Embed Size (px)
Citation preview

UDC 597:591.4:554.3
Original scientific paper
PHE ETIC RELATIO SHIPS BETWEEN GOBIES (GOBIIDAE) BASEDON TRAITS OF EXTERNAL MORPHOLOGY
Predrag Sirnonovic! & Gabor Mesaros?
I Institute of Zoology, Faculty of Biology, University of Belgrade, Studentski trg 16,J 100J Belgrade, Yugoslavia
2 Department of Evolutionary Biology, Institute for Biological Research, University ofBelgrade, 29. Novembra 142, J1000 Belgrade, Yugoslavia
Sirnonovic P. & Mesaros G. (1998). Phenetic relationships between gobies(Pisces: Gobiidae) based on traits of their external morphology. Ichthyologia, Vol.30, o. 1, 43 - 50, Beograd.
Multivariate analysis that eccompassed five Euro-Mediterranean speciesiGobius spp. and Zosterisessor ophioccphalusy and five Ponto-Caspian species(Neogobius spp. and Proterorhinus marmoratusj of gobies and was based on 37external morphometric characters revealed that phenetic relationships betweengobies mainly do not conform to their systematic and zoogeographic status. Onlyone character, the interorbital distance, could be classified as zoogeographicallydiscriminative. The phenetic closeness between systematically and zoogeographi-cally distant species is likely to be determined by their affinities for a similar typeof habitat, with the prominent discriminative role of characters describing the cau-dal peduncle. The results imply that special attention should be paid to estimationof the role of homoplasy in the evolution of these gobiid taxa.
Key words: gobies, external morphology, phenetic relationships, zoogeogra-phy. ecology
Introduction
Morphological investigations of goby taxa are still the main source of data need-
Corresponding author: lnstitut za zoologiju, Bioloski fakultet, Univerzitet u Beogradu, 11000Beograd, Jugoslavija; tel. all 187266; e-mail:[email protected]

44 ICHTHYOLOGIA, Vol. 30, No.1, 43-50, 1998.
ed for making inferences about their relationships. The number of known gobiid taxa isestimated at up to 1800 species (Miller, 1986), but their worldwide distribution, diversi-ty, relatively small size, and usually cryptic life habits are reasons to expect that the pres-ent number is probably not even close to the real one. Several new go by species weredescribed just from European marine and fresh waters within the last decade(Economidis & Miller, 1990; Miller, 1992; Vasil'eva & Vasil'ev, 1994). In this prolifera-tive systematic group, a variety of morphological characteristics were used, ones of bothexternal morphology and osteology. Different approaches have been employed in pro-cessing external morphological characters of gobiid fishes. Miller (1986) and Bianco &Miller (1990) mostly relied on certain external traits established as specific to gobies,e.g., features of the pelvic disc and lateral line sensory system of the head. Economidis& Miller (1990) dealt with far greater variety of both external (e.g. body scalation, fea-tures of the perianal organ, meristic characters of fins, snout shape and structure) andinternal non-osteological traits (e.g., gonad shape and structure). As a result, certaincharacters have been established as "gobiid" ones, and their use has become commonin gobiid investigations, e.g., in McKay & Miller (1991). A comprehensive list of allexternal morphological traits used in gobiid taxa investigations on different classificationranks suggests that certain traits are specifically operational on particular classificationlevels.
As far as we know, relating gobies on the basis of their external continuous (mor-phometric) traits has not yet been done. Although gobies are very similar, any informa-tion obtained from results of analysis of external morphometric characters would con-tribute to the phylogenetic approach to gobiid systematics. Recent attempts at phyloge-netic analyses of gobies (e.g., Murdy, 1989; McKay & Miller, 1991) based on enzymevariability and morphology mostly confirmed the valid systematics of this group.However, it is to be expected that polarization in certain character states that could eas-ily distinguish particular gobiid taxa would lead to ordinary confirmation of the phenet-ic approach of experienced taxonomists (Abbot et al.; 1985). Hence the role of other,non-discriminative characters such as continuous morphometric traits could be usefulfor real testing of hitherto valid gobiid systematics.
