Embed Size (px)
Citation preview

304 Brain Research. ~31 ~ 1 ~)~).i ! 304 .! ~ 2 .c 1993 Elsevier Science Publishers B.V. Ali rights reserved 0006-,~'493 , ~ ~tl¢~i~!
BRES 19524
Estrogen-receptor immunoreactivity in hamster brain: preoptic area, hypothalamus and amygdala
Hui-Yun Li *, Jeffrey D. Blaustein, Geert J. De Vries, George N. Wade
Neuroscience and Behavior Program and Psychology Department, Uni~,ersity of Massachusetts, Amherst, MA 01003, USA
(Accepted 27 July 1993)
Key words: Estrogen receptor; Immunocytochemistry; Preoptic area; Hypothalamus; Amygdala; Hamster
The distribution of estrogen-receptor containing cells in the preoptic area, hypothalamus and amygdala of female Syrian hamster brain was studied by immunocytochemical methods. Dense populations of estrogen-receptor immunoreactive (ER-IR) cells were found in the medial preoptic area, the bed nucleus of the stria terminalis, amygdala, ventral and lateral parts of the hypothalamus, and the arcuate nucleus. Injection of estradiol caused a decrease in estrogen-receptor immunoreactivity (ERIR) containing cells within one hour, a decrease that may reflect a change in the ability of the occupied estrogen receptor to bind the particular antibody (H222) used rather than down-regulation of the estrogen receptor. Our findings on the distribution of estrogen-receptor containing cells in these areas using an immunocytochemical technique are consistent with and extend the findings of others using autoradiographic and in vitro binding techniques to study estrogen receptor distribution in hamster brain.
INTRODUCTION
Gonadal steroids regulate gonadotropin release s'18'43
and sexual behavior 6 at least in part by activating steroid hormone receptors in the central nervous sys- tem. As in other steroid hormone target tissues, estra- diol binds with estrogen receptors in the brain to form steroid-receptor complexes, causing changes in gene expression, leading to alterations in protein synthesis and consequently to alterations in cellular function 6. To begin to understand the molecular and cellular processes by which steroid hormones influence behav- ior, it is important to delineate the precise sites of hormone action, especially in neuroanatomical areas which are involved in the regulation of reproduction like the hypothalamus, preoptic area (POA) and amyg- dala.
Many sites containing sex steroid-concentrating ceils are involved in the neuroendocrine regulation of sexual behavior and other aspects of reproduction. For exam- ple, the results of a variety of experiments suggest that steroid receptor containing neurons in the ventrome- dial hypothalamus (VMH) are targets for estradiol
* Corresponding author. Fax: (1) (413) 545-0996.
action on female estrous behavior t2'j6'47. Implants of
estradiol in the VMH induce lordosis in response to subsequent progesterone injection 12"~6'47 in ovariec-
tomized hamsters, while lesions of the VMH attenuate the expression of lordosis in hormonally-primed ham- sters 3°. These results are consistent with the notion that estradiol acts on hypothalamic cells, altering the activity of those cells in such a way as to facilitate female sexual behavior.
Other forebrain areas beside the VMH have been implicated in the regulation of sexual behavior. While results of studies on the precise role of the medial preoptic area (MPA) have been contradictory, it is clear that ovarian steroids act at this site to influence estrous behavior 2"25'39. In addition, the amygdaloid area
is crucial in modulating and integrating sexual behavior in female rats. Activation of cells in the anterior part of the corticomedial nucleus of amygdala increases lor- dotic responses, while excitation of neurons in the posterior part of the lateral amygdaloid nucleus has the opposite effect 33. Moreover, the medial nucleus of amygdala (MeA) has direct connections with several brain sites that are involved in the regulation of behav- ioral and hormonal aspects of reproductive functions, such as the V M H 29'42, MPA and septal areas 15. These findings suggested that amygdaloid structures may play

a modulatory or integrative role in the control of sexual behavior in female rats.
Estradiol-concentrating neurons have been previ- ously demonstrated autoradiographically in a variety of species 23'36'45'46'50, and the distribution of estradiol-con- centrating neurons, which includes the POA, hypotha-
305
lamus and amygdala, is similar across a variety of species 37. A similar pattern of estrogen receptor mRNA has also been detected using in situ hybridization in rats 24'38'44. Studies using immunocytochemical tech- niques, which provides better cellular resolution, have confirmed that estrogen receptor-immunoreactive (ER-
/ ¸
\ ...... ,.
Io
~ . e ~"I',~ ' f E n
,:/ ...:S . : " \ % .F": .
. . . . : .. ~ " . ~ r . GP CPu . ,!.... ...., . .. ,...,, .;,,~,~'.',... . - , : . . .
