Embed Size (px)
Citation preview

Free Radical Biology & Medicine, Vol. 36, No. 5, pp. 641 –656, 2004Copyright D 2004 Elsevier Inc.
Printed in the USA. All rights reserved0891-5849/$-see front matter
doi:10.1016/j.freeradbiomed.2003.12.004
Original Contribution
EVIDENCE FOR INTRIGUINGLY COMPLEX TRANSCRIPTION OF HUMAN
THIOREDOXIN REDUCTASE 1
ANNA-KLARA RUNDLOF, * MAGNUS JANARD,* ANTONIO MIRANDA-VIZUETE,yand ELIAS S.J. ARNER*
*Medical Nobel Institute for Biochemistry, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm,Sweden and yCenter for Biotechnology, Department of Biosciences at NOVUM, Karolinska Institutet, Huddinge, Sweden
(Received 10 October 2003; Revised 4 December 2003; Accepted 5 December 2003)
Ad
Nobel
and Bi
Fax: +
Elias S
Abstract—Human thioredoxin reductase 1 (TrxR1, the TXNRD1 gene product) is a ubiquitously expressed selenoprotein
with many important redox regulatory functions. In this study, we have further characterized the recently identified core
promoter region of TXNRD1. One critical Sp1/Sp3 site was found to be important in A549 and HeLa cells, whereas
another Sp1/Sp3 site and one Oct1 site bound transcription factors but were, nonetheless, dispensable for transcription.
We also experimentally identified several 5V-region TXNRD1 transcript variants using 5V-RACE with cDNA derived from
different tissues, and we analyzed all available TXNRD1-derived EST sequences. The results show that the core promoter
governs transcription of the clear majority of TXNRD1 transcripts but also that alternative promoters may be activated
under rare conditions or in specific cell types. Furthermore, extensive alternative splicing occured in the 5Vregion of
TXNRD1. In total, 21 different transcripts were identified, potentially encoding five isoforms of TrxR1 carrying
alternative N-terminal domains. One isoform encompassed a glutaredoxin domain, whereas another encoded a predicted
mitochondrial localization signal. These results reveal that the human thioredoxin system is intriguingly complex. Cell-
specific transcription of the TXNRD1 gene encoding different isoforms of TrxR1 must be taken into account to fully
understand the functions of the human thioredoxin system. D 2004 Elsevier Inc. All rights reserved.
Keywords—Thioredoxin reductase, Alternative splicing, Promoter, Sp1, Sp3, Free radicals
INTRODUCTION
The thioredoxin system is found in most organisms and
consists of thioredoxin (Trx), thioredoxin reductase
(TrxR), and NADPH. Trx is a small, well-characterized,
ubiquitous redox-active protein that is reduced by TrxR
using NADPH as a source of reducing equivalents. Trx
contains an active site with two redox-active cysteine
residues and participates in many different types of
reactions including synthesis of deoxyribonucleotides,
redox control of transcription factors, and regulation of
apoptosis [1,2].
Mammalian thioredoxin reductases are selenocysteine-
containing oxidoreductase flavoproteins with a broad
substrate specificity, catalyzed by a carboxyterminal ac-
dress correspondence to: Dr. Anna-Klara Rundlof, Medical
Institute for Biochemistry, Department of Medical Biochemistry
ophysics, Karolinska Institute, SE-171 77 Stockholm, Sweden.
46-8-31-15-51; E-mail: [email protected]; or Dr.
.J. Arner, [email protected].
641
tive site structure involving a conserved Gly-Cys-Sec-Gly
tetrapeptide [3–7]. The enzymes reduce not only protein
disulfides, such as those in oxidized Trx, but also low
molecular weight disulfide compounds, such as DTNB [8]
and lipoic acid [9], as well as low molecular weight non–
disulfide compounds, such as selenite [10], alloxan [11],
or peroxides [12]. Three different human genes that
encode TrxR isoenzymes have been identified: (i) the
classical cytosolic TrxR1, (ii) a mitochondrial enzyme
(TrxR2) [13–15], and (iii) one isoenzyme expressed
mainly in testis [13]. These three proteins have the same
overall domain structure. However, the testis-specific
enzyme has an additional N-terminal glutaredoxin domain
with a monothiol-active site motif and can, in contrast to
TrxR1 and TrxR2, also catalyze the direct reduction of
both glutathione disulfide and thioredoxin. It has, there-
fore, been named ‘‘TGR’’ for thioredoxin and glutathione
reductase [16].
All mammalian TrxR transcripts contain a selenocys-
teine insertion sequence (SECIS) element in the 3V-un-translated region (UTR) necessary for selenocysteine

A. -K. RUNDLOF et al.642
incorporation [5,17–20]. The 3V-UTR of transcripts de-
rived from TXNRD11 also contains AU-rich elements
(AUREs) that lead to a rapid mRNA turnover [18,21].
AUREs are typically found in mRNA with rapid post-
transcriptional up- and downregulation, such as cytokine,
proto-oncogene, transcription factor, and other transiently
expressed mRNAs [22]. The fact that TXNRD1 transcripts
have functional AUREs is interesting because this en-
zyme is not transiently expressed only in certain cell types
or under specific growth conditions, rather it is widely
expressed (transcribed) in many diverse tissues [14,23,24]
and cells [25].
We recently reported on the initial characterization of
the proximal promoter for human TXNRD1 [26]. Deletion
constructs were made and revealed that the promoter
activity was maintained within the �115 to +167 region
in all cell lines tested. This region was, therefore, consid-
ered to be the core promoter region. The core promoter
lacked classical TATA or CCAAT boxes but had an
increased GC content. It contained a POU motif that was
shown to bind the Oct1 transcription factor and two GC-
rich regions that bound Sp1 and Sp3. Lack of classical
TATA or CCAAT boxes, an increased GC content with
functional Sp1 site(s) in the proximal promoter region, and
a predicted CpG island close to the transcriptional initia-
tion site are features typical of housekeeping genes
[27,28]. Also Oct1 is ubiquitously expressed and believed
to govern the transcription of many housekeeping genes
[29]. A housekeeping-type promoter in combination with
AUREs in the 3V-UTR of TXNRD1 transcripts suggest an
intricate regulation pattern for this enzyme [26]. Further-
more, a second level of regulation may derive from
alternative splicing events.
We and others have shown that mammalian TrxR1
proteins may exhibit alternative splicing around the first
exon [30–32]. At least three forms of transcripts differing
in the 5Vend have hitherto been identified in mice and rats,
and five such forms have hitherto been identified in
humans [30–32]. One of the alternative murine splice
variants contains an additional upstream in-frame ATG,
which could encode an N–terminally elongated protein of
67 kDa instead of the common 55 kDa form [31].
However, that mRNA form could not be found in humans
[32], although a human TrxR1 variant protein with an
1 The approved gene symbol encoding human TrxR1 is TXNRD1
[33]. The nomenclature for thioredoxin reductase has varied throughout
the years; however, in most studies, the enzyme is generally referred to as
TrxR1. In this report, we refer to the gene as TXNRD1 and the protein
product in general discussion as TrxR1. Due to the large number of
transcript variants, though, the specific isoforms of TrxR1 encoded by the
different splice variants described herein are referred to as TXNRD1_v1
to TXNRD1_v5, according to the recommendations for isoform
nomenclature given by the Human Genome Organization [33].
apparent mass of 67 kDa was detected in the JPX9 cell line
of T-cell origin [31].
In this study, we continued to analyze the human
TXNRD1 core promoter with point mutations in the
previously studied Sp1/Sp3- and Oct1-binding sites, and
we identified one of the two Sp1/Sp3 sites as being
important for the basal promoter activity. We also
cloned and sequence-determined 5V-RACE variants of
human TXNRD1 transcripts originating from different
tissues and analyzed TXNRD1-specific EST sequences.
We showed that the clear majority of all TXNRD1
transcripts initiated at the site of the core promoter.
However, we also found an extensive alternative splic-
ing pattern in the 5V region of TXNRD1. In total, we
identified 21 different exon combinations and evidence
for rare alternative promoters. Moreover, we identified
five different N-terminal domains that may be encoded
by these mRNA variants, three of which have not been
described previously. These results reveal that, despite a
housekeeping-type core promoter, transcription of
TXNRD1 seems to be regulated in an exceedingly
complex manner.
MATERIALS AND METHODS
5V-RACEs of TrxR1-related transcripts from human
cDNA libraries
Marathon-ready cDNA libraries (Clontech, Palo Alto,
CA, USA) from four different tissues (mammary gland,
ovary, testis, and thymus) were used to amplify the 5Vendsof human TrxR1. The libraries were used together with
three gene-specific primers (P1, 5V-GCT TCT ATC ATT
CTG TCC CAATCATG-3V; P2, 5V-GAC ATG CTGAAG
CTT TGTGTGACC-3V; and P3, 5V-GCAACC CACATT
CAC ACA TGT TCC-3V) complementary to different
parts of the TXNRD1 cDNA, as described in the second
section of Results. The obtained products were analyzed
on 1% agarose gels isolated, and cloned into pGEM-T
vectors (Promega, Madison, WI, USA). The plasmids
were propagated in XL1-Blue Supercompetent cells
(Stratagene, La Jolla, CA, USA) and sequenced. The
sequences were deposited in GenBank with accession
numbers given in Table 1.
Sequence analyses
The GenBank EST database was searched for
TXNRD1-specific sequences using the here-identified
5V-RACE products as queries, which were aligned using
the Pairwise BLAST Tool [National Center for Biotech-
nology Information (NCBI), Bethesda, MD, USA] and/
or the GeneJockeyII Sequence Processor (Biosoft, Cam-
bridge, England). Protein characteristics were analyzed
with PEPSTATS (http://www.ebi.ac.uk/emboss/pepinfo/