A closer insight into a patterns of within and between species variability of exter-nal morphology in gobies might throw some light on ambiguities of evolutionary diver-gence in this group. The overlappings of ranges occurring for characters in gobiidspecies should be analyzed in order to check the amount of homoplasy vs. the homolo-gy acting in phylogeny of those species. Similar overlappings noticed in certain kary-ological characteristics (Vasil'ev & Grygorjan, 1992) were processed for the same pur-pose by Sirnonovic (1996). Thus, the phenetic approach could be an overture to theimplementation of phylogenetic analysis on external continuous morphological features.
Material and methods
Material used for analysis comprised 191 individuals of 10 gobiid species from afour genera. The samples of black goby Gobius niger Linnaeus, 1758 (12 individuals),Bucchich's go by Gobius bucchichi Steindachner, 1877 (two individuals), and red-mouthed goby Gobius cruentatus Gmelin, 1789 (one individual), were collected inAugust 1994 in the Boka Kotorska Bay, on a flat sandy bottom, a bottom of boulders,and a rocky bottom, respectively. The sample of rock go by Gobius paganellus Linnaeus,I 758 (10 individuals) was collected in August 1995 at Buljarice, in the southern part of

Sirnonovic P. & Mesaros G.: PHENETIC RELATIONSHIPS
east Adriatic coast, on a mosaic sand-rock bottom. They were collected by breath div-ing, using a hand trap filled with crushed mussels and sea urchins. The one individualof grass goby Zosterisessor ophiocephalus (Pallas, 1811-1814) was obtained from theRazelm Lake, eastern Romania. The samples of sand goby Neogobius jluviatilis (Pallas,1811) (40 individuals), bighead goby Neogobius kessleri (Gunther, 1861) (45 individuals)and tubenose goby Proterorhinus marmoratus (Pallas, 1811) (33 individuals) were caughton an open sand-mud bottom, a bottom of stones and boulders, and a hard soil bottomcovered, densely with macrophytic vegetation, respectively. They were taken from theSava and Danube Rivers in the Belgrade area using hand nets, with electrofishing gear,and by angling. The sample of round goby Neogobius melanostomus (Pallas, 1811) (30individuals) was obtained from the Gdansk Bay of the Baltic Sea. Two specimens ofracer goby Neogobius gymnotrachelus (Kessler, 1857) were caught in 1992 in the DanubeRiver in the Brza Palanka area (884 km) on a soft bottom overgrown with vegetation,using electrofishing, and 15 specimens of that species were from the Razelm Lake, east-ern Romania.
Each individual was measured using a millimeter caliper to the nearest tenth ofmillimeter under the binoculars (4 fold magnification). The measuring design (Fig. 1)comprised linear distances between homologous landmarks 00 three projections: ventral(A), dorsal (B) and lateral (C). This design the "truss box network" scheme accordingto Bookstein et aL (1985), was chosen in order to process the shape of fish as compre-hensively as possible, and it comprised 37 linear distances.
The source data set was log transformed in order to satisfy the main condition forusing parametric statistics. It was then analyzed by MANOVA using the Statistica pack-age, Release 4.5, from StatSoft, Inc., 1993 with species and sexes as factors (interactionsincluded) in order to check the homogeneity of species samples. The pattern of betweenspecies variability was processed by Canonical Discrimination using the SAS package(PROC CANDISC) (SAS Inc, 1991). This method favors group (species) variability andcorrelates variability of morphometric traits with group appurtenance. Mahalanobis dis-tances D2(i,j)= (Xi-Xj)-1 COY (Xi-Xj) (Sneath & Sakal, 1973) between centroids ofscattered individuals of particular species were clustered using an agglomerativeUPGMA clustering method (Sneath & Sokal, 1973). The dendrogram of relationsbetween species was generated using NTSYS-pc package V. 1.50 (Rohlf, 1988).
Results
MANOVA revealed that only the variability between species was significant (R=43.183; dfspecies=36; dfsex=740; p<O.OOI). Insignificant variability of sexes (R=1.942,dfspecies=6;dfsex=168; p>0.05) and that of the interaction of species with sex (R=0.857;dfspecies=36; dfsex=740; p>0.05) approved using a common sample of all 10 species(comprising 191 individuals) for Canonical Discrimination.
