Embed Size (px)
Citation preview

International Journal of Food Microbiology xxx (2010) xxx–xxx
FOOD-05054; No of Pages 9
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
ARTICLE IN PRESS
Evaluation of the genetic polymorphism among Lactococcus lactis subsp. cremorisstrains using comparative genomic hybridization and multilocus sequence analysis
Amel Taïbi a, Nassra Dabour a,b, Maryse Lamoureux c, Denis Roy a, Gisèle LaPointe a,⁎a STELA Dairy Research Centre, Institute of Nutraceuticals and Functional Foods, Pavillon des services, Université Laval, Québec, QC, Canada, G1V 0A6b Department of Dairy Science and Technology, Faculty of Agriculture, University of Alexandria, Alexandria, Egyptc Agropur, Inc., 4700 Armand-Frappier Street, St. Hubert, QC, Canada, J3Z 1G5
⁎ Corresponding author. Tel.: +1 418 656 2131x3100E-mail address: [email protected] (G. LaP
0168-1605/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2010.02.020
Please cite this article as: Taïbi, A., et al., Ecomparative genomic hybridization and...,
a b s t r a c t
a r t i c l e i n f oAvailable online xxxx
Keywords:Lactococcus lactis subsp. cremorisStartersCheeseGenetic diversityMicroarray-based comparative genomichybridizationMultilocus sequence analysis
Genetic diversity of Lactococcus lactis subsp. cremoris provides an important reservoir of industrial functions.Knowledge of strain diversity is an important step for the selection of starter cultures, because technologicaland sensorial attributes are strain-dependent and it may help to distinguish strains with particulartechnological properties and performances. In the present study, microarray-based comparative genomichybridization (CGH) and multilocus sequence analysis (MLSA) were used to investigate the genetic variationamong eight strains of Lactococcus lactis subsp. cremoris. The CGH analysis allows strain grouping andidentification of absent or divergent genes involved in metabolism, amino acid biosynthesis, osmoregulationand proteolysis. The MLSA clustering of strains based on the partial sequence of eight genes shows goodcorrelation with the CGH grouping. Strains HP, ATCC 19257 and Wg2 were clustered together, followed byE8, and finally SK11 was in a separate cluster. The combined information provides genetic markers fordistinguishing between strains and their variants. By selecting strains according to their genetic profiles,compatible and complementary mixed starters can be formulated, opening new avenues for industrialapplications.
; fax: +1 418 656 3353.ointe).
l rights reserved.
valuation of the genetic polymorphism amoInternational Journal of Food Microbiology (2
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
In the dairy industry, lactococci are widely used for the productionof a large variety of fermented milks and cheeses as a principlecomponent of the starter culture (Macura and Townsley, 1983;Oberman and Libudzisz, 1998). Their importance resides mainly inlactic acid formation, the production of minor flavor and preservationcomponents and proteolysis (Broom and Limsowtin, 1998; Kieronc-zyk et al., 2003). Lactococcal cultures play a key role for determiningthe quality of fermented dairy products with respect to shelf-life andsensory quality (Smit et al., 2005). To date, two principal subspecies oflactococci are used intensively in dairy starters, Lactococcus lactissubsp. lactis and Lactococcus lactis subsp. cremoris. For CanadianCheddar cheese manufacture, L. lactis subsp. cremoris are preferredover L. lactis subsp. lactis because of their improved contribution toflavor production through degradation of bitter peptides (Salamaet al., 1991; Sandine, 1988; Vedamuthu et al., 1966).
The diversity of lactococcal strains provides an important reservoirfor industrial starters and may contribute to flavor differences andspecific features detected for the same product. Many L. lactis subsp.lactis and cremoris strains are known to carry plasmids encoding
important traits such as lactose catabolism, citrate utilization,proteinase production, bacteriocin production and immunity, bacte-riophage resistance, exopolysaccharide production, as well as heavymetal resistance (Davidson et al., 1996; McKay, 1983). As mobileelements, plasmids can be lost or acquired and contribute to animportant polymorphism among lactococcal strains, even within thesame culture. Moreover, genomic diversity is the origin of differencesin strain performance. Knowledge about strain diversity at thesubspecies level is an important step for starter culture selection,because technological and sensorial potential can be strain-depen-dent characters.
Several molecular techniques, including DNA–DNA hybridization,small subunit (SSU) rRNA gene sequencing, and PCR fingerprintanalysis have been used to differentiate between L. lactis subsp. lactisand L. lactis subsp. cremoris (Erlandson and Batt, 1997; Mangin et al.,1999; Pu et al., 2002; Salama et al., 1991). However, little is knownabout the genomic diversity at the subspecies level of the L. lactissubsp. cremoris strains. Genomic comparison at the intra-subspecieslevel was performed between L. lactis subsp. cremoris strains of theNCDO712 family using pulsed-field gel electrophoresis (PFGE)(Campo et al., 2002). Identical profiles were reported for most ofthe strains tested, suggesting that the differences can be attributed toplasmid content and minor chromosomal mutation or rearrange-ments (Campo et al., 2002). Samarzija et al. (2002) investigated thegenetic diversity of new isolates of L. lactis subsp. cremoris strains from
ng Lactococcus lactis subsp. cremoris strains using010), doi:10.1016/j.ijfoodmicro.2010.02.020

Table 1L. lactis subsp. cremoris strains used in the present study.
Strain Relevant characteristics References Source
SK11 (NCIMB702004)
Prt+ (PIII), phage-resistantAM1 derivative
(Exterkate et al., 1993;Makarova et al., 2006;Obis et al., 2001)
a
ATCC19257
NCDO 607T, type strain, Prt+
(PI)(HP strain deposited in NCDOthen in ATCC)
(Corroler et al., 1998) b
HPS NCDO 607T, type strain, Prt+
(PI)(Corroler et al., 1998) c
HPL Prt− variant of HPS (St-Gelais et al., 1992) c
E8S Prt+ (PII/PIII), wild-typestrain
(St-Gelais et al., 1992) c
E8L Prt− variant of E8S (St-Gelais et al., 1992) c
Wg2S Prt+ (PI/PIII), wild-type strain (St-Gelais et al., 1992) c
Wg2L Prt− derivative of Wg2S (St-Gelais et al., 1992) c
Prt+=proteinase-positive phenotype; Prt−=proteinase-negative phenotype.a NCIMB National Collection of Industrial, Marine and Food Bacteria (Aberdeen,
Scotland, UK).b ATCC: American Type Culture Collection (Manassas, Virginia, USA).c Netherlands Institute for Dairy Research (NIZO) (Ede, The Netherlands).
2 A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
various artisanal cultures using randomly amplified polymorphic DNA(RAPD) (Samarzija et al., 2002). They detected diversity among 40isolates and identified two major DNA patterns. Although PFGE andRAPD techniques have been more appropriate than other typingtechniques, they provide limited knowledge about the completegenome and specific sequence differences.
Recently, the availability of the complete genomic sequences of L.lactis subsp. lactis IL1403 as well as L. lactis subsp. cremoris MG1363and SK11 (Bolotin et al., 2004; Makarova et al., 2006; Wegmann et al.,2007) enabled genome-wide comparison between and within thesubspecies level. L. lactis subsp. lactis strain IL1403 is a plasmid-curedvariant of strain IL594, which was obtained from a cheese starterculture (Chopin et al., 1984). MG1363 is a plasmid-free L. lactis subsp.cremoris strain derived from NCDO 712 by eliminating the large andunstable complement of plasmid DNA using UV treatment andprotoplast-curing strategies (Gasson, 1983). Both IL1403 andMG1363 are widely used in laboratory studies of lactic acid bacteriagenetics. Strain SK11 has been used extensively in cheese-making bythe dairy industry as it has a debittering effect that improves cheeseflavor (Lee et al., 1996). The microarray approach has allowed thisgenetic diversity to be probed more fully, and offers severaladvantages over classical probe based methods: high throughput,parallelism, miniaturization and automation. High-density DNAmicroarrays have been used in a broad variety of applications suchas transcriptomics, comparative genome hybridization (CGH), rese-quencing and microbial community characterization (Huyghe et al.,2009). Microarray-based CGH is a powerful tool for estimatinggenomic diversity and can be used to determine the minimal core ofcommon genes among L. lactis subsp. cremoris strains, which may beuseful to predict and interpret responses to the stress conditions offood processing.