5Vvariants of TXNRD1 transcripts 643
index.html), and prediction of cellular localization was
analyzed with iPSORT and/or PSORT II (http://psort.
ims.u-tokyo.ac.jp/). Intron-Exon boundaries were
inspected manually using the ‘‘show sequence’’ feature
of the NCBI map viewer display of the human genome
(http://www.ncbi.nlm.nih.gov/mapview/maps.cgi).
Construction of Sp1 and Oct1 mutants of the TXNRD1
core promoter
To construct mutants of the Sp1/Sp3 and the Oct1 sites
in the core promoter of TXNRD1, the following primers
were used (with the nucleotides shown in italics intro-
ducing the desired point mutations): (i) Sp1A, 5V-GCCCGC TCG GCG CAG TTT GTG GCT TCT CGT-3V; (ii)Sp1B, 5V-GCC CGC TCG GTT TAG GGC GTG GCT
TCT CGT-3V; (iii) Sp1C, 5V-GCC CGC TCG GCT TAG
TTC GTG GCT TCT CGT-3V; and (iv) Oct1, 5V-AGC TTA
CTA GGC AGC GAG CAT AGG TTG CC-3V. The
QuickChange Site-Directed Mutagenesis kit (Stratagene)
was used and a polymerase chain reaction (PCR) was run
with the core promoter–luciferase construct called HA
[26] as the template and using 16 cycles of 95jC for 40 s,
55jC for 1 min, and 68jC for 12 min, according to the
manufacturer’s protocol. The PCR products were treated
with DpnI, religated, and propagated in XL1-Blue Super-
competent cells (Stratagene); the sequences and desired
mutations were verified.
Cell cultures
The A549 (human lung carcinoma) and HeLa (hu-
man cervix carcinoma) cell lines were grown in Dul-
becco’s modified Eagle medium, supplemented with
10% heat-inactivated fetal calf serum, 2 mM L-gluta-
mine, 100 units/ml penicillin, and 100 Ag/ml strepto-
mycin, in an atmosphere of 5% CO2 and 95% air at
37jC.
Determination of reporter gene activity
Approximately 3 � 104 cells per well in 24-well
dishes were seeded 24 h before transfection using the
mutated TXNRD1 promoter reporter constructs (0.5 Ag)and cotransfection with 0.5 ng pRL-CMV plasmid.
The transient transfections were performed using Lip-
ofectamine Plus Reagent (GIBCO, Grand Island, NY,
USA), with 2 Ag lipofectamine and 3 Al Plus reagent
for 4 h, according to the manufacturer’s protocol. After
24 h, the cells were washed with phosphate-buffered
saline and lysed in 100 Al Passive Lysis Buffer
(Promega, Madison, WI, USA) at room temperature
for 15 min. The firefly and renilla luciferase activities
in 10 Al of the cell lysates were determined using the
Dual-Luciferase Reporter Assay system (Promega),
according to the manufacturer’s instructions, and a
Turner Designs TD-20/20 luminometer.
Nuclear extracts
Nuclear extracts were prepared from 1 � 107 A549 or
HeLa cells, grown in large culture plates, which were
scraped into PBS and centrifuged for 10 min at 1800 � g.
The cells were resuspended in 600 Al hypotonic buffer [10mM HEPES (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 0.2
mM PMSF, and 0.5 mM DTT] and centrifuged at 1800 �g for 5 min at 4jC. The pellet was resuspended in 400
Al hypotonic buffer and incubated for 10 min on ice; next,
the cells were homogenized with 10 strokes using a
Dounce homogenizer with a B-type pestle. The homog-
enate was centrifuged for 15 min at 3300 � g at 4jC, thesupernatant was removed, and the nuclei in the pellet
were resuspended in 180 Al low-salt buffer [20 mM
HEPES (pH 7.9), 25% glycerol, 1.5 mM MgCl2, 10
mM KCl, 0.2 mM EDTA, 0.2 mM PMSF, and 0.5 mM
DTT]. KCl was added to a final concentration of 0.4 M
while stirring gently with a pipette tip. Nuclear proteins
were extracted for 30 min on ice with continuous gentle
mixing. After centrifugation for 30 min at 25,000� g and
4jC, the supernatant with extracted proteins was frozen
immediately at �80jC in aliquots, which were thawed
gently on ice at the time of use in electrophoretic mobility
shift assays (EMSA).
Electrophoretic mobility shift assays
The EMSA probes (Invitrogen, San Diego, CA, USA)
used were the following (sense sequence shown): (i) Sp1-
control, 5V-CCCGCTCGGC GCAGGGCGTG GCTTCT-
CGTA GCCATTAGGA-3V; (ii) Oct1-control, 5V-TCT-CAGCTTA CTAGGCAATT AGCATAGGTT
GCCAGGGCTG-3V; (iii) Sp1A-mut, 5V-CCCGCTCGGCGCAGTTTGTG GCTTCTCGTA GCCATTAGGA-3V;(iv) Sp1B-mut, 5V-CCCGCTCGGT TTAGGGCGTG G-
CTTCTCGTA GCCATTAGGA -3V; (v) Sp1C-mut, 5V-CCCGCTCGGC TTAGTTCGTGGCTTCTCGTA GCC-
ATTAGGA-3V; and (vi) Oct1-mut, 5V-TCTCAGCTTACTAGGCAGCG AGCATAGGTT GCCAGGGCTG-3V.Double-stranded oligonucleotides were generated by
annealing equimolar complementary oligonucleotides in
50 mM Tris-HCl (pH 7.4), 1 mM EDTA, 100 mM NaCl,
and 13 mM MgCl2 with the following temperature
program: 88jC for 2 min, 65jC for 10 min, 37jC for
10 min, and 25jC for 5 min. The double-stranded
oligonucleotides were end-labeled with [g-32P]ATP
(3000 mCi/mmol; DuPont NEN, Wilmington, DE,
USA) using T4 polynucleotide kinase, and the labeled
probes were purified by Chroma-spin 30 (Clontech). For
binding assays, a mixture was prepared containing [32P]-
labeled oligonucleotide (0.3 ng), 3 Ag nuclear proteins,

