Embed Size (px)
Citation preview

ELSEVIER
LifeSciewes,Vol.62, No.5,pp.39%412, W98CopyrightQ 1997EkvierScienceInc.PrintedintheUSA.Allrights-d
C024-3205/98$19.CO+.(XIPII SO024-3205(97)OIKBS
THIOREDOXIN FROM STREPTOMYCES AUREOFACIENS CONTROLSCOILING OF PLASMID DNA
Olga Golubnitchaya-Labudova*, Tana Horecka**, Marko Kapalla**, Dusan Perecko**,Eva Kutejova**, Gert Lubec*
*) University of Vienna, Dpt of Pediatrics ●’) Institute of Molecular Biology, SlovakAcademy of Sciences, Bratislava,
(Receivedin finalformNovemberl% 1997)
Summary
A number of potential functions of thioredoxin have been proposed inliterature, including a role for DNA replication. The aim of our study was toinvestigate the effects of thioredoxin from Streptomyces aureofaciens (TrxS.a.) on plasmid DNA. Trx S.a. was incubated with plasmid forms and theincubation product(s) characterized on agarose gels. To compare Trxactivity with enzymes with known DNA modifying activities, topoisomerase1,II (gyrase) and T4 DNA Iigase were incubated with plasmid DNA inparallel. For the demonstration of nick removal aPCR technique was used.Trx S.a. bound non-specifically to plasmid DNA relaxing supercooledcircleclosed form (CCC form) with subsequent formation of the circle closedform (CC form) as a major product. The amplification of a specific DNAtemplate, possible only after nick removal, took place following incubationwith Trx. The effect of topoisomerase I on plasmid DNA resembled Trx S.a.activity. We propose the foliowing mechanism for CCC relaxation: Bindingof Trx leads to a break of one strand and CC is formed by stepwiserelaxation, ending with nick removal. The concomitant finding of opencircle form (OC form) generation after incubation with Trx may indicate thegeneration of an intermediate due to the postulated strand break atinitiation. This control of coiling may play a role in the DNA replicationmachinery, providing CC as a readily available substrate for DNApolymerases. In addition, Trx may serve in DNA repair mechanisms by itsnonspecific binding to DNA and nick removing activity.
KeyW&: IhioredoxinjSbeptomyes aunofaciens, coilingj plasmid-DNAjnick-removingactivity
Correspondence:Prof.Dr.GerlLubec,CChem,FRSC(UK),UniversityofVienna,WaehringerGuertel18,A1090Vienna,Austria,fax:x43.I.404003238

39s ThioredoxinControlsPlasmidDNA Coiling Vol. 64 No. 5, 199S
Thioredoxin (TRX) is mainly known as serving in biological redox systems and wasoriginally defined as a hydrogen donor for ribonucleotide reductase (l). This findingpointed to an important role of thioredoxin for DNA synthesis, moreover, the inhibition ofDNA synthesis was predicted to be caused by a reduction in the deoxyribonucleotidepools. Muller (2) clearly showed that deleting both TRX1 and TRX2 genes in yeast didnot change levels of ITP, dCTP, dATP and dGTP but reduced the rate of DNAreplication. Although this observation can be easily explained by earlier work showingthat other enzyme systems can replace ribonucleotide reductase reduction activity ofTRX (3), TRX must play another role in DNA synthesis/replication: thioredoxin deficiencyin yeast prolongs S phase and shortens the G1 interval of the cell cycle (4) suggestingthat in the absence of TRX1 and TRX2 a slow DNA replication rate inhibits the normalprogress of cellular reproduction. Muller (2) proposed two models, one that proposesthat thioredoxin is required for the reduction of a subset of ribonucleotide reductasemolecules that are part of a complex channeling nucleotides directly to the replicationmachinery. The second proposes that an unknown component, the “replicationcomponent activated by thioredoxin”, RAT, of this machinery is activated by thioredoxin.The latter model is supported by findings of Hartman et al. (5) who described thatinjection of heterologous thioredoxin into fertilized Xenopus eggs blocks DNA synthesisin the presence of abundant deoxyribonucleotide and from Salz et al. (6) describingthe disturbed DNA synthesis in P-element disruption of Drosophila deadhead locus,which encodes thioredoxin.