DNA sequence analysis of selected genes is called multilocussequence analysis (MLSA) or typing (MLST), which has been shown tobe powerful technique for diversity analysis of dairy and non dairylactococcal strains (Rademaker et al., 2008). Indeed, MLSA methodshave brought a new dimension to elucidate the partial genomicrelatedness at inter- and intra-specific levels, providing microbiolo-gists with tools to search for phylogenetic markers (Maiden et al.,1998). However, in order to distinguish strains at the subspecies level,the genes selected should contain some polymorphism instead of theconserved regions useful for phylogenetic classification.
The aim of this work was to study the diversity of L. lactis subsp.cremoris by microarray-based CGH and MLSA methods applied to eightcommonly studied strains. This work provides new insights into thesequence divergence among L. lactis subsp. cremoris strains, andintroduces the application of microarray-based CGH for predicting theabsence or presence of genes for strain selection and complementation.
2. Material and methods
2.1. Bacterial strains, growth conditions and DNA manipulation
L. lactis subsp. cremoris strains used in this study are listed inTable 1. All strains were stored in 20% glycerol/Elliker broth at−80 °Cand sub-cultured at least two times at 24 h intervals before use inexperiments. The strains were grown aerobically at 22 °C for 16 h inElliker broth (Difco Laboratories, Sparks, MD, USA) supplementedwith 0.5% yeast extract (Difco) and 0.08% Tween 80 (Sigma-AldrichCanada Ltd, Oakville, ON, Canada). Lactococcal genomic DNA waspurified using standard phenol–chloroform–isoamyl alcohol extrac-tion as described by Hill et al. (Hill et al., 1991).
2.2. Design of an oligonucleotide-based microarray
A DNA microarray was designed based on the published L. lactissubsp. cremoris SK11 genome sequence (Genbank accession no.
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
CP000425). Oligonucleotide probes were designed for 1030 geneswith annotated functions using ROSO software (http://pbil.univ-lyon1.fr/roso/Home.php) (Reymond et al., 2004). The criteria used foroligonucleotide candidate selection were length (60 mer), meltingtemperature (60 to 65 °C, calculated based on a 50 mM saltconcentration), GC percentage (40 to 60%), absence of significantsecondary structures (free energy limit −4.00 kcal/mol), absence ofsmall repetitions of four nucleotides, starting and ending with G or Cand a maximum similarity of 70% with non target regions. A total of2561 oligonucleotide probes (including controls) were selected. Outof these, 2537 probes were specific for gene targets annotated ashaving roles in lactococcal general metabolism such as proteindegradation or in carbohydrate, fatty acid, nucleic acid and aminoacid metabolism (Makarova et al., 2006). The remaining 24 probeswere negative controls including Arabidopsis thaliana genes (rbcL,RCP1 and PPKase), and genes coding for HDAC2 (Mus musculus),ARPC5 (Bos taurus) and GAPD (Bos taurus) as well as lipA and lipBgenes from Escherichia coli. All array probes were printed by Agilent(Agilent Technologies, Palo Alto, CA, USA) with four repetitions usingthe 2×11 K format of two arrays per slide, each array containing10,807 features (4×2561 probes plus 19 empty spots and 544 spotsreserved for Agilent quality controls).
2.3. DNA labeling and microarray hybridization
Total DNA (50 µg) was digested with HincII (New England Biolabs,Pickering, ON, Canada), then randomly labeled by incorporatingaminohexylacrylamido-dUTP (aha-dUTP) during DNA synthesis usingthe Klenow fragment of DNA polymerase I (exo-Klenow) with theBioPrime Plus Array Indirect Genomic Labeling System kit (InvitrogenLife Technologies, Carlsbad, CA, USA) following the manufacturer'sinstructions. Labeled DNA was purified using QIAquick PCR Purifica-tion columns (Qiagen, Mississauga, ON, Canada) followed by couplingwith fluorescent dye Alexa Fluor 555 or 647. The DNA was suspendedin Agilent hybridization buffer and hybridized to microarrays for 17 hat 65 °C according to the Agilent protocol. For each strain, twoindependent hybridizations were performed.
2.4. Microarray data generation and analysis
The arrays with hybridized probes were scanned using aChipreader laser scanner (BioRad, Mississauga, ON, Canada). Thescanned imageswere analyzed first using ArrayPro software and spotsof poor quality determined by visual inspection were removed from
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020

3A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
further analysis. Lowess normalization was carried out on data fromeach channel and net signal intensities were obtained by subtractingthe background. The average of probes and replicates was calculatedusing BRB ArrayTools (http://linus.nci.nih.gov/BRB-ArrayTools.html)(Simon et al., 2007) and the hybridization signals were normalized onthe sample average for all genes with AMADA (Xia and Xie, 2001).Average probe hybridization for each gene was classed as eitherpresent (normalized signal above the average negative control signalof−2, where the average hybridization signal has a normalized valueof 0) or as non-hybridizing genes that are absent or divergent(normalized signal below −2). The normalized data were analyzedusing the TMev package (Saeed et al., 2003).
2.5. PCR amplification and DNA sequencing
PCR was performed with primers designed for eight genes (acmA,ftsE, infB, metE, msmK, nusA, oppA and prtP) of the L. lactis subsp.cremoris SK11 sequence using Primer Express software version 2.0(Applied Biosystems Technologies, Streetsville, ON, Canada) (Table 2).LACR_0276 (acmA) is annotated as a pseudogene in the SK11 GenBankentry CP000425, containing a deletion at 241 bp that changes thereading frame. This gene is designated acmA because it has 98%nucleotide identity with llmg_280 from strain MG1363 and 99% withacmA from strain 2250 (GenBank entry AF036720.1). The eight geneswere amplified using the genomic DNA of the eight tested strains astemplate. The PCR reaction volume of 25 µl contained 50 ng oftemplate DNA, 2.5 mM concentration of each deoxynucleosidetriphosphate, 10 mM concentration of each primer and 2.5 U of TaqDNA polymerase (New England Biolabs). PCR amplification wasperformed with a Tgradient (Biometra, Montreal Biotech, Montréal,QC, Canada) using the following cycles. After 1 cycle at 94 °C for 30 sto denature template DNA, 35 cycles were carried out with thefollowing conditions: denaturation at 94 °C for 30 s, primer annealingat 60 °C for 30 s, and DNA extension at 72 °C for 1 min, finishingwith afinal extension of 5 min at 72 °C then cooled to 4 °C until analysis. ThePCR amplifications were separated by electrophoresis on 0.8% agarose(Invitrogen) gel in 1X TAE buffer containing 40 mM Tris-acetate,1 mM EDTA and, after staining in ethidium bromide, visualized by UVphotography (Gel Doc, BioRad, Mississauga, ON, Canada). Theamplification products were purified by adding 2 µl of the ExoSAP-ITenzyme (USB, Cleveland, OH, USA) to 5 µl of PCR product, the mixturewas incubated 15 min at 37 °C to degrade remaining primers andnucleotides, then 15min at 80 °C for enzyme inactivation. AutomatedDNA sequence analysis was then carried out on both strands of purifiedPCR products with the ABI PRISM BigDye Terminator version 3.1 cyclesequencing kit (Applied Biosystems).
Table 2Primers used for MLSA of L. lactis subsp. cremoris strains.
Locus tag Description Sequence 5′→3′
LACR_0276(pseudogene)
N-acetylmuramidase AcmAa CGCTGTTCTTATTGCCGGAATTGTTTGAA
LACR_1058 (ftsE) Cell-division ATP binding proteinFtsE
CTCGCTCGAAATTGTGGGTCGCCATCAA
LACR_0817 (infB) Translation initiation factor IF-2 GTTGCTGATGACGGTGTGCAAGCGGATG
LACR_1368(metE)
5-methionine synthase GTAACCGAGGACGCTTGGCAAATGCGAA
LACR_0474 (msmK) Multiple sugar ABC transporterATP binding protein
GCCACTCAATATGCCGACAACTTCTGCCT
LACR_0814 (nusA) Transcription termination proteinNusA
TGAGCAAGCTATCAGCCAGACTTGAACC
LACR_D17 (oppA) Oligopeptide ABC transportersubstrate binding protein
CTGGTCCTGGTGGAGGTCTTGCAATGGC
LACR_C42 (prtP) Subtilisin-like serine protease CCGCTTTAATACCCTTGGATGCAGTGGCT
a LACR_0276 is annotated as a pseudogene in SK11; similar sequences (98 to 99%) are an
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
2.6. Multilocus sequence analysis (MLSA)
Sequences of each locus were aligned by using the CLUSTALWalgorithm of the MEGA4 software (Tamura et al., 2007). Statistics foreach locus, such as the number and proportion of polymorphic sites,were calculated using the START2 program (Jolley et al., 2001). ThedN/dS ratios (the ratios of non-synonymous to synonymous substitu-tions) were calculated by using the method of Nei and Gojobori inSTART2 (Nei and Gojobori, 1986).