A. -K. RUNDLOF et al.644
and 1 Ag poly(dIdC) (Amersham, Arlington Heights, IL,
USA), adjusted to 20 Al with binding buffer [20 mM
HEPES (pH 7.9), 1.5 mM MgCl2, 0.2 mM EDTA, 1 mM
DTT, 0.1 mM PMSF, 7.5% glycerol]. Binding reactions
were allowed for 15 min at 28jC; next, a 15 Al aliquot ofeach reaction was loaded onto a 4% nondenaturing
polyacrylamide gel and run in 1� Tris–Glycine buffer
at 120 V. Following electrophoresis, gels were dried and
autoradiographed.
RESULTS
Transcriptional activity of the human TXNRD1 core
promoter with point-mutated Sp1/Sp3 and Oct1 sites
In a previous study, we cloned and characterized the
promoter region of human TXNRD1 [26] and found a
predicted POU domain (�98 to �90) in the core promot-
er, which was functional in binding the Oct1 transcription
factor. Immediately upstream of the transcription start
point, a GC-rich region was present (�17 to �7), which
also bound the Sp1 and Sp3 transcription factors [26]. In
this study, we wished to verify the function of these
transcription factor motifs in human cells. Therefore, the
Oct1-binding site (�98 to �90) was mutated from
ATTAGCATA to GCGAGCATA (Oct1m); and, in case
of the Sp1/Sp3 tandem–binding site (�17 to �7), three
different mutants were made, with the sequence changed
from GCGCAGGGCGT to GCGCAGTTTGT (Sp1Am),
to GTTTAGGGCGT (Sp1Bm), or to a combination of the
two mutated consensus motifs GCTTAGTTCGT
(Sp1Cm).
The mutated oligonucleotides were first analyzed using
EMSA with nuclear extracts from either A549 or HeLa
cells. The electrophoretic mobility shift caused by Oct1
was abolished using the Oct1mut oligonucleotide, where-
as the shifts caused by Sp1/Sp3 could still be detected with
the Sp1Bmut variant but not with Sp1Amut or Sp1Cmut
(Fig. 1). Reporter constructs for the core promoter having
the corresponding mutations were then used for trans-
fections of A549 and HeLa cells, and the transcriptional
activity of the luciferase reporter gene was measured. As a
negative control, a construct (known to be transcription-
ally inactive [26]) with the upstream region cloned in
reverse orientation (HB) was used, while the wild-type
reporter construct (HA) served as a positive control. In
both cell lines, the Sp1Bmut and Oct1mut constructs
showed a luciferase activity comparable with the wild-
type HA construct, whereas Sp1Amut and Sp1Cmut gave
reduced transcriptional activities. The luciferase activities
of Sp1Amut and Sp1Cmut in A549 cells were more
decreased (50%) than in HeLa cells (25%; Fig. 2). These
results show that the POU site and the second Sp1/Sp3
consensus motif covered by the Oct1mut and Sp1Bmut
constructs are dispensable for basal TXNRD1 transcription
in A549 and HeLa cells. The Sp1/Sp3 site covered by the
Sp1Amut, on the other hand, contributes to a major part of
the basal TXNRD1 transcriptional activity in these cells.
Cloning of cDNA variants of TXNRD1-derived
transcripts differing in the 5Vregion
As mentioned in the Introduction, we and others have
previously reported the existence of up to five separate
splice variants of the 5V-UTR of human TXNRD1. Those
variants were mostly discovered only on the basis of
database searches with identification of EST sequences
derived from high-throughput screening projects. Since
different 5V-UTR sequences in theory may reveal the
existence of alternative promoters or transcriptional start
sites, we decided to focus on the possible presence of 5V-UTR variants using an experimental approach. The
results confirmed that the core promoter described to
date is the major promoter for human TXNRD1, but they
also revealed a surprisingly high number of alternative 5Vvariants.
To examine human TXNRD1 5V-region variants, we
screened four separate cDNA libraries from different
tissues (mammary gland, testis, ovary, and thymus). Three
primers complementary to three strategic parts of the
TXNRD1 sequence were designed for these 5V-RACEs.One primer (P1) was based on the region downstream of
the well-known ATG (present in what we subsequently
named Exon III); another primer (P2) was based on an
exon covering an alternative upstream in-frame ATG
(Exon I), resulting in the previously published ‘‘KDRF’’
variant of TrxR1 [21]. Finally, the third primer (P3) was
complementary to the nucleic acid sequence encoding the
N-terminal disulfide/dithiol–active site CVNVGC (Exon
V). These three primers generated several separate
TXNRD1-specific RACE products with different lengths
and pattern in the examined tissues. In total, we charac-
terized 44 different PCR products, the analysis of which
revealed 18 separate types of transcripts for TXNRD1. We
then complemented this information with an extensive
GenBank database search that identified EST clones that
supported these results. Furthermore, some EST sequen-
ces indicated the presence of additional splice variants that
were not identified in our experimental approach, which
together gave 21 separate splice variants. All exons could
be identified in the human genomic sequence covering the
TXNRD1 gene on chromosome 12. To discuss and de-
scribe these identified transcript variants, we have used the
following nomenclature:
(1) Exons present in the most common transcripts are
given roman numerals (I, II, III, IV, V, and so on) in

Fig. 1. EMSAwith Sp1/Sp3 and Oct1 motif mutants. The core promoter, with previously identified transcription factor–binding regions[26], is shown schematically in the upper part of the figure with the denoted transcription start site indicated by an arrow. Double-stranded 40-mer DNA oligonucleotides were made with either wild-type sequence or carrying a mutated Oct1-binding site (Oct1m), orseparate mutations of the Sp1/Sp3-binding sites (Sp1Am, Sp1Bm, and Sp1Cm). The relevant transcription factor–binding motifs areshown beneath the scheme of the promoter, with the mutated nucleotides boxed. EMSAwere performed with 0.3 ng 32P-labeled double-stranded oligonucleotides and 1 Ag poly(dI � dC). Lanes A are negative controls without the addition of nuclear extract; in lanes B and C,3 Ag protein of nuclear extract from A549 cells were added; in lanes C, an excess (20 ng) of the homologous but unlabeledoligonucleotide was also added for competition to show specific binding. No shifts could be detected using the Oct1m, Sp1Am, orSp1Cm oligonucleotides, whereas Sp1Bm generated three shifts identical to the wild-type Sp1 control. An identical pattern was obtainedusing nuclear extract from HeLa cells (not shown).
5Vvariants of TXNRD1 transcripts 645
order of appearance in the genome. The transcrip-
tional start site is indicated in parentheses after the
first exon, with ‘‘1’’ either being the previously
denoted transcriptional start site for exon I [26] or
the first nucleotide after a splice site of a regular
exon, in case a transcript initiates with an exon
other than exon I. Variant exons extended at the 5Vend are denoted with a ‘‘v’’ prefix (e.g., vII),
whereas exons with alternative donor splice sites
are denoted with a small letter suffix (e.g., the exon

Fig. 2. Transcriptional activity of the mutated luciferase reporter constructs. The wild-type luciferase reporter construct (HA) contained0.8 kb of the upstream region of TXNRD1, with the reverse orientation of the sequence named HB, which was used as a negative control.Mutated luciferase reporter constructs (Sp1Am, Sp1Bm, Sp1Cm, and Oct1m) carrying the same point mutations as analyzed in Fig. 1were also made. Luciferase activities were subsequently measured after transfections of either A549 or HeLa cells. The activities shownare the combined mean values F SD of the firefly luciferase measurements from three different experiments performed in triplets. A50% decrease of the luciferase activity was observed with the Sp1Am and Sp1Cm constructs in A549 cells and a 25% reduction wasseen in HeLa cells. The Oct1m and Sp1Bm constructs had the same activities as the positive HA control in both A549 and HeLa cells.
A. -K. RUNDLOF et al.646
Ia is a shorter version of Ib due to the use of an
alternative donor splice site).
(2) Rare exons are named with Greek letters and
consecutive subscript roman number (e.g., gI, gII,
and gIII, orh-I,h-II, andh-III), where the number gives
the order of the exons as counted from the core
promoter and a negative number indicates an up-
stream location.
(3) Transcripts containing only the most common exons
are called a transcripts, those containing h exons are
named h transcripts, and g transcripts contain g
exons. Transcripts that differ due to alternative
splicing patterns are given consecutive numbers
(e.g., a1, a2, and a3).
(4) Potential protein isoforms encoded by the different
splice variants having alternative N-terminal do-
mains are specifically named as TXNRD1_v1,
TXNRD1_v2, and so on, following the guidelines
of the HUGO Gene Nomenclature Committee [33].
All the TXNRD1-derived 5V-variant transcripts identi-fied so far are listed in Table 1 and schematically drawn
in Fig. 3. The intron-exon boundaries are given in Table
2 and variant N-terminal domains are shown in Fig. 4.
Intron-exon boundaries
Genomic sequences of most intron-exon boundaries
followed splicing consensus, although several deviations
could also be noted (Table 2). Furthermore, sizes of exons
and introns for the ‘‘classical’’ splicing events of
TXNRD1, i.e., involving exons I–XV, as identified here
(Table 2) deviate in several instances from a prior similar
analysis [32]. A possible reason for these deviations could
be the continued curation of the available genomic
sequence information. The data presented in this study
is based on the April 14, 2003 release (see ‘‘Build 33’’ on
www.ncbi.nlm.nih.gov/genome/guide/human/release_
notes.html), which is considered to encompass an essen-
tially completed chromosome 12 sequence.
Variants of first exon and transcriptional start site
It became evident that the clear majority of the
TXNRD1 transcripts began with exon I and were initiated
by the core promoter. Of the transcripts initiating with
exon I, several variants were identified. Two major
alternative transcription start sites seem to be present,
either around nt �2 to 20 (transcripts a1, a7, a10, or
a12) or around nt 50 to 70 (transcripts a2, a8, a9, a11,
and g3). The longest variants having a transcriptional
start site at �2 to 20 are derived from the region
immediately following the tandem Sp1/Sp3 site, which
we have further characterized here in Figs. 1 and 2.
Exon I showed alternative splicing patterns, apparently
with three separate donor splice sites. The first donor site,
resulting in the Ia or vIa forms, was used in transcriptsa6–
a11 and g2 and g3; it seems to be the most common splice

