Embed Size (px)
Citation preview

Vol. 7, No. 11MOLECULAR AND CELLULAR BIOLOGY, Nov. 1987, p. 4075-40810270-7306/87/114075-07$02.00/0Copyright © 1987, American Society for Microbiology
Simultaneous Expression of Two P-Glycoprotein Genes in Drug-Sensitive Chinese Hamster Ovary Cells
JANE A. ENDICOTT, PETER F. JURANKA, FARIDA SARANGI, JAMES H. GERLACH,tKATHRYN L. DEUCHARS, AND VICTOR LING*
The Ontario Cancer Institute, The Princess Margaret Hospital, and The Department of Medical Biophysics, University ofToronto, Toronto, Ontario M4X 1K9 Canada
Received 22 May 1987/Accepted 7 August 1987
Overexpression of P-glycoprotein is characteristic of multidrug-resistant cells. We analyzed four P-glycoprotein transcripts that are simultaneously expressed in a drug-sensitive Chinese hamster ovary cell line.We concluded that these transcripts are encoded by two distinct members of a P-glycoprotein multigene family,each of which has two alternative polyadenylation sites. A comparison of the two hamster sequences with thesingle reported human and mouse P-glycoprotein cDNA sequences demonstrates that P-glycoprotein is a highlyconserved protein, that the hamster multigene family is undergoing concerted evolution, and that differencesbetween gene family members are maintained across species. These conserved differences suggest that theremay be functional differences between P-glycoprotein molecules.
The presence of multidrug-resistant cells in human tumorsand their selection and proliferation during chemotherapymay be major obstacles to cancer treatment (9). Mammaliancell lines selected for resistance to a single cytotoxic agentoften develop pleiotropic resistance to unrelated drugs (9).Since many of the drugs involved are commonly used inchemotherapy, such cell lines have proven useful as in vitromodels for the study of multidrug resistance in humantumors.A consistent alteration in multidrug-resistant cells is the
overexpression of P-glycoprotein, an integral membraneprotein of 170 kilodaltons (9). A P-glycoprotein cDNA clone,pCHP1 (19), has been isolated by using a P-glycoprotein-specific monoclonal antibody (14). This clone was used toisolate the cDNA clones discussed in this paper. Indepen-dently, an in-gel renaturation technique (20) has also beenused to isolate amplified sequences from a drug-resistanthamster cell line (21). These sequences were used to isolatehomologous human (2) and mouse (10) cDNA clones. Thehuman cDNA sequence (mdrl) was reconstructed fromseveral overlapping clones isolated from a multidrug-re-sistant cell line cDNA library. The mouse sequence wasdetermined from a single full-length cDNA clone isolatedfrom a drug-sensitive cell line. The human cDNA sequence(mdrl) encodes a protein detected by P-glycoprotein-specificmonoclonal antibodies (26). We show that the human andmouse genes are homologs of different hamster P-glyco-protein (pgp) genes. We refer to the human and mouse mdrsequences as pgp genes in this communication to facilitatecomparison of the P-glycoprotein genes across species.
P-glycoprotein contains a tandem duplication in structure,with each half consisting of six transmembrane domains anda cytoplasmic domain containing the consensus sequencesfor an ATP-binding site (Fig. 1) (2, 8, 10; J.A.E. and P.F.J.,unpublished observations). Amino acid and structural ho-mology extends throughout each half of P-glycoprotein toHlyB, a membrane transport protein that is required for the
* Corresponding author.t Present address: The Ontario Cancer Treatment and Research
Foundation, Kingston Regional Cancer Centre, Kingston, OntarioK7L 2V7, Canada.
transport of alpha-hemolysin (a 107-kilodalton protein) frombacterial cells (8, 16). This homology is sufficiently strikingthat P-glycoprotein can be regarded as a tandem duplicationof the HlyB molecule (8). P-glycoprotein is thought to be amembrane "pump" which actively transports drug mole-cules from the cell, thereby reducing intracellular drugconcentrations in multidrug-resistant cells (2, 8, 10).We present the partial cDNA and deduced amino acid
sequences of four different P-glycoprotein transcripts thatare simultaneously expressed in drug-sensitive Chinese ham-ster ovary (CHO) cells. We conclude that these transcriptsare encoded by two distinct members of a P-glycoproteinmultigene family (pgpl and pgp2), each of which has twoalternative polyadenylation sites. This is the first directconfirmation of the P-glycoprotein multigene family hypoth-esis (4, 19). The gene family members are distinguished bynucleotide sequence differences in both the protein-codingand 3'-untranslated regions. We also compare the two ham-ster P-glycoprotein cDNA sequences with the single P-glycoprotein cDNA sequences isolated from mouse andhuman cells (2, 10). This comparison shows that the humanP-glycoprotein sequence (mdrl) is more homologous tohamster pgpl and that the mouse sequence has greaterhomology to hamster pgp2.
MATERIALS AND METHODScDNA cloning. Clones were isolated from an Okayama and
Berg pCD vector library prepared from the drug-sensitiveCHO cell line E29Pro' (7). This cell line exhibits no charac-teristics of pleiotropic drug resistance and does not over-express P-glycoprotein. pCHP1 (19) was used as a P-glycoprotein-specific probe to screen approximately 106clones in two independent screenings of the cDNA library. Atotal of 24 pCHP1-positive clones were isolated. Cloneswere initially identified by Maxam and Gilbert sequencing(17) that extended from the poly(A) tail. Four clones (pL2,pL20, pL28, and pL34) (Fig. 1) were subcloned into eitherM13mp8 or M13mp9 phage as PstI or BamHI fragments.Single-stranded DNA was sequenced by primer extension,using the dideoxy method of Sanger et al. (22). The entirenucleotide sequence was determined on both strands for twoindependent subclones. The nucleotide sequences were as-
4075

4076 ENDICOTT ET AL.
2 34 56 A B
1968bp
1689 bp
2 321 bp
826bppL2 1- 1
FIG. 1. Schematic representation of P-glycoprotein. The pro-posed transmembrane regions are marked by numbered boxes. Thetwo consensus sequences that form the potential ATP-binding foldcommon to P-glycoprotein, HlyB, and the bacterial periplasmictransport systems are marked by solid boxes lettered A and B. Thepositions of the four cDNA clones, pL34, pL28, pL20, and pL2, arealso shown. bp, Base pairs.
sembled by using the data base programs of Staden (24).Clones were classified as either pgpl or pgp2 transcripts onthe basis of sequence homologies in both the 3'-coding and3'-untranslated regions characteristic of each gene.