~" " ~ ; : ~i' ~;~
Fig. 1A
Fig. 1. Camera lucida drawings of sections, rostral to caudal, through the preoptic area (Fig. 1A) and hypothalamus and amygdala (Fig. 1B) of the estrogen receptor-immunoreactive cells in the hamster brain. Each dot represents a ceil. Abbreviations: 3V, 3rd ventricle; AA, anterior amygdaloid area; ac, anterior commissure; ACo, anterior cortical amygdaloid nu; AH, anterior hypoth area; AHiAL, amygdalohip area, anterolat; Arc, arcuate hypoth nu; BM, basomedial amygdaloid nu; BMP, basomedial amygdaloid nu, post; BSTIA, bed nu st, intraamyg div; BSTL, bed nu stria ter, lat div; BSTM, bed nu stria ter, med div; CeM, central amyg nu, med div; CPu, caudate putamen; DMD, dorsomedial hypoth nu, diffuse; DM, dorsomedial hypoth nu; f, fornix; GP, globus pallidus; ic, internal capsule; LA, lateroanterior hypoth nu; LH, lateral hypoth area; LHb, lateral habenular nu; lo, lateral olfactory tract; LPO, lateral preoptic area; LSI, lateral septal nu, intermediate; LV, lateral ventricle; MD, mediodorsal thai nu; ME, median eminence; MeA, medial amygdaloid nu, anterior; MHb, medial habenular nu; MPA, medial preoptic area; mt, mammillothalamic tract; MTu, medial tuberal nu; opt, optic tract; ox, optic chiasm; PaAP, paravent hypotha nu, ant parvo; PCo, posterial cortical amygda nu; PeF, perifornical nu; PMCo, posteromed cortical amyg nu; RCh, retrochiasmatic area; SCh, suprachiasmatic nu; SFi, septofimbria nu; SI, substantia innominata; sm, stria medullaries thai; SO, supraoptic nucleus; SStr, substriatal area; st, stria terminalis;
VEn, ventral endopiriform nu; VMH, ventromedial hypoth nu; VP, ventral pallidum. From Paxinos and Watson, 1986.

306
\ L.-" i:~:~l ~:;±(:>I /'-~ 8 t ~ -T ~ /
PaAP ~_~ Cl::~u
Fig. IA (continued).
o ,~ / ~
Fig. 1B (continued).

. t'.~.2.. I ..K,. ~ ",. ~" : ~ .
0 'o."°" - ~~"--S~'"~ c:, . , . . . ,,.,~, ,.~ ~, ~.~ ,.,. .~ ,'~ / ~ . ~ :~ ~ . ... , ~ , c~. , ~...,
"~ . .~ " ' . " , • . , ~:~,,':I
..-i;."~ -..~r
" " • .. c > :..." •
/ ~ i '~ / ~ ~ ~ ~ ?
. . . . . r ~ 6
Fig. 1C (continued).
307
IR) cells are distributed within the hypothalamic, pre- optic, and amygdaloid areas as well as midbrain 7'14'49. For example, dense populations of ER-IR cells have been observed in the MPA, the bed nucleus of the stria terminalis (BST), the arcuate nucleus (ARC), medial amygdala, and the VMH in rats 3'u'2°, guinea pig 7, o p o s s u m s 17, musk shrews 13 and ferrets 48. While one immunocytochemical study of hamster brain has been reported, it was limited to the medial preoptic area 26.
Recently, in vitro binding studies showed that in female hamsters, the MPA, mediobasal hypothalamus, and lateral hypothalamus (LH) contain high concentra- tions of estrogen receptors 9. This technique, however is limited in that it does not afford single-cell resolution. In the present experiment, we developed a sensitive, immunocytochemical technique using a monoclonal an- tibody to human estrogen receptors in order to enable us to map ER-IR cells throughout the hypothalamus, POA, and amygdala of hamster brains. This technique which is more sensitive than [3H]estradiol autoradi- ography 23, allows the precise mapping of the location of ER-IR cells under a variety of physiological condi- tions.
M A T E R I A L S A N D M E T H O D S
Animals
Female Syrian hamsters weighing 90-100 g were purchased from Charles River Breeding Laboratories (Wilmington, MA) and housed in stainless-steel wire-bottom cages (17.7 x 17.5 x 25 cm) at a room temperature of 22+ I°C. Hamsters were housed in a long photope- riod (LD 16:8 h; lights on at 05.00 h) and given ad libitum access to Purina Laboratory Rodent Chow (No. 5001) pellets and tap water.
Two to six weeks prior to each experiment, animals were bilaterally ovariectomized through dorsal incisions under sodium pentobarbital anesthesia (80 mg/kg i.p.).