Important information about the involvement of thioredoxin in DNA replication camefrom previous work showing that thioredoxin was a processivity factor for the gene 5protein (7-9): E. coli thioredoxin binds tightly and stoechiometrically to T7 polymerase;thioredoxin in its reduced form stabilizes the binding of gene 5 protein to a primertemplate by up to 80 fold thus increasing the processivity of polymerization by 1000 fold(10,11). Recent studies (12-15) have revealed another major function of thioredoxin inthe DNA replication system: the fidelity of thioredoxin controls coiling of plasmid DNA.DNA synthesis can be controlled by thioredoxin-thioredoxin either enhances or reducesfidelity depending on the type of error considered.
We provide evidence for an additional mechanism participating in the DNA replicationmachinery: in streptomyces thioredoxin is removing nicks, protects dsDNA againstDNAse 1,and induces DNA relaxation suggesting a role for replication.
Methods
Materials
Chemicals-Electrophoretic agarose was from Sigma; restriction endonucleases, PNK,PCR kit were from Promega; oligonucleotides were synthesized by MWG (Germany);HEPES, acrylamide, bisacrylamide, TEMED, NADPH, DTNB were from Serva(Germany); Sephactyl S- 300, Phenyl-Sepharose 4B, Mono Q HR 5/5 were fromPharmacia (Sweden). All other chemicals were purchased from common commercialsources. Streptomyces aureofaciencs 3239 (10762) and E.coli DH 5 alpha wereobtained from ATCC. Media-Niedercorn medium (16) was used for the cultivation ofstreptomyces aureofaciens. LB medium and LB medium solidified with Difco agar andsupplemented with ampicillin (100 rig/ml) according to Sambrook et al. (17).Topoisomerase 1 was from Gibco BRL (38042-024), DNA Gyrase was from Gibco

Vol. 62 No. 5, 1998 ThioredoxinControlsPlasmidDNA Coiling 399
(8043SB). T4 DNA Iigase was from Boehringer Mannheim (481 220). The RandomPrimer Fluorescein Labeling kit was from Dupont (NEL 803), the Plasmid purification kitwas from Quiagen (QUI 676) an PUC18was purchased from Boehringer ( 885 797).
Cultivation of Streptomyces aureofaciens (S.a.)
The first generation of cells was grown at 28°C for 24 hrs on a rotary shaker in 500 mlflasks containing 100 ml of Niedercorn medium. 5 ml of cells from the first generationwere used to seed 100 ml of medium 16 (0.3’Yosaccharose, 1.50/.dextrine, 0.01YO urea,0.05 Y. NaCl and 0.65 Y. K2HPOd.3H20)in a 500 ml flask. After 20 hrs growth on a rotaryshaker at 28°C cells were harvested by centrifugation at 2000 g for 15min. The pelletwas washed once with 0.9 Y. saline. Washed cells were stored frozen at -20°C until use.
Isolation and characterization of thioredoxin (Trx) from S.a.