3. Results
3.1. Hierarchical cluster analysis of CGH profiles of L. lactis subsp.cremoris strains
Based on the normalized microarray hybridization data, thedendrogram shows overall genomic diversity for the eight strains(Fig. 1). Among the three major clusters identified, L. lactis subsp.cremoris SK11 (NCIMB 702004) formed cluster one by itself whereasE8S and E8L strains were grouped together in cluster two. Theremainingmajority of strains (including HPS, Wg2S and their isogenicPrt negative mutants HPL andWg2L) were grouped together with theATCC 19257 strain in cluster three.
3.2. Distribution of variable genes identified by CGH
Themicroarray used in this study covered 997 chromosomal genesand 33 plasmid-coded genes from the genomic sequence of L. lactissubsp. cremoris SK11. These include 98.4% of the genes known orproposed by similarity to be involved in bacterial functions (carbo-hydrate, fatty acid and phospholipids, protein, nucleotide, central,intermediary and energy metabolism, cellular processes, replication,transcription and regulatory functions, translation and biosynthesis ofamino acids, cofactors, transport and cell envelope). The SK11 strainused in this study (NCIMB 702004) was obtained from NCIMB(National Collection of Industrial, Marine and Food Bacteria, UK) andthe plasmid profile does not differ from that published by de Vos (deVos et al., 1984). Hybridization of SK11 DNA revealed divergence in0.2 % of the genes (n=2) compared to the SK11 genome (designatedSK11_R for the reference genomic sequence CP000425 obtained by theJoint Genome Institute group, USA) (Makarova et al., 2006). However,these two genes (hisK and trpD) coding for histidinol phosphatase andanthranilate phosphoribosyltransferase respectively, had at least oneprobe on the array showing significant hybridization to SK11 DNA.
Overall, the tested strains revealed 8.1% low-hybridizing genes,compared to the reference genome SK11_R. The remaining 91.9%(947/1030 genes) were common for all eight strains while 2 to 56
5′ start position PCR product length (bp) Gene ID
CGGAACCACCTG
96 808 4434429
AAGCGAATAGTC
69 522 4432491
TGTTCAACT
1581 907 4433120
GATATTGATGGT
520 530 4432557
CGTTGAACAATGG
46 916 4433016
CAGCAGCATCAGCA
81 834 4433117
GGTCAAGTTTTAC
254 800 4405870
CCGCCCAAC
690 1010 4405830
notated as acmA in other lactococcal strains.
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020
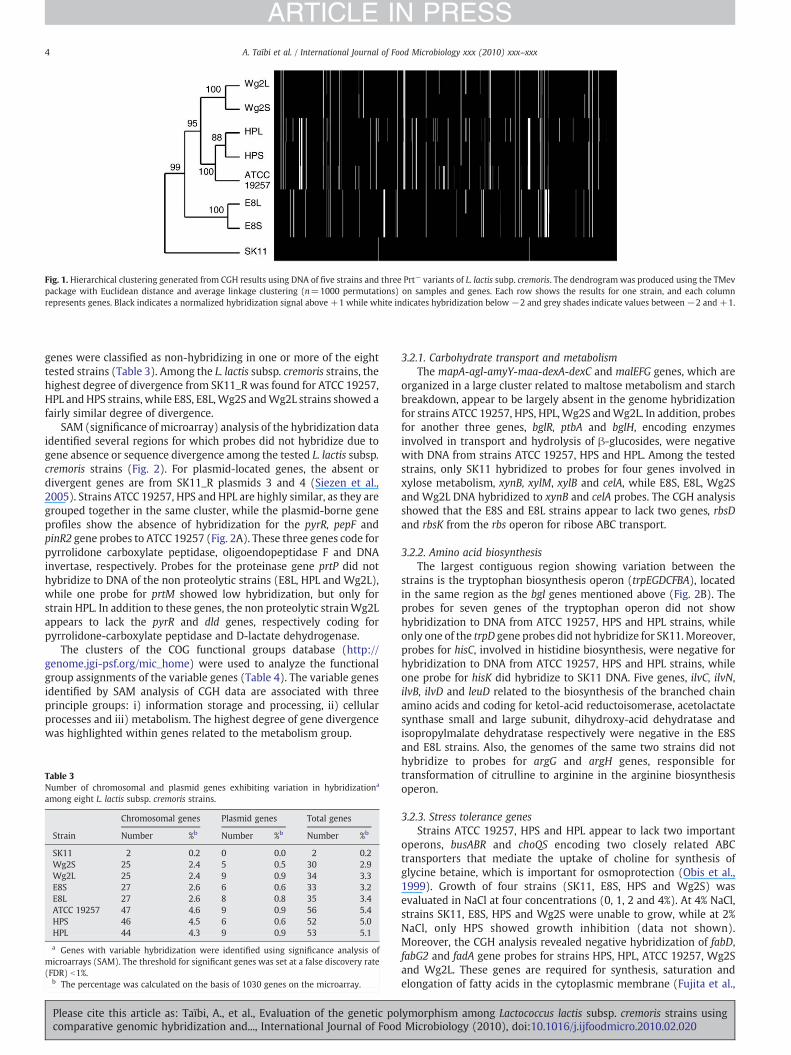
Fig. 1. Hierarchical clustering generated from CGH results using DNA of five strains and three Prt− variants of L. lactis subp. cremoris. The dendrogram was produced using the TMevpackage with Euclidean distance and average linkage clustering (n=1000 permutations) on samples and genes. Each row shows the results for one strain, and each columnrepresents genes. Black indicates a normalized hybridization signal above +1 while white indicates hybridization below −2 and grey shades indicate values between −2 and +1.
4 A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
genes were classified as non-hybridizing in one or more of the eighttested strains (Table 3). Among the L. lactis subsp. cremoris strains, thehighest degree of divergence from SK11_R was found for ATCC 19257,HPL and HPS strains, while E8S, E8L,Wg2S andWg2L strains showed afairly similar degree of divergence.
SAM (significance of microarray) analysis of the hybridization dataidentified several regions for which probes did not hybridize due togene absence or sequence divergence among the tested L. lactis subsp.cremoris strains (Fig. 2). For plasmid-located genes, the absent ordivergent genes are from SK11_R plasmids 3 and 4 (Siezen et al.,2005). Strains ATCC 19257, HPS and HPL are highly similar, as they aregrouped together in the same cluster, while the plasmid-borne geneprofiles show the absence of hybridization for the pyrR, pepF andpinR2 gene probes to ATCC 19257 (Fig. 2A). These three genes code forpyrrolidone carboxylate peptidase, oligoendopeptidase F and DNAinvertase, respectively. Probes for the proteinase gene prtP did nothybridize to DNA of the non proteolytic strains (E8L, HPL and Wg2L),while one probe for prtM showed low hybridization, but only forstrain HPL. In addition to these genes, the non proteolytic strainWg2Lappears to lack the pyrR and dld genes, respectively coding forpyrrolidone-carboxylate peptidase and D-lactate dehydrogenase.
The clusters of the COG functional groups database (http://genome.jgi-psf.org/mic_home) were used to analyze the functionalgroup assignments of the variable genes (Table 4). The variable genesidentified by SAM analysis of CGH data are associated with threeprinciple groups: i) information storage and processing, ii) cellularprocesses and iii) metabolism. The highest degree of gene divergencewas highlighted within genes related to the metabolism group.
Table 3Number of chromosomal and plasmid genes exhibiting variation in hybridizationa
among eight L. lactis subsp. cremoris strains.
Chromosomal genes Plasmid genes Total genes
Strain Number %b Number %b Number %b
SK11 2 0.2 0 0.0 2 0.2Wg2S 25 2.4 5 0.5 30 2.9Wg2L 25 2.4 9 0.9 34 3.3E8S 27 2.6 6 0.6 33 3.2E8L 27 2.6 8 0.8 35 3.4ATCC 19257 47 4.6 9 0.9 56 5.4HPS 46 4.5 6 0.6 52 5.0HPL 44 4.3 9 0.9 53 5.1
a Genes with variable hybridization were identified using significance analysis ofmicroarrays (SAM). The threshold for significant genes was set at a false discovery rate(FDR) b1%.
b The percentage was calculated on the basis of 1030 genes on the microarray.