Perfusion
Hamsters were deeply anesthetized with sodium pentobarbital (80 mg/kg i.p.). Five thousand units of sodium heparin dissolved in 1.0 ml saline were injected directly into the left ventricle followed by cutting the right atrium of the heart. A cannula was inserted into the left ventricle. Normal saline (75-100 ml) preceded the flow of fixative (2% paraformaldehyde in 0.1 M sodium phosphate buffer containing 0.4% glutaraldehyde). Perfusion pressure was maintained at approximately I00 mmHg, and flow rate was maintained at ap- proximately 25 ml per minute for 10 min. Brains were removed from the cranium and stored at 4°C overnight in 0.1 M sodium phosphate buffer containing 20% sucrose.
Immunocy tochemis t ry
The immunocytochemical technique developed for estrogen-re- ceptor immunoreactivity (ERIR) in guinea pigs 7 was used with some modifications. Forty-/zm coronal sections were cut through the pre- optic area and hypothalamus on a freezing microtome, and free- floating sections were rinsed three times for 5 min each with 0.05 M Tris-buffered saline (TBS; pH 7.6) followed by 1% sodium borohy- dride (in TBS) for 10 rain to reduce residual aldehydes. Sections were then rinsed with 0.05 M TBS four times for 5 rain each followed by incubation for 20 min in TBS with 1% hydrogen peroxide, 1% bovine serum albumin, and 20% normal rabbit serum to deplete endogenous peroxidase activity and to decrease non-specific staining.
The sections were incubated in H222 antibody, a rat monoclonal antibody to human estrogen receptors (Abbott Laboratories, North Chicago, IL) at a concentration of 1 /~g/ml in TBS containing 0.5% Triton X-100, 0.1% gelatin and 0.02% sodium azide, pH 7.6 at 4°C for 72 h. The sections were then rinsed with the same buffer for three times for 5 min each followed by incubation for 90 rain at room temperature in secondary antiserum (biotinylated rabbit anti-rat immunoglobulins, Vector Laboratories, Burlingame, CA), diluted in the same buffer as the primary antibody at a concentration of 6 /zg/ml. Following two washes with and one without Triton X-100 (5 min each), the sections were incubated for 90 min in the avidin DH/biotinylated horseradish peroxidase H complex (1:100, Vectas- tain Elite ABC Kit, Vector Laboratories, Burlingame, CA) diluted at
308
P
t l
7~

309
a concentration of 1:100 in TBS. Following rinsing with TBS contain- ing 0.5% Triton X-100 and 0.02% sodium azide at pH 7.6 two times and TBS once, sections were incubated with diaminobenzidine (0.05% in TBS) for 10 min in the presence of hydrogen peroxide (0.05%) and rinsed in TBS.
In the present report, we studied the distribution of ERIR in the brain of female hamsters using the H222 antibody which is directed against the steroid binding domain of the receptor 19. Previous stud- ies which have demonstrated the specificity of the monoclonal anti- body H222 for estrogen receptors in guinea pigs have included preadsorption of the antibody with a estrogen receptor-rich cellular extract 5, omission of the primary antibody and acute pretreatment of animals with estradiol to decrease ERIR 4'7. Specificity of the anti- body has been demonstrated by preadsorption with estrogen recep- tor-rich extracts in rats tl, opossums 17, musk shrews t3 and sheep (Lehman, M.N., personal communication). In the present experi- ment, the specificity of immunostaining was determined by omission of the primary antibody during immunocytochemistry. In addition, ovariectomized hamsters were given subcutaneous injection with 50 gg of estradiol-17/3 (i.p., dissolved in 0.1 ml of 10% ethanol) or oil vehicle at 1 h prior to perfusion to determine if the ERIR is rapidly influenced by estradiol and, therefore, likely to be the estrogen receptor.
Camera lucida drawings Drawings of the preoptic area, hypothalamus and amygdala im-
munostained for estrogen receptors were made using bright-field illumination to map ER-IR cells and dark-field illumination to delineate cell groups. Drawings were made every 160 /zm through the preoptic area, hypothalamus and amygdala. The nomenclature from the rat atlas of Paxinos and Watson was adopted 35, because no equivalent atlas is available for the hamster brain.
R E S U L T S
Consistent with data from [3H]estradiol autoradiog- raphy in female hamsters 23, dense population of ER-IR cells were found in the MPA, BST, periventricular area, ARC, ventrolateral-VMH, medial tuberal nu- cleus, and amygdala (Fig. 1). However, we observed ERIR in other areas in which estrogen receptors were previously undetected using autoradiography. These areas include the lateral preoptic area (LPO), substan- tia innominata (SI), ventral endopiriform nucleus (VEn), and dorsomedial hypothalamic nucleus (DM).
