and Engin, 2008), or have extremely limited distributions(Proterorhinus tataricus; Freyhof and Naseka, 2007). We include se-ven Gobiinae outgroups (Chromogobius zebratus, Gobius auratus, G.bucchichi, G. fallax, G. niger, Pomatoschistus minutus and Zosterisess-or ophiocephalus) that range throughout the Black and Mediterra-nean Seas and are members of the hypothesized sister lineage ofthe Ponto-Caspian gobiids (Miller, 1990). We also utilize five spe-cies of tadpole gobies, including Benthophilus (B. abdurahmanovi,
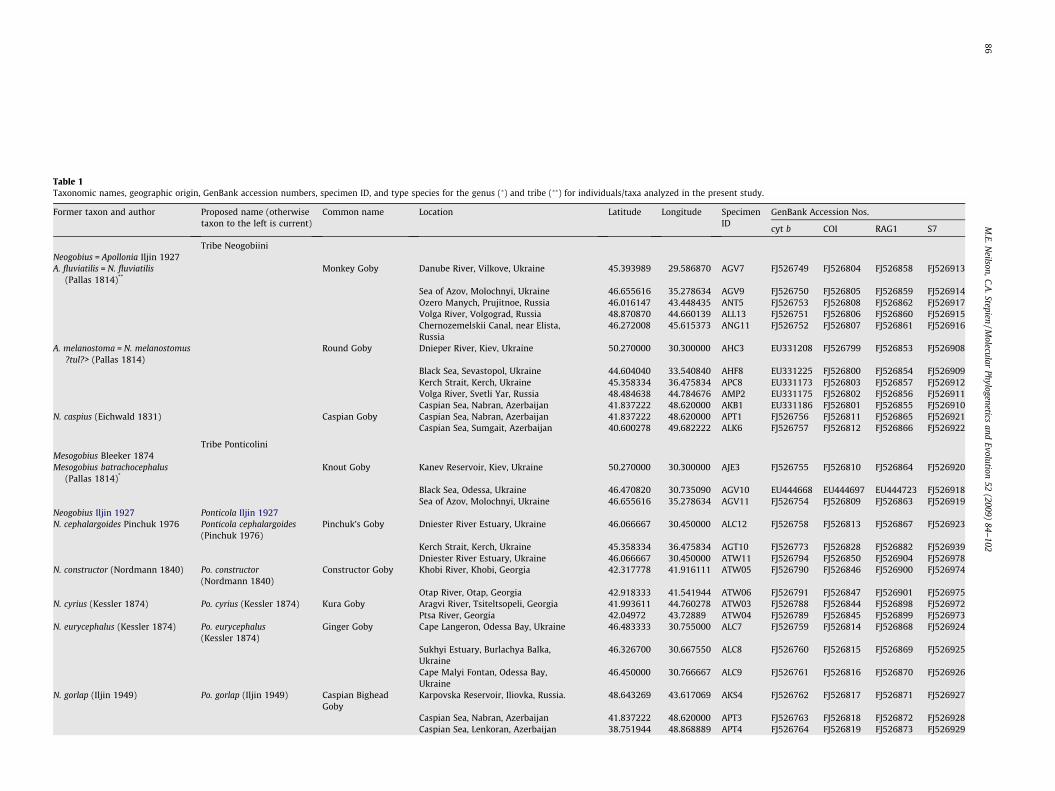
able 1axonomic names, geographic origin, GenBank accession numbers, specimen ID, and type species for the genus (�) and tribe (��) for individuals/taxa analyzed in the present study.
ormer taxon and author Proposed name (otherwisetaxon to the left is current)
Common name Location Latitude Longitude SpecimenID
GenBank Accession Nos.
cyt b COI RAG1 S7
Tribe Neogobiinieogobius = Apollonia Iljin 1927. fluviatilis = N. fluviatilis
(Pallas 1814)**Monkey Goby Danube River, Vilkove, Ukraine 45.393989 29.586870 AGV7 FJ526749 FJ526804 FJ526858 FJ526913
Sea of Azov, Molochnyi, Ukraine 46.655616 35.278634 AGV9 FJ526750 FJ526805 FJ526859 FJ526914Ozero Manych, Prujitnoe, Russia 46.016147 43.448435 ANT5 FJ526753 FJ526808 FJ526862 FJ526917Volga River, Volgograd, Russia 48.870870 44.660139 ALL13 FJ526751 FJ526806 FJ526860 FJ526915Chernozemelskii Canal, near Elista,Russia
46.272008 45.615373 ANG11 FJ526752 FJ526807 FJ526861 FJ526916
. melanostoma = N. melanostomus?tul?> (Pallas 1814)
Round Goby Dnieper River, Kiev, Ukraine 50.270000 30.300000 AHC3 EU331208 FJ526799 FJ526853 FJ526908
Black Sea, Sevastopol, Ukraine 44.604040 33.540840 AHF8 EU331225 FJ526800 FJ526854 FJ526909Kerch Strait, Kerch, Ukraine 45.358334 36.475834 APC8 EU331173 FJ526803 FJ526857 FJ526912Volga River, Svetli Yar, Russia 48.484638 44.784676 AMP2 EU331175 FJ526802 FJ526856 FJ526911Caspian Sea, Nabran, Azerbaijan 41.837222 48.620000 AKB1 EU331186 FJ526801 FJ526855 FJ526910
. caspius (Eichwald 1831) Caspian Goby Caspian Sea, Nabran, Azerbaijan 41.837222 48.620000 APT1 FJ526756 FJ526811 FJ526865 FJ526921Caspian Sea, Sumgait, Azerbaijan 40.600278 49.682222 ALK6 FJ526757 FJ526812 FJ526866 FJ526922
Tribe Ponticoliniesogobius Bleeker 1874esogobius batrachocephalus
(Pallas 1814)*Knout Goby Kanev Reservoir, Kiev, Ukraine 50.270000 30.300000 AJE3 FJ526755 FJ526810 FJ526864 FJ526920
Black Sea, Odessa, Ukraine 46.470820 30.735090 AGV10 EU444668 EU444697 EU444723 FJ526918Sea of Azov, Molochnyi, Ukraine 46.655616 35.278634 AGV11 FJ526754 FJ526809 FJ526863 FJ526919
eogobius Iljin 1927 Ponticola Iljin 1927. cephalargoides Pinchuk 1976 Ponticola cephalargoides
(Pinchuk 1976)Pinchuk’s Goby Dniester River Estuary, Ukraine 46.066667 30.450000 ALC12 FJ526758 FJ526813 FJ526867 FJ526923
Kerch Strait, Kerch, Ukraine 45.358334 36.475834 AGT10 FJ526773 FJ526828 FJ526882 FJ526939Dniester River Estuary, Ukraine 46.066667 30.450000 ATW11 FJ526794 FJ526850 FJ526904 FJ526978
. constructor (Nordmann 1840) Po. constructor(Nordmann 1840)
Constructor Goby Khobi River, Khobi, Georgia 42.317778 41.916111 ATW05 FJ526790 FJ526846 FJ526900 FJ526974
Otap River, Otap, Georgia 42.918333 41.541944 ATW06 FJ526791 FJ526847 FJ526901 FJ526975. cyrius (Kessler 1874) Po. cyrius (Kessler 1874) Kura Goby Aragvi River, Tsiteltsopeli, Georgia 41.993611 44.760278 ATW03 FJ526788 FJ526844 FJ526898 FJ526972
Ptsa River, Georgia 42.04972 43.72889 ATW04 FJ526789 FJ526845 FJ526899 FJ526973. eurycephalus (Kessler 1874) Po. eurycephalus
(Kessler 1874)Ginger Goby Cape Langeron, Odessa Bay, Ukraine 46.483333 30.755000 ALC7 FJ526759 FJ526814 FJ526868 FJ526924
Sukhyi Estuary, Burlachya Balka,Ukraine
46.326700 30.667550 ALC8 FJ526760 FJ526815 FJ526869 FJ526925
Cape Malyi Fontan, Odessa Bay,Ukraine
46.450000 30.766667 ALC9 FJ526761 FJ526816 FJ526870 FJ526926
. gorlap (Iljin 1949) Po. gorlap (Iljin 1949) Caspian BigheadGoby
Karpovska Reservoir, Iliovka, Russia. 48.643269 43.617069 AKS4 FJ526762 FJ526817 FJ526871 FJ526927
Caspian Sea, Nabran, Azerbaijan 41.837222 48.620000 APT3 FJ526763 FJ526818 FJ526872 FJ526928Caspian Sea, Lenkoran, Azerbaijan 38.751944 48.868889 APT4 FJ526764 FJ526819 FJ526873 FJ526929
86M
.E.Neilson,C.A
.Stepien/M
olecularPhylogenetics
andEvolution
52(2009)
84–102
TT
F
NA
A
N
MM
NN
N
N
N
N
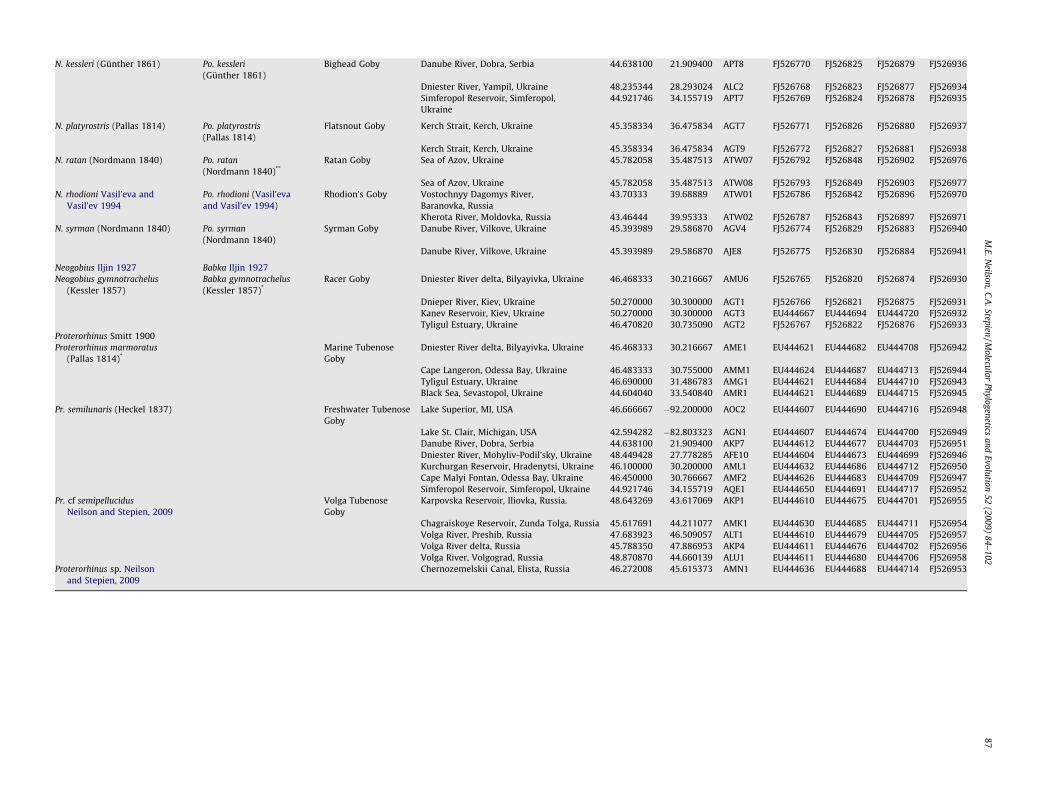
N. kessleri (Günther 1861) Po. kessleri(Günther 1861)
Bighead Goby Danube River, Dobra, Serbia 44.638100 21.909400 APT8 FJ526770 FJ526825 FJ526879 FJ526936
Dniester River, Yampil, Ukraine 48.235344 28.293024 ALC2 FJ526768 FJ526823 FJ526877 FJ526934Simferopol Reservoir, Simferopol,Ukraine
44.921746 34.155719 APT7 FJ526769 FJ526824 FJ526878 FJ526935
N. platyrostris (Pallas 1814) Po. platyrostris(Pallas 1814)
Flatsnout Goby Kerch Strait, Kerch, Ukraine 45.358334 36.475834 AGT7 FJ526771 FJ526826 FJ526880 FJ526937
Kerch Strait, Kerch, Ukraine 45.358334 36.475834 AGT9 FJ526772 FJ526827 FJ526881 FJ526938N. ratan (Nordmann 1840) Po. ratan
(Nordmann 1840)**Ratan Goby Sea of Azov, Ukraine 45.782058 35.487513 ATW07 FJ526792 FJ526848 FJ526902 FJ526976
Sea of Azov, Ukraine 45.782058 35.487513 ATW08 FJ526793 FJ526849 FJ526903 FJ526977N. rhodioni Vasil’eva and
Vasil’ev 1994Po. rhodioni (Vasil’evaand Vasil’ev 1994)
Rhodion’s Goby Vostochnyy Dagomys River,Baranovka, Russia
43.70333 39.68889 ATW01 FJ526786 FJ526842 FJ526896 FJ526970
Kherota River, Moldovka, Russia 43.46444 39.95333 ATW02 FJ526787 FJ526843 FJ526897 FJ526971N. syrman (Nordmann 1840) Po. syrman
(Nordmann 1840)Syrman Goby Danube River, Vilkove, Ukraine 45.393989 29.586870 AGV4 FJ526774 FJ526829 FJ526883 FJ526940
Danube River, Vilkove, Ukraine 45.393989 29.586870 AJE8 FJ526775 FJ526830 FJ526884 FJ526941
Neogobius Iljin 1927 Babka Iljin 1927Neogobius gymnotrachelus
(Kessler 1857)Babka gymnotrachelus(Kessler 1857)*
Racer Goby Dniester River delta, Bilyayivka, Ukraine 46.468333 30.216667 AMU6 FJ526765 FJ526820 FJ526874 FJ526930
Dnieper River, Kiev, Ukraine 50.270000 30.300000 AGT1 FJ526766 FJ526821 FJ526875 FJ526931Kanev Reservoir, Kiev, Ukraine 50.270000 30.300000 AGT3 EU444667 EU444694 EU444720 FJ526932Tyligul Estuary, Ukraine 46.470820 30.735090 AGT2 FJ526767 FJ526822 FJ526876 FJ526933
Proterorhinus Smitt 1900Proterorhinus marmoratus
(Pallas 1814)*Marine TubenoseGoby
Dniester River delta, Bilyayivka, Ukraine 46.468333 30.216667 AME1 EU444621 EU444682 EU444708 FJ526942
Cape Langeron, Odessa Bay, Ukraine 46.483333 30.755000 AMM1 EU444624 EU444687 EU444713 FJ526944Tyligul Estuary, Ukraine 46.690000 31.486783 AMG1 EU444621 EU444684 EU444710 FJ526943Black Sea, Sevastopol, Ukraine 44.604040 33.540840 AMR1 EU444621 EU444689 EU444715 FJ526945
Pr. semilunaris (Heckel 1837) Freshwater TubenoseGoby
Lake Superior, MI, USA 46.666667 �92.200000 AOC2 EU444607 EU444690 EU444716 FJ526948
Lake St. Clair, Michigan, USA 42.594282 �82.803323 AGN1 EU444607 EU444674 EU444700 FJ526949Danube River, Dobra, Serbia 44.638100 21.909400 AKP7 EU444612 EU444677 EU444703 FJ526951Dniester River, Mohyliv-Podil’sky, Ukraine 48.449428 27.778285 AFE10 EU444604 EU444673 EU444699 FJ526946Kurchurgan Reservoir, Hradenytsi, Ukraine 46.100000 30.200000 AML1 EU444632 EU444686 EU444712 FJ526950Cape Malyi Fontan, Odessa Bay, Ukraine 46.450000 30.766667 AMF2 EU444626 EU444683 EU444709 FJ526947Simferopol Reservoir, Simferopol, Ukraine 44.921746 34.155719 AQE1 EU444650 EU444691 EU444717 FJ526952
Pr. cf semipellucidusNeilson and Stepien, 2009
Volga TubenoseGoby
Karpovska Reservoir, Iliovka, Russia. 48.643269 43.617069 AKP1 EU444610 EU444675 EU444701 FJ526955
Chagraiskoye Reservoir, Zunda Tolga, Russia 45.617691 44.211077 AMK1 EU444630 EU444685 EU444711 FJ526954Volga River, Preshib, Russia 47.683923 46.509057 ALT1 EU444610 EU444679 EU444705 FJ526957Volga River delta, Russia 45.788350 47.886953 AKP4 EU444611 EU444676 EU444702 FJ526956Volga River, Volgograd, Russia 48.870870 44.660139 ALU1 EU444611 EU444680 EU444706 FJ526958
Proterorhinus sp. Neilsonand Stepien, 2009
Chernozemelskii Canal, Elista, Russia 46.272008 45.615373 AMN1 EU444636 EU444688 EU444714 FJ526953
M.E.N
eilson,C.A.Stepien
/Molecular
Phylogeneticsand
Evolution52
(2009)84–
10287

Fig. 1. Current range (excluding introduced range in North America) of nominal species of the subfamily Benthophilinae (hatched area; based on Miller, 2003), and locationsof taxa sampled in the present study. K-M = Kumo-Manych Depression.