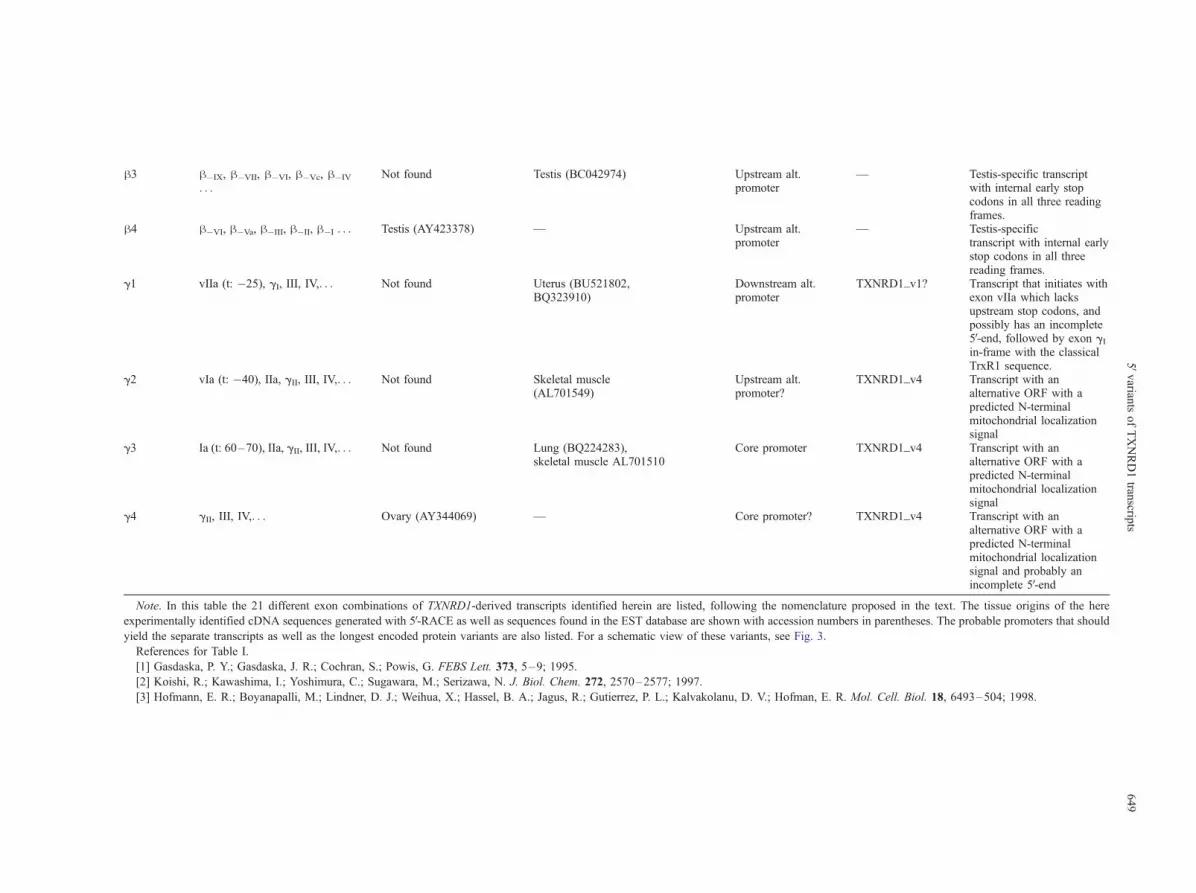
Table 1. TXNRD1 cDNA 5V-End Variants
TXNRD1transcript
Exon combination in 5V-region Tissue origins of cDNAlibrary for transcripts
experimentally identifiedin this study(accession no.)
Tissue origins ofcorresponding ESTs
(accession no.)
Probable promoter Longest encodedTrxR1 isoform
Comments
a1 Ib (t: �2–20), IIa, III, IV,. . . Thymus (AY344075,AY344076), mammarygland (AY344072,AY344073, AY344074)
Testis (BQ048924) Core promoter TXNRD1_v2 Transcript which initiatesclose to the Sp1/Sp3 sitestudied herein.
a2 Ib (t: 50–70), IIa, III, IV,. . . Testis (AY344077),mammary gland(AY344070, AY344071)
Uterus (BQ222639), skin(BQ678794), skeletalmuscle (BX470432,AL701384, BX470447,AL701440, AL701548),unknown (AU100288)
Core promoter TXNRD1_v2 Same as published cDNAfor human TrxR1, derivedfrom placenta [1].
a3 Ib (t: 150–300), IIa, III, IV,. . . Ovary (AY344078,AY344079)
Skin (BU166571), lung(BF182740), uterus(BM476786)
Core promoter? TXNRD1_v2 If t: 255, then same as‘‘KDRF’’ (cDNA frombone-marrow-derivedstromal cell line [2]).
a4 Ic (t: 50), IIa, III, IV,. . . Not found Embryonic stem cell(CD652517)
Core promoter TXNRD1_v1 Use of rare alternativedonor splice site in exon I.
a5 Ib (t: 60–70), III, IV,. . . Not found Unknown (AU077310),bone (BQ014595)
Core promoter TXNRD1_v1 Rare transcript whichinitiates at a secondtranscriptional start site ofthe core promoter.
a6 vIa (t: �96;�40), IIa, III,IV,. . .
Testis (AY344083) Testis (BG771986,BG771648)
Upstream alt.promoter?
TXNRD1_v1 Rare upstreamtranscriptional start site.
a7 Ia (t: �2–20), IIa, III, IV,. . . Testis (AY344081,AY344082), thymus(AY344087), mammarygland (AY344090,AY344091)
Pre-B-cell (BE247591),neuroepithelium (AA173096),nasopharynx(CD697218), thymus(BX436752), testis(BM463380), mixed(BU601103)
Core promoter TXNRD1_v1 Common transcript, whichresults in the classicalamino acid sequence andinitiates close to the Sp1/Sp3 site studied herein.
a8 Ia (t: 50–70), IIa, III, IV,. . . Testis (AY344080,AY344677), ovary(AY344085, AY344086,AY344678, AY344680),thymus (AY344088,AY344089), mammarygland (AY344092,AY344093, AY344674)
Brain (BQ070241,BE314677, BF206572),uterus (BU522017),embryonic stem cell(CD652543, AU280829),mixed (BU940667),cervix (BI088157),skeletal muscle(AL705247),teratocarcinoma (AU128350), lung(BE785986, BE789006,BG331212, AA376292,AA375636), smallintestine (BM801989),
Core promoter TXNRD1_v1 Common transcript,encodes the classical aminoacid sequence and initiatesat a second transcriptionalstart site within the corepromoter.
(continued on next page)
5Vvarian
tsofTXNRD1tran
scripts
647