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
3.2.1. Carbohydrate transport and metabolismThe mapA-agl-amyY-maa-dexA-dexC and malEFG genes, which are
organized in a large cluster related to maltose metabolism and starchbreakdown, appear to be largely absent in the genome hybridizationfor strains ATCC 19257, HPS, HPL,Wg2S andWg2L. In addition, probesfor another three genes, bglR, ptbA and bglH, encoding enzymesinvolved in transport and hydrolysis of β-glucosides, were negativewith DNA from strains ATCC 19257, HPS and HPL. Among the testedstrains, only SK11 hybridized to probes for four genes involved inxylose metabolism, xynB, xylM, xylB and celA, while E8S, E8L, Wg2Sand Wg2L DNA hybridized to xynB and celA probes. The CGH analysisshowed that the E8S and E8L strains appear to lack two genes, rbsDand rbsK from the rbs operon for ribose ABC transport.
3.2.2. Amino acid biosynthesisThe largest contiguous region showing variation between the
strains is the tryptophan biosynthesis operon (trpEGDCFBA), locatedin the same region as the bgl genes mentioned above (Fig. 2B). Theprobes for seven genes of the tryptophan operon did not showhybridization to DNA from ATCC 19257, HPS and HPL strains, whileonly one of the trpD gene probes did not hybridize for SK11.Moreover,probes for hisC, involved in histidine biosynthesis, were negative forhybridization to DNA from ATCC 19257, HPS and HPL strains, whileone probe for hisK did hybridize to SK11 DNA. Five genes, ilvC, ilvN,ilvB, ilvD and leuD related to the biosynthesis of the branched chainamino acids and coding for ketol-acid reductoisomerase, acetolactatesynthase small and large subunit, dihydroxy-acid dehydratase andisopropylmalate dehydratase respectively were negative in the E8Sand E8L strains. Also, the genomes of the same two strains did nothybridize to probes for argG and argH genes, responsible fortransformation of citrulline to arginine in the arginine biosynthesisoperon.
3.2.3. Stress tolerance genesStrains ATCC 19257, HPS and HPL appear to lack two important
operons, busABR and choQS encoding two closely related ABCtransporters that mediate the uptake of choline for synthesis ofglycine betaine, which is important for osmoprotection (Obis et al.,1999). Growth of four strains (SK11, E8S, HPS and Wg2S) wasevaluated in NaCl at four concentrations (0, 1, 2 and 4%). At 4% NaCl,strains SK11, E8S, HPS and Wg2S were unable to grow, while at 2%NaCl, only HPS showed growth inhibition (data not shown).Moreover, the CGH analysis revealed negative hybridization of fabD,fabG2 and fadA gene probes for strains HPS, HPL, ATCC 19257, Wg2Sand Wg2L. These genes are required for synthesis, saturation andelongation of fatty acids in the cytoplasmic membrane (Fujita et al.,
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020

Fig. 2.Normalized hybridization levels of SK11 plasmid (A) and chromosomal (B) gene probes to DNA from eight lactococcal strains. Genes were identified using significance analysisof microarray (SAM). Contiguous genes are grouped according to their organization in the reference genome of strain SK11_R. The status of each gene is indicated as follows: black,present (normalized signal above +1); white, absent (signal below −2); grey, low hybridization (signal between −2 and +1).
5A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
2007; Wang and Cronan, 2004). Probes for two genes, mtsB andmtsC,did not hybridize to DNA from ATCC 19257, HPS and HPL. ThemtsABCoperon encodes ABC transporters of manganese and iron, which areimportant for resistance to oxidative stress in Gram-positive bacteria(Janulczyk et al., 2003).
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
3.2.4. Cell wall biosynthesisProbes for the rgpABEF and ycbF genes, all part of the operon
involved in cell wall localization and side chain formation ofrhamnose-glucose polysaccharides (RGP), showed low to no hybrid-ization with DNA from strains ATCC 19257, HPS, HPL, Wg2S and
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020
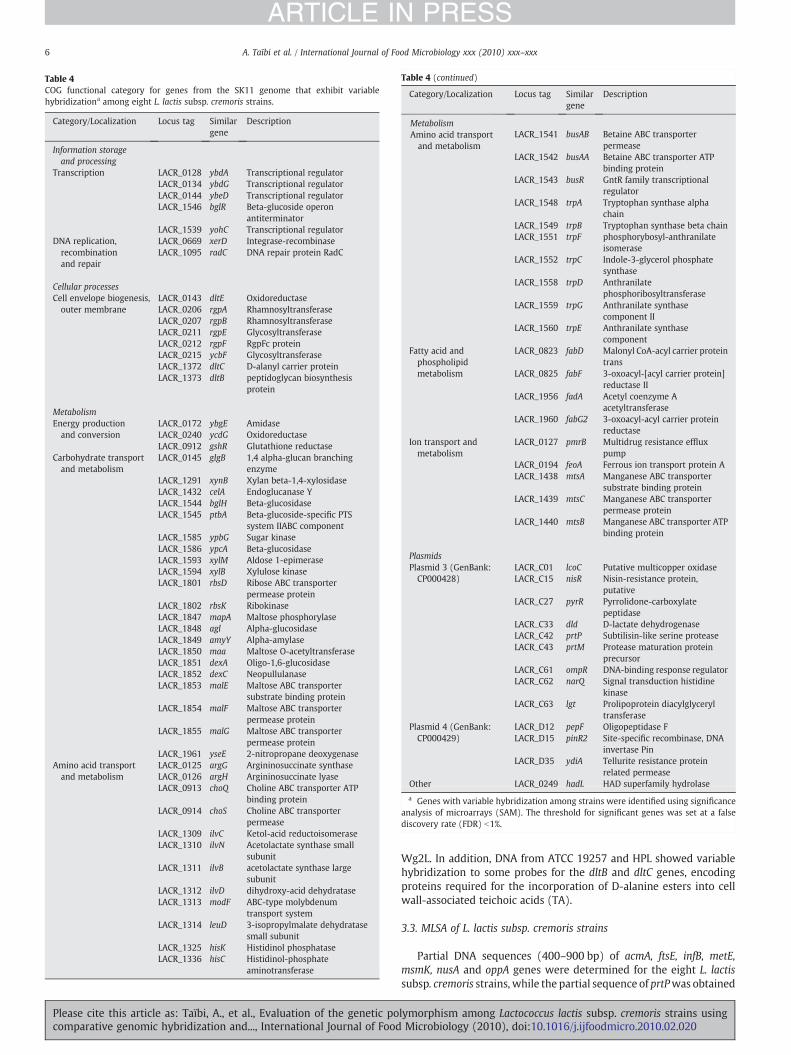
Table 4COG functional category for genes from the SK11 genome that exhibit variablehybridizationa among eight L. lactis subsp. cremoris strains.