88 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
B. granulosus, B. mahmudbejovi, and B. stellatus) and Caspiosoma cas-pium. The tadpole gobies constitute a second Ponto-Caspian ende-mic goby group that is hypothesized to be closely related to‘‘neogobiins” and their relatives (Ahnelt, 2003). Specimens werepreserved immediately following capture either in 95% ethanolfor molecular analyses or in 10% formalin (with removal of rightpectoral fin for preservation in 95% ethanol and molecular analy-ses) for future morphological analyses.
2.2. DNA analysis
Genomic DNA was isolated from fin clips or caudal muscle tis-sue using a Qiagen DNEasy tissue kit (Valencia, CA) following man-ufacturer’s protocols. Two mitochondrial genes (cyt b and COI) andtwo nuclear genes (RAG1 and S7) were amplified via the polymer-ase chain reaction (PCR) using the following primers: cyt b—AJG15(Akihito et al., 2000), H15343goby (Neilson and Stepien, 2009),L15162goby (Neilson and Stepien, 2009), and H5 (Akihito et al.,2000); COI—L6486, H7127, L7057, and H7696 (Thacker, 2003);RAG1—RAG1F1 (López et al., 2004), RAG1-R811goby (Neilson andStepien, 2009), RAG1-F709 (50-CTTATGTCTGCACGCTCTGC-30, thisstudy), and RAG1R1 (López et al., 2004); and S7—S7RPEX1F andS7RPEX2R (Chow and Hazama, 1998). PCR amplifications were per-formed in 25 lL volumes containing 10 mM Tris–HCl pH 8.3,50 mM KCl, 1.5 mM MgCl2 (2.5 mM for COI), 0.001% (w/v) gelatin,200 lM each dNTP, 0.5 lM each primer, 1.5 units of Taq polymer-ase, and �100 ng (1–3 lL) of template DNA. The PCR profile for cytb and RAG1 included an initial denaturation of 94 �C for 2 min, 40cycles of 94 �C for 45 s, gene specific annealing temperature (cytb—52�; RAG1—50�; S7—60�) for 30 s, and 72 �C for 60 s, with a finalextension of 72 �C for 3 min. The cycling profile for COI included aninitial denaturation of 94 �C for 3 min, 35 cycles of 94 �C for 30 s,53 �C for 30 s, and 72 �C for 60 s, with a final extension of 72 �C
for 2 min. The profile for S7 included an initial denaturation of94 �C for 3 min, 40 cycles of 94 �C for 60 s, 60 �C for 45 s, and72 �C for 120 s, with a final extension of 72 �C for 5 min. PCR reac-tions were checked on 1% agarose gels stained with ethidium bro-mide, and excess primers and unincorporated nucleotides wereremoved from successful reactions with spin column purificationkits (QIAquick PCR Purification Kit, Qiagen; or QuickStep 2 PCRPurification Kit, Edge Biosystems, Gaithersburg, MD).
For all genes, amplicons were sequenced in both directionsusing dye-labeled terminators and PCR primers, and resolved onan ABI 3730 (Applied Biosystems, Foster City, CA) genetic analyzerat the Cornell University Life Sciences Core Laboratories Center.Forward and reverse sequences for each gene per individual werealigned in our laboratory, a contiguous sequence was created withBioEdit (Hall, 1999), and sequences for each gene were globallyaligned using Clustal X (cyt b, COI, RAG1; Larkin et al., 2007) orT-Coffee (S7; Notredame et al., 2000).
2.3. Phylogenetic analyses
We used parsimony, likelihood, and Bayesian approaches toreconstruct phylogenies, with PAUP* v4.0b10 (Swofford, 2003),PhyML v2.4.4 (Guindon and Gascuel, 2003) and MrBayes v3.1(Ronquist and Huelsenbeck, 2003), respectively. Parsimony analy-ses were performed using unweighted heuristic searches, withstarting trees obtained by random addition (100 replications) hold-ing 10 trees per replicate, and tree-bisection-reconnection branchswapping. Branch support was calculated for the inferred branchesvia non-parametric bootstrapping (2000 replications) (Table 2).
For likelihood and Bayesian analyses, ModelTest v3.7 (Posadaand Crandall, 1998) was employed to determine the simplestbest-fit model of evolution for each gene under the Akaike infor-mation criterion. For the cyt b data, the best-fit model was
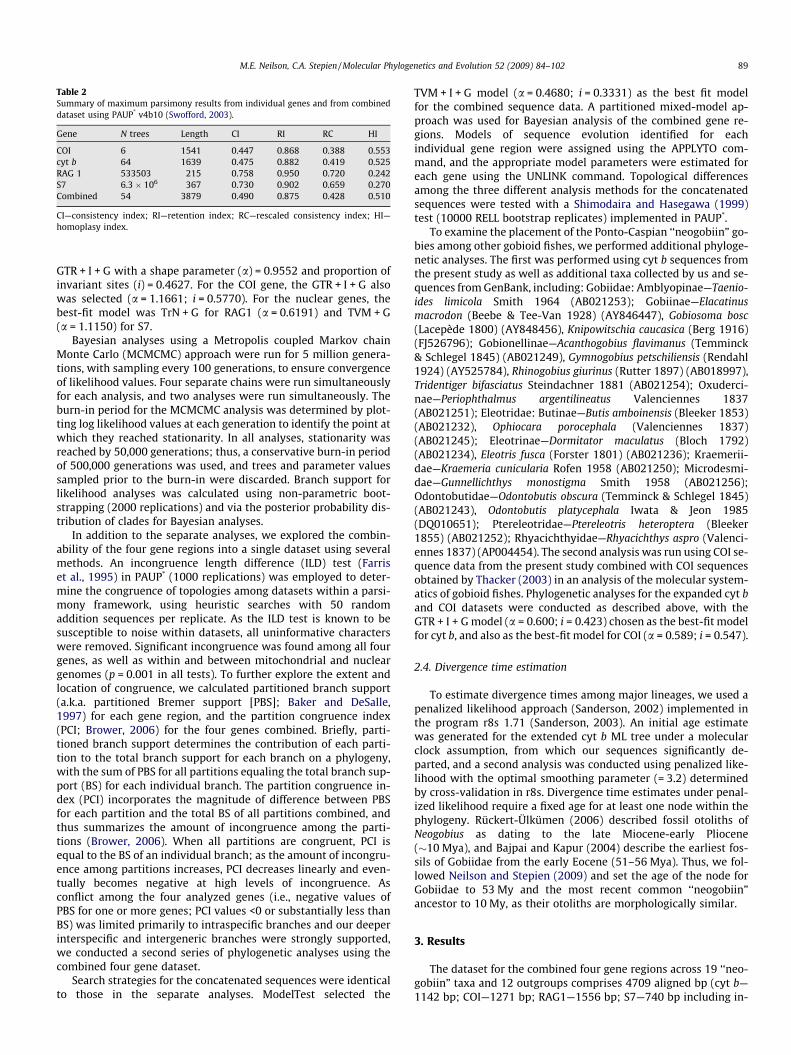
Table 2Summary of maximum parsimony results from individual genes and from combineddataset using PAUP* v4b10 (Swofford, 2003).
Gene N trees Length CI RI RC HI
COI 6 1541 0.447 0.868 0.388 0.553cyt b 64 1639 0.475 0.882 0.419 0.525RAG 1 533503 215 0.758 0.950 0.720 0.242S7 6.3 � 106 367 0.730 0.902 0.659 0.270Combined 54 3879 0.490 0.875 0.428 0.510
CI—consistency index; RI—retention index; RC—rescaled consistency index; HI—homoplasy index.
M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 89
GTR + I + G with a shape parameter (a) = 0.9552 and proportion ofinvariant sites (i) = 0.4627. For the COI gene, the GTR + I + G alsowas selected (a = 1.1661; i = 0.5770). For the nuclear genes, thebest-fit model was TrN + G for RAG1 (a = 0.6191) and TVM + G(a = 1.1150) for S7.
Bayesian analyses using a Metropolis coupled Markov chainMonte Carlo (MCMCMC) approach were run for 5 million genera-tions, with sampling every 100 generations, to ensure convergenceof likelihood values. Four separate chains were run simultaneouslyfor each analysis, and two analyses were run simultaneously. Theburn-in period for the MCMCMC analysis was determined by plot-ting log likelihood values at each generation to identify the point atwhich they reached stationarity. In all analyses, stationarity wasreached by 50,000 generations; thus, a conservative burn-in periodof 500,000 generations was used, and trees and parameter valuessampled prior to the burn-in were discarded. Branch support forlikelihood analyses was calculated using non-parametric boot-strapping (2000 replications) and via the posterior probability dis-tribution of clades for Bayesian analyses.
In addition to the separate analyses, we explored the combin-ability of the four gene regions into a single dataset using severalmethods. An incongruence length difference (ILD) test (Farriset al., 1995) in PAUP* (1000 replications) was employed to deter-mine the congruence of topologies among datasets within a parsi-mony framework, using heuristic searches with 50 randomaddition sequences per replicate. As the ILD test is known to besusceptible to noise within datasets, all uninformative characterswere removed. Significant incongruence was found among all fourgenes, as well as within and between mitochondrial and nucleargenomes (p = 0.001 in all tests). To further explore the extent andlocation of congruence, we calculated partitioned branch support(a.k.a. partitioned Bremer support [PBS]; Baker and DeSalle,1997) for each gene region, and the partition congruence index(PCI; Brower, 2006) for the four genes combined. Briefly, parti-tioned branch support determines the contribution of each parti-tion to the total branch support for each branch on a phylogeny,with the sum of PBS for all partitions equaling the total branch sup-port (BS) for each individual branch. The partition congruence in-dex (PCI) incorporates the magnitude of difference between PBSfor each partition and the total BS of all partitions combined, andthus summarizes the amount of incongruence among the parti-tions (Brower, 2006). When all partitions are congruent, PCI isequal to the BS of an individual branch; as the amount of incongru-ence among partitions increases, PCI decreases linearly and even-tually becomes negative at high levels of incongruence. Asconflict among the four analyzed genes (i.e., negative values ofPBS for one or more genes; PCI values <0 or substantially less thanBS) was limited primarily to intraspecific branches and our deeperinterspecific and intergeneric branches were strongly supported,we conducted a second series of phylogenetic analyses using thecombined four gene dataset.
Search strategies for the concatenated sequences were identicalto those in the separate analyses. ModelTest selected the
TVM + I + G model (a = 0.4680; i = 0.3331) as the best fit modelfor the combined sequence data. A partitioned mixed-model ap-proach was used for Bayesian analysis of the combined gene re-gions. Models of sequence evolution identified for eachindividual gene region were assigned using the APPLYTO com-mand, and the appropriate model parameters were estimated foreach gene using the UNLINK command. Topological differencesamong the three different analysis methods for the concatenatedsequences were tested with a Shimodaira and Hasegawa (1999)test (10000 RELL bootstrap replicates) implemented in PAUP*.
To examine the placement of the Ponto-Caspian ‘‘neogobiin” go-bies among other gobioid fishes, we performed additional phyloge-netic analyses. The first was performed using cyt b sequences fromthe present study as well as additional taxa collected by us and se-quences from GenBank, including: Gobiidae: Amblyopinae—Taenio-ides limicola Smith 1964 (AB021253); Gobiinae—Elacatinusmacrodon (Beebe & Tee-Van 1928) (AY846447), Gobiosoma bosc(Lacepède 1800) (AY848456), Knipowitschia caucasica (Berg 1916)(FJ526796); Gobionellinae—Acanthogobius flavimanus (Temminck& Schlegel 1845) (AB021249), Gymnogobius petschiliensis (Rendahl1924) (AY525784), Rhinogobius giurinus (Rutter 1897) (AB018997),Tridentiger bifasciatus Steindachner 1881 (AB021254); Oxuderci-nae—Periophthalmus argentilineatus Valenciennes 1837(AB021251); Eleotridae: Butinae—Butis amboinensis (Bleeker 1853)(AB021232), Ophiocara porocephala (Valenciennes 1837)(AB021245); Eleotrinae—Dormitator maculatus (Bloch 1792)(AB021234), Eleotris fusca (Forster 1801) (AB021236); Kraemerii-dae—Kraemeria cunicularia Rofen 1958 (AB021250); Microdesmi-dae—Gunnellichthys monostigma Smith 1958 (AB021256);Odontobutidae—Odontobutis obscura (Temminck & Schlegel 1845)(AB021243), Odontobutis platycephala Iwata & Jeon 1985(DQ010651); Ptereleotridae—Ptereleotris heteroptera (Bleeker1855) (AB021252); Rhyacichthyidae—Rhyacichthys aspro (Valenci-ennes 1837) (AP004454). The second analysis was run using COI se-quence data from the present study combined with COI sequencesobtained by Thacker (2003) in an analysis of the molecular system-atics of gobioid fishes. Phylogenetic analyses for the expanded cyt band COI datasets were conducted as described above, with theGTR + I + G model (a = 0.600; i = 0.423) chosen as the best-fit modelfor cyt b, and also as the best-fit model for COI (a = 0.589; i = 0.547).