Table 1 (continued)
TXNRD1transcript
Exon combination in 5V-region Tissue origins of cDNAlibrary for transcripts
experimentally identifiedin this study(accession no.)
Tissue origins ofcorresponding ESTs
(accession no.)
Probable promoter Longest encodedTrxR1 isoform
Comments
prostate (BQ936015),adipose (CA405944),bone (AI751930), skin(BF692051), stomach(BM826421), eye(BE779822)
a9 Ia (t: 60–70), IIb Not found Stomach (BM826421) Core promoter — Alternative exon IIextended downstream365 nt.
a10 Ia (t: �2–20), III, IV,. . . Mammary gland(AY344097)
Placenta (CB989925,CB989312)
Core promoter TXNRD1_v1 Transcript with rare splicingevent which originates at asecond transcriptional startsite within the corepromoter.
a11 Ia (t: 50–70), III, IV,. . . Ovary (AY344084,AY344669, AY344673),mammary gland(AY344675)
Lymph (BM477984),teratocarcinoma (AU132293), eye(BE618239), lung(BE786133), brain(BX461036)
Core promoter TXNRD1_v1 Transcript with rare splicingevent which originates atthe Sp1/Sp3 site studiedherein.
a12 IIa (t: 1–20), III, IV,. . . Ovary (AY344094),thymus (AY344095),mammary gland(AY344096, AY344678,AY344679)
Placenta (BE408950),uterus (BU507940),embryo (AA333346)
Downstream alt.promoter or corepromoter?
TXNRD1_v1 Transcript originating withexon II and which is thesame as the GRIM-12sequence derived frombreast carcinoma [3].
a13 vIIa (t: -120–30), III, IV,. . . Testis (AY344670,AY344671, AY344672)
Testis (BG772373),trachea (AK128515)
Downstream alt.promoter
TXNRD1_v5 Transcript initiating with arare form of exon II thatresults in a previouslyunknown ORF for analternative N-terminaldomain.
h1 h�VIII, h�VI, h�Va, IIa,III, IV,. . .
Testis (AY057105) Testis (BG772375,BG717223)
Upstream alt.promoter
TXNRD1_v3 An apparently testis-specifictranscript originating froman upstream alternativepromoter, which encodes analternative ORF with anadditional N-terminalGrx-like domain.
h2 h�IX, h�VII, h�VI, h�Vb . . . Not found Testis (BI559668,BI559562)
Upstream alt.promoter
— Testis-specific transcriptwith internal early stopcodons in all three readingframes.
A.-K.RUNDLOF
eta
l.648
h3 h�IX, h�VII, h�VI, h�Vc, h�IV
. . .Not found Testis (BC042974) Upstream alt.
promoter— Testis-specific transcript
with internal early stopcodons in all three readingframes.
h4 h�VI, h�Va, h�III, h�II, h�I . . . Testis (AY423378) — Upstream alt.promoter
— Testis-specifictranscript with internal earlystop codons in all threereading frames.
g1 vIIa (t: �25), gI, III, IV,. . . Not found Uterus (BU521802,BQ323910)
Downstream alt.promoter
TXNRD1_v1? Transcript that initiates withexon vIIa which lacksupstream stop codons, andpossibly has an incomplete5V-end, followed by exon gIin-frame with the classicalTrxR1 sequence.
g2 vIa (t: �40), IIa, gII, III, IV,. . . Not found Skeletal muscle(AL701549)
Upstream alt.promoter?
TXNRD1_v4 Transcript with analternative ORF with apredicted N-terminalmitochondrial localizationsignal
g3 Ia (t: 60–70), IIa, gII, III, IV,. . . Not found Lung (BQ224283),skeletal muscle AL701510
Core promoter TXNRD1_v4 Transcript with analternative ORF with apredicted N-terminalmitochondrial localizationsignal
g4 gII, III, IV,. . . Ovary (AY344069) — Core promoter? TXNRD1_v4 Transcript with analternative ORF with apredicted N-terminalmitochondrial localizationsignal and probably anincomplete 5V-end
Note. In this table the 21 different exon combinations of TXNRD1-derived transcripts identified herein are listed, following the nomenclature proposed in the text. The tissue origins of the here
experimentally identified cDNA sequences generated with 5V-RACE as well as sequences found in the EST database are shown with accession numbers in parentheses. The probable promoters that should
yield the separate transcripts as well as the longest encoded protein variants are also listed. For a schematic view of these variants, see Fig. 3.
References for Table I.
[1] Gasdaska, P. Y.; Gasdaska, J. R.; Cochran, S.; Powis, G. FEBS Lett. 373, 5–9; 1995.
[2] Koishi, R.; Kawashima, I.; Yoshimura, C.; Sugawara, M.; Serizawa, N. J. Biol. Chem. 272, 2570–2577; 1997.
[3] Hofmann, E. R.; Boyanapalli, M.; Lindner, D. J.; Weihua, X.; Hassel, B. A.; Jagus, R.; Gutierrez, P. L.; Kalvakolanu, D. V.; Hofman, E. R. Mol. Cell. Biol. 18, 6493–504; 1998.
5Vvarian
tsofTXNRD1tran
scripts
649

Fig. 3. Genomic organization of TXNRD1 and transcript variants with different exon combinations. In the upper part of the figure, the genomic organization of human TXNRD1 is shown,with TXNRD1-specific exons indicated by different colors. Unrelated exons (see Discussion) are shown in black and indicated with their Unigene cluster identification numbers. Location ofthe previously characterized core promoter is indicated with a filled arrow; hollow arrows show possible alternative promoter regions. Also indicated above the genomic scheme are thespecific exons encoding the N-terminal CVNVGC redox-active motif and the large exon XV encoding the C-terminal GCUG tetrapeptide and the 3V-UTR. (A–C) Transcript variantscontaining different exon combinations in the 5Vregion are shown with possible ORFs indicated by black arrows. The encoded protein variant is indicated to the right of each transcript. (A)All variants containing solely the classical a exons; (B) transcripts containing upstream h exon; and (C) the transcripts carrying alternative g exons. See Table 1 for further information on theseparate transcripts and Fig. 4 for amino acid sequences.
A.-K.RUNDLOF
eta
l.650

Table 2. Sizes of Exons and Introns of the Transcript Variants of TXNRD1 and Sequences at Splice Site Boundaries
Donor exona (nt) Acceptor
exona (nt)
Intron
sizeb (kb)
Sequences from EXON/intron/EXONc (splice site consensus:
. . .AGjguragu. . .(y)10 – 15nyagjG. . .)
Ia (50–120) IIa (110) 1.81 . . .AUUCUUjguaagc. . .ugcuuuugugccacagjAGGACG. . .Ia (50–120) III (123) 24.1 . . .AUUCUUjguaagc. . . uguuauuuuucuuuagjGCUUAU. . .Ib (290–356) IIa (110) 1.58 . . .CAUGUGjguaggu. . .ugcuuuugugccacagjAGGACG. . .Ib (290–356) III (123) 23.9 . . .CAUGUGjguaggu. . .uguuauuuuucuuuagjGCUUAU. . .Ic (187) IIa (110) 1.71 AGCAGUjgugcgu. . .ugcuuuugugccacagjAGGACG. . .IIa (110) III (123) 22.2 . . .UUGAAGjguagga. . .uguuauuuuucuuuagjGCUUAU. . .IIa (110) gII (112) 1.95 . . .UUGAAGjguagga. . .acuuuuuuuucuuugajAGAUGG. . .vIIa (146) gI (79) 0.42 . . .UUGAAGjguagga. . .uuccucuuaauucucucagjGGGUAG. . .III (123) IV (73) 1.83 . . .GCUAAGjgcaagg. . .uaauuuuauuuuccagjGAGGCA. . .IV (73) V (120) 2.46 . . .AGAUGGjg!guaag. . .ucucaauguuguuguagjG!UCUCG. . .V (120) VI (143) 3.01 . . .AGACAGjguauga. . .guuuuauuuucuuauacagjUUAAGC. . .VI (143) VII (116) 0.41 . . .AUUAAGjguaauu. . .auugguuccuuuguagjGCAACA. . .VII (116) VIII (226) 1.50 CAGCAGjguaaag. . .uuuuuuuuucccccagjUGAUGA. . .VIII (226) IX (93) 4.01 . . .AUUAAAjguaagu. . .uuuaucacucuuacagjGUUGAA. . .IX (93) X (77) 0.91 . . .AAUACGjguaagg. . .auuaauaauuuuucagjGUGAUG. . .X (77) XI (161) 1.09 . . .UGAAAAjguaaga. . .cugaauaaaauugcagjGACUGG. . .XI (161) XII (103) 3.85 . . .UCAAG!GUGAjguguug. . .ccauuuuuaaacagu!gugajCUAUGA. . .XII (103) XIII (96) 1.50 . . .AUUGAGjguaagt. . .gcuuuuguaucuucuuagjGUUUAC. . .XIII (96) XIV (135) 4.80 . . .GACAAUjguaagu. . .uuuuuuuuugccuuagjGAACGU. . .XIV (135) XV (1932) 9.07 . . .GCAGAGjgugggu. . .uucucuuccccugcagjGUAUUC. . .h�IX (208) h�VII (87) 11.9 . . .CUCGCCjguaagu. . .uguuugaucaacccagjGAAUCU. . .h�VIII (115) h�VI (152) 35.6 . . .GGUCAGjguacga. . .cuuccauuacuucuagjCUAAAG. . .h�VII (87) h�VI (152) 26.6 . . .AGGCAGjguaaaa. . .cuuccauuacuucuagjCUAAAG. . .h�VI (152) h�V (62) 6.33 . . .ACUGAGjguaaag. . .uuuuucaacuucuuccagjGUAAAG. . .h�Va (62) II (110) 30.8 . . .AAACAGjguaagu. . .ugcuuuugugccacagjAGGACG. . .h�Va (62) h�III (82) 18.5 . . .AAACAGjguaagu. . .ucucuuuuccacuaacagjUCAAAA. . .h�Vc (1395) h�IV (1159) 1.02 .CAACAA!C!A!A!C!A!A|cagcaa. . .cuccagccuggg!c!a!a!c!a!ajGAGCACh�III (82) h�II (95) 6.19 . . .GCCAGGjguaaga. . .uuauaauuuuuuuuguagjAGACAA. . .h�II (95) h�I (131) 1.30 . . .ACAAGjguaagu. . .uuucuguguuuccuucagjCAAUGU. . .gI (79) III (123) 21.7 . . .CUUCAGjgugugu. . .uguuauuuuucuuuagjGCUUAU. . .gII (112) III (123) 20.2 . . .AGUGAGjguauag. . .uguuauuuuucuuuagjGCUUAU. . .
a The donor and acceptor exons are shown with the nomenclature given in the text and their sizes in nucleotides are shown in parentheses. For a
schematic figure showing splicing in the 5V-region, see Fig. 3.b Intron size resulting from the indicated splicing event is given in kilobases.c The boundaries of splice sites in the pre-mRNA sequence as inferred from the genomic DNA sequence are compared to the consensus of splice site
motifs, as compiled elsewhere [Bauren, G. (1996), Pre-mRNA processing in the polytene nuclei of Chironomus tentans, in Thesis, Department of Cell
and Molecular Biology, ISBN 91-628-2242-X, Karolinska Institute, Stockholm]. The four nucleotides surrounding the 5V-end of the intron at the splice
site are shown with bold letters, as are the three nucleotides at the 3V-side, with the splice site indicated with a vertical line. Alternative splice sites are
shown with exclamation marks. The polypyrimidine tract is given in italics. Deviations from the consensus are underlined.
5Vvariants of TXNRD1 transcripts 651
site of exon I. The rare, longer version of exon I (Ic), seen
only in transcript a4, was derived from an embryonic stem
cell. The third donor site gave exon Ib and was rather
common; in combination with exon II, this resulted in an
alternative and extended open reading frame. All splicing
events involving exon I conformed to splice site consensus
sequences in the intron, whereas the flanking exon sequen-
ces showed slight deviations from the general consensus
sequence (Table 2). Interestingly, transcript a6 (found in
cDNA from testis) and g2 (skeletal muscle) started with
exon I but initiated at �40 or �96, suggesting either that
the core promoter region (�115 to +167) may guide these
rare upstream transcriptional initiation sites or, alterna-
tively, that these forms are incomplete sequences repre-
senting transcripts initiating from alternative promoters
situated further upstream in the genome.
It should be noted that the variation of exon I length
varied depending on the primer that we had used in the 5V-
RACE as well as from which tissue the cDNA originated.
A complete long exon I (exon Ib) could only be obtained
with primer P2, which was complementary to the 3Vend ofexon Ib, likely indicating that exon Ib is only present in
rather rare transcripts, although it was also found in
numerous EST clones (see the a1, a3, and a5 transcripts;
Table 1).
Three clones that we amplified from the testis library
(transcript a13) were found to initiate with exon II
extending upstream of its common acceptor splice site,
and these were identical with parts of two ESTclones, one
originating from testis (accession number BG772373)
and one derived from trachea (accession number
AK128515). Moreover, one clone amplified from the
ovarian cDNA library initiated with a first exon that
normally is located within the second intron (exon gII,
see transcript g4). This exon contained an AluJ-like
element and was also found in three EST clones that also