Within the POA, ER-IR cells were densely packed surrounding the rostral part of the periventricular area and gradually disappeared at more caudal levels. A dense population of ER-IR cells was observed within the MPA, with the densest concentration of cells in the anterior portion (Fig. 2A and B). Large numbers of ER-IR cells were also observed extending dorsolater- ally towards the fornix and stria terminalis, especially the medial portion of the BST.
Within the VMH, ERIR was seen most abundantly in the lateral division and its surround, extending later- ally into the medial tuberal nucleus (Figs. 1B, 2C and 2D). ER-IR cells were also found in the ARC of the hypothalamus (Figs. 1B and 2E). As has been observed previously, in most cases, the distribution of ER-IR cells did not follow boundaries of cell groups defined by Nissl staining. In fact, in the VMH, many ER-IR cells were found in the lateral area adjacent to the ventromedial hypothalamus. Within the medial and cortical nuclei of amygdala (Figs. 1B and 2F) ER-IR cells were found in the MeA, central nucleus of amyg- dala (CeA), anterior and posterior portions of cortical amygdaloid nucleus.
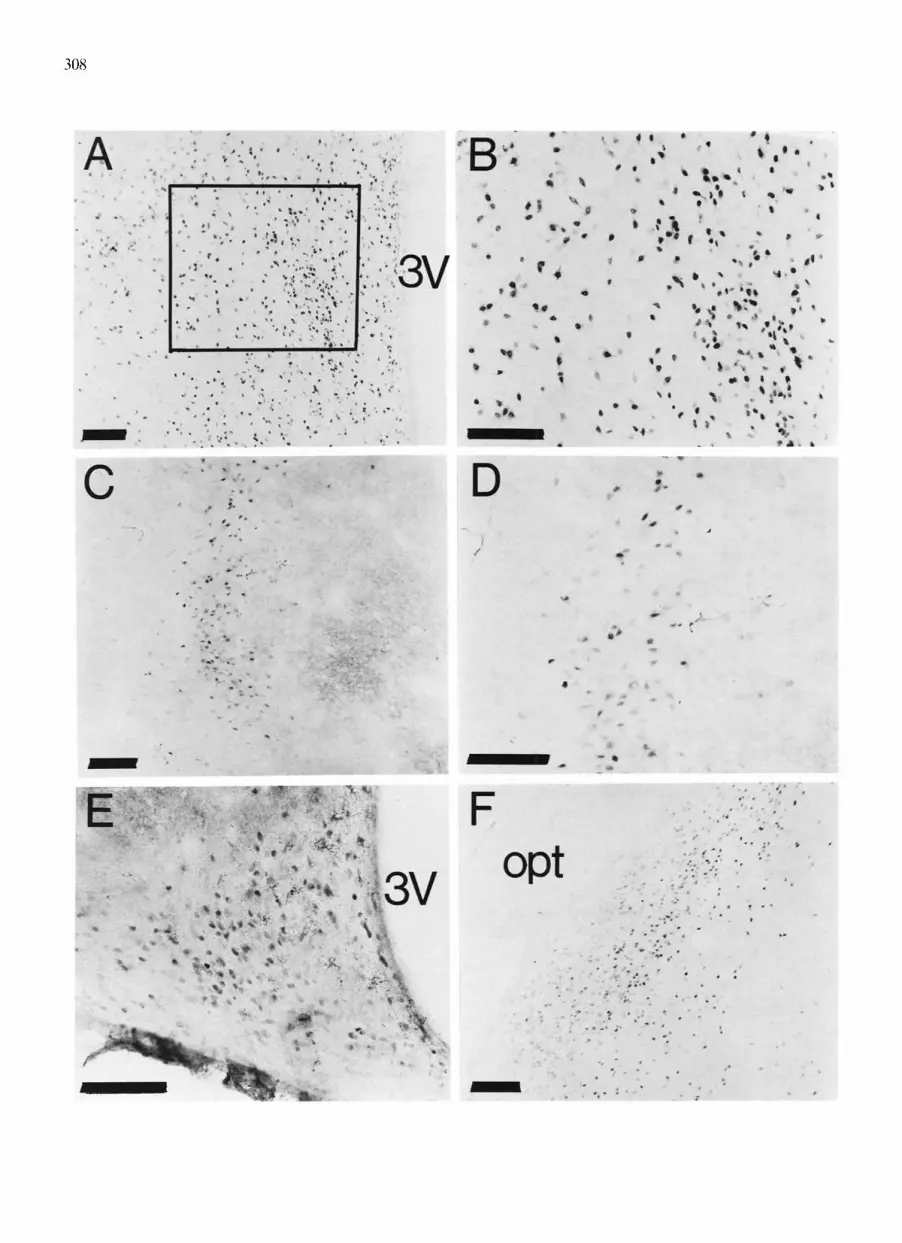
In all cases, especially within MPA, immunostaining was darkest in cell nuclei, with many cells having abundant ERIR in the cytoplasm (Fig. 3). Injection of estradiol one hour prior to perfusion resulted in a dramatic loss of ER-IR cells and lighter staining in most areas (Fig. 4A) compared to oil-treated animals (Fig. 4B). Omission of the primary antibody resulted in the absence of positively immunostained cells (Fig. 4C). Although red blood cells are seen occasionally, they are easily distinguishable at high-power magnification.