Nucleic acid sequence analysis. The 3'-untranslated regionswere analyzed by using the National Biomedical ResearchFoundation RELATE program (3) and the Staden DIAGONprogram (24). The nucleotide sequence divergence of theprotein-coding region was analyzed by the method of Perleret al. (18).
RESULTS
Two P-glycoprotein genes simultaneously expressed in drug-sensitive CHO cells. The partial nucleotide sequences of twoP-glycoprotein genes from drug-sensitive CHO cells areshown in Fig. 2. pgpl is constructed from the overlappingclones pL28 and pL34, and pgp2 is constructed from pL20and pL2. P-glycoprotein has a tandem internal duplication instructure, with each half encoding six potential transmem-brane domains and a cytoplasmic domain containing theconsensus sequences for a putative ATP-binding site (Fig.1). The two gene transcripts encode the six potential trans-membrane regions and the cytoplasmic domain of the C-terminal half of P-glycoprotein. The highest concentrationsof nucleotide differences in the coding region are seen at the5' ends of the cDNAs in the sequences that encode the sixpotential transmembrane regions and the short linkingpeptides between them. The nucleotide sequence that en-codes the cytoplasmic domain is highly conserved; only 36changes are found between the two hamster transcripts in atotal sequence length of 873 bases. To align the sequences atthe 3' end, an insertion of six bases has to be made in thepgp2 sequence.
Transcripts from pgpl and pgp2 show alternate poly-adenylation, producing 3'-untranslated regions of four dif-ferent lengths (Fig. 2). There are hexamer sequences 10 to 30nucleotides 5' to each polyadenylation site that may act as
signal sequences for polyadenylation (1). Downstream fromthe pL20 and pL34 polyadenylation sites, pL2 and pL28have a G+T-rich sequence which is believed to be animportant consensus sequence on the 3' side of a potentialpolyadenylation site (1).For pgpl, 14 clones were polyadenylated at site 1 (pL34-
like) and 2 clones were polyadenylated at site 2 (pL28-like).For pgp2, 3 clones were polyadenylated at site 1 (pL20-like)and 5 clones were polyadenylated at site 2 (pL2-like). Twoscreenings of the cDNA library failed to produce any clonesthat were not either pgpl or pgp2. This finding suggests that
these sequences are the major P-glycoprotein transcripts inthis drug-sensitive cell line. It is possible that othermultigene family members may be expressed at very lowlevels. The distribution of differences between the pgpl andpgp2 sequences strongly suggests that they are transcriptsfrom two different P-glycoprotein genes and do not resulteither from differential splicing of a primary RNA transcriptor from genomic rearrangement of a single gene.
Nucleotide sequence homology with human and mouseP-glycoprotein gene transcripts. A comparison of the singleP-glycoprotein cDNA sequences isolated from human andmouse cells with the two hamster P-glycoprotein gene tran-scripts shows that the P-glycoprotein multigene family ishighly conserved across species. This homology can bedemonstrated by comparing both the protein-coding and3'-untranslated regions.The 3'-untranslated regions of the human and mouse
P-glycoprotein gene transcripts were compared with the twohamster P-glycoprotein cDNA sequences in the correspond-ing region by using the Staden DIAGON program (resultsnot shown). The mouse 3'-untranslated region was shown tobe more homologous to that of hamster pgp2, while thehuman 3'-untranslated region was more homologous to thehamster pgpl sequence. On the basis of this analysis, thehuman sequence was tentatively identified as the humanhomolog of hamster pgpl, and the mouse sequence wasidentified as the mouse equivalent of hamster pgp2. Toquantify the homology between the four 3'-untranslatedregions, the sequences were also analyzed by using theNational Biomedical Research Foundation RELATE pro-gram (Table 1). The segment comparison scores were calcu-lated by using the unitary matrix with a bias of +2, afragment length of 15, and 500 random runs per comparison.The scores are expressed in units of standard deviation (SD),the larger values corresponding to greater homology. Fournucleotide sequences were used: hamster pgpl (1716 to1968), hamster pgp2 (1966 to 2321), human pgpl (3841 to4222), and mouse pgp2 (3829 to 4189). The DIAGON resultswere confirmed; the hamster pgpl 3'-untranslated regionshowed greater homology to the human 3'-untranslatedregion (score = 13.0 SD units), and the hamster pgp2sequence showed strong homology to the mouse 3'-untranslated region (score = 17.1 SD units). The two ham-ster sequences have much less homology between their3'-untranslated regions (score = 3.2 SD units). The proba-bility of obtaining a score of >6 SD units is <10-9, whereasthe probability of a score of >10 SD units is <10-23.
Interestingly, a large insertion had to be made in thehamster pgp2 3'-untranslated region to maximize alignmentwith the mouse sequence. The hamster pgp2 sequence doesnot contain the (AAAC)6 repeat present in the mouse 3'-untranslated region. It is probable that the mouse has gainedthis sequence since the gene family diverged.The nucleotide sequence divergence in the protein-coding
region was analyzed as described by Perler et al. (18), amethod which is designed to consider differences in nucleo-tide sequence between two closely related proteins. Theanalysis considers nucleotide changes that produce a changein an amino acid (replacement substitutions) separately fromthose changes that produce a synonymous codon (silentsubstitutions). The nucleotide sequence encoding the C-terminal half of P-glycoprotein was used in each comparison,which in each case was taken to begin at the first potentialtransmembrane domain of the C-terminal half of the dupli-cation. To avoid having to make a six-base insertion in thepgp2 sequences to align the last 24 nucleotides of the coding
NH2-[1 2 34 56 A B
pL34 1-
pL28
pL20 F
MOL. CELL. BIOL.