Category/Localization Locus tag Similargene
Description
Information storageand processing
Transcription LACR_0128 ybdA Transcriptional regulatorLACR_0134 ybdG Transcriptional regulatorLACR_0144 ybeD Transcriptional regulatorLACR_1546 bglR Beta-glucoside operon
antiterminatorLACR_1539 yohC Transcriptional regulator
DNA replication,recombinationand repair
LACR_0669 xerD Integrase-recombinaseLACR_1095 radC DNA repair protein RadC
Cellular processesCell envelope biogenesis,outer membrane
LACR_0143 dltE OxidoreductaseLACR_0206 rgpA RhamnosyltransferaseLACR_0207 rgpB RhamnosyltransferaseLACR_0211 rgpE GlycosyltransferaseLACR_0212 rgpF RgpFc proteinLACR_0215 ycbF GlycosyltransferaseLACR_1372 dltC D-alanyl carrier proteinLACR_1373 dltB peptidoglycan biosynthesis
protein
MetabolismEnergy productionand conversion
LACR_0172 ybgE AmidaseLACR_0240 ycdG OxidoreductaseLACR_0912 gshR Glutathione reductase
Carbohydrate transportand metabolism
LACR_0145 glgB 1,4 alpha-glucan branchingenzyme
LACR_1291 xynB Xylan beta-1,4-xylosidaseLACR_1432 celA Endoglucanase YLACR_1544 bglH Beta-glucosidaseLACR_1545 ptbA Beta-glucoside-specific PTS
system IIABC componentLACR_1585 ypbG Sugar kinaseLACR_1586 ypcA Beta-glucosidaseLACR_1593 xylM Aldose 1-epimeraseLACR_1594 xylB Xylulose kinaseLACR_1801 rbsD Ribose ABC transporter
permease proteinLACR_1802 rbsK RibokinaseLACR_1847 mapA Maltose phosphorylaseLACR_1848 agl Alpha-glucosidaseLACR_1849 amyY Alpha-amylaseLACR_1850 maa Maltose O-acetyltransferaseLACR_1851 dexA Oligo-1,6-glucosidaseLACR_1852 dexC NeopullulanaseLACR_1853 malE Maltose ABC transporter
substrate binding proteinLACR_1854 malF Maltose ABC transporter
permease proteinLACR_1855 malG Maltose ABC transporter
permease proteinLACR_1961 yseE 2-nitropropane deoxygenase
Amino acid transportand metabolism
LACR_0125 argG Argininosuccinate synthaseLACR_0126 argH Argininosuccinate lyaseLACR_0913 choQ Choline ABC transporter ATP
binding proteinLACR_0914 choS Choline ABC transporter
permeaseLACR_1309 ilvC Ketol-acid reductoisomeraseLACR_1310 ilvN Acetolactate synthase small
subunitLACR_1311 ilvB acetolactate synthase large
subunitLACR_1312 ilvD dihydroxy-acid dehydrataseLACR_1313 modF ABC-type molybdenum
transport systemLACR_1314 leuD 3-isopropylmalate dehydratase
small subunitLACR_1325 hisK Histidinol phosphataseLACR_1336 hisC Histidinol-phosphate
aminotransferase
Table 4 (continued)
Category/Localization Locus tag Similargene
Description
LACR_1541 busAB Betaine ABC transporterpermease
LACR_1542 busAA Betaine ABC transporter ATPbinding protein
LACR_1543 busR GntR family transcriptionalregulator
LACR_1548 trpA Tryptophan synthase alphachain
LACR_1549 trpB Tryptophan synthase beta chainLACR_1551 trpF phosphorybosyl-anthranilate
isomeraseLACR_1552 trpC Indole-3-glycerol phosphate
synthaseLACR_1558 trpD Anthranilate
phosphoribosyltransferaseLACR_1559 trpG Anthranilate synthase
component IILACR_1560 trpE Anthranilate synthase
componentFatty acid andphospholipidmetabolism
LACR_0823 fabD Malonyl CoA-acyl carrier proteintrans
LACR_0825 fabF 3-oxoacyl-[acyl carrier protein]reductase II
LACR_1956 fadA Acetyl coenzyme Aacetyltransferase
LACR_1960 fabG2 3-oxoacyl-acyl carrier proteinreductase
Ion transport andmetabolism
LACR_0127 pmrB Multidrug resistance effluxpump
LACR_0194 feoA Ferrous ion transport protein ALACR_1438 mtsA Manganese ABC transporter
substrate binding proteinLACR_1439 mtsC Manganese ABC transporter
permease proteinLACR_1440 mtsB Manganese ABC transporter ATP
binding protein
PlasmidsPlasmid 3 (GenBank:CP000428)
LACR_C01 lcoC Putative multicopper oxidaseLACR_C15 nisR Nisin-resistance protein,
putativeLACR_C27 pyrR Pyrrolidone-carboxylate
peptidaseLACR_C33 dld D-lactate dehydrogenaseLACR_C42 prtP Subtilisin-like serine proteaseLACR_C43 prtM Protease maturation protein
precursorLACR_C61 ompR DNA-binding response regulatorLACR_C62 narQ Signal transduction histidine
kinaseLACR_C63 lgt Prolipoprotein diacylglyceryl
transferasePlasmid 4 (GenBank:CP000429)
LACR_D12 pepF Oligopeptidase FLACR_D15 pinR2 Site-specific recombinase, DNA
invertase PinLACR_D35 ydiA Tellurite resistance protein
related permeaseOther LACR_0249 hadL HAD superfamily hydrolase
a Genes with variable hybridization among strains were identified using significanceanalysis of microarrays (SAM). The threshold for significant genes was set at a falsediscovery rate (FDR) b1%.
MetabolismAmino acid transportand metabolism
6 A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
Wg2L. In addition, DNA from ATCC 19257 and HPL showed variablehybridization to some probes for the dltB and dltC genes, encodingproteins required for the incorporation of D-alanine esters into cellwall-associated teichoic acids (TA).
3.3. MLSA of L. lactis subsp. cremoris strains
Partial DNA sequences (400–900 bp) of acmA, ftsE, infB, metE,msmK, nusA and oppA genes were determined for the eight L. lactissubsp. cremoris strains, while the partial sequence of prtPwas obtained
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020

Fig. 3. Dendrogram showing the sequence similarity of L. lactis strains. A) Clusteranalysis of seven partial gene sequences of L. lactis subsp. cremoris strains with L. lactissubsp. lactis IL1403 as the outgroup. Bar, 0.01 nucleotide substitutions per site. B)Cluster analysis of eight concatenated gene sequences (adding prtP) from five L. lactissubsp. cremoris dairy strains compared with the reference gene sequences of SK11_R.The trees were constructed using the neighbor-joining algorithm based on the Jukes–Cantor model of nucleotide substitution (MEGA 4). Bootstrap percentages (≥50) after500 simulations are shown. Bar, 0.0005 nucleotide substitutions per site.
7A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
for five strains (excluding Prt− strains HPL, E8L and Wg2L). Thesequences showed a high percent of identity (98 to 100%), withpolymorphic nucleotides in all eight genes (Table 5). The percentage ofpolymorphic sites varied between 0.2 (metE) to 3.0% (prtP). For eachgene, each sequence variation was designated as an allele and eachunique combination of allele numbers represents one sequence type(ST). The number of defined alleles varied from 2 to 6 (Table 5 andSupplementary material, Fig. S1). Our results show that all the testedPrt− strains had the same sequence type (ST) as the Prt+ parent strain.
The proportion of nucleotide alterations that changed the aminoacid sequence (non-synonymous substitutions, dN) and the propor-tion of silent changes (synonymous substitutions, dS) were calculatedfor each gene to give an acceptance rate (dN/dS ratio). This ratiomeasures the selective pressure at the protein level, dN/dS=1meaning neutral mutations, dN/dSb1 purifying selection, and dN/dSN1 diversifying selection. The dN/dS ratio varied from 0 formetE to2.0 for acmA (Table 5). The metE locus had only one nucleotidesubstitution that was synonymous and did not affect the amino acidcomposition (Table 5). The lower dN/dS ratio of infB (0.48) could alsoindicate that the degree of selection is stronger than for the remaininggenes. Moreover, the acmA and oppA genes presented dN/dS valueshigher than 1, which indicates the insensitivity of these two genes tonucleotide substitutions leading to the variation of certain aminoacids.
The concatenated gene sequences of seven loci (without prtP)revealed the close similarity of L. lactis subsp. cremoris strains inrelation to L. lactis subsp. lactis IL1403 as an outlier (Fig. 3A). Withinthe L. lactis subsp. cremoris group, strain MG1363 showed distinctsequence divergence from the dairy strains. The prtP gene was addedto the concatenated gene sequences giving eight loci for comparingdairy strains of L. lactis subsp. cremoris (Fig. 3B). This addition did notchange the branching points compared to the dendrogram con-structed using seven loci (Fig. 3A). The eight-gene dendrogramreveals that strain SK11 (NCIMB 702004) forms a single cluster withthe SK11_R reference genome sequence (Fig. 3B). The Wg2S strainappears closer to the ATCC 19257 and HPS strains, while the E8S strainappears intermediate.
4. Discussion
For a better control of starter composition used in cheese-making,improved knowledge of the variability among strains is necessary.Strain SK11 is recognized for contributing to nonbitter flavor (Lee
Table 5Sequence variation of selected genes from L. lactis subsp. cremoris.