Fig. 4. N-terminal domain variants of TXNRD1. Amino acid sequences of the five different N-terminal domains encoded by the TXNRD1gene are aligned in this figure, with shared identical motifs boxed. The corresponding exons are indicated above the protein sequences.The atypical active site of the glutaredoxin domain found in TXNRD1_v3 is underlined and the predicted cleavage site of themitochondrial localization signal in TXNRD1_v4 is indicated with an upward arrow. Amino acid numbering for each variant is given tothe right. Only sequences upstream of the shared classical N-terminal dithiol-active site motif are shown. Note that alternative splicingmay also occur in the 3Vregion of the gene (see text). The calculated properties of the unique peptides serving as N-terminal additions tothe classical TrxR1 amino acid sequence are shown in the lower right corner of the figure.
A. -K. RUNDLOF et al.652
contained exons II and Ia or vIa (transcripts g2 and g3).
Finally, rare transcripts that were derived only from testis
initiated far upstream of the core promoter (the h1–h4transcripts). The presence of these alternative first exons
in rare TXNRD1 transcripts show that alternative pro-
moters may be activated in certain cells.
Possible alternative open reading frames and protein
isoforms of TrxR1
When we examined the nucleotide sequences of the
different variant transcripts, we observed that some of
these encoded alternative 5V-extended open reading frames
in frame with the classical TrxR1 sequence. In total, five
potential N-terminal variants can be encoded. We called
these isoforms TXNRD1_v1 to TXNRD1_v5; the amino
acid sequences and their encoding exons are shown in Fig.
4 and are described briefly.
TXNRD1_v1. This is the classical form of mammalian
TrxR1 as generally purified from tissues such as liver,
thymus, or placenta [8,34,35]. It may be encoded by all
TXNRD1-derived transcripts carrying the combination of

5Vvariants of TXNRD1 transcripts 653
exons III, IV, V, and so on, in consecutive order (i.e., all a
transcripts except a9, all g transcripts, and possibly
transcript h1), with the initiating methionine codon being
located in exon III.
TXNRD1_v2. This form is identical to the previously
described KDRF and contains a potential N-terminal
elongation while still keeping the classical initiating
ATG in exon III. It may be encoded by transcripts a1 to
a3, carrying exons Ib, II, III, IV, V, and so on, in
consecutive order. Transfection of mammalian cells with
a construct identical to the a3 transcript showed that both
initiating ATGs (i.e., in exons Ib or III) could be func-
tional, resulting in a mixture of products between 55 kDa
and 60 kDa [21].
TXNRD1_v3. This form is encoded by the h1 transcript
and has not been described before. Interestingly, it
encodes a 70 kDa form carrying a large N-terminal
glutaredoxin domain with an atypical CTRC-active site
sequence. The properties of this protein will be described
elsewhere.
TXNRD1_v4. This form is encoded by the g2 to g4
transcripts and has not been described before. The
combination of exons gII and III yields an alternative
open reading frame with a 44 amino acid N-terminal
elongation while still keeping the classical initiating
ATG in exon III. When analyzed using PSORT, this
form is predicted with high probability to carry a
mitochondrial localization signal. This variant could
theoretically complement the function of mitochondrial
TrxR2 [13–15] and will be studied elsewhere in further
detail.
TXNRD1_v5. This variant represents an N-terminal
elongation encoded by exon vII, which is present in
transcript a13. The a13 transcript contained an extend-
ed ORF with possible predicted N-terminal signals for
both mitochondrial and nuclear localization (using
PSORT). An EST clone originating from trachea (ac-
cession number AK128515), which carried the 5Vend of
a13, was a long transcript that also encoded an extend-
ed and alternative C-terminal region of the protein as
the result of an intron retention in the 3V region. Thatinsertion generated a frame shift with a potential ex-
tended ORF harboring three in-frame UGA codons,
followed by an intact 3V-UTR containing the SECIS
element. If such a variant would be translated in
trachea, it could thereby contain an alternative C-termi-
nal domain carrying three selenocysteine residues but
not the classical C-terminal tetrapeptide motif of TrxR1.
The properties of this protein will be described in
further detail elsewhere.
DISCUSSION
In this study, we have continued our analysis of the
core promoter region of TXNRD1 with mutational anal-
ysis of the previously identified [26] Oct1- and Sp1/Sp3-
binding motifs. It is clear from the results presented here
that the first Sp1/Sp3-binding site in the tandem motif at
�17 to�7 contributes to about 50% of the transcriptional
activity in A549 and HeLa cells, while the other analyzed
Sp1/Sp3- and Oct1-binding motifs were dispensible for
transcriptional activity in these cells. It is possible that the
genomic regions covered by nucleotides +35 to +75 or
+125 to +167 could be involved in guiding the remaining
transcription [26] as well as two antioxidant-responsive
elements recently shown to be functional [36]. The
promoter-driven regulation of TXNRD1 transcription cer-
tainly needs to be studied further in diverse cellular
contexts and including distant regulatory elements. We
can conclude, however, that, as shown by the analysis of
cDNA variants reported herein, the core promoter has
proven to be the most common regulator of transcription
of TXNRD1, as the clear majority of the TXNRD1-derived
transcripts initiated at this site.
We were surprised by the extensive alternative
splicing and/or alternative transcriptional start sites
of human TXNRD1 that we identified in this study.
Notably, two intervening sequences in the TXNRD1
gene are nearly identical to a large number of cDNA
clones that seem not to be related to TrxR1. In
between the core promoter and exon h-I, a section
of the gene encodes a sequence identical to that of
ribosomal protein L18A (UniGene cluster Hs.414074).
However, that protein is transcribed from chromosome
19 and this sequence in the TXNRD1 gene is likely to
be an insertion of a pseudogene. A second set of
sequences relates to the two ‘‘exons’’ located between
exons gII and III of TXNRD1 (see Fig. 3) and found
in two mRNA and 33 EST clones as collected in
UniGene cluster Hs.313413. Those transcripts were
derived from a range of tissues and encode a hypo-
thetical protein of unknown function. As we, however,
could not find any TXNRD1-specific sequences con-
taining both or parts of these hypothetical exons, their
relation to TXNRD1 transcription, if any, remains
obscure.
The multitude of TXNRD1 transcripts with different
5V ends probably reflects alternative post-transcription-
al regulation as well as regulation by cell- or growth
condition–specific switches to transcription from al-
ternative promoters. A differential splicing could pos-
sibly yield mRNA with yet unidentified regulatory
elements in the 5V-UTR, which could be tissue-specif-
ic or responding to various cellular signals, in addi-
tion to generating the variant TrxR1 proteins with