VOL. 7. 1987 TWO P-GLYCOPROTEIN GENES 4077
LTGC AGA CTT GTC ATG ATG CAG ACT AGA GCA MAT GAA GTT GAA CTG GCA ACT GAA GCT GAT GGA TCC CAG ACT GAC ACC ATI GCC TCT GAA hIG ALT ICA GAA GAA ITT MAA TCC CCT TCA 120[CYS ARG LEU VAL MET M4ET GIN THR ARG GILY ASN GLU VAL GLU LEU GILY SER GLU ALA ASP GILY SIR GLN SIR ASP THR ILE ALA SER GLU LEU THR SER GLU GLU PIIE ITS SER PRO SIR 40pL 20GTG AGG MAA TCA ACT TGT ACA AGC ATC TGT GGT TCC CAA GAC CAA GAG AGG ACA GTC ACT GTG MAA GAG GCC CAG GAI GAA CAT GIG CCI CTG CTT TCC TIT TGG GCG ATC CIG MAG CTA 240VAL ARC ITS SER THR CYS ARC SER ILE CYS GLY SIR GIN ASP GIN GLU ARG ARG VAL SIR VAL ITS GLU AL,. GIN ASP GLU ASP VAL PRO LEU VAL SIR PHE TRP GLY ILE LEU ITS LEU 60
TILE ILE ILEITSA T G G A T C C A AG GCC T A CA A A TMA G TG A AC 107
MAC ATA ACT GAA IGG CCT TAT TTA GTT GTT GCT GTA CTT TGT GCT GTT ATA MAT GCA TGT ATG CAA CCC GIC TTC ICC AlA GTA TTT TCA GGG ATC ATA GCC GTT TTC ACA AGA CAT CAT 360ASN ILE THR GILU TRP PRO TYR LEU VAI VAL GILY VAI LEU CYS ALA VAL TILE ASN GLY CYS MET GLN PRO VAL PHE SER ILE VAL PHE SIR GILY ILE TILE GILY VAL PHE THR MRG ASP ASP 120
ASP CLU ARC HIS ASP SIR LEU ILE LEU VAL SIR ILE PHE LEU 75T GATG C T G A C C C TA G A C T CGGC C A T T CT 227
GCA CCC MAA ACC MAA CAA CAG MAT TGT MAT TTG TTT TCA CTG TTT TTT CTG GTC ATG GGC ATG ATT TGT TTT GTT ACA TAT TTC TTC CAG CCC TTC ACA TTT CCC MAA GCT GCA GAG ATC 460ASP PRO ITS THR ITS GLN GLN ASN CYS ASN LEU PHE SER LEU PHE PHE LEU VAL M4ET GILY MET ILE CYS PHE VAL THR TYR PHI PHI GLN CIY PHI THR PHE CIT LYS ALA CIT GILU ILE 160
VAL ASN PRO LYS THR ASN GILY IIIGCG ACC A C C A C AC TCG347
CTC ACC AAC CGA CTC CGA TAC ATG GTT TTC MAA TCC ATG CTG AGA CAC CAT ATA ACC TGG TTT CAT CAT CAT ACA MAT ACC ACT CCC GCA TTC ACC ACC ACC CTA CCC ACT CAT CCC GCT 600LEU THR ITS ARC LEU ARC TYR MET VAL PHE ITS SIR MET LEU ARC GIN ASP ILE SIR TRP PHE ASP ASP HIS ARC ASN SER THR CIT ALA LEU THR THR ARC LEU ALA SIR ASP ALA ALA 200
GIN THR CIT ALA VAL F-TILE ILE LEU ALA 155C A CACGAGIC A TC IA A C C CT CA 467MIT GTT MAA CCC GCT ATG ACT TCA AGG CTT GCCT CCA ATT ACC CAG MTGTA GCIA MAT CTT CCC ACA CCA ATC ATC ATA TCC CTA GTC TAT CCC TGG CAG TTA ACA CTT TTA CTT GTA GIG 720ASN VAL ITS CIT ALA MET SIR SIR ARC LEU ALA CIY ILE THR GIN ASN VAL ALA ASN LEU CIT THR CIT ILE TILE ILE SIR LEU VAL TYR CIT TRP GIN LEU THR LEU LEU LEU VAL VAL 240
VAL ILE ALA ILE ALA VAL VAL MET CIT 195TC C A GCCA GCA AC CGT A A A A T C CT 5607
ATT GCCI CCA CTC ATT AlA CTA ACC CCC AIG ATG GAG AIG MAA GTG TTG TCT CCT CAA CCT CTG MAA GAC MAG MAA GAG CIA GAA GTC TCT CCC MAG ATT CCT ACT GAA GCA AlA GAG MAC 640ILE ALA PRO LEU ILE ILE LEU SER CIT MET MET CLU M4ET ITS VAL LEU SIR CIT GIN ALA LEU ITS ASP ITS IYS GLU LEU GLU VAL SIR CIT ITS ILE ALA THR GLU ALA ILE GLU ASN 260O
235707
TIC CCC ACT GTC GTC TCT TTG ACT CCC GAG CAG MAG TTT GAA MAT ATG TAT CCC CAG ACC TTC CAG ATA CCA TAC ACA MAT GCT CTG MAG MAA CCA CAC GTC TTT CCC ATC ACA TTC TCC 960PHE ARC THR VAL VAL SER LEU THR ARC CIU GIN LYS PHE CIU ASN MET TYR ALA GIN SER LEU GIN ILE PRO TTR ARC ASN ALA LEU ITS LYS ALA HIS VAL PHE CIT TILE THR PHI SIR 320
ARC CLU LEU LEU ILE 27?5T CCCG T A A C A A C A T627
TTC ACC CAG CCC ATG ATG TAT TTT TCG TAT CCT CCT TGT TIC CCC TTT CCT CCC TAC TTG GTG CCA CAT CAA ATC ATG ACT ITT GAA MAT CIT AIG TIC CIA TIT TCA GCCI CIT GTC TIC 1060PHI THR GIN ALA MET MET TTR PHIE SER TTR ALA ALA CYS PHE ARC PHE CIT ALA TTR LEU VAL ALA HIS GIN TILE MET THR PHI CIU ASN VAL MET LEU VAL PHI SIR ALA VAL VAL PHE 360
MET VAL GIN VAL MET ILE VAL 315C T CCI T C T T C C 947