2.4. Divergence time estimation
To estimate divergence times among major lineages, we used apenalized likelihood approach (Sanderson, 2002) implemented inthe program r8s 1.71 (Sanderson, 2003). An initial age estimatewas generated for the extended cyt b ML tree under a molecularclock assumption, from which our sequences significantly de-parted, and a second analysis was conducted using penalized like-lihood with the optimal smoothing parameter (= 3.2) determinedby cross-validation in r8s. Divergence time estimates under penal-ized likelihood require a fixed age for at least one node within thephylogeny. Rückert-Ülkümen (2006) described fossil otoliths ofNeogobius as dating to the late Miocene-early Pliocene(�10 Mya), and Bajpai and Kapur (2004) describe the earliest fos-sils of Gobiidae from the early Eocene (51–56 Mya). Thus, we fol-lowed Neilson and Stepien (2009) and set the age of the node forGobiidae to 53 My and the most recent common ‘‘neogobiin”ancestor to 10 My, as their otoliths are morphologically similar.
3. Results
The dataset for the combined four gene regions across 19 ‘‘neo-gobiin” taxa and 12 outgroups comprises 4709 aligned bp (cyt b—1142 bp; COI—1271 bp; RAG1—1556 bp; S7—740 bp including in-
90 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
dels). GenBank accession numbers are EU331173, EU331175,EU331186, EU331208, EU331225, EU444604, EU444607,EU444610–EU444612, EU444621, EU444624, EU444626,EU444630, EU444632, EU444636, EU444650, EU444667,EU444668, EU444670, and FJ526747–FJ526795 for cyt b;EU444673–EU444677, EU444679–EU444680, EU444682–EU444691, EU444694, EU444697–EU444698, and FJ526797–FJ526850 for COI; EU444699–EU444700, EU444702–EU444703,EU444705–EU444706, EU444708–EU444717, EU444720,
Fig. 2. Maximum likelihood phylogeny (PhyML; Guindon and Gascuel, 2003) of the subfaNumbers at nodes indicate likelihood bootstrap support (2000 pseudoreplications), with
EU444723–EU444724, and FJ526851–FJ526904 for RAG1; andFJ526905–FJ526978 for S7 (Table 1, Appendix A; 61 individuals).Base composition is stationary across taxa for each gene(v2 > 31.6; df = 180; p > 0.29).
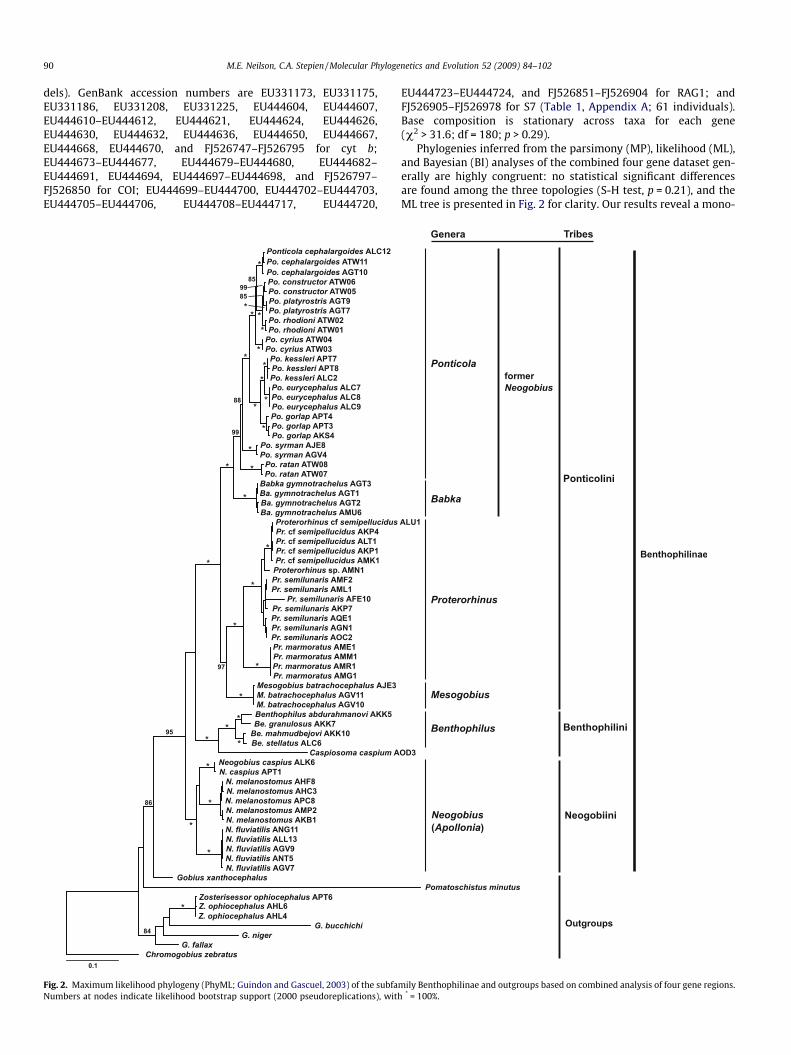
Phylogenies inferred from the parsimony (MP), likelihood (ML),and Bayesian (BI) analyses of the combined four gene dataset gen-erally are highly congruent: no statistical significant differencesare found among the three topologies (S-H test, p = 0.21), and theML tree is presented in Fig. 2 for clarity. Our results reveal a mono-
mily Benthophilinae and outgroups based on combined analysis of four gene regions.* = 100%.
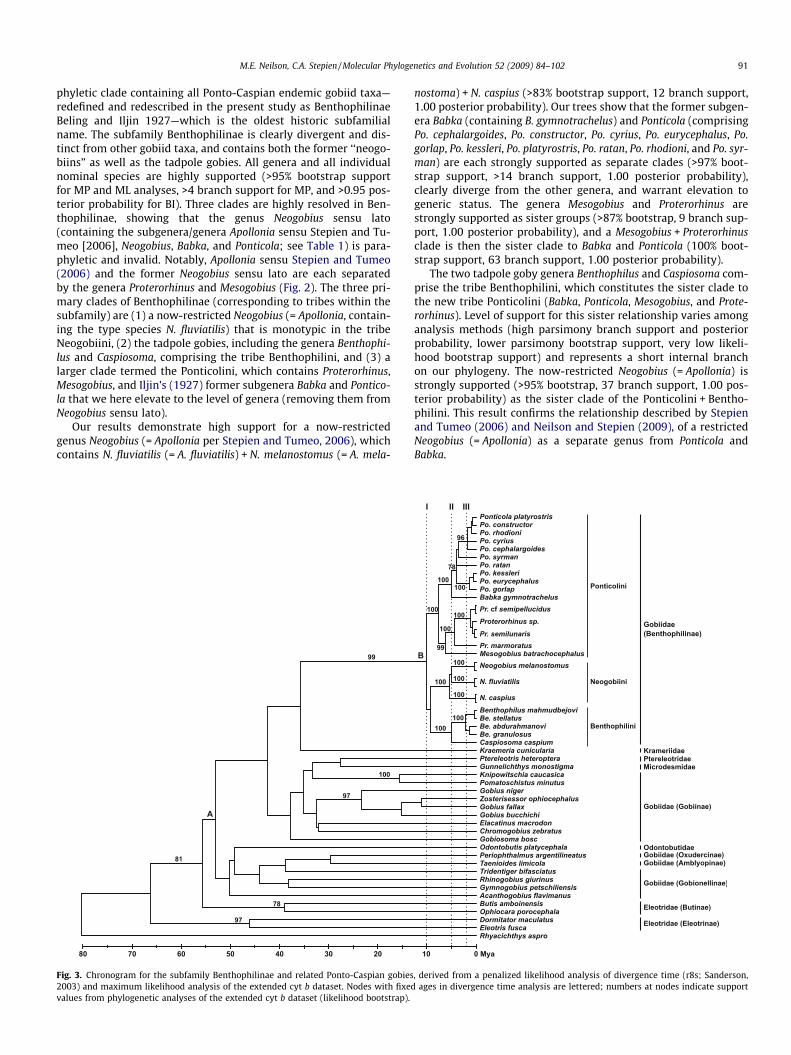
M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 91
phyletic clade containing all Ponto-Caspian endemic gobiid taxa—redefined and redescribed in the present study as BenthophilinaeBeling and Iljin 1927—which is the oldest historic subfamilialname. The subfamily Benthophilinae is clearly divergent and dis-tinct from other gobiid taxa, and contains both the former ‘‘neogo-biins” as well as the tadpole gobies. All genera and all individualnominal species are highly supported (>95% bootstrap supportfor MP and ML analyses, >4 branch support for MP, and >0.95 pos-terior probability for BI). Three clades are highly resolved in Ben-thophilinae, showing that the genus Neogobius sensu lato(containing the subgenera/genera Apollonia sensu Stepien and Tu-meo [2006], Neogobius, Babka, and Ponticola; see Table 1) is para-phyletic and invalid. Notably, Apollonia sensu Stepien and Tumeo(2006) and the former Neogobius sensu lato are each separatedby the genera Proterorhinus and Mesogobius (Fig. 2). The three pri-mary clades of Benthophilinae (corresponding to tribes within thesubfamily) are (1) a now-restricted Neogobius (= Apollonia, contain-ing the type species N. fluviatilis) that is monotypic in the tribeNeogobiini, (2) the tadpole gobies, including the genera Benthophi-lus and Caspiosoma, comprising the tribe Benthophilini, and (3) alarger clade termed the Ponticolini, which contains Proterorhinus,Mesogobius, and Iljin’s (1927) former subgenera Babka and Pontico-la that we here elevate to the level of genera (removing them fromNeogobius sensu lato).
Our results demonstrate high support for a now-restrictedgenus Neogobius (= Apollonia per Stepien and Tumeo, 2006), whichcontains N. fluviatilis (= A. fluviatilis) + N. melanostomus (= A. mela-
Fig. 3. Chronogram for the subfamily Benthophilinae and related Ponto-Caspian gobies2003) and maximum likelihood analysis of the extended cyt b dataset. Nodes with fixedvalues from phylogenetic analyses of the extended cyt b dataset (likelihood bootstrap).
nostoma) + N. caspius (>83% bootstrap support, 12 branch support,1.00 posterior probability). Our trees show that the former subgen-era Babka (containing B. gymnotrachelus) and Ponticola (comprisingPo. cephalargoides, Po. constructor, Po. cyrius, Po. eurycephalus, Po.gorlap, Po. kessleri, Po. platyrostris, Po. ratan, Po. rhodioni, and Po. syr-man) are each strongly supported as separate clades (>97% boot-strap support, >14 branch support, 1.00 posterior probability),clearly diverge from the other genera, and warrant elevation togeneric status. The genera Mesogobius and Proterorhinus arestrongly supported as sister groups (>87% bootstrap, 9 branch sup-port, 1.00 posterior probability), and a Mesogobius + Proterorhinusclade is then the sister clade to Babka and Ponticola (100% boot-strap support, 63 branch support, 1.00 posterior probability).
The two tadpole goby genera Benthophilus and Caspiosoma com-prise the tribe Benthophilini, which constitutes the sister clade tothe new tribe Ponticolini (Babka, Ponticola, Mesogobius, and Prote-rorhinus). Level of support for this sister relationship varies amonganalysis methods (high parsimony branch support and posteriorprobability, lower parsimony bootstrap support, very low likeli-hood bootstrap support) and represents a short internal branchon our phylogeny. The now-restricted Neogobius (= Apollonia) isstrongly supported (>95% bootstrap, 37 branch support, 1.00 pos-terior probability) as the sister clade of the Ponticolini + Bentho-philini. This result confirms the relationship described by Stepienand Tumeo (2006) and Neilson and Stepien (2009), of a restrictedNeogobius (= Apollonia) as a separate genus from Ponticola andBabka.
, derived from a penalized likelihood analysis of divergence time (r8s; Sanderson,ages in divergence time analysis are lettered; numbers at nodes indicate support

92 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
Results of extended cyt b analyses are very similar to the com-bined four gene analyses (ML topology and support values inFig. 3), with slight placement differences for Ponticola ratan andPo. syrman, and in grouping the Benthophilini (Benthophilus + Cas-piosoma) with the Neogobiini rather than with the Ponticolini.There is strong support for the subfamily Benthophilinae as a cladedistinct from the remainder of the Gobiidae, as well as for genericseparation of Neogobius (= Apollonia), Babka, and Ponticola. Diver-gence time estimates from the extended cyt b ML tree are reportedin Table 3. Separation of the subfamily Benthophilinae from otherGobiinae taxa occurred �39 million years ago (Mya).
The genera Neogobius, Mesogobius, Proterorhinus, Babka, Pontico-la, Benthophilus, and Caspiosoma) have similar dates of origin, rang-ing from 4.29–6.25 Mya. Proterorhinus diverged from Mesogobius�6.18 Mya. Among the remaining genera, the three Neogobius(= Apollonia) species are separated by an estimated 5.47 My, Prote-rorhinus species by �4.29 My, and Babka and Ponticola by�5.08 My. Within Ponticola, a split (�4.07 My) into two primaryclades occurred: a ‘‘kessleri” group containing Po. eurycephalus,Po. gorlap, and Po. kessleri, versus a clade containing the remainingPonticola species. The ‘‘kessleri” group began diverging from oneanother �1.37 Mya. Within the second Ponticola clade, Po. ratanand Po. syrman branched off soon after separating from the kesslerigroup; and the remaining species (Po. cephalargoides, Po. construc-tor, Po. cyrius, Po. platyrostris, and Po. rhodioni) comprise a second‘‘platyrostris” group that radiated from one another �1.82 Mya.
Extended COI analysis trees (Fig. 4) are very similar to the com-bined four gene analyses, and also are generally congruent with theextended cyt b trees; with all identifying Benthophilinae as a dis-tinct subfamily from other gobiin taxa. The extended COI and ex-tended cyt b analyses vary in degree of separation between theBenthophilinae and members of the putative subfamily Gobiinae,as well as in designating its sister taxa. The extended COI analysisdepicts a clade containing Gobius + Zosterisessor as the sister cladeof Benthophilinae, which are more distantly related in the ex-tended cyt b analysis. Both extended analyses have similar supportvalues; including high support for the subfamily Benthophilinaeand its component taxa, and low support for most deeper gobiidbranches outside of the Benthophilinae (Figs. 3 and 4).