D I S C U S S I O N
The distribution of ERIR containing cells in the POA, hypothalamus and amygdala of hamsters is simi- lar to the distribution of [3H]estrogen-concentrating cells observed using autoradiography 23. Presumably due to the increased sensitivity of the immunocytochemical technique, ERIR containing cells were also observed in other areas of the brain, including the lateral preoptic area, substantia innominata, ventral endopiriform nu- cleus and dorsomedial hypothalamic nucleus.
Many of the neuroanatomical sites containing high concentrations of ER-IR cells in the hamster brain has been demonstrated to regulate certain aspects of re- production in previous studies. The POA 3°, anterior hypothalamus t°, V M H 12'16'30'47 and amygdala 33 have all
been implicated in the control of estrous behavior in female hamsters. For example, lesions at the lateral aspect of the VMH significantly suppress lordosis, and this is correlated with our finding that dense ER-IR cells are confined in the lateral portion of the VMH.
or--
Fig. 2. ERIR in various neuroanatomical areas immunostained with the H222 antibody and a vectastain Elite procedure, using diaminobenzidine as the chromogen. A: medial preoptic area. B: higher magnification of the medial preoptic area from the region indicated by the box in A. C: ventromedial nucleus of hypothalamus. D: higher magnification of the ventromedial nucleus of hypothalamus. E: arcuate nucleus. F: amygdala.
Magnification bars = 100 p,m.

310
~,! (i!!~)~ ~' i ̧~̧;̧̧ ̧ : ! ~iiiiiiii!,~,, ~!:~iii~ ~:̧̧
Fig. 3. Higher power magnification of ERIR showing cytoplasmic immunostaining in the medial preoptic area of an ovariectomized hamster visualized by the diaminobenzidine-peroxidase method.
Magnification bars = 10/.~m.
Unlike the VMH, one role of estradiol in the POA seems to be inhibition of female sexual behavior. For example, in female rats and guinea pigs, lesions in the MPA facilitate sexual behavior 25'39"4°, while in female
rats and hamsters, electrical stimulation of the POA area dramatically inhibits lordosis 3t'34. However, estra-
diol implants in the MPA also facilitate lordosis sug- gesting that the inhibitory effect of the MPA on sexual behavior can be masked by estradiol t27'32'41"53. Using
estradiol implants, the neural region in and about the arcuate nucleus was found to be most sensitive to estradiol for the inhibition of ovulation 28. Similarly,
electrical stimulation in amygdaloid nuclei suggested that amygdala is important in regulation of the behav- ioral and endocrine aspects of reproductive function in female rats 33'52.
The fact that estradiol dramatically decreased ERIR in most areas in the hamster brain within sixty minutes is in agreement with previous work in rats and guinea pigs 4'7. It has been suggested that this rapid decrease in rats is unlikely be due to down-regulation of estro- gen receptors, because it occurs within twenty minutes of estradiol injection 4. Rather, it is more likely that the loss of ERIR is due to some consequence of occupa- tion of estrogen receptors by estradiol. The rapid loss of E R I R observed in rats occurs with the H222 anti- body, but not with an antibody directed at the N- terminus of the receptor protein 4 suggesting that it is likely to be due to an occupation-induced conforma- tional change in estrogen receptors. Nevertheless, the rapid modulation of ERIR after estradiol provides
B
¢gl
I
Fig. 4. ERIR in the ventromedial nucleus of hypothalamus from ovariectomized hamsters that were (A) estradiol treated or (B) injected with oil vehicle. Sections were immunostained with the H222 antibody and a vectastain Elite procedure, using diaminobenzi- dine as the chromogen. C: omission of the H222 results in an
elimination of immunostaining. Magnification bars = 100 #m.

indirect evidence that the present immunocytochemical technique labels estrogen receptors, per se.