CCT CCC AlA CCA GCCI CCC MT CCC ACC TCA TTT GCCI CCT GAC TAT CCC MAA CCC MAA GIG TCT CCA ICC CAT ATC ATC AGG ATC ATG GAA MAA ATC CII TCC AlT GAC ACT TAC ACC ACC 1200CLY ALA ILE ALA ALA CIT ASN ALA SER SER PHI ALA PRO ASP TTR ALA ITS ALA ITS VAL SIR ALA SIR HIS ILE TILE ARC TILE MET GLU ITS ILE PRO SER ILE ASP SER TYR SIR THR 400
CIT THR ASN 355C ACA C C C A 1067ACA CCC CTG MAG CCT MAT TGG TIC GAA CCA MAT GIG MAA TTT MAT GAA GTT GTG TIC MAC TAT CCC ACC CGA CCI GAC ATC CCA GTG CTT CAG CCC CTG ACC CTG GAG GIG MCG MAG CCC 1320ARC CIY LEU ITS PRO ASN TRP LEU CIU CIT ASN VAL ITS PHE ASN GLU VAL VAL PHI ASN TYR PRO THR ARC PRO ASP ILE PRO VAL LEU GIN CIT LEU SIR LEU CIU VAL ITS ITS CIT 440
VAL ASN 395
CAC ACG CTC CCC CTG GIG CCC ACC ACT CCC ICC CCC MAG ACC ACA GIG GTC CAG CTG CTC GAG CGG TIC TAC GAC CCC ATG CCC CCA ACA GIG TTI CIA CAT CCC MAA GAA ATA MC CMA 1440CLN THR LIU ALA LEU VAL CIT SIR SER CIT CYS CIT LYS SIR THR VAL VAL GIN LEU LEU GLU ARC PHE TYR ASP PRO MET ALA CIT THR VAL PHI LEU ASP CIT ITS GLU ILE ITS GIN 460
435A C C T 1307
CTG MAT GTC CAC ICC CTC CGA GCA CAl TTA CCC All GIG ICC CAA GAG CCC ATC CTG ITT GAC TGC ACC AlT GCT GAG MAC ATC CCC TAT CCA GAC MAC ACC CCC GTC GIG ICC CAG GAl 1560LEU ASN VAL GIN TRP LEU ARC ALA HIS LIU CIT ILE VAL SER GIN GLU PRO ILE LEU PHI ASP CYS SER TILE ALA GLU ASN ILE ALA TYR CIT ASP ASN SER ARC VAL VAL SIR GIN ASP 520
475C A A 1427
CAC ATC GAG AGG GCCC CCC MCG GAG GCCC CAAC CAC CAG TIC All GAG TCT CTG CCI GAC MAA TAC MAC ACC ACA CIA CCA GAC MAA CCC ACT CAC CTG ICC CCI CCC CAC MAA CAA CCC 1660GLU ILE CLU ARC ALA ALA ITS CLU ALA ASN HIS GIN PHI ILE CIU SIR LEU PRO ASP ITS TIN ASN THR ARC VAL CIT ASP ITS CIT THR GIN LEU SER CIT CIT GIN ITS GIN ARC 560
515I A 1547
ATC CCC AlA GCA CCC CCC CTC CTC ACA lAG CCI CAC All TIC CTC CTG CAT GAA CCA ACA ICA CCC CTG CAT ACA GAG ACT GAA MCG CII GTC CAC GAA CCC CTG GAC MAA CCC AGG GAA 16000TILE ALA ILE ALA ARC ALA LEU VAL ARC GIN PRO HIS ILE LEU LEU LEU ASP CLU Al2A THR SIR ALA LEU ASP THR GLU SIR GLU ITS VAL VAL GIN GILU ALA LEU ASP ITS ALA ARC GIU 600
555C 1667
CCC CCC ACC ICC ATl GIG ATC GCCI CAC CCC CTG ICC ACC ATC CAG MAC GCA GAC TIC ATC GTC CII All CAG MIT CCC MAG GTC MCG GAG CAT CCC ACC CAC CAG CAG CIC CTG GCA CAG 1920CIT ARC THR CYS ILE VAL ILE ALA HIS ARC LEU SIR THR ILE GIN ASN ALA ASP LIU TILE VAL VAL ILE GIN ASN CIT ITS VAL ITS GLU HIS CIT THR HIS GIN CLN LEU LEU ALA GIN 640
SIR VAL 571ACT GIG A IL,A ACTGTGTTMAATAITITTTTTMTATTTGIATIMACACATACCATTTAMCCMAAGITMAMAACGAATACGTACCAGAMTACCAGTT 1606
MAA CCC ATC TAT TIC TCC ATGGCCI-----CAG GCCI CCA GCA MCG CCC TIG TAA ACTGTGACTGTTTGMAATGCCTAAGTGTTTTTAATATCTCTTCTTMAAATCCGCACTTAACCAMAGTTTMACAAGTGAACACTGTTG 2055ITS CIT ILE TYR PHI SIR MET VAL - GIN ALA CLY ALA ITS ARC LEU..655
pL34 -M pL126
CAGITTTMTTATTCMACCCTACTT TGTTMAATGATTATMAAACTTGTAMAATGTACTTMAAATATCCATAMAGTCCCTATMMAACTAMATGITTCAIATATMTGCATCTTGTCTGITCTGTTGTAITMATTTATATATTTTTCCAMACCAGATCCT 2373PL120
CIMATMITITACCI- ~~~~~~~~~~~~~~~~~~~~~~~~2409GAAATTACTTCATAMAATGGATTTTA!g
FIG. 2. Sequences of two P-glycoprotein genes isolated from drug-sensitive CHO cells. pgpl is constructed from the cDNA clones pL34and pL28, and pgp2 is constructed from pL20 and pL2. For pgpl, only the residues that differ from pgp2 are marked. The ends of each cloneare indicated. pgpl is the upper sequence and begins at residue 254 in the lower pgp2 sequence. To align pgp2 with pgpl. a six-base insertionhad to be made in pgp2 at the 3' end, which is marked with dashes in the nucleotide sequence. ***, Termination codon; _, potentialpolyadenylation signal.