Table 3Divergence times for major lineages/nodes within phylogeny of the subfamily Benthophlikelihood in r8s (Sanderson, 2003). Nodes representing fixed ages (A and B) and major ge
Node Ea
MRCA of Gobiidae (A; fixed at 53.00) 5MRCA of Neogobius sensu stricto, Babka + Ponticola, and Proterorhinus (B; fixed at
10.00)1
Tribe Neogobiini (Neogobius) + Tribe Benthophilini ‘‘tadpole gobies”(Benthophilus + Caspiosoma)
Tribe Ponticolini (Babka + Ponticola + Mesogobius + Proterorhinus)
Proterorhinus + Mesogobius
Tribe Neogobiini = Neogobius sensu stricto (N. fluviatilis, N. melanostomus, and N.caspius)
Babka (B. gymnotrachelus) + Ponticola (Po. cepharlargoides, Po. constructor, Po. cyrius,Po. eurycephalus, Po. gorlap, Po. kessleri, Po. platyrostris, Po. ratan, Po. rhodioni, andPo. syrman)
Tribe Benthophilini ‘‘tadpole gobies” (Benthophilus + Caspiosoma)marine and freshwater tubenose gobies ProterorhinusPonticolaBenthophilusPonticola ‘‘platyrostris group” (Po. cephalargoides, Po. constructor, Po. cyrius, Po.
platyrostris, and Po. rhodioni)Ponticola ‘‘kessleri group” (Po. eurycephalus, Po. gorlap, and Po. kessleri)freshwater Proterorhinus
Phylogenies inferred from the parsimony (MP), likelihood(ML), and Bayesian (BI) analyses primarily differ only in thebranching order of individual specimens within species. A singleexception for high congruent support in our trees occurs in thefreshwater tubenose goby Proterorhinus semilunaris. The evolu-tionary and phylogeographic history of Proterorhinus recentlyevaluated by us (Neilson and Stepien, 2009) is very similar tothe relationships seen in the present study except for placementof a single individual, Proterorhinus sp. AMN1. In Neilson andStepien (2009) and the ML and BI analyses here, AMN1 (collectedin the Kumo-Manych Depression—a lowland between the RussianPlain and the northern foothills of the Caucasus Mountains; seeFig. 1) groups closely with Proterorhinus from the Caspian Sea/Volga River clade (Pr. cf semipellucidus); whereas in our currentMP analysis, it clusters with the Black Sea freshwater species(Pr. semilunaris). In addition, Pr. semilunaris and Pr. cf semipelluci-dus are not distinguished as clades in our present MP analysis(Fig. 2); and instead form a single large clade with Pr. semilunarislocated basally. This difference likely results from the addition ofnuclear S7 intron data, which were not previously sequenced(Neilson and Stepien, 2009).
4. Discussion
Phylogenetic analysis of our molecular data for 19 Ponto-Cas-pian ‘‘neogobiin” species yields a robust phylogeny that generallyagrees with prior molecular and morphological data, yet deviatesfrom some earlier morphological hypotheses. Our results revealthat the ‘‘neogobiin” and tadpole ‘‘benthophilin” gobies togethercomprise a clade that markedly diverges from other gobiid taxa.We thus resurrect and redescribe the subfamily Benthophilinae Il-jin 1927 to encompass three tribes; Neogobiini (Neogobius = Apol-lonia), Ponticolini (containing Babka, Mesogobius, Ponticola, andProterorhinus), and Benthophilini (the tadpole gobies Benthophilus,Caspisoma, etc.) Our results support the primary findings of Stepienand Tumeo (2006; findings #1–3) and Neilson and Stepien (2009;#2–3) Beling and, which (1) distinguish a restricted genus Neogo-bius (= Apollonia) comprising the monotypic tribe Neogobiini thatis differentiated from the remainder of the Benthophilinae (justify-
ilinae, showing ages estimated for the extended cyt b tree (Fig. 3) using penalizedologic events in the Ponto-Caspian basin (I–III) are indicated on Fig. 3.
stimatedge (Mya)
Major geologic event in Ponto-Caspian basin (Reid and Orlova, 2002)
3.000.00 Separation of Ponto-Caspian and Pannonian basins (�12.5–10 Mya;
I)9.18
7.58 Intermittent connections with World Ocean, with introgression ofmarine fauna (8.3–6.4 Mya)
6.25 Brief reconnection with Pannonian basin and immigration ofendemic Pannonian fauna (6.4–5.8 Mya)
5.47 Separation of Black and Caspian basins (5.8–5.0 Mya—coincides withMessinian salinity crisis in paleo-Mediterranean and Black Seabasins; II)
5.08 Single, large lake in southern Caspian basin (5.2–2.5 Mya, II)
5.044.294.072.17 Black and Caspian basins connected via Kumo-Manych depression,
faunal exchange between basins; glacially-driven fluctuations inwater levels (2.6–0.7 Mya; III)
1.82
1.371.18
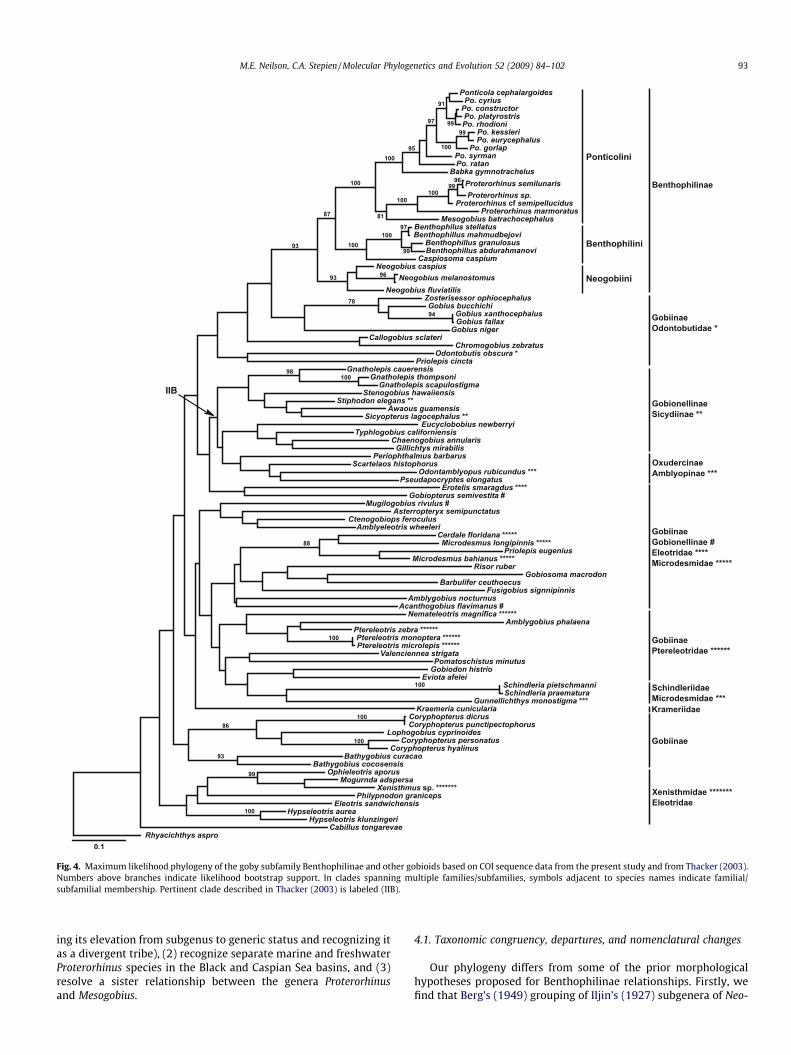
Fig. 4. Maximum likelihood phylogeny of the goby subfamily Benthophilinae and other gobioids based on COI sequence data from the present study and from Thacker (2003).Numbers above branches indicate likelihood bootstrap support. In clades spanning multiple families/subfamilies, symbols adjacent to species names indicate familial/subfamilial membership. Pertinent clade described in Thacker (2003) is labeled (IIB).
M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 93
ing its elevation from subgenus to generic status and recognizing itas a divergent tribe), (2) recognize separate marine and freshwaterProterorhinus species in the Black and Caspian Sea basins, and (3)resolve a sister relationship between the genera Proterorhinusand Mesogobius.
4.1. Taxonomic congruency, departures, and nomenclatural changes
Our phylogeny differs from some of the prior morphologicalhypotheses proposed for Benthophilinae relationships. Firstly, wefind that Berg’s (1949) grouping of Iljin’s (1927) subgenera of Neo-

94 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
gobius (Neogobius, Apollonia, Babka, and Ponticola) into a singlegenus is paraphyletic and invalid. Our trees demonstrate clear phy-logenetic separation of a restricted genus Neogobius (= Apollonia) inthe tribe Neogobiini from the tribe Ponticolini; which includes thenewly elevated genera Babka and Ponticola, along with the generaProterorhinus and Mesogobius. Berg (1949), as first reviser, selectedNeogobius sensu lato as the generic name, and thereby N. fluviatilisbecame the type species for the genus Neogobius sensu lato. Sincethe genus name Neogobius must remain as priority for the cladecontaining N. fluviatilis (W. Eschmeyer, personal communication),we hereby synonymize the generic name Apollonia with Neogobius.
In addition, we resolve Neogobius caspius (Eichwald 1831) asbelonging to the now-restricted genus Neogobius (N. fluviatilisand N. melanostomus; see Stepien and Tumeo, 2006), which com-prises a strongly-supported clade (Figs. 2–4). Neogobius caspiusonce was placed in a separate (monotypic) subgenus Eichwaldiella(Whitley 1930), and later incorrectly was moved (W. Eschmeyer,personal communication) without justification to a monotypicsubgenus Neogobius by Miller and Vasil’eva (2003). Its position rel-ative to other Neogobius/Apollonia species thus was in questionprior to our study. Pinchuk (1991) suggested that N. caspiusgrouped together with N. fluviatilis + N. melanostomus on the basisof mouth size (small in the three species vs. large for taxa now con-tained in Ponticola and Babka) as well as tooth size distribution onthe dentary, but regarded N. caspius as distinct in the forward posi-tion of its anterior and posterior nostrils. Miller and Vasil’eva(2003), in describing Iljin’s (1927) subgenera, presented the diag-nostic character of an absent metapterygoid/quadrate bridge asuniting N. fluviatilis and N. melanostomus. We discern that themetapterygoid/quadrate bridge likewise is absent in N. caspius,and thus is synapomorphic for our restricted Neogobius clade.Strong support of our molecular data for this restricted Neogobius(= Apollonia) clade (N. caspius + N. fluviatilis + N. melanostomus),combined with several morphological similarities and the nomen-clatural changes described above, leads to our redefinition of a re-stricted Neogobius (in synonymy with Apollonia) in the tribeNeogobiini.
The molecular phylogenies presented here, as well as in Stepienand Tumeo (2006) and Neilson and Stepien (2009), are congruentin identifying large separation between Ponticola/Babka and Neogo-bius sensu stricto/Apollonia. In addition, pronounced genetic diver-gence between Babka and Ponticola (subgenera delineated by Iljin[1927]), along with their morphological separation and autapo-morphies (Miller and Vasil’eva 2003), supports their elevation togeneric level. Our phylogenetic trees reflect this new nomencla-ture. Our results show that the monotypic Babka contains the racergoby B. gymnotrachelus and is the sister species to a strongly-sup-ported monophyletic Ponticola clade, which diverged �4.51–4.86 Mya (Table 3). The Ponticola + Babka clade is the sister groupof Mesogobius + Proterorhinus, with high support; which togetherform the tribe Ponticolini. Historically, Babka once was hypothe-sized to be closely related to the knout goby Mesogobius batracho-cephalus (Pallas, 1814) based on early studies of morphology (Berg,1949) and protein electrophoresis (Dobrovolov et al., 1995),although Vasil’ev and Grigoryan (1992) concluded that the twowere not congeners based on chromosomal morphology; whichis further confirmed by their generic separation in our study.
Departures of our phylogeny from former systematic hypothe-ses occur for the clade Ponticola. Notably, Vasil’eva et al. (1993)suggested two distinct groups within Ponticola based on cranialmorphometry: one containing Po. gorlap (Iljin 1949), Po. kessleri,Po. ratan, and Po. syrman (Nordmann 1840), and the other encom-passing Po. cephalargoides, Po. eurycephalus (Kessler 1874), Po.platyrostris, and the Caucasian freshwater gobies [Po. constructor(Nordmann 1840), Po. cyrius (Kessler 1874), and Po. rhodioni (Vasi-l’eva and Vasil’ev 1994)]. Based on our molecular data, Po. ratan is
basal to all other Ponticola species; whose branching order differsslightly from that suggested by Vasil’eva et al. (1993). In addition,we resolve two species groups different than those proposed byVasil’eva et al. (1993): the first group contains Po. eurycephalus,Po. gorlap, and Po. kessleri (designated as the ‘‘kessleri” group in Ta-ble 3); the second comprises Po. cephalargoides, Po. constructor, Po.cyrius, Po. platyrostris, and Po. rhodioni (the ‘‘platyrostris” group inTable 3).
4.2. Relationships among Ponto-Caspian endemic gobiid groups
Inclusion of the tadpole gobies Benthophilus and Caspiosoma is anovel feature of our molecular phylogeny. Although some recentstudies have considered their osteology and taxonomy (e.g., Ahneltet al., 2000; Ahnelt, 2003; Boldyrev and Bogutskaya, 2007), none ofthe recent larger-scale studies of goby morphological (Birdsonget al., 1988; Pezold, 1993) or molecular (Akihito et al., 2000; Thack-er, 2003) systematics included any tadpole gobies (e.g., Anatiro-strum, Benthophiloides, Benthophilus, and Caspiosoma) or‘‘neogobiin” taxa (Babka, Mesogobius, Neogobius, Ponticola, and Pro-terorhinus). Although the ‘‘neogobiins” and tadpole gobies wereposited to be sister groups based on shared geography and similarpostcranial osteology (Ahnelt, 2003), ours is the first study toincorporate both in a comprehensive phylogenetic analysis.
All of our analyses strongly support a monophyletic clade com-prising the ‘‘neogobiins” and tadpole gobies (Figs. 2–4), for whichwe resurrect the historic name Benthophilinae (Beling and Iljin1927), as a subfamily of Gobiidae. Subfamily Benthophilinae con-tains three distinctive and divergent clades—designated here asthe tribes Benthophilini (the tadpole gobies), Neogobiini (mono-typic for the genus Neogobius), and Ponticolini (Babka, Mesogobius,Ponticola, and Proterorhinus). Placement of the tribe Benthophiliniis inconsistent among some of our trees. All combined sequencedata analyses resolve Benthophilini as the sister clade to Pontico-lini, however this relationship has mixed support (1.00 posteriorprobability, <84% bootstrap support, 13 branch support) and rela-tively short branch lengths. In the extended cyt b analysis Bentho-philini is found as the sister clade to Neogobiini (but with shortbranch length and no support), whereas in the extended COI anal-ysis Benthophilini again groups with Ponticolini. Additional genet-ic, morphological, and taxonomic sampling of Benthophilini isrecommended to further resolve its relationships. One potentialindication of a close relationship between Neogobiini and Bentho-philini is their shared loss of the metapterygoid bridge, in contrastto its presence in the Ponticolini. However, presence of the metap-terygoid bridge is widely considered a pleisiomorphic trait withingobiids (Miller, 1973).
4.3. Higher taxonomic placement of the subfamily Benthophilinae
Relationship of the newly-defined Benthophilinae to other gobi-ids was contentious prior to our study. Morphological studiesplaced members of the group either in the Gobiinae (Ahnelt,2003), divergent from other Gobiinae taxa but related to them(Simonovic, 1999), or in the Gobionellinae (Pezold, 1993). Our ex-tended cyt b and COI analyses further resolve this question. In bothanalyses, the Benthophilinae comprises a true taxon, removedfrom all other gobiin taxa. In addition, both the cyt b and COI anal-yses suggest a non-monophyletic Gobiinae similar to that found byThacker (2003) using the mt COI, ND1, and ND2 genes (who did notexamine any ‘‘neogobiins”). Analysis of the COI data from our studycombined with Thacker’s (2003) COI data (Fig. 4) yields a similarresult to our extended cyt b dataset, depicting a clade comprisingthe Benthophilinae plus Gobius + Zosterisessor (Gobiinae) nestingas the sister group of one of Thacker’s (2003) Gobiinae clades(IIB). Our results yield high support for the subfamily Benthophil-

M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 95
inae and its three component tribes, which are the focus of ourstudy, and show less resolution for deeper branches separatingother gobioid families. Further research will be necessary to fullyidentify the arrangement of the Benthophilinae within the high-er-order framework of gobioid systematics, which is not our focus.