Also in agreement with recent work in rats 3, guinea pigs 7'14, and a variety of other species 13'17'48 is the finding that ERIR was detected in the cytoplasm as well as in cell nuclei. This does not conflict with an abundance of other work in other reproductive tissues which has suggested that the estrogen receptor is pri- marily a nuclear protein22'5~; in fact, the darkest im- munostaining in hamster brain is seen in cell nuclei. However, while the occurrence of cytoplasmic estrogen receptor immunostaining has been observed in a vari- ety of species and by a variety of investigators, others (e.g. refs. 11,21) have reported little or no cytoplasmic immunostaining of estrogen receptors in rat brain. This discrepancy may be due to the fact that sensitivity of the ER-immunocytochemical technique decreases markedly as a consequence of postfixation; in fact, it has been observed that the longer the postfixation, the weaker the ER-immunostaining of both cytoplasm and nucleus (Blaustein, J.D., unpublished). In the present experiment, brains were perfused for 10 minutes and then placed immediately into sucrose-phosphate buffer without postfixation.
Based on our results, the H222 monoclonal antibody to ER can be used for immunocytochemical detection of ERIR containing cells in hamster brain. This should be a useful technique to study the neuroanatomical distribution of cells containing estrogen receptors which may be involved in the actions of estradiol. Moreover, a comprehensive understanding of the neuroanatomi- cal distribution of ERIR in certain areas in the brain sheds light on the behavioral and physiological aspects of steroid functions in different brain loci.
Acknowledgements. This research was supported by the following grants: NS 10873 and DK 32976 from the NIH (G.N.W.), RSDA 00885 from the NIMH (J.D.B.), and NS 19327 from the NIH (J.D.B.). We thank Robin Lempicki and Jay Alexander for their expert technical assistance and Marc Tetel, Terrie Thomas and Joanne Turcotte for their helpful suggestions and comments on the manuscript. We are grateful to Abbott Laboratories, Diagnostic Division, Abbott Park, North Chicago, IL and Geoffrey Greene for providing the H222 monoclonal antibody.
R E F E R E N C E S
1 Barfield, R.J. and Chen, J.J., Activation of estrous behavior in ovariectomized rats by intracerebral implants of estradiol ben- zoate, Endocrinology, 101 (1977) 1716-1725.
2 Bast, J.D., Hunts, C., Renner, K.J., Morris, R.K. and Quadagno, D.M., Lesions in the preoptic area suppress sexual receptivity in ovariectomized rats with estrogen implants in the ventromedial hypothalamus, Brain Res. Bull., 18 (1987) 153-158.
3 Blaustein, J.D., Cytoplasmic estrogen receptors in rat brain: immunocytochemical evidence using three antibodies with dis- tinct epitopes, Endocrinology, 131 (1992) 1336-1342.
4 Blaustein, J.D., Estrogen receptor-immunoreactivity in rat brain:
311
rapid effects of estradiol injection, Endocrinology, 1342 (1993), in press.
5 Blaustein, J.D., Lehman, M.N., Turcotte, J.C. and Greene, G., Estrogen receptors in dendrites and axon terminals in the guinea pig hypothalamus, Endocrinology, 131 (1992) 281-290.
6 Blaustein, J.D. and Olster, D.H., Gonadal steroid hormone re- ceptor and social behaviors. In J. Balthazart (Ed), Advances in Comparative and Environmental Physiology, Springer-Verlag, Berlin/Heidelberg, 1989, pp. 31-104.
7 Blaustein, J.D. and Turcotte, J.C., Estrogen receptor-immuno- staining of neuronal cytoplasmic processes as well as cell nuclei in guinea pig brain, Brain Res., 495 (1989) 75-82.
8 Brann, D.W. and Mahesh, V.B., Regulation of gonadotropin secretion by steroid hormone, Frontiers Neuroendocrinol., 12 (1991) 165-207.
9 Chen, T.J. and Tu, W.W., Sex difference in estrogen and andro- gen receptors in hamster brain, Life Sci., 50 (1992) 1639-1647.
10 Ciaccio, L.A. and Lisk, R.D., Central control of estrous behavior in the female golden hamster, Neuroendocrinology, 13 (1973/74) 21-28.
11 Cintra, A., Fuxe, K., Harfstrand, A., Aganti, L.F., Miller, L.S., Greene, J.L. and Gustafsson, J.A., On the cellular localization and distribution of estrogen receptors in the rat tel-and dien- cephalon using monoclonal antibodies to human estrogen recep- tor, Neurol. Chem., 8 (1986) 587-595.
12 DeBold, J.F., Malsbury, C.W., Harris, V.S. and Malenka, R., Sexual receptivity: brain sites of estrogen action in female ham- sters, Physiol. Behav., 29 (1982) 589-593.