4078 ENDICOTT ET AL.
TABLE 1. Segment comparison scores for P-glycoprotein3'-untranslated regions
Homology (SD units)P-glycoprotein
gene Hamster Hamster Human Mousepgpl pgp2 pgpl pgp2
Hamster pgpl 3.2 13.0 7.4Hamster pgp2 8.7 17.1Human pgpl 9.9Mouse pgp2
region, we truncated each sequence after the triplet encodingvaline (see Fig. 3). The sequences used were hamster pgpl(residues 3 to 1691), hamster pgp2 (256 to 1944), human pgpl(2125 to 3813), and mouse pgp2 (2119 to 3807).The corrected replacement-site percent divergence scores
(Table 2) confirm the results shown in Table 1; the hamsterpgpl sequence has greater homology to the human sequence(5.2%) than to the mouse sequence (8.3%), and the hamsterpgp2 sequence has greater homology to the mouse sequence(5.1%) than to the human sequence (8.4%). Of interest arethe corrected percent divergence results generated by con-sidering only silent nucleotide changes. Since silent substi-tutions produce no change in the amino acid sequence, suchchanges would not be selected for or against at the proteinlevel and should reflect the time since divergence of thegenes. Minor variations may result from variations in theinherent rate of mutation in different species (15). It wouldbe expected that the divergence scores for comparisons ofpgpl and pgp2 would be similar, reflecting the time since thegeneration of these genes. The corrected silent-site percentsequence divergence scores are consistent with this predic-tion for the comparisons of hamster pgpl with mouse pgp2(47.6%), hamster pgp2 with human pgpl (46.4%), and humanpgpl with mouse pgp2 (55.6%) but not for the comparison ofhamster pgpl with hamster pgp2 (21.6%). This findingstrongly suggests that the two hamster genes have notevolved independently. Concerted evolution (27) has beenreported for the alpha-globin (13) and mouse immunoglobu-lin gamma-chain (12) multigene families. Several mecha-nisms, such as gene conversion and double unequal cross-ing-over, have been postulated for this process (12).
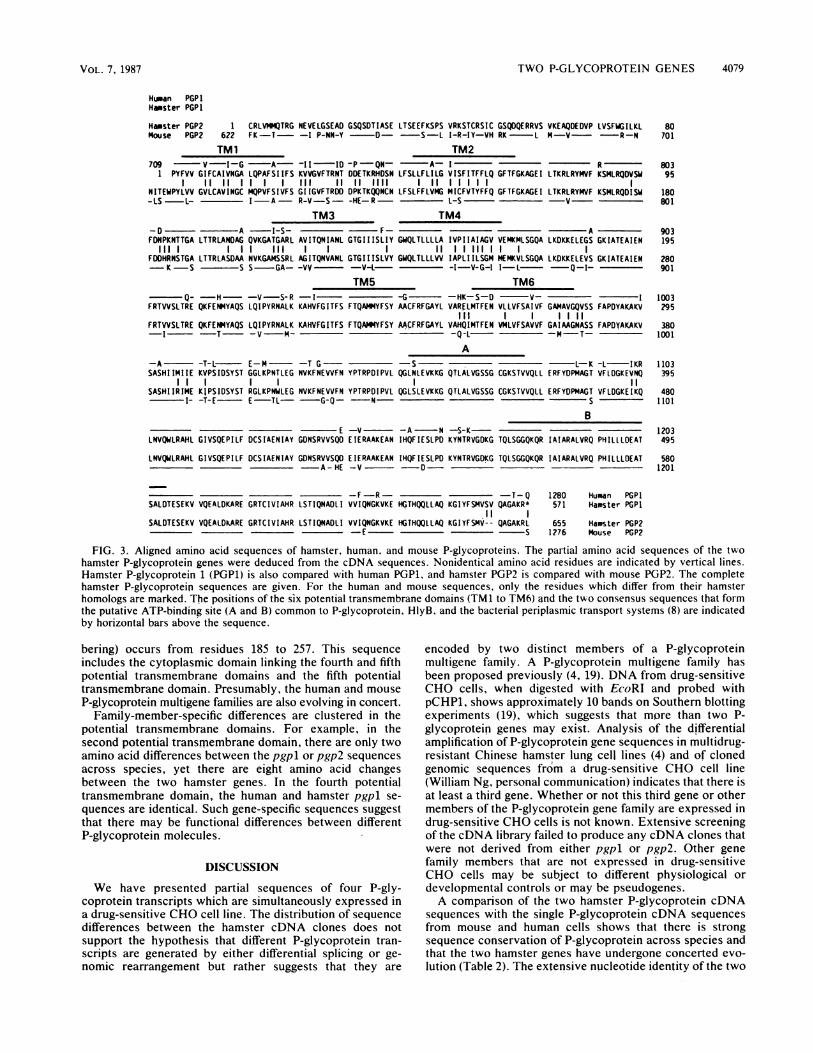
Protein sequence homology. The partial amino acid se-quences of two P-glycoproteins from drug-sensitive CHOcells are shown in Fig. 2. These amino acid sequencescontain six putative transmembrane domains (8). The orien-tation of P-glycoprotein in the plasma membrane has beendetermined by localizing the C terminus to the cytoplasmwith monoclonal antibodies (14). The longer stretches ofamino acids that join transmembrane regions 2 to 3 and 4 to5 are in the cytoplasm, and only short stretches of aminoacids are outside the cell (see Fig. 3).The regions coding for the six potential transmembrane
domains contain the highest concentrations of nucleotidedifferences between the two hamster genes. A comparison ofthe deduced amino acid sequences, however, shows thatmost of these changes either are silent (i.e., producing nochange in the protein sequence) or result in replacement witha conserved amino acid. Within the potential transmembranedomains, only five amino acid changes are not conservedchanges (as defined by the mutation data matrix for aminoacid replacements [3]).The C-terminal cytoplasmic domain of P-glycoprotein,
which is even more highly conserved, contains two blocks ofamino acid sequence (marked A and B in Fig. 3), each of
MOL. CELL. BIOL.
which contains consensus sequences thought to form anucleotide-binding fold. In the C-terminal cytoplasmic do-main, there are only 13 amino acid changes between the twohamster sequences. Three of these changes occur at the Cterminus. pgp2 lacks the bases coding for pgpl serine-564and valine-565, and the codon for the terminal leucine (TTG)is a stop codon (TAG) in pgpl. Seven amino acid changesare clustered close to the sixth potential transmembranedomain. The threonine-to-tryptophan exchange at position322 in pgpl is interesting because it requires a change in eachbase of the codon.Homology with mouse and human P-glycoproteins. Figure 3
shows the alignment of the amino acid sequences of thehamster P-glycoproteins with those of the human and mouseP-glycoproteins in the corresponding region. The most ex-tensive sequence conservation is within the cytoplasmicdomain. Differences among the four P-glycoprotein se-quences are clustered into three regions. The first regionfollows the sixth potential transmembrane domain, the sec-ond region is shorter and follows the glycine-rich sequenceof the potential ATP-binding fold, and the third regionprecedes the second consensus sequence of the putativeATP-binding site.