A. -K. RUNDLOF et al.654
different N-terminal domains identified herein. More-
over, in addition to transcripts differing in the 5V end,those with different 3V ends might also be expressed.
This seemed to be the case with the a13 transcript
encoding TXNRD1_v5, and other variations at the 3V-UTR of additional transcripts may possibly exist (but
were not studied here). Interestingly, transcript a13
encoding TXNRD1_v5 was derived from testis and
trachea. Since the thioredoxin-like 2 protein was only
found in testis and cilia of lung airways [37], this
could possibly indicate a functional link between
these rare members of the human thioredoxin system.
The thioredoxin system seems to be particularly
complex in testis with many recently identified iso-
enzymes of thioredoxin [37–41]. The fact that the htranscripts of TXNRD1 found in this work were only
present in testis-derived material could possibly relate
to one or several testis-specific thioredoxins.
The regulation and function of the different TXNRD1
variants must be the subject of future detailed studies.
Furthermore, it should be noted that alternative splicing
also has been reported for the human mitochondrial
isoenzyme TrxR2 [20] and for TrxR from Drosophila
[31]. The sequences of some EST clones indicate that
the human testis-specific TGR may display alternative
first exons [31]. Interestingly, for TGR of Echinococcus
granulosus both cis- and trans-splicing events, which have
not yet been seen in the human transcripts, have been
reported [42]. For the different splicing sites identified in
this study, most conformed to splice site consensuses with
slight deviations (Table 2). These deviations may indicate
the presence of sequence-determined regulation of splic-
ing, the detailed molecular mechanisms of which are as of
yet unclear [43]. Generally, alternative splicing is known
to be an important mechanism for controlling gene ex-
pression and is a common event in higher organisms [44].
Although the specific mechanisms that control use of
different splice sites are still relatively undefined, regula-
tory elements for alternative splicing have been identified
for several human gene products, and these elements may
be localized within the splice site consensus sequences of
exons or introns or much further up- or downstream in the
pre-mRNA [43]. Alternative splicing may also be con-
trolled by the concentration and activity of splicing
regulatory proteins, the activities of which in turn can be
regulated by signal transduction events [45]. Another level
of regulated splicing is controlled via tissue-specific
splicing factors, as has been shown for several testis-
specific splicing events [46]. Such factors are probably
also involved in the regulation of TXNRD1, since the htranscripts found in this work were only present in testis-
derived material. Clearly, both the mechanisms generating
the many alternative transcripts of TXNRD1 as well as
their diverse functions must be the subject of extensive
further studies.
In conclusion, we have shown that the core promoter
guides transcription of the majority of transcripts derived
from the TXNRD1 gene. Also, we have identified and
categorized a wide range of transcript variants and there-
by revealed an exceedingly complex picture of TXNRD1
function. We have proposed a nomenclature for the
organization of the different exons and transcripts. Final-
ly, we believe that the findings presented here can serve as
the basis for more detailed studies of TXNRD1 tissue-,
cell-, and growth condition–specific expression.
Acknowledgments—This study was supported by grants from TheSwedish Society for Medical Research (to A.-K. R.), the SwedishMedical Society (to E. S. J. A.), The Karolinska Institute (to E. S. J. A.and A. M.-V.), the Swedish Cancer Society (Projects 3775 and 4056 toE. S. J. A.), the Swedish Medical Research Council (Projects 03P-14096-01A and 03X-14041-01A to A. M.-V.), and the Ake WibergsStiftelse (A. M.-V.).
REFERENCES
[1] Arner, E. S. J.; Holmgren, A. Physiological functions of thiore-doxin and thioredoxin reductase. Eur. J. Biochem. 267:6102–6109; 2000.
[2] Nordberg, J.; Arner, E. S. J. Reactive oxygen species, antioxi-dants, and the mammalian thioredoxin system. Free Radic. Biol.Med. 31:1287–1312; 2001.
[3] Tamura, T.; Stadtman, T. C. A new selenoprotein from humanlung adenocarcinoma cells: purification, properties, and thiore-doxin reductase activity. Proc. Natl. Acad. Sci. USA 93:1006–1011; 1996.
[4] Gladyshev, V. N.; Jeang, K.-T.; Stadtman, T. C. Selenocysteine,identified as the penultimate C-terminal residue in human T-cellthioredoxin reductase, corresponds to TGA in the human placen-tal gene. Proc. Natl. Acad. Sci. USA 93:6146–6151; 1996.
[5] Zhong, L.; Arner, E. S. J.; Ljung, J.; Aslund, F.; Holmgren, A.Rat and calf thioredoxin reductase are homologous to gluta-thione reductase with a carboxyl-terminal elongation containinga conserved catalytically active penultimate selenocysteine resi-due. J. Biol. Chem. 273:8581–8591; 1998.
[6] Zhong, L.; Holmgren, A. Essential role of selenium in the cata-lytic activities of mammalian thioredoxin reductase revealed bycharacterization of recombinant enzymes with selenocysteine mu-tations. J. Biol. Chem. 275:18121–18128; 2000.
[7] Zhong, L.; Arner, E. S. J.; Holmgren, A. Structure and mechanismof mammalian thioredoxin reductase: the active site is a redox-active selenolthiol/selenenylsulfide formed from the conservedcysteine-selenocysteine sequence. Proc. Natl. Acad. Sci. USA97:5854–5859; 2000.
[8] Holmgren, A. Bovine thioredoxin system. Purification of thiore-doxin reductase from calf liver and thymus and studies of itsfunction in disulfide reduction. J. Biol. Chem. 252:4600–4606;1977.
[9] Arner, E. S. J.; Nordberg, J.; Holmgren, A. Efficient reduction oflipoamide and lipoic acid by mammalian thioredoxin reductase.Biochem. Biophys. Res. Commun. 225:268–274; 1996.
[10] Kumar, S.; Bjornstedt, M.; Holmgren, A. Selenite is a substratefor calf thymus thioredoxin reductase and thioredoxin and elicits alarge non-stoichiometric oxidation of NADPH in the presence ofoxygen. Eur. J. Biochem. 207:435–439; 1992.
[11] Holmgren, A.; Lyckeborg, C. Enzymatic reduction of alloxan by