13 Dellovade, T.L., Blaustein, J.D. and Rissman, E.F., Neural distri- bution of estrogen receptor-immunoreactivity cells in the female musk shrew, Brain Res., 595 (1992) 189-194.
14 DonCarlos, L.L., Monroy, E. and Morrell, J.I., Distribution of estrogen receptor-immunoreactivity cells in the forebrain of the female guinea pig, J. Comp. Neurol., 305 (1991) 591-612.
15 Dudley, C.A., Lee, Y. and Moss, R.L,, Electrophysiological iden- tification of a pathway from the septal area the medial amygdala: sensitivity to estrogen and luteinizing hormone-releasing hor- mone, Synapse, 6 (1990) 161-168.
16 Floody, O.R., Blinn, N.E., Lisk, R.D. and Vomachka, A.J., Local- ization of hypothalamic sites for the estrogen-priming of sexual receptivity in female hamsters, Behav. Neurosci., 101 (1987) 309- 314.
17 Fox, C.A., Ross, L.R., Handa, R.J. and Jacobson, C.D., Localiza- tion of cells containing estrogen receptor-like immunoreactivity in the Brazilian opossum brain, Brain Res., 546 (1991) 96-105.
18 Gorski, J., Welshons, W.V., Sakai, D., Hansen, J., Walent, J., Kassis, J., Shull, J., Stack, G. and Campen, C., Evolution of a model of estrogen action, Recent Prog. Horm. Res., 42 (1986) 297-329.
19 Greene, G.L., Sobel, N.B., King, W.J. and Jensen, E.V., Immuno- cytochemical studies of estrogen receptors, J. Steroid Biochem., 20 (1984) 51-56.
20 Henry, W.W., Medlock, K.L., Sheehan, D.M. and Scallet, A.C., Detection of estrogen receptor (ER) in the rat brain using rat anti-ER monoclonal IgG with the unlabeled antibody method, Histochemistry, 96 (1991) 157-162.
21 Kallo, I., Liposits, Z., Flerko, B. and Coen, C.W., Immunocyto- chemical characterization of afferents to estrogen receptor-con- taining neurons in the medial preoptic area of the rat, Neuro- science, 50 (1992) 299-308.
22 King, W.J. and Greene, G.L., Monoclonal antibodies localize oestrogen receptor in the nuclei of target cells, Nature, 307 (1984) 745-749.
23 Krieger, M.S., Morrel, J.I. and Pfaff, D,W., Autoradiographic localization of estradiol-concentrating cells in the female hamster brain, Neuroendocrinology, 22 (1976) 193-205.
24 Lauber, A.H., Romano, G.J., Mobbs, C.V. and Pfaff, D.W., Estradiol regulation of estrogen receptor messenger ribonucleic acid in rat mediobasal hypothalamus: an in situ hybridization study, J. Neuroendocrinol., 2 (1990)606-611.
25 Law, T. and Meagher, W., Hypothalamic lesions and sexual behavior in the female rat, Science, 128 (1958) 1626-1627.

312
26 Lawson, N.O., Wee, B.E.F., Blask, D.E., Castles, C.G. Spriggs, L.L. and Hill, S.M., Melatonin decreases estrogen receptor ex- pression in the medial preoptic area of inbred (LSH/SsLak) golden hamsters, Biol. Reprod., 47 (1992) 1082-1090.
27 Lisk, R.D., Diencephalic placement of estradiol and sexual recep- tivity in the female rat, Am. J. Physiol., 203 (1962) 493-496.
28 Lisk, R.D., and Ferguson, D.S., Neural localization of estrogen- sensitive sites for inhibition of ovulation in the golden hamsters Mesocricetus auratus, Neuroendocrinology, 12 ( 19731 153- 160.
29 MacBride, R.L. and Sutin, J., Amygdaloid and pontine projection to the ventromedial nucleus of the hypothalamus. J. Comp. Neurol., 174 (1977) 377-396.
30 Malsbury, C.W., Kow, L-M. and Pfaff, D.W., Effects of medial hypothalamic lesions on the lordosis response and other behav- iors in female golden hamsters, Physiol. Behaz,., 19 (19771223-237.
31 Malsbury, C.W., Pfaff, D.W. and Malsbury, A.M., Suppression of sexual receptivity in the female hamster: neuroanatomical projec- tions from preoptic and anterior hypothalamic electrode sites, Brain Res., 181 (1981) 267-284.
32 Morin, L.P. and Feder, H.H., Intracranial estradiol benzoate implants and lordosis behavior of ovariectomized guinea pig, Brain Res., 70 (19741 95-102.
33 Masco, D.H. and Carter, H.F., Sexual receptivity in female rats after lesion or stimulation in different amygdaloid nuclei, Physiol. Behaz,., 24 (1980) 1073-1080.