4.4. Biogeographic patterns
Our analysis of divergence times among lineages of Benthophil-inae is in general concordance with major geological events in thePonto-Caspian basin (Fig. 4 and Table 3). The basin has experienceda tumultuous geological history since the mid-Miocene epoch(�15 Mya), including multiple large sea-level changes and inter-mittent connection with the World Ocean, and associated inter-ba-sin connections between the Black and Caspian Sea basins(Mandych, 1995; Reid and Orlova, 2002). These fluctuating waterlevels and connections caused salinity levels within the basins torange 1–30 ppt over the last 5 My (Reid and Orlova, 2002), result-ing in lineage separations on multiple temporal scales.
The initial separation of the Black and Caspian Sea basins�5 Mya coincides with the diversification of most Neogobi-ini + Ponticolini genera (Neogobius, Babka, Mesogobius, Ponticola,and Proterorhinus), as well as diversification within Benthophilini(separation of Benthophilus and Caspiosoma; Fig. 3). Congruently,Cristescu et al. (2003, 2004) identified late Miocene divergences(�5.0–7.9 Mya) for benthic amphipods, and Audzijonyte et al.(2008) found a �5 Mya split between Paramysis lineages. Thesedivergences within diverse Ponto-Caspian fauna occurred on asimilar time scale as large-scale desiccation events in the Mediter-ranean Sea basin (Messinian Salinity Crisis �5.9 Mya) and in theeastern Paratethys/early Black Sea basin �5.5 Mya (Hsü and Gio-vanoli, 1979; Gillet et al., 2007). Desiccation of the Black Sea basinduring this period dramatically reduced water levels and increasedsalinity, enhancing isolation among tributaries within the basin.This led to allopatric separation of taxa residing in these freshwaterareas and increased speciation within the more saline basin.
In addition to older divergences within Ponto-Caspian taxa, sev-eral recent separation events are identified. Onset of the Pleisto-cene glaciations created additional fluctuations in water levelswithin the Ponto-Caspian basin (Reid and Orlova, 2002). Severalradiation events occurred 1–2 Mya among the Ponticolini, duringthe midst of these Pleistocene glacial cycles. Notably, in the Ponti-cola ‘‘platyrostris” species group both Po. cephalargoides and Po.cyrius diverged early �1.82 Mya, and are distributed at oppositeends of the Ponto-Caspian basin today (northwest Black Sea/AzovSea and the Kura River basin flowing into the Caspian Sea, respec-tively). Ponticola constructor, Po. platyrostris, and Po. rhodioni thenseparated �1.3 Mya and are found in the central portion of thePonto-Caspian basin (marine and freshwater areas of the easternBlack Sea). A similar distribution pattern occurs within the Pontico-la ‘‘kessleri” species group; with Po. gorlap occupying marine andfreshwater areas of the Caspian Sea basin and Don River, Po. eury-cephalus inhabiting marine areas of the northwest Black Sea andAzov Sea, and Po. kessleri primarily found in freshwater drainagesof the northwest Black Sea (Dnieper, Dniester and Danube Rivers).The freshwater species of Proterorhinus also occupy an analogousdistribution, with Pr. semilunaris occurring in freshwater basins ofthe northwest Black Sea, Proterorhinus sp. found in the Kumo-Manych Depression (Don/Manych River basin), and Pr. cf semipellu-cidus inhabiting the upper and lower Volga River basin and delta(Neilson and Stepien, 2009). These three species groups demon-strate a congruent biogeographic pattern: initial isolation and sep-aration of a broadly distributed taxon following closure of aninterbasin connection �1.7–2.0 Mya (Apsheron connectionthrough Kumo-Manych Depression; Kaplan, 1995; Reid and Orl-ova, 2002), succeeded by isolation and further radiation within
the Black Sea basin due to glacially-associated fluctuations inwater levels and basin shape. In addition, recent water level trans-gressions and separations within the Pleistocene coincide withlineage divergences of the two subspecies of Neogobius melanosto-mus (N. m. melanostomus in the Black Sea and N. m. affinis in theCaspian Sea; see Brown and Stepien, 2008; Fig. 3). This pattern ofPleistocene-aged phylogenetic and phylogeographic breaks amongBlack/Caspian Sea basins is echoed in a variety of taxa ranging fromother fishes (Rutilis frisii; Kotlík et al., 2008) to benthic and plank-tonic aquatic invertebrates (cladocerans—Cristescu et al., 2003,2004; dreissenid mussels—Stepien et al., 2003, 2002; Gelembiuket al., 2006; and mysids—Audzijonyte et al., 2006, 2008).
Our analysis of divergence times generally is congruent withevolutionary hypotheses proposed for European gobiids, primarilyin origins of the ‘‘transverse gobies” (Atlantic-MediterraneanGobius, Caffrogobius, Nematogobius, Mauligobius, Padogobius, andZosterisessor) and the ‘‘sand gobies” (Economidichthys, Knipowits-chia, and Pomatoschistus; Miller, 2003a). Penzo et al. (1998), usingportions of the mt 12S and 16S rRNA genes, estimated a separa-tion time of the transverse and sand gobies of �48 Mya; in thepresent study using cyt b, we resolved a clade containing Gobi-us + Zosterisessor (mostly identical to Penzo et al.’s study) separat-ing from a clade containing the sand gobies Knipowitschia andPomatoschistus �35 Mya (Fig. 3). In addition, McKay and Miller(1997) found a close relationship between the sand gobies andwestern Pacific gobiids using morphology and isozymes, indicat-ing an earlier separation from the transverse gobies. This associ-ation also is seen in our study, with the sand gobies appearingclosely related to the western Pacific microdesmids (andptereleotrids (cyt b; Fig. 3) or to other western Pacific gobiids(COI; Fig. 4). Although Miller (2003a) groups our Ponto-CaspianBenthophilinae as members of the transverse gobiids, it appearsthat they diverged much earlier from the transverse + sand gobyancestor, �42 Mya. Since the Benthophilinae shares the trans-verse pattern of cephalic neuromasts with other Atlantic-Mediter-ranean gobiids, the ‘‘transverse gobies” group is paraphyletic,retaining this ancestral character trait across multiple evolution-ary lineages.
5. Conclusion
The goby subfamily Benthophilinae represents an understudiedyet important component of the Ponto-Caspian fish fauna. Wepresent the most complete phylogenetic and biogeographic studyof the group, and clarify outstanding taxonomic issues presentfor the last 20 years. The Benthophilinae constitutes a unique radi-ation of gobiid fishes, and is a separate subfamily from the remain-der of the Gobiidae. Its evolutionary history has been driven by thedynamic geologic and hydrologic evolution of the Ponto-Caspianbasin.
6. Systematic conclusions
Benthophilinae Beling and Iljin 1927:309.Type genus Benthophilus Eichwald 1831.
Distinguishing features: small to moderate gobiids with infraor-bital neuromast organs (comprised of sensory papillae) in 6–7transverse rows, four before and 2–3 above hyomandibularrow b, and lacking row a. Dorsal supraorbital rows o showingseparation along dorsal midline. Tubular anterior nostrils, lack-ing process from the rim. Posterior nostril generally near orbit.Uppermost pectoral fin rays contained within membrane.Swimbladder not present. Moderate to large oligoplasmaticeggs; no pelagic larval stage. Benthophilinae can be separated

96 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
from the Gobiinae (where it was formerly included) by gener-ally increased number of total (628) and caudal (18–22) verte-brae. Primarily found in the Azov, Black, and Caspian Sea basinsand adjacent river drainages; several species introduced intocentral and northern Europe and the North American GreatLakes.
Tribe Benthophilini Beling and Iljin 1927:309.Type genus Benthophilus Eichwald 1831.
Distinguishing features: small gobiids with infraorbital neuro-mast organs in 6–7 rows, four before and 2–3 above hyoman-dibular row b, and lacking row a. Benthophilini can beseparated from other members of the Benthophilinae by thecombination of complete loss of all head canals, and reductionor complete loss of scales.
Genus Anatirostrum Iljin 1930: 48.Type species Benthophilus profundorum Berg 1927.Included species
Anatirostrum profundorum (Berg 1927). Original name: Bentho-philus profundorum Berg 1927.
Genus Benthophiloides Beling and Iljin 1927: 309.Synonym: Asra Iljin 1941: 384.Type species Benthophiloides brauneri Beling and Iljin 1927.Included species:
Benthophiloides brauneri Beling and Iljin 1927.Benthophiloides turcomanus (Iljin 1941). Original name: Asra tur-comanus Iljin 1941.
Genus Benthophilus Eichwald 1831: 77.Synonyms: Bentophilus Eichwald 1838: 102; Hexacanthus Nord-mann 1838: 332; Doliichthys Sauvage 1874: 336.Type species Gobius macrocephalus Pallas 1787.Included species:
Benthophilus abdurahmanovi Ragimov 1978. Original name:Benthophilus magistri abdurahmanovi Ragimov 1978.Benthophilus baeri Kessler 1877.Benthophilus casachicus Ragimov 1978. Original name: Bentho-philus stellatus casachicus Ragimov 1978.Benthophilus ctenolepidus Kessler 1877. Synonym: Benthophilusmagistri lencoranicus Ragimov 1982.Benthophilus durrelli Boldyrev and Bogutskaya 2004.Benthophilus granulosus Kessler 1877. Synonym: Benthophilussquamatus Baer in Lukina 1984.Benthophilus grimmi Kessler 1877.Benthophilus kessleri Berg 1927. Original name: Benthophilusgrimmi kessleri Berg 1927.Benthophilus leobergius Berg 1949. Original name: Bentho-philus stellatus leobergius Berg 1949. Synonym: Benthophilusaculeatus Baer in Lukina 1984.Benthophilus leptocephalus Kessler 1877.Benthophilus leptorhynchus Kessler 1877.Benthophilus macrocephalus (Pallas 1787). Original name: Gobiusmacrocephalus Pallas 1787. Synonym: Hexacanthus macrocepha-lus (Nordmann 1838).Benthophilus magistri Iljin 1927.Benthophilus mahmudbejovi Ragimov 1976.Benthophilus nudus Berg 1898. Original name: Benthophilus mac-rocephalus nudus Berg 1898. Synonym: Benthophilus macroceph-alus ponticus Berg 1916.
Benthophilus pinchuki Ragimov 1982. Original name: Benthophi-lus ctenolepidus pinchuki Ragimov 1982.Benthophilus ragimovi Boldyrev and Bogutskaya 2004.Benthophilus spinosus Kessler 1877.Benthophilus stellatus (Sauvage 1874). Original name: Doliich-thys stellatus Sauvage 1874. Synonyms: Benthophilus macroceph-alus maeotica Kuznetsov 1888; Benthophilus monstrosusKuznetsov 1888.Benthophilus svetovidovi Pinchuk and Ragimov 1979.
Genus Caspiosoma Iljin 1927: 129.Type species Gobiosoma caspium Kessler 1877.Included species:
Caspiosoma caspium (Kessler 1877). Original name: Gobiosomacaspium Kessler 1877.
Tribe Neogobiini new tribe, Neilson and StepienType genus Neogobius Iljin 1927.
Distinguishing features: moderate gobiids with infraorbitalneuromast organs in seven rows, four before and three abovehyomandibular row b, and lacking row a. Neogobiini can be sep-arated from other members of the Benthophilinae by the fol-lowing characters: head width about equal to depth;metapterygoid bridge absent; dentary with generally smallteeth, largest in the outer row.
Genus Neogobius Iljin 1927: 135.Synonyms: Apollonia (subgenus of Gobius) Iljin 1927: 133; Neo-gobius (subgenus of Gobius) Iljin 1927: 135.Type species: Gobius fluviatilis Pallas 1814.Included species:
Neogobius fluviatilis (Pallas 1814). Original name: Gobius fluvia-tilis Pallas 1814. Synonyms: Gobius sordidus Bennett 1835; Gobi-us lacteus Nordmann 1840; Gobius stevenii Nordmann 1840;Gobius niger Eichwald 1841 (not of Linnaeus 1758); Gobius flu-viatilis nigra Kessler 1859; Gobius fluviatilis pallasi Berg 1916;Gobius caspius Ragimov 1967. Other combination: Apollonia flu-viatilis (Stepien and Tumeo 2006).Neogobius melanostomus (Pallas 1814). Original name: Gobiusmelanostomus Pallas 1814. Synonyms: Gobius cephalarges Pallas1814; Gobius chilo Pallas 1814; Gobius melanio Pallas 1814;Gobius virescens Pallas 1814; Gobius exanthematosus Pallas1814; Gobius affinis Eichwald 1831; Gobius sulcatus Eichwald1831; Gobius lugens Nordmann 1840; Gobius grossholzii Steind-achner 1894; Gobius marmoratus Antipa 1909. Other combina-tions: Gobius melanostomus affinis Navozov 1912; Apolloniamelanostomus (Stepien and Tumeo 2006).Neogobius caspius (Eichwald 1831). Original name: Gobius casp-ius Eichwald 1831. Other combinations: Gobius (Eichwaldia)caspius Smitt 1900; Neogobius (Eichwaldia) caspius (Gaibova1952).
Tribe Ponticolini new tribe, Neilson and StepienType genus Ponticola Iljin 1927.
Distinguishing features: moderate gobiids with infraorbitalneuromast organs in generally seven rows, four before andthree above hyomandibular row b, and lacking row a. Pontico-lini can be separated from other members of the Benthophilinaeby the following characters: metapterygoid bridge present;hyomandibular generally narrow (breadth generally <100%length).

M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 97
Genus Babka Iljin 1927: 132.Synonym: Babka (subgenus of Gobius) Iljin 1927: 132.Type species Gobius gymnotrachelus Kessler 1857.Included species:
Babka gymnotrachelus (Kessler 1857). Original name: Gobiusgymnotrachelus Kessler 1857. Synonyms: Gobius macropus DeFilippi 1863; Gobius burmeisteri Kessler 1877; Gobius macroph-thalmus Kessler 1877; Mesogobius gymnotrachelus otschakovinusZubovitch 1925. Other combinations: Mesogobius gymnotrache-lus (Berg 1916); Gobius (Babka) gymnotrachelus Iljin 1927; Gobi-us (Mesogobius) gymnotrachelus Sözer 1941; Mesogobiusgymnotrachelus macrophthalmus (Berg 1949); Gobius (Babka)gymnotrachelus gymnotrachelus Banarescu 1964; Gobius gymno-trachelus macrophthalmus Ragimov 1967; Neogobius gymnotra-chelus (Miller 1973); Neogobius gymnotrachelus gymnotrachelus(Pinchuk 1977); Neogobius gymnotrachelus macrophthalmus(Pinchuk 1977).