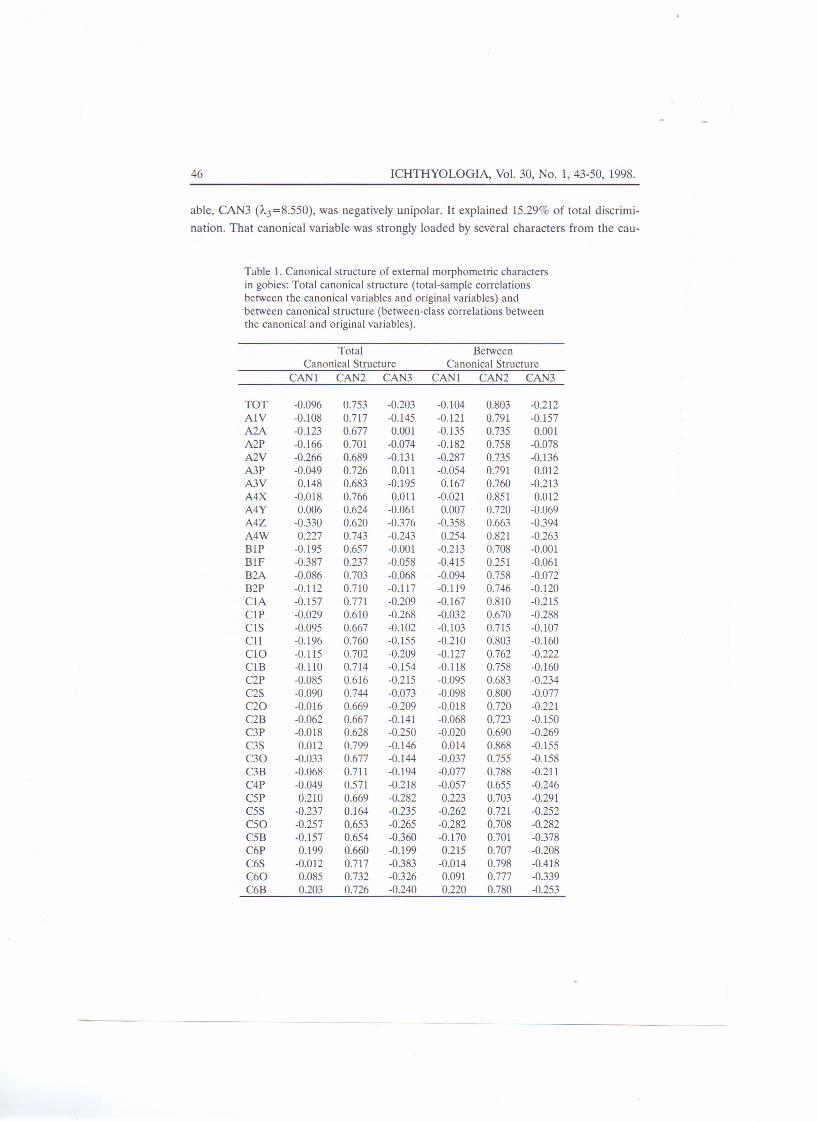
The first three canonical variables (CANI-CAN3) explained 77.38% of the wholediscrimination between species. The first canonical variable, CANI (1"1 =20.963), wasbipolar. It explained 37.49% of total discrimination between species. The strongest char-acter on that canonical variable was interorbital distance, i.e Blf .. The second canonicalvariable, CAN2 (A2=13.750), was positively unipolar. It explained 24.59% of total dis-crimination. All characters except interorbital distance loaded that canonical variablestrongly, implying their strong determination by overall size. The third canonical vari-
45
46 ICHTHYOLOGIA, Vol. 30, No.1, 43-50, 1998.
able, CAN3 (/1.3=8.550), was negatively unipolar. It explained 15.29% of total discrimi-
nation. That canonical variable was strongly loaded by several characters from the cau-
Table 1. Canonical structure of external morphometric charactersin gobies: Total canonical structure (total-sample correlationsbetween the canonical variables and original variables) andbetween canonical structure (between-class correlations betweenthe canonical and original variables).