Locus tag Genelength(bp)
Amplifiedfragmentlength (bp)
Analyzedfragmentlength (bp)
No. ofpolymorphicsites (%)
No. ofalleles
dN/dSa
LACR_0276(acmA)
1313 808 748 15 (2.0) 5 2.08
LACR_1058(ftsE)
693 522 464 4 (0.8) 3 0.81
LACR_0817(infB)
2853 907 812 5 (0.6) 3 0.48
LACR_1368(metE)
2274 530 504 1 (0.2) 2 –
LACR_0474(msmK)
1137 916 725 8 (1.1) 4 0.84
LACR_0814(nusA)
1149 834 747 17 (2.3) 6 1.00
LACR_D17(oppA)
1803 800 645 8 (1.2) 5 1.80
LACR_C42(prtP)
5889 1010 561 17 (3.0) 5 1.07
a The dN/dS ratio is calculated by dividing non-synonymous substitutions by thenumber of synonymous substitutions.
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
et al., 1996), while HP is non-autolytic and is responsible for thedevelopment of bitterness in Cheddar cheese (Pillidge et al., 2002). St-Gelais et al. demonstrated that strain Wg2 exhibited a differentproteolytic activity and the least lactic acid production compared tostrains HP and E8, but E8 produced more lactic acid than other strains(St-Gelais et al., 1992; St-Gelais et al., 1993). The four strains differ onthe basis of the protease PrtP sequence and specificity of casein αS1cleavage sites (Exterkate et al., 1993). Other studies also showed thatthese four strains do not have the same autolytic and proteolyticcapacity and consequently, vary in technological potential (Pillidgeet al., 2002; Sheehan et al., 2005; St-Gelais et al., 1992). Thus, thesestrains can be regarded as examples of strains originating fromstarters that have been used in the cheese industry.
In this study, we used CGH and MLSA as two different methods foranalysis of the gene diversity of L. lactis subsp. cremoris strains andtheir variants. Using CGH, hybridization of the DNA from eight strainsrevealed the presence of sequences similar to more than 91% of genesfrom the reference genome of SK11. As the hybridization temperaturewas 65 °C, and themelting temperature of the probes was between 60and 65 °C, a very low tolerance for mismatches was allowed.Therefore, any divergence in sequence would be enough to lead to aweak hybridization signal for some probes. Very few microarrayprobes did show weak hybridization signals for the DNA from thestrain of SK11 originating from the NCIMB collection. The variationsobserved between the genome of the strain SK11 used in this studyand the genome of the reference strain (SK11_R) available in NCBIcould be due to genetic drift of the strain in two locations.
Among genes identified by CGH analysis as absent or divergent,some are responsible for the metabolism of sugars. These genes codefor enzymes of sugar degradation such as xylose and maltose.Previous studies showed that strains having these genes are of plantorigin (Erlandson et al., 2000; Siezen et al., 2008), but the presence ofthese genes does not necessarily imply their capacity to develop in thepresence of these sugars. Moreover, even though the xylose geneswere detected, strain SK11 was not able to develop in the presence of
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020

8 A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
xylose, probably due to deletions in the xyl genes (Siezen et al., 2008).The divergence of these metabolic genes should not have an influenceon the starter response during the conditions of cheese manufacture.
The strains of dairy origin have acquired systems which enabledthem to adapt to growth in milk, such as their capacity to use lactose(de Vos and Gasson, 1989) and to degrade casein using the PrtPprotease (Kok, 1990). On the other hand, strains of dairy origin havelargely lost their capacity to synthesize the branched chain aminoacids (BCAA) and histidine (Deguchi and Morishita, 1992; Godon etal., 1993). Indeed, in this study, E8S and E8L seem to lack genes ofBCAA biosynthesis, while HPS, HPL and ATCC 19257 show divergencein genes of histidine and tryptophan synthesis. The aromatic aminoacid tryptophan can lead to the production of off-flavor compoundsduring cheese-making (van Kranenburg et al., 2002). Dairy L. lactissubsp. cremoris strains have been shown to be auxotrophic for sixamino acids (Glu, Met, Leu, Ile, Val, and His) while strains of plantorigin are prototrophic for all the amino acids (Chopin, 1993; Cocaign-Bousquet et al., 1995). In Streptococcus thermophilus, the branchedchain amino acid (BCAA) pathway is considered as essential foroptimal growth in milk, as a strain with mutated BCAA genes is notable to develop (Garault et al., 2000). However, in the case of strain E8(BCAA negative), growth in milk does not seem to be affectedcompared to HP or Wg2 (St-Gelais et al., 1992; St-Gelais et al., 1993),probably due to the presence of specific transporters for free aminoacids and peptides. In strains E8S and E8L, the genes for argininebiosynthesis also appear to have been lost, and arginine is anonessential amino acid for lactococcal strains growing in milk.
Strains HPS, HPL and ATCC 19257 also lack genes of osmoprotec-tion, busABR and choQS, as well as some fab genes for fatty acidbiosynthesis. The fab genes are responsible in particular for theelongation and the saturation of phospholipids (Heath and Rock,1995). The absence of the fab genes might lead to a lower amount ofsaturated fatty acids and less elongated fatty acids in the cellmembrane, making it less densely packed. A modification of thefatty acid composition could lead to membranes that may differ inresponse to stress conditions. The genes responsible for osmoregu-lation are also considered as thermoprotectors (Caldas et al., 1999);their absence would reduce tolerance to salting and cooking.According to Guillot et al., an osmotic shock can also be responsiblefor themodification of the composition in fatty acids of themembrane(Guillot et al., 2000). Our findings suggest the low resistance of HPS toosmotic stress conditions as growth was inhibited at 2% NaCl. Indeed,HP failed to accumulate betaine in a previous study (Obis et al., 2001).In the case of HPS and ATCC 19257, the absence of hybridization ofthese genes may indicate variant or nonfunctional gene products thatcould lead to cellular lysis in response to a combination of osmotic andthermal stress. Studies are underway to evaluate differences in geneexpression profiles of these strains under the conditions of Cheddarcheese cooking temperature and salting.
Certain genes from the rgp operon are also lacking or low-hybridizing in ATCC 19257, HPS, HPL, Wg2S and Wg2L. These genesinvolved in biosynthesis and transport of cell wall polysaccharides arealso important for adsorption of certain phages to the cell surface.Dupont et al. (2004) demonstrated that rpg mutants were phage-resistant. This phenomenon seems to be gene-dependent, underliningdifferences in phage sensitivity. This was established for strainsIL1403 and Wg2, where the genes responsible for phage adsorptionwere shown to be different (Dupont et al., 2004).
The MLSA provides classification of strains on the basis of partialgene sequences, which show some polymorphic sites between strainsSK11 and SK11_R. This observation can be explained by genetic driftbetween the variants that corroborates the microarray hybridizationresults. Dairy strains of L. lactis subsp. cremoris are tightly grouped incomparison to L. lactis subsp. lactis IL1403 sequences, and divergefrom L. lactis subsp. cremoris MG1363. Within the dairy strains, agradient of divergence is apparent from SK11 towards HPS, with
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
strainWg2S closer to HPS. There are four genes with a dN/dS ratio of 1or higher (prtP, oppA, acmA and nusA) which means that they are notvery sensitive to nucleotide mutations leading to the variation ofcertain amino acids. These genes could be used to discriminatebetween the L. lactis subsp. cremoris strains through SNP (singlenucleotide polymorphism) detection.
The strains used in this study constitute only a small sample of thevarious profiles of strains used in the cheese industry. These strainshave been the subject of several studies of microbiological andbiochemical characterization, and the analysis of their genetic profileshas made it possible to validate the use of DNAmicroarrays and MLSAfor typing L. lactis subsp. cremoris strains. The identification ofpolymorphic sites by MLSA allows an initial classification andselection of strains whose DNA can then be submitted to completegenomic profiling by microarray hybridization. The characterizationof the genomic diversity of L. lactis subsp. cremoris strains used incheese-making is advanced through the use of this unique DNAmicroarray, which allows the detection of the presence or absence ofgenes associated with metabolic activities of industrial interest. Thecombination of the information obtained by CGH and MLSA on strainclassification and the identification of the source of diversity opensnew avenues for industrial applications based on selection of strainsaccording to their genetic profiles. The identification of non-hybridizing genes will allow the elimination of strains which lackgenes responsible for paramount functions, the prediction of theirresponse or behavior during cheese manufacture and the selection ofthe strains which could be compatible in mixed starter cultures. Thesemolecular tools will help to classify strains in function of theirperformance, in order to avoid defects such as bitterness and todetermine strain compatibility, for example, in terms of proteolyticactivity. This will allow the selection of the best strain combinationsby the presence and type of specific characters such as proteolysis,peptidases and acidification. However, genomic analyses alone do notprovide a full understanding of starter diversity, as the second step forbetter comprehension of genetic variation is the investigation of geneexpression of L. lactis subsp. cremoris strains under Cheddar cheeseconditions. Transcriptomics will improve this knowledge in order todevelop tools for screening strains exhibiting the desired properties.Biomarkers could then be identified that could be used to predict theperformance of Lactococcus lactis subsp. cremoris during Cheddarcheese manufacture.