5Vvariants of TXNRD1 transcripts 655
thioredoxin and NADPH-thioredoxin reductase. Proc. Natl. Acad.Sci. USA 77:5149–5152; 1980.
[12] Bjornstedt, M.; Hamberg, M.; Kumar, S.; Xue, J.; Holmgren, A.Human thioredoxin reductase directly reduces lipid hydroperox-ides by NADPH and selenocystine strongly stimulates the reac-tion via catalytically generated selenols. J. Biol. Chem. 270:11761–11764; 1995.
[13] Sun, Q. A.; Wu, Y.; Zappacosta, F.; Jeang, K. T.; Lee, B. J.; Hat-field, D. L.; Gladyshev, V. N. Redox regulation of cell signaling byselenocysteine in mammalian thioredoxin reductases. J. Biol.Chem. 274:24522–24530; 1999.
[14] Miranda-Vizuete, A.; Damdimopoulos, A. E.; Pedrajas, J. R.;Gustafsson, J. A.; Spyrou, G. Human mitochondrial thioredoxinreductase cDNA cloning, expression, and genomic organization.Eur. J. Biochem. 261:405–412; 1999.
[15] Lee, S. R.; Kim, J. R.; Kwon, K. S.; Yoon, H. W.; Levine, R. L.;Ginsburg, A.; Rhee, S. G. Molecular cloning and characterizationof a mitochondrial selenocysteine-containing thioredoxin reduc-tase from rat liver. J. Biol. Chem. 274:4722–4734; 1999.
[16] Sun, Q. A.; Kirnarsky, L.; Sherman, S.; Gladyshev, V. N.Selenoprotein oxidoreductase with specificity for thioredoxinand glutathione systems. Proc. Natl. Acad. Sci. USA 98:3673–3678; 2001.
[17] Fujiwara, N.; Fujii, T.; Fujii, J.; Taniguchi, N. Functional expres-sion of rat thioredoxin reductase: selenocysteine insertion se-quence element is essential for the active enzyme. Biochem. J.340:439–444; 1999.
[18] Gasdaska, J. R.; Harney, J. W.; Gasdaska, P. Y.; Powis, G.;Berry, M. J. Regulation of human thioredoxin reductase expres-sion and activity by 3V-untranslated region selenocysteine inser-tion sequence and mRNA instability elements. J. Biol. Chem.274:25379–25385; 1999.
[19] Kawai, H.; Ota, T.; Suzuki, F.; Tatsuka, M. Molecular cloning ofmouse thioredoxin reductases. Gene 242:321–330; 2000.
[20] Lescure, A.; Gautheret, D.; Carbon, P.; Krol, A. Novel selenopro-teins identified in silico and in vivo by using a conserved RNAstructural motif. J. Biol. Chem. 274:38147–38154; 1999.
[21] Koishi, R.; Kawashima, I.; Yoshimura, C.; Sugawara, M.; Seriza-wa, N. Cloning and characterization of a novel oxidoreductaseKDRF from a human bone marrow–derived stromal cell lineKM-102. J. Biol. Chem. 272:2570–2577; 1997.
[22] Chen, C. Y.; Shyu, A. B. AU-rich elements: characterization andimportance in mRNA degradation. Trends Biochem. Sci. 20:465–470; 1995.
[23] Rozell, B.; Hansson, H. A.; Luthman, M.; Holmgren, A. Immu-nohistochemical localization of thioredoxin and thioredoxin re-ductase in adult rats. Eur. J. Cell Biol. 38:79–86; 1985.
[24] Gasdaska, J. R.; Gasdaska, P. Y.; Gallegos, A.; Powis, G. Humanthioredoxin reductase gene localization to chromosomal position12q23-q24.1 and mRNA distribution in human tissue. Genomics37:257–259; 1996.
[25] Berggren, M.; Gallegos, A.; Gasdaska, J. R.; Gasdaska, P. Y.;Warneke, J.; Powis, G. Thioredoxin and thioredoxin reductasegene expression in human tumors and cell lines, and the ef-fects of serum stimulation and hypoxia. Anticancer Res. 16:3459–3466; 1996.
[26] Rundlof, A.-K.; Carlsten, M.; Arner, E. S. J. The core promoter ofhuman thioredoxin reductase 1: cloning, transcriptional activity,and Oct1, Sp1, and Sp3 binding reveal a housekeeping-type pro-moter for the AU-rich element-regulated gene. J. Biol. Chem.376:30542–30551; 2001.
[27] Swick, A. G.; Blake, M. C.; Kahn, J. W.; Azizkhan, J. C. Func-tional analysis of GC element binding and transcription in thehamster dihydrofolate reductase gene promoter. Nucleic AcidsRes. 17:9291–9304; 1989.
[28] Jensen, D. E.; Black, A. R.; Swick, A. G.; Azizkhan, J. C.Distinct roles for Sp1 and E2F sites in the growth/cell cycleregulation of the DHFR promoter. J. Cell. Biochem. 67:24–31;1997.
[29] Ryan, A. K.; Rosenfeld, M. G. POU domain family values: flex-
ibility, partnerships, and developmental codes. Genes Dev.11:1207–1225; 1997.
[30] Rundlof, A. K.; Carlsten, M.; Giacobini, M. M.; Arner, E. S. J.Prominent expression of the selenoprotein thioredoxin reductasein the medullary rays of the rat kidney and thioredoxin reductasemRNA variants differing at the 5Vuntranslated region. Biochem. J.347:661–668; 2000.
[31] Sun, Q. A.; Zappacosta, F.; Factor, V. M.; Wirth, P. J.; Hatfield,D. L.; Gladyshev, V. N. Heterogeneity within animal thioredoxinreductases. Evidence for alternative first exon splicing. J. Biol.Chem. 276:3106–3114; 2001.
[32] Osborne, S. A.; Tonissen, K. F. Genomic organisation and alter-native splicing of mouse and human thioredoxin reductase 1genes. BMC Genomics 2:10; 2001.
[33] Wein, H. M.; Bruford, E. A.; Lovering, R. C.; Lush, M. J.; Wright,M. W.; Povey, S. Guidelines for human gene nomenclature. Ge-nomics 79:464–470; 2002.
[34] Luthman, M.; Holmgren, A. Rat liver thioredoxin and thioredoxinreductase: purification and characterization. Biochemistry 21:6628–6633; 1982.
[35] Oblong, J. E.; Gasdaska, P. Y.; Sherrill, K.; Powis, G. Purificationof human thioredoxin reductase: properties and characterizationby absorption and circular dichroism spectroscopy. Biochemistry32:7271–7277; 1993.
[36] Hintze, K. J.; Wald, K. A.; Zeng, H.; Jeffery, E. H.; Finley, J.W. Thioredoxin reductase in human hepatoma cells is transcrip-tionally regulated by sulforaphane and other electrophiles viaan antioxidant response element. J. Nutr. 133:2721–2727;2003.
[37] Sadek, C. M.; Jimenez, A.; Damdimopoulos, A. E.; Kieselbach,M.; Nord, M.; Gustafsson, J. A.; Spyrou, G.; Davis, E. C.; Oko,R.; van der Hoorn, F. A.; Miranda-Vizuete, A. Characterization ofhuman thioredoxin-like 2. A novel microtubule-binding thiore-doxin expressed predominantly in the cilia of lung airway epithe-lium and spermatid manchette and axoneme. J. Biol. Chem.278:13133–13142; 2003.
[38] Miranda-Vizuete, A.; Ljung, J.; Damdimopoulos, A. E.; Gustafs-son, J. A.; Oko, R.; Pelto-Huikko, M.; Spyrou, G. Characteriza-tion of Sptrx, a novel member of the thioredoxin familyspecifically expressed in human spermatozoa. J. Biol. Chem.276:31567–31574; 2001.
[39] Miranda-Vizuete, A.; Tsang, K.; Yu, Y.; Jimenez, A.; Pelto-Huik-ko, M.; Flickinger, C. J.; Sutovsky, P.; Oko, R. Cloning anddevelopmental analysis of murid spermatid-specific thioredoxin-2 (Sptrx-2), a novel sperm fibrous sheath protein and autoantigen.J. Biol. Chem. 278(45):44874–44885; 2003.
[40] Sadek, C. M.; Damdimopoulos, A. E.; Pelto-Huikko, M.; Gustafs-son, J. A.; Spyrou, G.; Miranda-Vizuete, A. Sptrx-2, a fusionprotein composed of one thioredoxin and three tandemly repeatedNDP-kinase domains, is expressed in human testis germ cells.Genes Cells 6:1077–1090; 2001.
[41] Yu, Y.; Oko, R.; Miranda-Vizuete, A. Developmental expressionof spermatid-specific thioredoxin-1 protein: transient associationto the longitudinal columns of the fibrous sheath during sperm tailformation. Biol. Reprod. 67:1546–1554; 2002.
[42] Agorio, A.; Chalar, C.; Cardozo, S.; Salinas, G. AlternativemRNAs arising from trans-splicing code for mitochondrialand cytosolic variants of Echinococcus granulosus thioredox-in glutathione reductase. J. Biol. Chem. 278:12920–12928;2003.
[43] Ladd, A. N.; Cooper, T. A. Finding signals that regulate alterna-tive splicing in the post-genomic era. Genome Biol. 3:re-views0008; 2002.
[44] Black, D. L. Mechanisms of alternative pre-messenger RNA splic-ing. Annu. Rev. Biochem. 72:291–336; 2003.
[45] Stamm, S. Signals and their transduction pathways regulatingalternative splicing: a new dimension of the human genome.Hum. Mol. Genet. 11:2409–2416; 2002.
[46] Venables, J. P. Alternative splicing in the testes. Curr. Opin. Gen-et. Dev. 12:615–619; 2002.

A. -K. RUNDLOF et al.656
ABBREVIATIONS
AURE—AU-rich elements
DTNB—dithionitrobenzoic acid
EST—expressed sequence tag
ORF—open reading frame
POU—Pit-Oct-Unc
RACE—rapid amplification of cDNA ends
ROS—reactive oxygen species
SECIS—selenocysteine incorporation sequence
Trx— thioredoxin
TrxR—thioredoxin reductase
UTR—untranslated region





