Total BetweenCanonical Structure Canonical Structure
CAN 1 CAN2 CAN3 CAN 1 CAN2 CAN3
TOT -0.096 0.753 -0.203 -0.104 0.803 -0.212A1V -0.108 0.717 -0.145 -0.121 0.791 -0.157A2A -0.123 0.677 0.001 -0.135 0.735 0.001A2P -0.166 0.701 -0.074 -0.182 0.758 -0.078A2V -0.266 0.689 -0.131 -0.287 0.735 -0.136A3P -0.049 0.726 0.011 -0.054 0.791 0.012A3V 0.148 0.683 -0.195 0.167 0.760 -0.213A4X -0.018 0.766 0.011 -0.021 0.851 0.012A4Y 0.006 0.624 -0.061 0.007 0.720 -0.069A4Z -0.330 0.620 -0.376 -0.358 0.663 -0.394A4W 0.227 0.743 -0.243 0.254 0.821 -0.263B1P -0.195 0.657 -0.001 -0.213 0.708 -0.001B1F -0.387 0.237 -0.058 -0.415 0.251 -0.061B2A -0.086 0.703 -0.068 -0.094 0.758 -0.072B2P -0.112 0.710 -0.117 -0.119 0.746 -0.120CIA -0.157 0.771 -0.209 -0.167 0.810 -0.215CIP -0.029 0.610 -0.268 -0.032 0.670 -0.288CIS -0.095 0.667 -0.102 -0.103 0.715 -0.107Cll -0.196 0.760 -0.155 -0.210 0.803 -0.160ClO -0.115 0.702 -0.209 -0.127 0.762 -0.222ClB -0.110 0.714 -0.154 -0.118 0.758 -0.160C2P -0.085 0.616 -0.215 -0.095 0.683 -0.234C2S -0.090 0.744 -0.073 -0.098 0.800 -0.077C20 -0.016 0.669 -0.209 -0.018 0.720 -0.221C2B -0.062 0.667 -0.141 -0.068 0.723 -0.150C3P -0.018 0.628 -0.250 -0.020 0.690 -0.269C3S 0.012 0.799 -0.146 0.014 0.868 -0.155C30 -0.033 0.677 -0.144 -0.037 0.755 -0.158C3B -0.068 0.711 -0.194 -0.077 0.788 -0.211C4P -0.049 0.571 -0.218 -0.057 0.655 -0.246C5P 0.210 0.669 -0.282 0.223 0.703 -0.291C5S -0.237 0.164 -0.235 -0.262 0.721 -0.252C50 -0.257 0.653 -0.265 -0.282 0.708 -0.282C5B -0.157 0.654 -0.360 -0.170 0.701 -0.378C6P 0.199 0.660 -0.199 0.215 0.707 -0.208C6S -0.012 0.717 -0.383 -0.014 0.798 -0.418C60 0.085 0.732 -0.326 0.091 0.777 -0.339C6B 0.203 0.726 -0.240 0.220 0.780 -0.253

Sirnonovic P. & Mesaros G.: PHENETIC RELATIONSHIPS 47
dal part of the body, namely: analfin base length (A4z), diagonal dis-tance between the beginning of theanal fin base and end of the seconddorsal fin base (C5b), dorsal lengthof the caudal peduncle(C6s), anddiagonal distance between the endof second dorsal fin base and begin-ning of the ventral part of the cau-dal fin base (C60) (Tab. 1).
The canonical structure ofdiscrimination revealed thatbetween-group canonical structurewas more prominent than the with-in-group one (Table 1) in all of thefirst three canonical variables,implying that all three canonicalvariables discriminated mostlyspecies, with negligible interferenceof intraspecific sources of variabilityon the overal discrimination.