34 Moss, R.L., Paloutzian, R.F. and Law, O.T., Electrical stimula- tion of forebrain structures and its effect on copulatory as well as stimulus-bound behavior in ovariectomized hormone-primed rats, Physiol. Behau., 12 (1974) 997-1004.
35 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordi- nates, Academic Press, New York, 1986.
36 Pfaff, D.W., Keiner, D.M., Atlas of estradiol-concentrating cells in the central nervous system of the female rat, J. Comp. Neurol., 151 (1973) 121-158.
37 Pfaff, D.W. and Schwartz-Giblin, S., Cellular mechanisms of female reproductive behaviors. In E. Knobil and J. Neill (Eds.), The Physiology of Reproduction, Raven Press, New York, 1988, pp. 1487-1568.
38 Pelletier, G., Liao., N., Follea, N. and Govindan, M.V., Mapping of estrogen-producing cells in the rat brain by in situ hybridiza- tion, Neurosci. Lett., 94 (1988) 23-28.
39 Powers, B. and Valenstein, E.S., Sexual receptivity: facilitation by medial preoptic lesions in female rats, Science, 175 (1972) 1003- 1005.
40 Rodriguez-Sierra, J.F. and Terasawa, E., Lesions of the preoptic
area facilitate lordosis behavior in male and female guinea pig,,, Brain Res. Bull., 4 (1979) 513-517.
41 Ross, J., Claybaugh, C., Clemens, L.G. and Gorski, R.A., Shorl latency of estrous behavior with intracerebraI gonadal hormones in ovariectomized rats, Endocrinology, 89 (19711 32-38.
42 Saper, C.B., Swanson, LW. and Cowan, W.M., r i le efferent connections of the ventromcdial nucleus of the hypotbalamus of the rat, J. Comp. Neurol., 169 (1976) 409-442.
43 Schaison, G. and Couzinet, B., Steroid control of gonadotropin secretion, J. Steroid Biochem. Mol. BioL, 40 (1991) 417-420.
44 Simerly, R.B., Chang, C., Muramatsu, M and Swanson, L.W.. Distribution of androgen and estrogen receptor mRNA-contain- ing cells in the rat brain: an in situ hybridization study. Z Comp. Neurol., 294 (1990) 76-95.
45 Stumpf. W.E., Sar, M. and Keefer, D.A., Atlas of estrogen target cells in rat brain. In W.E. Stumpf and LD. Grant (Eds.), Anatomical Neuroendocrinoh)gy, Karger, Basel, 1975, pp. 104-109.
46 Stumpf, W.E., Morin, J.K., Ennis, B.W., Zielinski, J.E. and Hochberg, R.B., Utility of [16a-1251]iodoestradiol for autoradiog- raphy for the study of cellular and regional distribution of recep- tors, J. Histochem. Cytochem,, 35 (19871 87-92.
47 Takahashi, L.K. and Lisk, R,D., Estrogen action in anterior and ventromedial hypothalamus and the modulation of heterosexual behavior in female golden hamsters, Physiol. Behat:., 34 (19851 233-239.
48 Tobet, S.A., Basham, M., Fox, T.O. and Baum, M.J., Estrogen- receptor immunoreactive neurons in the forebrain of adult male and female ferrets, Soc. Neurosci. Abstr., 561.20, (1991).
49 Turcotte, J.C. and Blaustein, J.D., Immunocytochemical localiza- tion of mesencepbalic estrogen and progestin receptor-containing cells in female guinea pig, J. Comp. Neurol., 328 (19931 76-87,
50 Warembourg, M., Radioautograpbic localization of estrogen-con- centrating cells in the brain and pituitary, of the guinea pig, Brain Res., 123 (19771 357-362.
51 Welshons, W.V., Liberman, M.E. and Gorski, J., Nuclear local- ization of unoccupied oestrogen receptors, Nature, 307 (19841 747-749.
52 Wong, M. and Moss, R.L., Modulation of single-unit activity in the rat medial amygdala by neurotransmitters, estrogen priming, and synaptic inputs from the hypothalamus and midbrain, Synapse, 10 (19921 94-102.
53 Yanase, M. and Gorski, R.A., Sites of estrogen and progesterone facilitation of lordosis in the spayed female rat, Biol. Reprod.. 15 (19761 536-543.



![Modulation Of [35S]Tert Butylbicyclophosphorothionate Binding By Somatostatin In Rat Hypothalamus](https://img.pdfslide.net/doc/110x75/631ba0d07abff1d7c20b0671/modulation-of-35stert-butylbicyclophosphorothionate-binding-by-somatostatin-in.jpg)

