Genus Mesogobius Bleeker 1874: 317.Synonym: Mesogobius (subgenus of Gobius) Bleeker 1874: 317.Type species Gobius batrachocephalus Pallas 1814.Included species:
Mesogobius batrachocephalus (Pallas 1814). Original name: Gobi-us batrachocephalus Pallas 1814. Synonym: Gobius batracho-cephalus borysthenis Pinchuk 1963. Other combinations:Gobius (Mesogobius) batrachocephalus Bleeker 1874; Gobiusbatrachocephalus batrachocephalus Smitt 1900.Mesogobius nigronotatus (Kessler 1877). Original name: Gobiusnigronotatus Kessler 1877.Mesogobius nonultimus (Iljin 1936). Original name: Gobiusnonultimus Iljin 1936. Other combination: Mesogobius batracho-cephalus nonultimus (Miller 1986).
Genus Ponticola Iljin 1927: 134.Synonym: Ponticola (subgenus of Gobius) Iljin 1927: 134.Type species: Gobius ratan Nordmann 1840.Included species:
Ponticola bathybius (Kessler 1877). Original name: Gobius bathy-bius Kessler 1877. Other combinations: Neogobius (Chasar)bathybius (Berg 1949); Neogobius fluviatilis pallasi (Berg 1949);Gobius (Chasar) bathybius Ragimov 1967a; Gobius bathybius Pin-chuk 1976; Neogobius bathybius (Pinchuk and Ragimov 1985).Ponticola cephalargoides (Pinchuk 1976). Original name: Neogo-bius cephalargoides Pinchuk 1976. Synonyms: Gobius syrmanKessler 1859; Gobius constructor Kessler 1874; Gobius cephalarg-es Chichkoff 1912; Gobius (Ponticola) cephalarges Borcea 1934;Neogobius cephalarges (Georghiev Aleksandrova and Nikolayev1960); Gobius ratan Pinchuk 1963; Gobius (Ponticola) cephalarg-es cephalarges Banarescu 1964; Neogobius ratan (Zambriborshch1968); Neogobius cephalarges cephalarges (Smirnov 1986).Ponticola constructor (Nordmann 1840). Synonyms: Gobius con-structor Nordmann 1840; Gobius platyrostris cyrius Kessler1879; Gobius platyrostris Berg 1916; Gobius platyrostris cyriusBerg 1923; Gobius cephalarges Iljin (1926) 1927; Gobius (Ponti-cola) platyrostris cyrius Iljin 1927a; Gobius cephalarges construc-tor Iljin 1927b; Neogobius cephalarges constructor (Berg 1949);Neogobius platyrostris constructor (Pinchuk 1977). Other combi-nation: Neogobius constructor (Vasil’eva and Vasil’ev 1994).Ponticola cyrius (Kessler 1874). Synonyms: Gobius cyrius Kessler1874; Gobius weidemanni Kessler 1874; Gobius platyrostris cyri-us (Berg 1916); Gobius constructor Berg 1923; Gobius platyrostris
Berg 1923; Gobius cephalarges constructor Iljin 1927; Neogobiuscephalarges constructor (Berg 1949); Neogobius platyrostris con-structor (Pinchuk 1977). Other combination: Neogobius cyrius(Vasil’eva and Vasil’ev 1994).Ponticola eurycephalus (Kessler 1874). Original name: Gobiuseurycephalus Kessler 1874. Synonyms: Gobius cephalarges Nord-mann 1840; Gobius platyrostris Ul’janin 1871; Gobius constructorKessler 1874; Gobius (Ponticola) cephalarges Iljin 1927; Neogobi-us cephalarges (Berg 1949); Gobius cephalarges Pinchuk 1963;Gobius (Ponticola) cephalarges cephalarges Banarescu 1964; Neo-gobius platyrostris (Georgiev 1966); Neogobius cephalarges(Bogachik and Remez 1970); Neogobius platyrostris eurycephalus(Pinchuk 1977); Neogobius platyrostris odessicus Pinchuk 1977.Other combinations: Gobius eurycephalus eurycephalus Smitt1900; Neogobius eurycephalus (Miller 1986).Ponticola gorlap (Iljin in Berg 1949). Original name: Neogobiuskessleri gorlap Iljin in Berg 1949. Synonyms: Gobius batracho-cephalus Eichwald 1841; Gobius kessleri Kessler 1874; Gobiusplatyrostris cyrius Derhavin 1926; Gobius cephalarges constructorDerzhavin 1934; Gobius kessleri gorlap Chugunova 1946; Neogo-bius cephalarges constructor (Berg 1949); Neogobius kessleri Oliva1960; Neogobius iljini Vasil’eva and Vasil’ev 1996. Other combi-nations: Neogobius (Ponticola) kessleri gorlap (Gaibova 1952);Gobius gorlap Iljin 1956.Ponticola kessleri (Günther 1861). Original name: Gobius kessleriGünther 1861. Synonyms: Gobius platyrostris Nordmann 1840;Gobius platycephalus Kessler 1857; Gobius cephalarges Steind-achner 1870; Gobius batrachocephalus platycephalus Smitt1900; Gobius trautvetteri Antipa 1909; Gobius (Ponticola) platy-rostris Borcea 1934. Other combinations: Gobius (Ponticola)kessleri Iljin 1927; Neogobius kessleri (Berg 1949); Neogobiuskessleri kessleri (Pinchuk 1977).Ponticola platyrostris (Pallas 1814). Original name: Gobius platy-rostris Pallas 1814. Synonyms: Gobius cephalarges platyrostrisSmitt 1900; Gobius cephalarges Smirnov 1959. Other combina-tions: Gobius (Ponticola) platyrostris Iljin 1927; Neogobius platy-rostris (Berg 1949); Neogobius platyrostris platyrostris (Pinchuk1977).Ponticola ratan (Nordmann 1840). Original name: Gobius ratanNordmann 1840. Synonyms: Gobius bogdanowi Kessler 1874;Gobius goebelii Kessler 1874; Gobius trautvetteri Kessler 1874.Other combinations: Gobius cephalarges ratan Smitt 1900; Gobi-us cephalarges bogdanowi Smitt 1900; Gobius cephalarges goebel-ii Smitt 1900; Gobius rotan Iljin 1927a; Gobius (Ponticola) ratanIljin 1927b; Neogobius ratan (Berg 1949); Neogobius ratan goe-beli (Berg 1949); Neogobius bogdanowi (Berg 1949); Gobius ratangoebeli Iljin 1956; Gobius ratan Pinchuk 1963; Neogobius (Ponti-cola) ratan ratan (Banarescu 1964); Neogobius ratan (Zam-briborshch 1968); Neogobius ratan ratan (Pinchuk 1976).Ponticola rizensis (Kovacic and Engín 2008). Original name: Neo-gobius rizensis Kovacic and Engín 2008.Ponticola rhodioni (Vasil’eva and Vasil’ev 1994). Original name:Neogobius rhodioni Vasil’eva and Vasil’ev 1994. Synonyms: Gobi-us constructor Nordmann 1840; Gobius platyrostris cyrius Kessler1879; Gobius platyrostris Berg 1923; Gobius platyrostris cyriusBerg 1923; Gobius cephalarges Iljin (1926) 1927; Gobius (Ponti-cola) platyrostris cyrius Iljin 1927a; Gobius cephalarges construc-tor Iljin 1927b; Neogobius cephalarges constructor (Berg 1949);Neogobius platyrostris constructor (Pinchuk 1977).Ponticola syrman (Nordmann 1840). Original name: Gobius syr-man Nordmann 1840. Synonyms: Gobius trautvetteri Kessler1859; Gobius eurystomus Kessler 1877; Gobius constructor Bor-cea 1934. Other combinations: Gobius (Ponticola) syrman Iljin1927; Neogobius syrman (Berg 1949); Neogobius syrman eurysto-mus (Berg 1949); Neogobius (Ponticola) syrman eurystomus(Gaibova 1952); Gobius (Ponticola) syrman eurystomus Iljin
98 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
1956; Gobius (Ponticola) syrman syrman Banarescu 1964; Gobiussyrman eurystomus Ragimov 1967; Neogobius syrman syrman(Smirnov 1986).Ponticola turani (Kovacic and Engín 2008). Original name: Neo-gobius turani Kovacic and Engín 2008.
Genus Proterorhinus Smitt 1900: 544.Synonym: Proterorhinus (subgenus of Gobius) Smitt 1900: 544.Type species Gobius marmoratus Pallas 1814.Included species:Proterorhinus marmoratus (Pallas 1814). Original name: Gobiusmarmoratus Pallas 1814. Synonyms: Gobius quadricapillus Pallas1814; Gobius macropterus Nordmann 1840.Proterorhinus nasalis (De Fillipi 1863). Original name: Gobiusnasalis De Fillipi 1863. Synonym: Gobius blennioides Kessler1877.Proterorhinus semilunaris (Heckel 1837). Original name: Gobiussemilunaris Heckel 1837. Synonym: Gobius rubromaculatus Kri-esch 1873.
Proterorhinus cf semipellucidus (Kessler 1877). Synonyms: Gobi-us semipellucidus Kessler 1877.Proterorhinus tataricus Freyhof and Naseka 2008.
Acknowledgments
We thank N. Bogutskaya, V. Boldyrev, L. Corkum, I. Grigorovich,J. Herler, S. Ibrahimov, H. Jenner, J. Kornichuk, V. Kovac, Y. Kvach, A.Naseka, J. Ram, S. Rudnicka, M. Sapoto, P. Simonovic, Y. Slynko, A.Smirnov, and C. Wiesner for specimen collection; V. Boldyrev andE. Vasil’eva for assistance with specimen identification; and D.Murphy for technical advice in the laboratory. This work wasfunded by a grant from the National Science Foundation (DEB-0456972) to C.A.S. We thank J. Banda, J. Brown, A. Haponski, D.Murphy, L. Pierce, R. Lohner, and O. Sepulveda-Villet for valuablecomments on the manuscript and N. Bogutskaya, W. Eschmeyer,J. Nelson, and E. Vasil’eva for discussions about the phylogeny. Thisis publication 2009-005 from the Lake Erie Center.
Appendix A
Geographic origin, GenBank accession numbers, and specimen ID for outgroup taxa analyzed in the present study.
Taxon and author
Location Latitude Longitude SpecimenIDGenBank Accession Nos.
cyt b
COI RAG1 S7Benthophilus abdurahmanoviRagimov 1978
Volga River delta, Russia
46.265635 49.093737 AKK5 FJ526777 FJ526832 FJ526886 FJ526960B. granulosus Kessler 1877
Volga River delta, Russia 46.265635 49.093737 AKK7 FJ526778 FJ526833 FJ526887 FJ526961 B. mahmudbejovi Ragimov1976
Volga River delta, Russia 46.265635 49.093737 AKK10 FJ526779 FJ526834 FJ526888 FJ526962B. stellatus Sauvage 1874
Dniester River Delta,Bilyayivka, Ukraine46.468333
30.216667 ALC6 FJ526780 FJ526835 FJ526889 FJ526963Caspiosoma caspium Kessler1877
Volga River delta, Russia
46.265635 49.093737 AOD3 FJ526781 FJ526836 FJ526890 FJ526964Chromogobius zebratusKolombatovic 1981
Adriatic Sea, Isle of Cres,Croatia
44.822745
14.337957 AQB2 FJ526783 FJ526838 FJ526892 FJ526966Gobius bucchichi Steindachner1870
Adriatic Sea, Isle of Krk,Croatia
45.094134
14.436694 AQB6 FJ526784 FJ526839 FJ526893 FJ526967G. fallax Sarato 1889
Adriatic Sea, Isle of Cres,Croatia44.822745
14.337957 AQB8 FJ526785 FJ526840 FJ526894 FJ526968G. niger Linnaeus 1758
Yalta Bay, Yalta, Ukraine 46.937577 37.258114 AOD8 FJ526782 FJ526837 FJ526891 FJ526965 G. xanthocephalus Heymerand Zander 1992
Atlantic Ocean, Arrabida,Portugal38.468233
�8.986918 AQB10 FJ526795 FJ526841 FJ526895 FJ526969Pomatoschistus minutusPallas 1770
Budaksky Lagoon, Ukraine
46.000000 30.500000 AOD1 FJ526776 FJ526831 FJ526885 FJ526959Zosterisessor ophiocephalusPallas 1814
Black Sea, Odessa, Ukraine
46.470820 30.735090 AHL6 FJ526747 FJ526797 FJ526851 FJ526906 Tyligul Estuary, Ukraine 46.690000 31.486783 AHL4 EU444670 EU444698 EU444724 FJ526905 Kerch Strait, Kerch,Ukraine45.358334
36.475834 APT6 FJ526748 FJ526798 FJ526852 FJ526907Appendix B
Reference list of taxonomic authorities for all taxa included inthe present study.
Antipa, G., 1909. Fauna Ichtiologica a Romaniei. RomanianEducational Academy, Bucharest.Beling, D.E., Iljin, B.S., 1927. Benthophiloides brauneri n.g. n. sp. Ein für das Schwarzmeerbassin neuer Vertreterder Familie der Gobiidae. Trav. St. Biol. Dniepr. 2, 309–325.
Banarescu, P., 1964. Pisces-Osteichthyes (Pestii Ganoizi siOsosii). Academia Republicii Populare Romine, Bucharest.Beebe, W., Tee-Van, J., 1928. The fishes of Port-au-PrinceBay, Haiti, with a summary of the known species of marinefishes of the island of Haiti and Santo Domingo. Zool. Sci.Contrib. N.Y. Zool. Soc. 10, 1–279.Bennett, E.T., 1835. Characters of several previously unde-scribed fishes form Trebizond. Proc. Zool. Soc. Lond. 3,91–92.Berg, L.S., 1898. On a collection of fishes from Bessarabia.Dnev. Zool. Otdel. Obshch. Estestv.2, 34.