P-glycoprotein shows extensive homology to the proposedATP-binding domain of one of the proteins of bacterialperiplasmic binding-protein-dependent transport systems(V. Ling, P. F. Juranka, J. A. Endicott, K. L. Deuchars, andJ. H. Gerlach, in P. V. Wooley and K. D. Tew, ed.,Mechanisms ofDrug Resistance in Neoplastic Cells, vol. 9,in press). The extent of amino acid sequence conservation,however, suggests that further functional constraints, inaddition to bind'ing ATP, must exist (8). The cytoplasmicdomain of P-glycoprotein has greatest homology to HlyB, acomponent of the alpha-hemolysin active transport system(8), and thus may have evolved from a composite of ancientprotein domains capable of ATP binding and active molec-ular transport.
Analysis of the C-terminal nucleotide sequences of P-glycoprotein transcripts from human, mouse, and hamstercells strongly suggests that the two hamster P-glycoproteingenes have evolved by concerted evolution (Table 2). Thisconcerted evolution is reflected in the amino acid sequence(Fig. 3). In the cytoplasmic domain, the two hamster se-quences are identical from glutamine-395 in hamster pgpl tothe position of the two-amino-acid gap required to align thesequences close to the C terminus. This sequence includesthe region just proximal to the B region of the potentialATP-binding site where there is a clustering of differencesbetween the human and mouse sequences and the hamstersequence. A second example (with the hamster pgpl num-
TABLE 2. Sequence divergence within the P-glycoproteinmultigene familya
Sequence divergence (%)P-glycoprotein
gene Hamster Hamster Human Mousepgpl pgp2 pgpl pgp2
Hamster pgpl 6.8b 5.2 8.3Hamster pgp2 21.6 8.4 5.1Human pgpl 45.9 46.4 9.2Mouse pgp2 47.6 41.6 55.6
a The upper right side of the table shows the corrected percent divergencescores for replacement nucleotide changes; the lower left side of the tableshows the corrected percent divergence scores when only silent nucleotidechanges are considered.
I Corrected percent divergence (18).
TWO P-GLYCOPROTEIN GENES 4079
Human PGP1Hamster PGP1
Hamster PGP2 1 CRLVMMQTRG NEVELGSEAD GSQSDTIASE LTSEEFKSPS VRKSTCRSIC GSQDQERRVS VKEAQDEDVP LVSFWGILKLMouse PGP2 622 FK-T- -1 P-NN-Y D- -S-L I-R-IY-VH RK L M-V- -R-N
TM1 TM2709 V -I-G - A- I ID -P QN- A- I R
1 PYFVV GIFCAIVNGA LQPAFSIIFS KVVGVFTRNT DDETKRHDSN LFSLLFLILG VISFITFFLQ GFTFGKAGEI LTKRLRYMVF KSMLRQDVSWI 11 11 I I I I III 11 11 1111 I 1! I I I I
NITEWPYLVV GVLCAVINGC MQPVFSIVFS GIIGVFTRDD DPKTKQQNCN LFSLFFLVMG MICFVTYFFQ GFTFGKAGEI LTKRLRYMVF KSMLRQDISW-LS -L- I-A- R-V-S- -HE-R L-S V
TM3 TM4-D A -I-S- F- AFDNPKNTTGA LTTRLANDAG QVKGATGARL AVITQNIANL GTGIIISLIY GWQLTLLLLA IVPIIAIAGV VEMKMLSGQA LKDKKELEGS GKIATEAIEN
III I I III I 11 I I III I I IFDDHRNSTGA LTTRLASDAA NVKGAMSSRL AGITQNVANL GTGIIISLVY GWQLTLLLVV IAPLIILSGM MEMKVLSGQA LKDKKELEVS GKIATEAIEN-K-S S S-GA- -VV -V-L -I-V-G-I I-L -Q-I-
TM5 TM6Q- H- -V -S-R -I -G -HK-S-D V- I
FRTVVSLTRE QKFENMYAQS LQIPYRNALK KAHVFGITFS FTQAMMYFSY AACFRFGAYL VARFLMTFEN VLLVFSAIVF GAMAVGQVSS FAPDYAKAKVIII 11
FRTVVSLTRE QKFENMYAQS LQIPYRNALK KAHVFGITFS FTQAMMYFSY AACFRFGAYL VAHQIMTFEN VMLVFSAVVF GAIAAGNASS FAPDYAKAKV-I -T -V-M- -Q-I -M T-
A-A -T-L E-M -T G- -S L-K -L-IKRSASHIIMIIE KVPSIDSYST GGLKPNTLEG NVKFNEVVFN YPTRPDIPVL QGLNLEVKKG QTLALVGSSG CGKSTVVQLL ERFYDPMAGT VFLDGKEVNQ
II I I I I 11SASHIIRIME KIPSIDSYST RGLKPNWLEG NVKFNEVVFN YPTRPDIPVL QGLSLEVKKG QTLALVGSSG CGKSTVVQLL ERFYDPMAGT VFLDGKEIKQ
I- -T-E E-TL- -G-Q- -N- S
BE -V -A N -S-K-
LNVQWILRAHL GIVSQEPILF OCSIAENIAY GDNSRVVSQO EIERAAKEAN IHQFIESLPD KYNTRVGDKG TQLSGGQKQR IAIARALVRQ PHILlLDEAT
LNVQWLRAHL GIVSQEPILF DCSIAENIAY GDNSRVVSQD EIERAAKEAN IHQFIESLPD KYNTRVGDKG TQLSGGQKQR IAIARALVRQ PHILLLDEAT- A-HE -V - -D
-F-R- -T-QSALDTESEKV VQEALDKARE GRTCIVIAHR LSTIQNADLI VVIQNGKVKE HGTHQQLLAQ KGIYFSMVSV QAGAKR*
11SALDTESEKV VQEALDkARE GRTCIVIAHR LSTIQNADLI VVIQNGKVKE HGTHQQLLAQ KGIYFSMV-- QAGAKRL
-E S
1280 Human PGP1571 Hamster PGP1
655 Hamster PGP21276 Mouse PGP2
FIG. 3. Aligned amino acid sequences of hamster, human, and mouse P-glycoproteins. The partial amino acid sequences of the twohamster P-glycoprotein genes were deduced from the cDNA sequences. Nonidentical amino acid residues are indicated by vertical lines.Hamster P-glycoprotein 1 (PGP1) is also compared with human PGPI, and hamster PGP2 is compared with mouse PGP2. The completehamster P-glycoproteirn sequences are given. For the human and mouse sequences, only the residues which differ from their hamsterhomologs are marked. The positions of the six potential transmembrane domains (TM1 to TM6) and the two consensus sequences that formthe putative ATP-binding site (A and B) common to P-glycoprotein, H1yB, and the bacterial periplasmic transport systems (8) are indicatedby horizontal bars above the sequence.
bering) occurs from residues 185 to 257. This sequenceincludes the cytoplasmic domain linking the fourth and fifthpotential transmembrane domains and the fifth potentialtransmembrane domain. Presumably, the human and mouseP-glycoprotein multigene families are also evolving in concert.