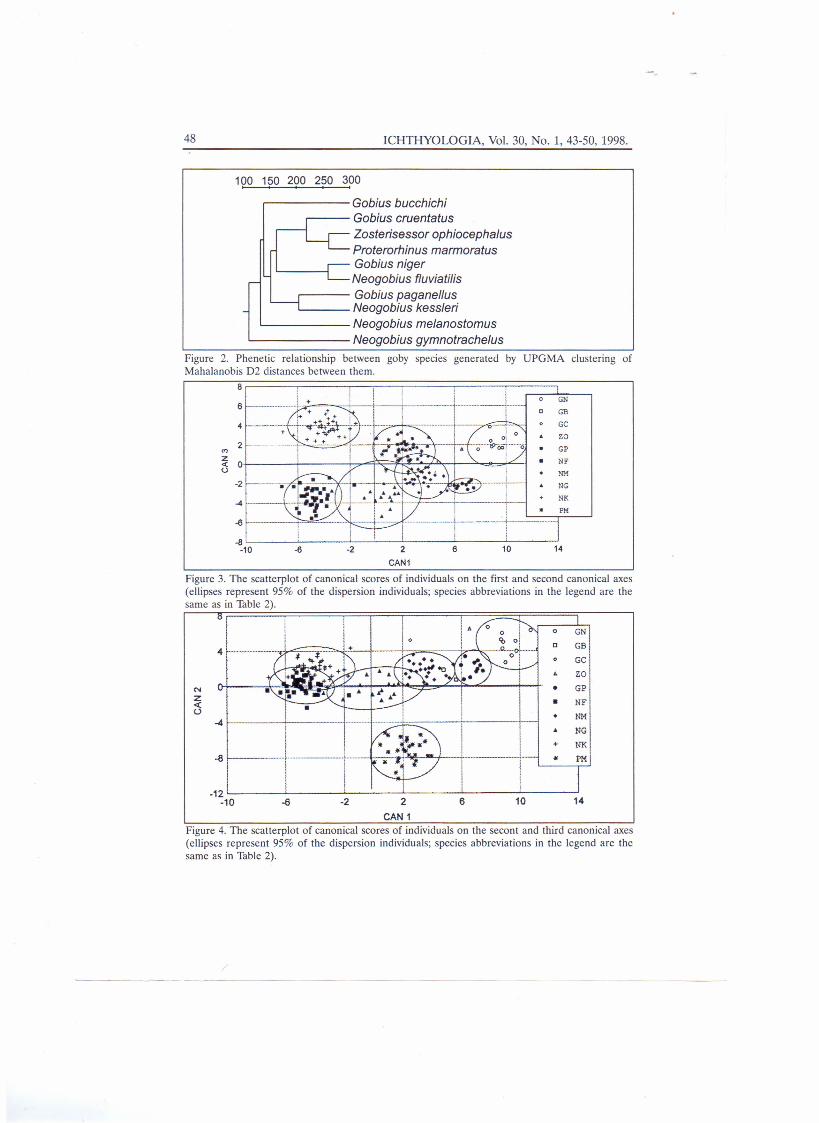
Mahalanobis distances be-tween centroids of scattered individ-ual scores of particular species (Tab.2), UPGMA clustering of species according to their Mahalanobis distances (Fig. 2), aswell as plotting of their canonical scores (Figs. 3 & 4) revealed that relationshipsbetween species are very difficult to interpret. On both the first and second canonicalvariables, the most similar two species are bighead go by and sand goby (Fig. 3).However, they are rather different on the third canonical variable (Fig. 4), meaning thattheir caudal peduncles are different. Their position on the phenogram, however, did notreveal any prominent closeness (Fig. 2).
A
z
Figure 1. The measuring design of external morpho-metric characters in gobies on the ventral (A), dorsal(8), and lateral (C) projections
Table 2. Squared Mahalanobis distance between goby species on their external morphometric traits(Cb - Gobius bucchichi; Cc - Gobius cruentatus; 20 - Zosterisessor ophiocephalus; CIl - Gobius niger;Cp - Gobius paganellus, Nf - Neogobius fluviatilis; Ng - Neogobius gymnotrachelus; Nk - Neogobiuskessleri; Nm - Neogobius melanostomus; Pm - Proterorhinus marmoratus'[.
Cb Cc Zo C•• CfJ NI Ng Nk Nm Pm
Cb 0
Cc 153.915 0
Zo 1016.317 187.845 0
C•• 117.041 151.392 55.370 0
CfJ 60.3015 178.938 122.143 97.492 0
NI 174.499 188.501 239.299 259.436 195.499 0
Ng 95.496 124.326 178.049 173.168 112.719 54.815 0
Nk 167.460 155.588 188.343 205.997 204.700 62.314 86.788 0
Nm 138.092 147.721 143.619 119.339 163.629 114.714 79.696 114.769 0
Pm 155329 250.020 260.871 212.578 182.162 132.010 97.677 1301.480 123.218 0
48 ICHTHYOLOGIA, Vol. 30, No.1, 43-50, 1998.
190 150 290 250 3PO
,------- Gobius bucchichi.---- Gobius cruentatus
Zosterisessor ophiocephalusProterorhinus marmoratusGobius niger
Neogobius f/uviatilis.----- Gobius paganellus'----- Neogobius kessleri
'------- Neogobius melanostomus'-------- Neogobius gymnotrachelus
Figure 2. Phenetic relationship between goby species generated by UPGMA clustering ofMahalanobis D2 distances between them.