M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 99
Berg, L.S., 1916. Freshwater Fishes of the Russian Empire.Izdatel’vesto Departmenta Zemledeliya, Moscow.Berg, L.S., 1923. Freshwater Fishes of Russia, 2nd edition.Gosizdat, Moscow.Berg, L.S., 1927. Notes on the Caspian Benthophilus (Gobii-dae). In: Collection of papers in honor of Prof. Knipowitsch1885–1927, vol. 1927. Moscow, pp. 331–344.Berg, L.S., 1949. Freshwater Fishes of the USSR and Adja-cent Countries. Izdatel’vesto Akademii Nauk SSSR, Moscow.Bleeker, P., 1853. Vierde bijdrage tot de kennis der ichthy-ologische fauna van Amboina. Natuur. Tijdsch. Ned. Ind. 5,317–352.Bleeker, P., 1855. Negende bijdrage tot de kennis der ich-thyologische fauna van Borneo. Zoetwatervisschen vanPontianak en Bandjermasin. Natuur. Tijdsch. Ned. Ind. 9,415–430.Bleeker, P., 1874. Esquisse d’un système naturel des Gobi-ides. Arch. Néerl. Sci. Exact. Nat. 9, 289–331.Bloch, M.E., 1792. Naturgeschichte der AusländischenFische, vol. 6. Berlin.Bloch, M.E., Schneider, J.G., 1801. Systema Ichthyologiae.Bogachik, T.A., Remez, Z.M., 1970. Morphology of the diges-tive apparatus of gobies and its significance for systemat-ics. In: Problems of Fishery Management of the Waters ofthe Ukraine. Naukova Dumka, Kiev, pp. 45–47.Boldyrev, V.S., Bogutskaya, N.G., 2004. Description of twonew species of tadpole-gobies (Teleostei: Gobiidae: Bentho-philus). Zoosystematica Rossica 13, 129–135.Borcea, I., 1934. Revision systematique et distribution geo-graphique des gobiides de la mer Noire et particulierementdes eaux Roumaines. Ann. Sci. Univ. Jassy 19, 1–236.Chugunova, N.I., 1946. The distribution of gobies in theNorth Caspian Sea. Zool. Zhurnal 25, 459–467.Cuvier, G., Valenciennes, A., 1837. Histoire Naturelle desPoissons.Derzhavin, A.N., 1926. Fishes of the Karasu River. Izv.Bakinsk. Ikhtyol. Lab. 2, 161–184.Derzhavin, A.N., 1934. Freshwater fishes from the southerncoast of the Caspian. Tr. Azerb. Otdel. Zak. Fiiiala Akad.Nauk SSSR, Zool. 7, 91–126.Eichwald, C.E., 1831. Zoologia Specialis quam Expositis Ani-malibus tum Vivis, Vol. 3. Vilna.Eichwald, C.E., 1838. Faunae Caspii Maris primitae. Bull.Soc. Nat. Moscou 11, 125–147.Eichwald, C.E., 1841. Fauna Caspio-Caucasica NonnullisObservationibus Novis Illustravit. St. Petersburg.Filippi, F. de, 1863. Nuove o poco note specie di animalivertebrati raccolte in viaggio in Persia nell’estate dell’anno1862. Arch. Zool. Anat. Filiol. Genova 2, 377–394.Freyhof, J., Naseka, A.M., 2007. Proterorhinus tataricus, anew tubenose goby from Crimea, Ukraine (Teleostei, Gobii-dae). Ichthyol. Explor. Freshw. 18, 325–334.Gaibova, R.A., 1952. Gobies from the Schihovo-Karadagskiiregion of the Caspian Sea. Tr. Inst. Zool. Akad. Nauk Azerb.SSR 15, 53–106.Georghiev, J.M., 1966. Composition d’espèce et caractéris-tique des Gobiides (Pisces) en Bulgaria. Izv. Nauchn. –Issled. Inst. Rib. Stop. Okeanogr. Varna 7, 159–228.Günther, A., 1861. Catalogue of the AcanthopterygianFishes in the Collection of the British Museum. Vol. 3;Gobiidae, Discoboli, Pediculati, Blenniidae, Labyrinthici,Mugilidae, Notacanthi. British Museum Press, London.Heckel, J.J. 1837. Ichthyologische Beiträge zu den Familiender Cottoiden, Scorpaenoiden, Gobioiden und Cyprinoiden.Ann. Wien. Mus. Natur. 2, 143–164.
Iljin, B.S., 1927. Bermerkunger über die pontischen Gobi-iden (Pisces in der Sammlung der Zoolgischen Museumsder Akademie der Wisenchaften. Yezhegodn. Zool. MuzeyaAN SSSR 27 (1926), 382–387.Iljin, B.S., 1927a. Regarding a new record of Gobius (Pontico-la) platyrustris cyrius Kessler. Tr. Kerch. Nauch. Rybokhoz.St. 1, 179–183.Iljin, B.S., 1927b. A guide to the gobies (family Gobiidae) ofthe Azov and Black seas. Tr. Azov. –Chernom. Nauch. –Prom. Eksped. 2, 128–143.Iljin, B.S., 1930. Le système des Gobiidés. Trab. Inst. Esp.Oceanogr. Madrid. 2, 1–63.Iljin, B.S., 1936. A new goby from the Caspian Sea Gobiusnonultimus sp. n. (Pisces, Gobiidae). Dokl. Acad. Nauk SSR4, 325–327.Iljin, B.S., 1941. Asra turcomanus, a new genus and speciesof goby (Gobiidae) from the Caspian Sea. Izv. Akad. NaukSSSR Biol. 1941, 385–390.Iljin, B.S., 1956. Remarks and corrections to suborderGobioidei in the book of L. S. Berg ‘‘Freshwater fishes ofthe USSR and neighboring countries”, 4th edit., 1948–1949, pp. 1055–1125. Vopr. Ikhtiol. 7, 185–192.Iwata, A., Jeon, S.-R., Mizuno, N., Choi, K.-C., 1985. A revi-sion of the eleotrid goby genus Odontobutis in Japan, Koreaand China. Jpn. J. Ichthyol. 31, 373–388.Kessler, K.T., 1857. Nachträge zur ichthyologie des süd-westlichen Russlands. Bull. Soc. Imp. Natur. Moscow 30,453–481.Kessler, K.T., 1859. Systematische Übersicht der Stachelf-losser (Acanthopteri Müller), welche im nordwestlichenTheile des schwarzen Meeres und in den Mündungen derin derselben sich ergießenden südrussischen Flüsse vork-ommen. Bull. Soc. Imp. Natur. Moscow 32, 186–268.Kessler, K.T., 1874. A description of fishes belonging to thefamilies common to both the Black and the Caspian seas.Tr. St. Petersb. Obsh. Estest. 5, 191–324.Kessler, K.T., 1877. Fishes of the Aralo-Caspio-Pontine ich-thyological region. Tr. Aral. –Kasp. Eksped. 4, 1–360.Kessler, K.T., 1879. Notiz über die Fische des FlussesTuapse. Bull. Soc. Nat. 54, 424–428.Kovacic, M., Engin, S., 2008. Two new species of Neogobiusfrom northeastern Turkey. Cybium 32, 73–80.Kolombatovic, G., 1891. Glamoci (Gobii) spljetskog pomor-skog okruzja u Dalmaciji. [Gobii della Dalmazia.] Spalato.Glamoci (Gobii) spljetskog pomorskog okruzja u Dalmaciji.[Gobii della Dalmazia.]: 3–29.Kriesch, A., 1873. Ein neuer Gobius. Verh. Zool. Bot. Ges.Wien 23, 369–376.Kuznetsov, I.D., 1888. Percarina and Benthophilus of the Seaof Azov. Tr. St. Petersb. Obshch. Estestv. 19, 189–212.Linnaeus, C., 1758. Systema Naturae, Ed. X.Lukina, T.A. (Ed.), 1984. Scientific Heritage, vol. 9. CaspianExpedition of K. M. Baer in 1853–1857. Diaries and Materi-als. Nauka, Leningrad.Miller, P.J., 1973. Gobiidae. In: Hureau, J.C., Monod, T. (Eds.),Check-List of the Fishes of the North-eastern Atlantic andof the Mediterranean. UNESCO, Paris.Miller, P.J. 1986. Gobiidae. In: Whitehead, P.J.P., Bauchot,M.L., Hureau, J.C., Nielsen, J., Tortonese, E. (Eds.), Fishes ofthe North-eastern Atlantic and the Mediterranean, Volume3. UNESCO, Paris, pp. 1019–1085.Navozov, N.P., 1912. Materials for an ichthyofauna of theUral River basin. Vestn. Ryb. 1912, 252–283.Nordmann, A., 1838. Bericht an die Kaiserliche Akademieder Wissenschaften über eine neue Fischgattung (Hexacan-

100 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
thus) aus der Familie der Gobioiden. Izv. Imp. Akad. Nauk.3, 328–332.Nordmann, A., 1840. Observations sur la fauna pontique.In: de Démidoff, A. (Ed.), Voyage dans la Russie méridionaleet la Crimée. Vol. III. Paris, pp. 353–635.Oliva, O., 1960. Remarks on the distribution of gobies at thewest coasts of the Middle and south Caspian. Izv. Akad.Nauk Azerb. SSR. 4, 47–50.Pallas, P.S., 1770. Spicilegia Zoologica quibus novae impri-mis et obscurae animalium species iconibus, descriptioni-bus atque commentariis illustrantur.Pallas, P.S., 1787. Piscium novae species descriptae. NovaActa Acad. Sci. Imp. Petropol. 1 (Hist.), 258–259.Pallas, P.S., 1814. Zoographia Rosso-Asiatica, sistens omni-um animalium in extenso Imperio Rossico et adjacentibusmaribus observatorum recensionem, domicilia, mores etdescriptiones anatomen atque icones plurimorum. Vol. 3.Petropoli.Pinchuk, V.I. 1963. Sea and lagoon forms of the knout gobyof the north-western part of the Black Sea. Dopov. NaukURSR 1, 126–128.Pinchuk, V.I., 1976. The taxonomy of gobies of the generaGobius Linné (native species), Neogobius Iljin and Mesogobi-us Bleeker. Vopr. Ikhtiol. 16, 600–609.Pinchuk, V.I., 1977. The taxonomy of gobies of the generaGobius Linné (native species), Neogobius Iljin and Mesogobi-us Bleeker. Vopr. Ikhtiol. 17, 587–596.Pinchuk, V.I., Ragimov, D.B., 1979. Benthophilus svetovidoviPinchuk and Ragimov, sp. n. (Pisces, Gobiidae) from theCaspian Sea and a key of species of the genus Benthophilus.Zool. Zhurnal 58, 515–519.Pinchuk, V.I., Ragimov, D.B., 1985. The lateral line system intwo endemic species of gobies of the Caspian Sea. Zool.Zhurnal 64, 562–566.Ragimov, D.B., 1967. On the systematics of fishes belongingto the genus Gobius in the Caspian Sea. In: Biological Pro-ductivity of the Kurinsk-Caspian Fishing Region. AkademiiNauka Azerbaijanskoy SSR Institut Zoologicheskiy, Baku,pp. 252–277.Ragimov, D.B., 1976. Benthophilus mahmudbejovi sp. nov.(Pisces, Gobiidae) from the Caspian Sea. Zool. Zhurnal 55,1196–1200.Ragimov, D.B., 1978. On the taxonomical status ofsome species of the genus Benthophilus (Gobiidae)from the Caspian and Azov Seas. Vopr. Ikhtiol. 18,791–798.Ragimov, D.B., 1982. New subspecies of Caspian gobies(Gobiidae, Benthophilus). Zool. Zhurnal 61, 47–55.Rendahl, H., 1924. Beiträge zur Kenntniss der marinen Ich-thyologie von China. Ark. Zool. 16, 1–37.Risso, A., 1810. Ichthyologie de Nice, ou Histoire Naturelledes Poissons du Département des Alpes Maritimes. F. Scho-ell, Paris.Rofen, R.R., 1958. The marine fishes of Rennell Island. Nat.Hist. Rennell Is. 1, 149–218.Rutter, C.M., 1897. A collection of fishes obtained in Swa-tow, China, by Miss Adele M. Fielde. Proc. Acad. Nat. Sci.Philadelphia 49, 56–90.Sarato, C., 1889. Causerie scientifique. Gobius fallaz. Gaz.Nice Alpes Mar. 16, 3.Sauvage, H.E., 1874. Notices ichthyologiques. Rev. Mag.Zool. 2, 332–340.Smirnov, A.I., 1959. Materials on the biology of fishes fromthe Black Sea in the Karadag region. Tr. Karadagsk. Biol. Sta.15, 31–84.
Smirnov, A.I., 1986. Okuneobraznye (bychkovidnye), Sko-rpenoobraznye, Kambaloobraznye, Prisokoperoobraznye,udil’Tchikoobraznye. Fauna Ukrainy, vol. 8, Ryby. NaukovaDumka, Kiev.Smith, C.L., 1964. Taenioides limicola, a new goby fromGuam, Marianas Islands. Micronesica 1, 145–150.Smith, J.L.B., 1958. The gunnellichthid fishes with descrip-tion of two new species from East Africa and of Gunnellich-thys (Clarkichthys) bilineatus (Clark), 1936. Ichthyol. Bull. J.L. B. Smith Inst. Ichthyol. 9, 123–129.Smitt, F.A., 1899. Preliminary notes on the arrangement ofthe genus Gobius, with an enumeration of its European spe-cies. Öfversigt af Kongliga Veterskaps-Akademiens För-handlingar 56, 543–555.Sözer, F., 1941. Les Gobiides de la Turque. Rev. Fac. Sci.Univ. Istanbul, Series B 6, 128–169.Steindachner, F., 1870. Ichthyologische notizen (X). Sit-zungsber. Kaiser. Akad. Wiss. Math. Naturwiss. Kl. 61,623–642.Steindachner, F., 1881. Ichthyologische Beiträge (X). Sit-zungsber. Kaiser. Akad. Wiss. Math. Naturwiss. Kl. 83,179–219.Steindachner, F., 1894. Ichthyologische Beiträge (XVII). Sit-zungsber. Kaiser. Akad. Wiss. Math. Naturwiss. Kl. 103,444–464.Stepien, C.A., Tumeo, M.A., 2006. Invasion genetics of Pon-to-Caspian gobies in the Great Lakes: a ‘cryptic’ species,absence of founder effects, and comparative risk analysis.Biol. Invasions 8, 61–78.Temminck, C.J., Schlegel, H., 1843. Pisces. In: Fauna Japon-ica, sive descriptio animalium quae in itinere per Japoniamsuscepto annis 1823–30 collegit, notis observationibus etadumbrationibus illustravit P. F. de Siebold, Part 1, pp. 1–20.Vasil’eva, E.D., Vasil’ev, V.P., 1994. Systematics of the Cau-casian riverine gobies (Gobiidae) in the light of moderndata, with the description of a new species, Neogobius rho-dioni sp. nova. Vopr, Ikhtiol. 34, 747–758.Vasil’eva, E.D., Vasil’ev, V.P., 1996. The description of Neo-gobius iljini sp. nov. within the former N. kessleri (Gobiidae,Pisces). Acta Univ. Carol. Biol. 39, 261–270.Vasil’eva, E.D., Vasil’ev, V.P, Pinchuk, V.I., 1993. Craniologi-cal analysis of the goby subgenus Ponticola Iljin, 1927. III.Comparative morphological study of Neogobius kessleri, N.ratan, and additional findings on N. syrman relevant tothe diagnosis and content of the subgenus. Vopr. Ikthiol.33, 609–617.Zambriborshch, F.S., 1968. On the taxonomy of Gobiidae ofthe basins of the Black and Azov seas. Vest. Zool. 1968, 37–44.Zubovitch, P.O., 1925. On the Black Sea gobies (Gobiidae). I.Mesogobius batrachocephalus otschakovinus subsp. Tr.Gosud. Chernom. –Azovskoi Opty. Sta. 1, 189–202.
References
Ahnelt, H., 2003. The postcranial skeleton of the benthophiline gobiids Anatirostrumand Benthophilus (Teleostei: Gobiidae). Folia Zool. 52, 213–221.
Ahnelt, H., Duchkowitsch, M., 2004. The postcranial skeleton of Proterorhinusmarmoratus with remarks on the relationships of the genus Proterorhinus(Teleostei: Gobiidae). J. Nat. Hist. 38, 913–924.
Ahnelt, H., Abdoli, A., Naderi, M., Coad, B.W., 2000. Anatirostrum profundorum: a raredeep-water gobiid species from the Caspian Sea. Cybium 24, 139–159.
Akihito, Iwata, A., Kobayashi, T., Ikeo, K., Imanishi, T., Ono, H., Umehara, Y.,Hamamatsu, C., Sugiyama, K., Ikeda, Y., Sakamoto, K., Fumihito, A., Ohno, S.,Gojobori, T., 2000. Evolutionary aspects of gobioid fishes based on an analysis ofmitochondrial cytochrome b genes. Gene 259, 5–15.