Family-member-specific differences are clustered in thepotential transmembrane domains. For example, in thesecond potential transrpembrane domain, there are only twoamino acid differences between the pgpl or pgp2 sequencesacross species, yet there are eight amino acid changesbetween the two hamster genes. In the fourth potentialtransmembrane domain, the human and hamster pgpl se-
quences are identical. Such gene-specific sequences suggestthat there may be functional differences between differentP-glycoprotein molecules.
DISCUSSION
We have presented partial sequences of four P-gly-coprotein transcripts which are simultaneously expressed ina drug-sensitive CHO cell line. The distribution of sequencedifferences between the hamster cDNA clones does notsupport the hypothesis that different P-glycoprotein tran-scripts are generated by either differential splicing or ge-nomic rearrangement but rather suggests that they are
encoded by two distinct members of a P-glycoproteinmultigene family. A P-glycoprotein multigene family hasbeen proposed previously (4, 19). DNA from drug-sensitiveCHO cells, when digested with EcoRI and probed withpCHP1, shows approximately 10 bands on Southern blottingexperiments (19), which suggests that more than two P-glycoprotein genes may exist. Analysis of the differentialamplification of P-glycoprotein gene sequences in multidrug-resistant Chinese hamster lung cell lines (4) and of clonedgenomic sequences from a drug-sensitive CHO cell line(William Ng, personal communication) indicates that there isat least a third gene. Whether or not this third gene or othermembers of the P-glycoprotein gene family are expressed indrug-sensitive CHO cells is not known. Extensive screeningof the cDNA library failed to produce any cDNA clones thatwere not derived from either pgpl or pgp2. Other genefamily members that are not expressed in drug-sensitiveCHO cells may be subject to different physiological or
developmental controls or may be pseudogenes.A comparison of the two hamster P-glycoprotein cDNA
sequences with the single P-glycoprotein cDNA sequencesfrom mouse and human cells shows that there is strongsequence conservation of P-glycoprotein across species andthat the two hamster genes have undergone concerted evo-
lution (Table 2). The extensive nucleotide identity of the two
80701
80395
180801
903195
280901
1003295
3801001
1103395
4801101
1203495
5801201
VOL. 7, 1987

4080 ENDICOTT ET AL.
hamster P-glycoprotein sequences supports this view. T'heimpact of such evolution on the function of P-glycoprotein isnot known; however, the existence of the gene family couldbe a mechanism to help maintain the structural integrity ofan essential protein (6).The discovery that at least two P-glycoprotein genes are
simultaneously expressed in drug-sensitive cells is importantin determining the normal function of P-glycoprotein. It is ofinterest to determine whether different members have dis-tinct functions. Comparison of P-glycoprotein across speciesshows that the human cDNA sequence is more homologousto hamster pgpl and that the mouse sequence is morehomologous to hamster pgp2. This similarity inherentlysuggests that a human gene homologous to hamster pgp2 anda mouse equivalent to hamster pgpl must exist. Since thereare differences in amino acid sequence between the twohamster gene family members, and since these differencesare reflected in the human and mouse homologs, it is likelythat different P-glycoprotein isotypes may have distinctfunctions. It is possible that the normal function of P-glycoprotein in drug-sensitive cells requires the balancedexpression of different multigene family members. It wouldbe of interest to determine whether there is a similar simul-taneous expression of P-glycoprotein genes in drug-sensitivehuman and mouse cells.
Independent transformation experiments with DNA frommultidrug-resistant human or hamster cell lines consistentlyshow transfer of only a subset of the P-glycoprotein gene-specific DNA restriction fragments (5, 23). This findingsuggests that only a single P-glycoprotein gene family mem-ber is transferred and amplified. It is interesting that in bothcases, only pgpl-specific sequences were functionally trans-fected (23; K.L.D., unpublished observation). Perhaps pgp2is inactivated through rearrangement in the donor cell orcannot be amplified sufficiently to meet the stringent selec-tion conditions applied to the recipient cells in the transfor-mation protocol. However, transfection and expression indrug-sensitive cells of a full-length P-glycoprotein cDNAisolated from a drug-sensitive mouse cell line is able toconfer resistance to several drugs (11). This is the mousehomolog of hamster pgp2, and so it appears that either genefamily member, when functionally overexpressed, can causemultidrug resistance.
Multidrug-resistant cell lines often show P-glycoproteingene amplification leading to increased expression of P-glycoprotein. Southern blot analysis of genomic DNA frommultidrug-resistant cell lines shows that P-glycoprotein-specific sequences have undergone differential amplificationand rearrangement which leads to the production of "noveljoints" (19, 25). Aberrant P-glycoprotein mRNAs have beendetected in highly amplified cell lines (J.A.E. and P.F.J.,unpublished observations), which, if translated, would pro-duce variant P-glycoprotein molecules. An analysis of P-glycoprotein-specific cDNAs from the cell line CHRC5 hasshown that both pgpl- and pgp2-specific transcripts can besimultaneously expressed and amplified in multidrug-resistant cell lines. Thus, differential amplification and un-balanced expression of gene family members may contributeto the complexity of the phenotype and should be consideredin the study of multidrug resistance.
ACKNOWLEDGMENTSThis work was supported by the National Cancer Institute of
Canada and by Public Health Service grant CA37130 from theNational Institutes of Health. J.A.E. is the recipient of a MedicalResearch Council of Canada Studentship. and P.F.J. is a Medical
Research Council Fellow. K.L.D. is the recipient of a NationalCancer Institute of Canada Studentship.We thank P. Borst for his gift of cDNA clones from his CHRC5
cDNA library and E. Elliott for constructing the E29Pro' cDNAlibrary. We thank our colleagues for helpful discussions and espe-cially G. Bradley for critical reading of the manuscript.