8GN
6 c GBGCzo
'" GPz • NF«<.J NM
-2 NG-4 NK--.-
PM
-6 .....f--.-.----.--
-8-10 -2 2 6 10 14
CAN1
Figure 3. The scatterplot of canonical scores of individuals on the first and second canonical axes(ellipses represent 95% of the dispersion individuals; species abbreviations in the legend are thesame as in Table 2).
GNe
D GBe GC~ ZO• GP• NF• NH
I • NG
-i------- ....-. + NK,. PM
!
4
i-4 r--'---+I---
I .--·--·-·l·-····--··-------r···..-
_12~---L-------L-------L-------L------'------~-10 -6 -2 2 6 10 14
CAN 1
-8
Figure 4. The scatterplot of canonical scores of individuals on the secont and third canonical axes(ellipses represent 95% of the dispersion individuals; species abbreviations in the legend are thesame as in Table 2).

Simonovic P. & Mesaros G.: PHENETIC RELATIONSHIPS 49
DiscussionAlthough the obtained relationship between analyzed goby taxa did not match the
contemporary classification, the pattern of between-species variabilty based on theirexternal morphology indicated some interesting relations. The phenetic closeness ofzoogeographically discrete taxa (e.g., grass and tubenose goby, black and sand go by,stone and bighead goby) (Fig. 2) could correspond more to characteristics of their habi-tats. Thus, both grass and tubenose gobies inhabit a more or less weedy and soft bottomsubstrate, both black and sand goby inhabit an open sandy bottom, whereas both stoneand bighead goby inhabit either a pure or a mosaically stony bottom (Smirnov, 1986;Miller, 1986). This suggests that the direction of morphological divergence of gobiid taxamight be induced by adaptation to a complex of specific ecological conditions. The posi-tion of the red-mouthed goby and Bucchich's goby deviates from relationships consid-ered in this way, since they are confined to bottoms of rocks and boulders, respectivel-ly.
The prominent role of characters that describe the caudal peduncle on the thirdcanonical variable indicates that correspondence between tail shape and different habi-tat types should be studied for explanation of morphological (dis)similarity. However,deviations imply somewhat more complex determination of the relationships foundhere.
It was possible to explain the zoogeographic determinant of morphology by itsmatching to the first canonical variable determinant, interorbital distance (Blf). Thus,variation of this characteristic in Ponto-Caspian genera (Neogobius, Proterorhinus) andspecies was different in relation to Euro-Mediterranean genera iGobius, Zosterisessor)and species (Figs. 3 & 4). No clear relationships could be seen between characteristicsthat strongly weighted the other two canonical variables and the zoogeographic statusof these species.
Our results suggest that both Euro-Mediterranean and Ponte-Caspian gobies aremorphometrically very similar in the majority of traits. The results of phenetic analysisdid not conform to the classification in those two zoogeographically distinct groups. Itseems that consistent phylogenetic classification of gobies should take into account boththe level and the pattern of morphological similarity, which point to the occurrence ofhomoplasy during the evolution of those two zoogeographically both distinct and adja-cent gobiid groups, wich are still problematic systematically.
Acknowledgements
The authors are indebted to Petru Banarescu (Acad. Sci. Romania), KrzysztofSkora (Hel Mar. Station, Univ. Gdansk) and A1eksandar Hegedis (Center Multidisc.Studies, Univ. Belgrade) for the materials provided.
References
Abbot, L.A., Bisby, EA. & Rogers J.D. (1985): Taxonomic analysis in biology. Columbia UniversityPress, New York, pp. 336.
Bianco, P.G. & Miller J.P. (1990): Yugoslavian and other records of the Italian freshwater goby,Padogobius martensii, and a character polarisation in gobioid fishes. J. Nat. Hist. 24: 1289-1302
Bookstein, EL., Chernoff, B., Elder, R.L., Humphries, 1.M., Smith, G.R. & Strauss, E.R. (1985):Morphometries in evolutionary biology. The Acad. Nat. Sci. Philadelphia, Spec. Pub!. 15,pp.277.