Acknowledgements
This work was supported by a NSERC Strategic Project grantawarded to G. LaPointe and D. Roy in collaboration with Agropur, Inc.The authors gratefully acknowledge the financial contribution ofindustrial partnerships (Agropur Inc., Fromagerie Clément/DamafroInc., Novalait Inc, Parmalat Canada, Dairy Farmers of Canada, GroupeSaputo inc.) and NSERC (NSERC-Dairy Sector Industrial Research Chairin Cheese Technology and Typicity). We thank Marie-Hélène Lessardfor providing prtP sequence data.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ijfoodmicro.2010.02.020.
References
Bolotin, A., Quinquis, B., Sorokin, A., Ehrlich, D., 2004. Recent genetic transfer betweenLactococcus lactis and enterobacteria. Journal of Bacteriology 186, 6671–6677.
Broom, M.C., Limsowtin, G.K.Y., 1998. Starter peptidase activity in maturing cheese.Australian Journal of Dairy Technology 53, 79–82.
Caldas, T., Demont-Caulet, N., Ghazi, A., Richarme, G., 1999. Thermoprotection byglycine betaine and choline. Microbiology 145, 2543–2548.
Campo, N., Dias, M., Daveran-Mingot, M.L., Ritzenthaler, P., Le Bourgeois, P., 2002.Genome plasticity in Lactococcus lactis. Antonie van Leeuwenhoek 82, 123–132.
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020

9A. Taïbi et al. / International Journal of Food Microbiology xxx (2010) xxx–xxx
ARTICLE IN PRESS
Chopin, A., 1993. Organization and regulation of genes for amino acid biosynthesis inlactic acid bacteria. FEMS Microbiology Reviews 12, 21–38.
Chopin, A., Chopin, M.C., Moillo-Batt, A., Langella, P., 1984. Two plasmid-determinedrestriction and modification systems in Streptococcus lactis. Plasmid 11, 260–263.
Cocaign-Bousquet, M., Garrigues, C., Novak, L., Lindley, N.D., Loubiere, P., 1995. Rationaldevelopment of a simple synthetic medium for the sustained growth of Lactococcuslactis. Journal of Applied Bacteriology 79, 108–116.
Corroler, D., Mangin, I., Desmasures, N., Gueguen, M., 1998. An ecological study oflactococci isolated from raw milk in the Camembert cheese registered designationof origin area. Applied and Environmental Microbiology 64, 4729–4735.
Davidson, B.E., Kordias, N., Dobos, M., Hillier, A.J., 1996. Genomic organization of lacticacid bacteria. Antonie van Leeuwenhoek 70, 161–183.
de Vos, W.M., Gasson, M.J., 1989. Structure and expression of the Lactococcus lactis genefor phospho-β-galactosidase (lacG) in Escherichia coli and L. lactis. Journal ofGeneral Microbiology 135, 1833–1846.
de Vos, W.M., Underwood, H.M., Lyndon Davies, F., 1984. Plasmid encodedbacteriophage resistance in Streptococcus cremoris SK11. FEMS MicrobiologyLetters 23, 175–178.
Deguchi, Y., Morishita, T., 1992. Nutritional requirements in multiple auxotrophic lacticacid bacteria: genetic lesions affecting amino acid biosynthetic pathways inLactococcus lactis, Enterococcus faecalis and Pediococcus acidilactici. Bioscience,Biotechnology, and Biochemistry 56, 913–918.
Dupont, K., Janzen, T., Vogensen, F.K., Josephsen, J., Stuer-Lauridsen, B., 2004.Identification of Lactococcus lactis genes required for bacteriophage adsorption.Applied and Environmental Microbiology 70, 5825–5832.
Erlandson, K., Batt, C.A., 1997. Strain-specific differentiation of lactococci in mixedstarter culture populations using randomly amplified polymorphic DNA-derivedprobes. Applied and Environmental Microbiology 63, 2702–2707.
Erlandson, K.A., Park, O., El Khal, W., Kao, H., Basaran, P., Brydges, S., Batt, C.A., 2000.Dissolution of xylose metabolism in Lactococcus lactis. Applied and EnvironmentalMicrobiology 66, 3974–3980.
Exterkate, F.A., Alting, A.C., Bruinenberg, P.G., 1993. Diversity of cell envelopeproteinase specificity among strains of Lactococcus lactis and its relationship tocharge characteristics of the substrate-binding region. Applied and EnvironmentalMicrobiology 59, 3640–3647.
Fujita, Y., Matsuoka, H., Hirooka, K., 2007. Regulation of fatty acid metabolism inbacteria. Molecular Microbiology 66, 829–839.
Garault, P., Lefort, C., Juillard, V., Monnet, V., 2000. Branched-chain amino acidbiosynthesis is essential for optimal growth of Streptococcus thermophilus in milk.Applied and Environmental Microbiology 66, 5128–5133.
Gasson, M.J., 1983. Plasmid complements of Streptococcus lactis NCDO 712 and otherlactic streptococci after protoplast-induced curing. Journal of Bacteriology 154, 1–9.
Godon, J.J., Delorme, C., Bradowski, J., Chopin, M.C., Ehrlich, D., Renault, P., 1993. Geneinactivation in Lactococcus lactis: branched-chain amino acid biosynthesis. Journalof Bacteriology 175, 4383–4390.
Guillot, A., Obis, D., Mistou, M.Y., 2000. Fatty acid membrane composition andactivation of glycine-betaine transport in Lactococcus lactis subjected to osmoticstress. International Journal of Food Microbiology 55, 47–51.
Heath, R.J., Rock, C.O., 1995. Enoyl-acyl carrier protein reductase (fabI) plays adeterminant role in completing cycles of fatty acid elongation in Escherichia coli.Journal of Biological Chemistry 270, 26538–26542.
Hill, C., Massey, J., Klaenhammer, T.R., 1991. Rapid method to characterize lactococcalbacteriophage genomes. Applied and Environmental Microbiology 57, 283–288.
Huyghe, A., Francois, P., Schrenzel, J., 2009. Characterization of microbial pathogens byDNA microarrays. Infection, Genetics and Evolution 9, 987–995.
Janulczyk, R., Ricci, S., Bjorck, L., 2003. MtsABC is important for manganese and irontransport, oxidative stress resistance, and virulence of Streptococcus pyogenes.Infection and Immunity 71, 2656–2664.
Jolley, K.A., Feil, E.J., Chan, M.S., Maiden, M.C., 2001. Sequence type analysis andrecombinational tests (START). Bioinformatics 17, 1230–1231.
Kieronczyk, A., Skeie, S., Langsrud, T., Yvon, M., 2003. Cooperation between Lactococcuslactis and nonstarter lactobacilli in the formation of cheese aroma from aminoacids. Applied and Environmental Microbiology 692, 734–739.
Kok, J., 1990. Genetics of the proteolytic system of lactic acid bacteria. FEMSMicrobiology Reviews 87, 15–42.
Lee, K.D., Lo, C.G., Warthesen, J.J., 1996. Removal of bitterness from the bitter peptidesextracted from Cheddar cheese with peptidases from Lactococcus lactis ssp.cremoris SK11. Journal of Dairy Science 79, 1521–1528.
Macura, D., Townsley, P.M., 1983. Scandinavian ropy milk — identification andcharacterization of endogenous ropy lactic streptococci and their extracellularexcretion. Journal of Dairy Science 67, 735–744.
Maiden, M.C., Bygraves, J.A., Feil, E., Morelli, G., Russell, J.E., Urwin, R., Zhang, Q., Zhou, J.,Zurth, K., Caugant, D.A., Feavers, I.M., Achtman, M., Spratt, B.G., 1998. Multilocussequence typing: a portable approach to the identification of clones withinpopulations of pathogenic microorganisms. Proceedings of the National Academyof Sciences U.S.A. 95, 3140–3145.