LITERATURE CITED
1. Birnstiel, M. L., M1. Busslinger, and K. Strub. 1985. Transcrip-tion termination and 3' processing: the end is in site. Cell 41:349-359.
2. Chen, C., J. E. Chin, K. Ueda, D. P. Clark, I. Pastan, M. M.Gottesman, and I. B. Roninson. 1986. Internal duplication andhomology xvith bacterial transport proteins in the mndrl (P-glycoprotein) gene from multidrug-resistant human cells. Cell47:381-389.
3. Dayhoff, M. 0. 1978. Atlas of protein sequence and structure,vol. 5, suppl. 3. National Biomedical Research Foundation,Washington, D.C.
4. de Bruijn, M. H. L., A. NI. Van der Bliek, J. L. Biedler, and P.Borst. 1986. Differential amplification and disproportionateexpression of five genes in three multidrug-resistant Chinesehamster lung cell lines. Mol. Cell. Biol. 6:4717-4722.
5. Deuchars, K. L., R.-P. Du, M. Naik, D. Evernden-Porelle, N.Kartner, A. M. Van der Bliek, and V. Ling. 1987. Expression ofhamster P-glycoprotein and multidrug resistance in DNA-mediated transformants of mouse LTA cells. Mol. Cell. Biol. 7:718-724.
6. Dover, G. A. 1986. Molecular drive in multigene families: howbiological novelties arise, spread and are assimilated. Trends(Genet. 2:159-165.
7. Elliott, E. M., H. Okayama, F. Sarangi, G. Henderson, and V.Ling. 1985. Differential expression of three os-tubulin genes inChinese hamster ovary cells. Mol. Cell. Biol. 5:236-241.
8. Gerlach, J. H., J. A. Endicott, P. F. Juranka, G. Henderson, F.Sarangi, K. L. Deuchars, and V. Ling. 1986. Homology betweenP-glycopr-otein and a bacterial haemolysin transport proteinsuggests a model for multidruig resistance. Nature (London) 324:485-489.
9. Gerlach, J. H., N. Kartner, D. R. Bell, and V. Ling. 1986.Multidrug resistance. Cancer Surv. 5:25-46.
10. Gros, P., J. Croop, and D. Housman. 1986. Mammalianmultidrug resistance gene: complete cDNA sequence indicatesstr-ong homology to bacterial transport proteins. Cell 47:371-380.
11. Gros, P., Y. B. Neriah, J. NI. Croop, and D. E. Housman. 1986.Isolation and expression of a complementary DNA that confersmultidrug resistance. Nature (London) 323:728-731.
12. Hayashida, H., T. Miyata, Y. Yamawaki-Kataoka, T. Honjo, J.Wels, and F. Blattner. 1984. Concerted evolution of the mouseimmunoglobulin gamma chain genes. EMBO J. 3:2047-2053.
13. Higgs, D. R., A. V. S. Hill, D. K. Bowden, D. J. Weatherall, andJ. B. Clegg. 1984. Independent recombination events betweenthe duplicated human ot globin genes; implications for theirconcerted evolution. Nucleic Acids Res. 12:6965-6977.
14. Kartner, N., D. Evernden-Porelle, G. Bradley, and V. Ling.1985. Detection of P-glycoprotein in multidrug-resistant celllines by monoclonal antibodies. Nature (London) 316:820-823.
15. Li, W.-H., and M. Tanimura. 1987. The molecular clock runsmore slowly in man than in apes and monkeys. Nature (London)326:93-96.
16. Mackman, N., J.-M. Nicaud, L. Gray, and I. B. Holland. 1986.Secretion of haemolysin by Escherichia coli. Curr. Top. Micro-biol. Immunol. 125:159-181.
17. Maxam, A. M., and W. Gilbert. 1977. A new method forsequencing DNA. Proc. Natl. Acad. Sci. USA 74:560-564.
18. Perler, F., A. Efstatiadis, P. Lomedico, W. Gilbert, R. Kolodner,and J. Dodgson. 1980. The evolution of genes: the chickenpreproinsulin gene. Cell 20:555-566.
19. Riordan, J. R., K. Deuchars, N. Kartner, N. Alon, J. Trent, andV. Ling. 1985. Amplification of P-glycoprotein genes in mul-tidrug-resistant mammalian cell lines. Nature (London) 316:817-819.
MOL. CELL. BIOL.

TWO P-GLYCOPROTEIN GENES
20. Roninson, I. 1983. Detection and mapping of homologous,repeated and amplified DNA sequences by DNA renaturation inagarose gels. Nucleic Acids Res. 11:5413-5431.
21. Roninson, I. B., H. T. Abelson, D. E. Housman, N. Howell, andA. Varshavsky. 1984. Amplification of specific DNA sequences
correlates with multi-drug resistance in Chinese hamster cells.Nature (London) 309:626-628.
22. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-
ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
23. Shen, D.-W., A. Fojo, I. B. Roninson, J. E. Chin, R. Soffir, I.
Pastan, and M. M. Gottesman. 1986. Multidrug resistance ofDNA-mediated transformants is linked to transfer of the human
mdrl gene. Mol. Cell. Biol. 6:4039-4044.24. Staden, R. 1986. The current status and portability of our
sequence handling software. Nucleic Acids Res. 14:217-231.25. Stark, G. R., and G. M. Wahl. 1984. Gene amplification. Annu.
Rev. Biochem. 53:447-491.26. Ueda, K., M. M. Cornwell, M. M. Gottesman, I. Pastan, I.
Roninson, V. Ling, and J. R. Riordan. 1986. The ,ndrl gene,responsible for multidrug-resistance, codes for P-glycoprotein.Biochem. Biophys. Res. Commun. 141:956-962.
27. Zimmer, E. A., S. L. Martin, S. M. Beverley, Y. W. Kan, andA. C. Wilson. 1980. Rapid duplication and loss of genes codingfor the a chains of hemoglobin. Proc. Natl. Acad. Sci. USA 77:2158-2162.
VOL. 7, 1987 4081





