50 ICHTHYOLOGIA, Vol. 30, No.1, 43-50, 1998.
Economidis, P.S. & Miller, P.l. (1990): Systematics of the freshwater gobies from Greece (Teleostei:Gobiidae). 1. Zool. London 221: 125-170.
McKay, S.l. & Miller, P.J. (1991): Isozyme criteria in the testing of phyletic relationships betweenspecies of Gobius and related eastern Atlantic-Mediterranean genera (Teleostei:Gobiidae). 1. Fish BioI. 39 (Suppl. A): 291-299.
Miller, P.l. (1986): Gobiidae. pp. 1019-1085. In: Fishes of north-eastern Atlantic and theMediterranean (FNAM). UNESCO, Paris.
Miller, P.l. (1992): A new species of Didogobius (Teleostei: Gobiidae) from the Adriatic Sea. 1. Nat.Hist. 26: 1413-1419.
Murdy, E.O. (1989): A taxonomic revision and cladistic analysis of the oxudercine gobies(Gobiidae: Oxudercinae). Rec. Austral. Mus., Suppl. 11: 1-93.
Rohlf, F.l. (1988): NTSYS-pc: numerical taxonomy and multivariate analysis system. Version 1.50.Exeter Publishing Ltd. & Applied Biostatistics Inc., New York.
Sirnonovic, P.O. (1996): Karyological relationships between gobies (Gobiidae: Perciformes) ofEuro-Mediterranean and Ponto-Caspian districts. Acta BioI. Yugoslavica - Ichthyologia, 28:25-31.
Smirnov, AI. (1986): Perciformes (Gobioidei), Scorpaeniformes, Pleuronectiformes,Lophiiformes. In: Fauna Ukraini 8: 7-183.
Sneath, P.H.A & Sokal, R. R. (1973): Numerical taxonomy. W.H. Freeman & Co., San Francisco,pp.573.
Vasil'ev, v.P. & Grygoryan K. A (1992): Karyology of fish of the family Gobiidae. VOpL Ikhth.32: 27-40.
Vasil'cva, E.D. & Vasil'ev V. P. (1994): On the systematics of the Caucasian riverine gobies(Gobiidae): further craniological and karyological analyses and distribution of the popula-tions from the Black and Caspian Seas basins in biotopes. VOpL Ikhth. 34: 187-194.
Received: 26 May, 1998Accepted: 10 December, 1998
FENETICKI ODNOSI MEDU GLAVOClMA (GOBIIDAE) NA OSNOVUSVOJSTAVA SPOLJASNJE MORFOLOGIJE
Predrag Simonovic! i Gabor Mesaros-
I Institut za zoologiju, Bioloski fakultet, Univerzitet u Beogradu, 11000 Beograd2 Odeljenje za evolucionu biologiju, Institut za bioloska istraiivanja "Sinisa
Stanokovic", Univerzitet u Beogradu, 11000 Beograd, Jugoslavija
Sazetak
Multivarijatna feneticka analiza radena na pet evro-mediteranskih vrsta(Gobius spp. i Zosterisessor ophiocephalus) i pet ponto-kaspijskih vrsta (Neogobius spp. iProterorhinus marmoratus) glavoca, po 37 spoljasnjih morfometrijskih karaktera, pokaza-la je da feneticki odnosi izmedu tih vrsta glavoca u vecini ne odgovaraju sistematskim izoogeografskim vezama ustanovljenim medu njima. Sarno jedan karakter, interorbitalnorastojanje, mogao bi se okarakterisati zoogeografski diskriminativnim. Ustanovljenafeneticka bliskost izmedu sistematski i zoogeografski udaljenih vrsta verovatno jeodredena slicnoscu tipova stanista prema kojima pojedine vrste pokazuju afinitet, saizrazenorn diskriminativnom ulogom karaktera koji opisuju repnu drsku u odnosu natipove stanista. Ovakvi rezultati ukazuju da pri taksonomskim istrazivanjima trebaobratiti narocitu paznju na procenu uloge homoplazije u evoluciji ovih vrsta glavoca.
Primljeno: 26. maja 1998.Odobreno: 10. decembra 1998.
