Makarova, K., Slesarev, A., Wolf, Y., Sorokin, A., Mirkin, B., Koonin, E., Pavlov, A., Pavlova,N., Karamychev, V., Polouchine, N., Shakhova, V., Grigoriev, I., Lou, Y., Rohksar, D.,Lucas, S., Huang, K., Goodstein, D.M., Hawkins, T., Plengvidhya, V., Welker, D.,Hughes, J., Goh, Y., Benson, A., Baldwin, K., Lee, J.H., Díaz-Muñiz, I., Dosti, B.,Smeianov, V., Wechter, W., Barabote, R., Lorca, G., Altermann, E., Barrangou, R.,Ganesan, B., Xie, Y., Rawsthorne, H., Tamir, D., Parker, C., Breidt, F., Broadbent, J.,Hutkins, R., O'Sullivan, D., Steele, J., Unlu, G., Saier, M., Klaenhammer, T., Richardson,P., Kozyavkin, S., Weimer, B., Mills, D., 2006. Comparative genomics of the lacticacid bacteria. Proceedings of the National Academy of Sciences U.S.A. 42,15611–15616.
Please cite this article as: Taïbi, A., et al., Evaluation of the genetic pocomparative genomic hybridization and..., International Journal of Food
Mangin, I., Corroler, D., Reinhardt, A., Gueguen, M., 1999. Genetic diversity among dairylactococcal strains investigated by polymerase chain reaction with three arbitraryprimers. Journal of Applied Microbiology 86, 514–520.
McKay, L.L., 1983. Functional properties of plasmids in lactic streptococci. Antonie vanLeeuwenhoek 49, 259–274.
Nei, M., Gojobori, T., 1986. Simple methods for estimating the numbers of synonymousand nonsynonymous nucleotide substitutions. Molecular Biology and Evolution 3,418–426.
Oberman, H., Libudzisz, Z., 1998. Fermented milks, In: Wood, B.J.B. (Ed.), 2nd ed.Microbiology of Fermented Foods, vol. 1. Blackie Academic & Professional, London,pp. 308–350.
Obis, D., Guillot, A., Gripon, J.C., Renault, P., Bolotin, A., Mistou, M.Y., 1999. Genetic andbiochemical characterization of a high-affinity betaine uptake system (BusA) inLactococcus lactis reveals a new functional organization within bacterial ABCtransporters. Journal of Bacteriology 181, 6238–6246.
Obis, D., Guillot, A., Mistou, M.Y., 2001. Tolerance to high osmolality of Lactococcus lactissubsp. lactis and cremoris is related to the activity of a betaine transport system.FEMS Microbiology Letters 202, 39–44.
Pillidge, C.J., Rallabhandi, P.S.V.S., Tong, X.Z., Gopal, P.K., Farley, P.C., Sullivan, P.A., 2002.Autolysis of Lactococcus lactis. International Dairy Journal 12, 133–140.
Pu, Z.Y., Dobos, M., Limsowtin, G.K., Powell, I.B., 2002. Integrated polymerase chainreaction-based procedures for the detection and identification of species andsubspecies of the gram-positive bacterial genus Lactococcus. Journal of AppliedMicrobiology 93, 353–361.
Rademaker, J.L.W., Herbet, H., Starrenburg, M.J.C., Naser, S.M., Gevers, D., Kelly, W.J.,Hugenholtz, J., Swings, J., van Hylckama Vlieg, J.E.T., 2008. Diversity analysis of dairyand nondairy Lactococcus lactis isolates, using a novel multilocus sequence analysisscheme and (GTG)5-PCR fingerprinting. Applied and Environmental Microbiology73, 7128–7137.
Reymond, N., Charles, H., Duret, I., Calevro, F., Beslon, G., Fayard, J.M., 2004. ROSO:optimizing oligonucleotide probes for microarrays. Bioinformatics 20, 271–273.
Saeed, A.I., Sharov, V., White, J., Li, W.L., Bhagabati, N., Braisted, J., Klapa, M., Currier, T.,Thiagarajan, M., Sturn, A., Snuffin, M., Rezantsev, A., Popov, D., Ryltsov, A.,Kostukovich, E., Borisovsky, I., Liu, Z., Vinsavich, A., Trush, V., Quackenbush, J., 2003.TM4: a free, open-source system for microarray data management and analysis.Biotechniques 34, 374–378.
Salama, M., Sandine, W., Giovannoni, S., 1991. Development and application ofoligonucleotide probes for identification of Lactococcus lactis subsp. cremoris.Applied and Environmental Microbiology 57, 1313–1318.
Samarzija, D., Sikora, S., Redzepovik, S., Antunac, N., Havranek, J., 2002. Application ofRAPD analysis for identification of Lactococcus lactis subsp. cremoris strains isolatedfrom artisanal cultures. Microbiological Research 157, 13–17.
Sandine, W.E., 1988. New nomenclature of the non-rod-shaped lactic acid bacteria.Biochemistry 4, 519–521.
Sheehan, A., O'Loughlin, C., O'Cuinn, G., FitzGerald, R.J., Wilkinson, M.G., 2005.Cheddar cheese cooking temperature induces differential lactococcal cellpermeabilization and autolytic responses as detected by flow cytometry:implications for intracellular enzyme accessibility. Journal of Applied Microbi-ology 99, 1007–1018.
Siezen, R.J., Renckens, B., van Swam, I., Peters, S., van Kranenburg, R., Kleerebezem, M.,de Vos, W.M., 2005. Complete sequences of four plasmids of Lactococcus lactissubsp. cremoris SK11 reveal extensive adaptation to the dairy environment.Applied and Environmental Microbiology 71, 8371–8382.
Siezen, R.J., Starrenburg, M.J., Boekhorst, J., Renckens, B., Molenaar, D., van HylckamaVlieg, J.E.T., 2008. Genome-scale genotype-phenotype matching of two Lactococcuslactis isolates from plants identifies mechanisms of adaptation to the plant niche.Applied and Environmental Microbiology 74, 424–436.
Simon, R., Lam, A., Li, M.C., Ngan, M., Menenzes, S., Zhao, Y., 2007. Analysis of geneexpression data using BRB-array tools. Cancer Informatics 3, 11–17.
Smit, G., Smit, B.A., Engels, W.J., 2005. Flavour formation by lactic acid bacteria andbiochemical flavour profiling of cheese products. FEMS Microbiology Reviews 29,591–610.
St-Gelais, D., Roy, D., Haché, S., 1992. Growth and activities of Lactococcus lactis in milkenriched with low mineral retentate powders. Journal of Dairy Science 75,2344–2352.
St-Gelais, D., Roy, D., Haché, S., Desjardins, S., 1993. Growth of nonproteolyticLactococcus lactis in culture medium supplemented with different casein hydro-lyzates. Journal of Dairy Science 76, 3327–3337.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. Molecular Evolutionary GeneticsAnalysis (MEGA) software version 4.0. Molecular Biology and Evolution 24,1596–1599.
van Kranenburg, R., Kleerebezem, M., van Hylckama Vlieg, J.E.T., Ursing, B.M.,Boekhorst, J., Smit, B.A., Ayad, E.H.E., Smit, G., Siezen, R.J., 2002. Flavour formationfrom amino acids by lactic acid bacteria: predictions from genome sequenceanalysis. International Dairy Journal 12, 111–121.
Vedamuthu, E.R., Sandine,W.E., Elliker, P.R., 1966. Flavor and texture in cheddar cheese.I. Role of mixed strain lactic acid starter cultures. Journal of Dairy Science 49,144–150.
Wang, H., Cronan, J.E., 2004. Only one of the two annotated Lactococcus lactis fabG genesencodes a functional α-ketoacyl-acyl carrier protein reductase. Biochemistry 43,11782–11789.
Wegmann, U., O'Connell-Motherway, M., Zomer, A., Buist, G., Shearman, C., Canchaya,C., Ventura, M., Goesmann, A., Gasson, M., Kuipers, O.S., Inderen, D.V., Kok, J., 2007.Complete genome sequence of the prototype lactic acid bacterium Lactococcuslactis subsp. cremoris MG1363. Journal of Bacteriology 189, 3256–3270.
Xia, X., Xie, Z., 2001. AMADA: analysis of microarray data. Bioinformatics 17, 569–570.
lymorphism among Lactococcus lactis subsp. cremoris strains usingMicrobiology (2010), doi:10.1016/j.ijfoodmicro.2010.02.020





