M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102 101
Audzijonyte, A., Daneliya, M.E., Väinölä, R., 2006. Comparative phylogeography ofPonto-Caspian mysid crustaceans: isolation and exchange among dynamicinland sea basins. Mol. Ecol. 15, 2969–2984.
Audzijonyte, A., Daneliya, M.E., Mugue, N., Väinölä, R., 2008. Phylogeny of Paramysis(Crustacea: Mysida) and the origin of Ponto-Caspian endemic diversity:resolving power from nuclear protein-coding genes. Mol. Phylogenet. Evol. 46,738–759.
Baker, R.H., DeSalle, R., 1997. Multiple sources of character information and thephylogeny of Hawaiian drosophilids. Syst. Biol. 46, 654–673.
Berg, L.S., 1949. Freshwater Fishes of the USSR and Adjacent Countries. Izdatel’vestoAkademii Nauk SSSR, Moscow.
Birdsong, R.S., Murdy, E.O., Pezold, F.L., 1988. A study of the vertebral column andmedian fin osteology in gobioid fishes with comments on gobioid relationships.Bull. Mar. Sci. 42, 174–214.
Boldyrev, V.S., Bogutskaya, N.S., 2007. Revision of the tadpole-gobies of the genusBenthophilus (Teleostei: Gobiidae). Ichthyol. Explor. Freshw. 18, 31–96.
Brower, A.V.Z., 2006. The how and why of branch support and partitioned branchsupport, with a new index to assess partition incongruence. Cladistics 22, 378–386.
Brown, J.E., Stepien, C.A., 2008. Ancient divisions, recent expansions:phylogeography and population genetics of the round goby Apolloniamelanostoma. Mol. Ecol. 17, 2598–2615.
Charlebois, P.M., Marsden, J.E., Goettel, R.G., Wolfe, R.K., Jude, D.J., Rudnika, S., 1997.The Round Goby, Neogobius melanostomus (Pallas), A Review of European andNorth American Literature. Illinois-Indiana Sea Grant and Illinois NaturalHistory Survey, Champaign.
Chow, S., Hazama, K., 1998. Universal PCR primers for S7 ribosomal protein geneintrons in fish. Mol. Ecol. 7, 1255–1256.
Cohen, A., Carlton, J., 1998. Accelerating invasion rate in a highly invaded estuary.Science 279, 555–558.
Corkum, L.D., Sapota, M.R., Skora, K.E., 2004. The round goby, Neogobiusmelanostomus, a fish invader on both sides of the Atlantic Ocean. Biol.Invasions 6, 173–181.
Cristescu, M.E.A., Hebert, P.D.N., Onciu, T.M., 2003. Phylogeography of Ponto-Caspian crustaceans: a benthic-planktonic comparison. Mol. Ecol. 12, 985–996.
Cristescu, M.E.A., Witt, J.D.S., Grigorovich, I.A., Hebert, P.D.N., MacIsaac, H.J., 2004.Dispersal of the Ponto-Caspian amphipod Echinogammarus ischnus: invasionwaves from the Pleistocene to the present. Heredity 92, 197–203.
Crossman, E.J., Holm, E., Cholmondeley, R., Tuininga, K., 1992. First record forCanada of the rudd, Scardinius erythrophthalmus, and notes on the introducedround goby, Neogobius melanostomus. Can. Field Nat. 106, 206–209.
Dillon, A.K., Stepien, C.A., 2001. Genetic and biogeographic relationships of invasiveround (Neogobius melanostomus) and tubenose (Proterorhinus marmoratus)gobies in the Great Lakes versus Eurasian populations. J. Great Lakes Res. 27,267–280.
Dobrovolov, J.S., Georghiev, T.U., Dobrolova, S.G., 1995. Comparative electrophoreticinvestigations of the species of the family Gobiidae (Pisces) in the Bulgariansector of the Black Sea. Proc. Inst. Fish. Varna 23, 48–68.
Dougherty, J.D., Moore, W.S., Ram, J.L., 1996. Mitochondrial DNA analysis of roundgoby (Neogobius melanostomus) and tubenose goby (Proterorhinus marmoratus)in the Great Lakes basin. Can. J. Fish. Aquat. Sci. 53, 474–480.
Farris, J.S., Kallersjo, M., Kluge, A.G., Bult, C., 1995. Constructing a significance testfor incongruence. Syst. Biol. 44, 570–572.
Freyhof, J., Naseka, A.M., 2007. Proterorhinus tataricus, a new tubenose gobyfrom Crimea, Ukraine (Teleostei, Gobiidae). Ichthyol. Explor. Freshw. 18,325–334.
Gelembiuk, G.W., May, G.E., Lee, C.E., 2006. Phylogeography and systematics ofzebra mussels and related species. Mol. Ecol. 15, 1033–1050.
Gillet, H., Lericolais, G., Réhault, J.-P., 2007. Messinian event in the Black Sea:evidence of a Messinian erosional surface. Mar. Geol. 244, 142–165.
Greenwood, P.H., 1984. What is a species flock? In: Echelle, A.A., Kornfield, I. (Eds.),Evolution of Fish Species Flocks. University of Maine Press, Orono, pp. 13–19.
Guindon, S., Gascuel, O., 2003. A simple, fast, and accurate algorithm to estimatelarge phylogenies by maximum likelihood. Syst. Biol. 52, 696–704.
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor andanalysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98.
Hsü, K.J., Giovanoli, F., 1979. Messinian event in the Black Sea. Palaeogeogr.Palaeoclimatol. Palaeoecol. 29, 75–93.
Iljin, B.S., 1927. A guide to the gobies (family Gobiidae) of the Azov and Black seas.Tr. Azov. –Chernom. Nauch. –Prom. Eksped 2, 128–143.
Johns, G.C., Avise, J.C., 1998. Tests for ancient species flocks based on molecularphylogenetic appraisals of Sebastes rockfishes and other marine fishes.Evolution 52, 1135–1146.
Johnson, T.B., Allen, M., Corkum, L.D., Lee, V.A., 2005. Comparison of methodsneeded to estimate population size of round gobies (Neogobius melanostomus)in western Lake Erie. J. Great Lakes Res. 31, 78–86.
Jude, D.J., DeBoe, S.F., 1996. Possible impact of gobies and other introduced specieson habitat restoration efforts. Can. J. Fish. Aquat. Sci. 53, 136–141.
Jude, D.J., Jannsen, J., Crawford, G., 1995. Ecology, distribution, and impact of thenewly introduced round and tubenose gobies on the biota of the St. Clair &Detroit rivers. In: Munawar, M., Edsall, T., Leach, J. (Eds.), The Lake HuronEcosystem: Ecology, Fisheries and Management. SPB Academic Publishing, pp.447–460.
Kaplan, P., 1995. The Caspian: it’s past, present and future. In: Mandych, A.F. (Ed.),Enclosed Seas and Large Lakes of Eastern Europe and Middle Asia. SPB AcademicPublishing, pp. 71–117.
Kotlík, P., Marková, S., Choleva, L., Bogutskaya, N.G., Ekmekçi, F.G., Ivanova, P.P.,2008. Divergence with gene flow between Ponto-Caspian refugia in ananadromous cyprinid Rutilus frisii revealed by multiple gene phylogeography.Mol. Ecol. 17, 1076–1088.
Kovacic, M., Engin, S., 2008. Two new species of Neogobius from northeasternTurkey. Cybium 32, 73–80.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J.,Higgins, D.G., 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23,2947–2948.
Lockwood, J.L., Hoopes, M.F., Marchetti, M.P., 2006. Invasion Ecology. BlackwellPublishing, Malden, MA.
López, J.A., Chen, W.-J., Orti, G., 2004. Esociform phylogeny. Copeia 2004, 449–464.Mandych, A.F. (Ed.), 1995. Enclosed Seas and Large Lakes of Eastern Europe and
Middle Asia. SPB Academic Publishing, Amsterdam, The Netherlands.McKay, S.I., Miller, P.J., 1997. The affinities of European sand gobies (Teleostei:
Gobiidae). J. Nat. Hist. 31, 1457–1482.Miller, P.J., 1973. The osteology and adaptive features of Rhyacichtys aspro
(Teleostei: Gobioidei) and the classification of gobioid fishes. J. Zool. (London)171, 397–434.
Miller, P.J., 1990. The endurance of endemism: the Mediterranean freshwater gobiesand their prospects for survival. J. Fish. Biol. 37 (Suppl. A), 145–156.
Miller, P.J., 2003a. Family Gobiidae Risso, 1826. In: Miller, P.J. (Ed.), The FreshwaterFishes of Europe, vol. 8(1). AULA-Verlag, Wiesbaden, pp. 157–162.
Miller, P.J., 2003b. Chasar Vasil’eva, 1996. In: Miller, P.J. (Ed.), The Freshwater Fishesof Europe, vol. 8(2). AULA-Verlag, Wiesbaden, pp. 94–96.
Miller, P.J., Vasil’eva, E.D., 2003. Neogobius Iljin, 1927. In: Miller, P.J. (Ed.), TheFreshwater Fishes of Europe. AULA-Verlag, Wiesbaden, pp. 164–171.
Mills, E.L., Leach, J.H., Carlton, J.T., Secor, C.L., 1993. Exotic species in the GreatLakes: a history of biotic crises and anthropogenic introductions. J. Great LakesRes. 19, 1–54.
Mooney, H.A., Cleland, E.E., 2001. The evolutionary impact of invasive species. Proc.Natl. Acad. Sci. USA 98, 5446–5451.
Neilson, M.E., Stepien, C.A., 2009. Evolution and phylogeography of the tubenosegoby genus Proterorhinus (Teleostei: Gobiidae): evidence for new crypticspecies. Biol. J. Linn. Soc. 96(3), in press.
Notredame, C., Higgins, D.G., Heringa, J., 2000. T-Coffee: a novel method for fast andaccurate multiple sequence alignment. J. Mol. Biol. 302, 205–217.
Penzo, E., Gandolfi, G., Bargelloni, L., Colombo, L., Patarnello, T., 1998. Messiniansalinity crisis and the origin of freshwater lifestyle in western Mediterraneangobies. Mol. Biol. Evol. 15, 1472–1480.
Pezold, F.L., 1993. Evidence for a monophyletic Gobiinae. Copeia 1993, 634–643.Pinchuk, V.I., 1991. Species groupings in the genus Neogobius (Perciformes). Vopr.
Ikhtiol. 31, 380–393.Posada, D., Crandall, K.A., 1998. Modeltest: testing the model of DNA substitution.
Bioinformatics 14, 817–818.Ray, W.J., Corkum, L.D., 1997. Predation of zebra mussels by round gobies, Neogobius
melanostomus. Environ. Biol. Fishes 50, 267–273.Reid, D.F., Orlova, M.I., 2002. Geological and evolutionary underpinnings for the
success of Ponto-Caspian species invasions in the Baltic Sea and North AmericanGreat Lakes. Can. J. Fish. Aquat. Sci. 59, 1144–1158.
Ricciardi, A., MacIssac, H.J., 2000. Recent mass invasion of the North American GreatLakes by Ponto-Caspian species. Trends Ecol. Evol. 15, 62–65.
Ronquist, F., Huelsenbeck, J.P., 2003. MrBayes 3: Bayesian phylogenetic inferenceunder mixed models. Bioinformatics 19, 1572–1574.
Sax, D.F., Gaines, S.D., 2008. Species invasions and extinction: the future of nativebiodiversity on islands. Proc. Natl. Acad. Sci. USA 105, 11490–11497.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparisons of log-likelihoodswith applications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116.
Simberloff, D., Von Holle, B., 1999. Positive interactions of nonindigenous species:invasional meltdown? Biol. Invasions 1, 21–32.
Simonovic, P.D., 1999. Phylogenetic relationships of Ponto-Caspian gobies and theirrelationship to the Atlantic-Mediterranean Gobiinae. J. Fish. Biol. 54,533–555.
Stepien, C.A., Taylor, C.D., Dabrowska, K.A., 2002. Genetic variability andphylogeographic patterns of a nonindigenous species invasion: a comparisonof exotic versus native zebra and quagga mussel populations. J. Evol. Biol. 15,314–328.
Stepien, C.A., Taylor, C.D., Grigorovich, I.A., Shirman, S.V., Wei, R., Korniushin, A.V.,Dabrowska, K.A., 2003. DNA and systematic analysis of invasive and nativedreissenid mussels: is Dreissena bugensis really D. rostriformis? Aquat. Invaders14, 8–18.
Stepien, C.A., Tumeo, M.A., 2006. Invasion genetics of Ponto-Caspian gobies in theGreat Lakes: a ‘‘cryptic” species, absence of founder effects, and comparativerisk analysis. Biol. Invasions 8, 61–78.
Swofford, D.L., 2003. PAUP*. Phylogenetic Analysis Using Parsimony (* and othermethods). Sinauer Assoc., Cambridge.
Thacker, C.E., 2003. Molecular phylogeny of the gobioid fishes (Teleostei:Perciformes: Gobioidei). Mol. Phylogenet. Evol. 26, 354–368.
USGS, 2003. Nonindigenous Aquatic Species Database (http://nas.er.usgs.gov). USGeological Survey, Reston, Virginia.
Vanderploeg, H.A., Nalepa, T.F., Jude, D.J., MIlls, E.L., Holeck, K., Liebig, J.R.,Grigorovich, I.A., Ojaveer, H., 2002. Dispersal and emerging ecological impactsof Ponto-Caspian species in the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci.59, 1209–1228.

102 M.E. Neilson, C.A. Stepien / Molecular Phylogenetics and Evolution 52 (2009) 84–102
Vasil’eva, E.D., 1989. Cranial morphology of the round goby Gobius melanostomusand syrman goby Gobius syrman in relation to their position in the genus Gobiussensu lato. Vopr. Ikhtiol. 29, 186–197.
Vasil’eva, E.D., 1991. The morphology of the skull of goby Gobius gorlap Iljin inconnection with its place in the genus Gobius sensu lato (Gobiidae, Pisces). Biull.Mosk. Ova. Ispyt. Prir. (Biol) 96, 36–45.
Vasil’eva, E.D., 1999. Taxonomic status of the tube-nosed goby Gobiusmarmoratus Pallas (Gobiidae) based on its skull morphology. Vopr. Ikhtiol.39, 139–148.
Vasil’ev, V.P., Grigoryan, K.A., 1992. Karyology of fishes of the family Gobiidae. Vopr.Ikthiol. 32, 27–40.
Vasil’eva, E.D., Vasil’ev, V.P., 1994. Systematics of the Caucasian riverine gobies(Gobiidae) in the light of modern data, with the description of a new species,Neogobius rhodioni sp.. Nova. Vopr, Ikhtiol. 34, 747–758.
Vasil’eva, E.D., Vasil’ev, V.P., Pinchuk, V.I., 1993. Craniological analysis of the gobysubgenus Ponticola Iljin, 1927. III. Comparative morphological study ofNeogobius kessleri, N. ratan, and additional findings on N. syrman relevant tothe diagnosis and content of the subgenus. Vopr. Ikthiol. 33, 609–617.





