All operations were performed at O-4”C. 30 g of the cell pellet were suspended inapprox. 10 ml of buffer A (0.05 M Tris HCI, 0.002 M EDTA, pH 8.0) and the cellsuspension was sonicated seven times for 30 sec with one minute breaks kept in an icebath.The suspension was then centrifuged at 20.000 g for 20 min (all followingcentriguations were performed under the same conditions). 16 ml of 5’Yo streptomycinsulfate were added to 100 ml of the cell free extract. The suspension was stirred for 30min and the sediment was removed by centrifugation while the supernatant was heated2 min at 65°C. After rapid cooling insoluble proteins were removed from the supernatantby centrifugation and solid ammonium sulfate was added to 40?/. saturation underpermanent stirring. A sediment was removed after six hrs by centrifugation. Thesaturation of ammonium sulfate was brought to 60°/0, a sediment after 12 hrs wascollected by centrifugation and dissolved in 2 ml of buffer B (buffer A complemented withethylene glycol to ten percent) and applied to a 80 x 1.5 cm Sephacryi S-300 columnequilibrated with buffer B. Fractions containing thioredoxin activity (see enzyme assay)were pooled and dialyzed twice against buffer A and then against buffer C (buffer Acomplemented with ammonium sulfate up to 30Y0 saturation). The dialyzate wassubsequently applied to a 4.5 x 2.3 cm Phenyl-Sepharose column equilibrated withbuffer C, Trx was eluted with 100 ml of a linear gradient of 30-0 Y. saturated ammoniumsulfate and of 0-507. ethylene glycol in buffer A. Fractions containing Trx-activity werepooled and dialyzed twice against buffer D (0.025 M Tris.HCl, 0.002 M EDTA, pH 8.5)and then against buffer E (buffer D complemented with 10Yoethylene glycol). Fractionswith Trx activity were applied onto a Mono Q HR 5/5 column equilibrated with buffer E.The column was washed with 5 ml of buffer E. Trx was eluted with 40 ml of a lineargradient of 0.4 M NaCl in buffer E. The purification was monitored by a Trr assaymeasuring Trx activity and on 12Y0SDS-PAGE (18).
Assay for thioredoxin activity
Trx specific activity was determined using the insulin reduction assay of Holmgren (19)at a DIT concentration of 330 micromolar. After the last purification step Trx S.a. had aspecific activity of 5.0 A650/min/mg.
DNA modification by Trxs
DNA modification by Trxs was tested using pUC18 plasmid DNA as a template. DNAforms isolated from E.coli were separated on agarose gels, the individual bands cut out

4X) l’hioredoxinControls PlasmidDNA Coiling Vol, 62 No. 5, 199S
and characterized on agarose gels using DNA modification enzymes restrictionendonuclease Eco Rl, topoisomerase 1,II (gyrase) and T4 DNA Iigase (17),
a) 1 pg of plasmid DNA each was reacted with different concentrations (2-20 ng) of S.a.Trx in 50 mM Tris HCI buffer pH 8.0 containing lmM EDTA (TE), with and without DIT(under reducirig or non-reducing conditions) or ATP (excess of lmM, in order to showwhether binding activity was ATP requiring) for three hours at 30°C. The mixtures wererun on 17. agarose gels with subsequent ethidium bromide staining.
b) 10 ng of plasmid DNA were reacted with Trx, 2 fractions obtained during the final stepof purification of S.a. Trx (figure 3c), topoisomerase 1,II (gyrase), T4 DNA Iigase andDNAse 1,with and without DIT in the buffer described above for one hour at 37°C. Themixtures were run on IYo agarose gels with subsequent capillary blotting to nylonmembranes (Hybond N+, Dupont). Plasmid DNA was detected by hybridization withfluorescein labeled (Random Primer Fluorescein Labeling kit, Dupont, NEL 803)denaturated linearized pUC18 (by incubation of the CCC with Eco RI under optimalconditions) and chemiluminescence detection using the ReflectionTM AutoradiographyFilm (NEF 480, Dupont).
Rationale for the use of DNA modifying enzymes
Plasmid DNA exists in different forms: CCC (supercooled circle closed form, highestelectrophoretic mobility), OC (open or nicked circle form, slowest electrophoreticmobility), linear form (ds break, open form, intermediate electrophoretic mobility),different CC forms (circle closed form presenting with different turns leading to differentelectrophoretic mobility). In order to characterize the effect of Trx S.a. on the individualpiasmid forms we used a well-characterized and defined spectrum of DNA modifyingenzymes.
Eco RI induces nick formation in a stepwise manner acting primarily on on the OC withsubsequent generation of the linear form when incubated with the CCC form. Undernonoptimal conditions the process of linearization is only partial (production of OC andlinear forms). Under optimal conditions the linear form is produced exclusively,Topoisomerase I (DNA relaxing enzyme) relaxes both, negative and positivesuperhelical turns in covalently closed DNA by transient breakage and rejoining ofphosphate diester bonds. Incubation of this enzyme with the CCC form produces OCand a relaxed CC form. Topoisomerase II (gyrase, DNA supercooling enzyme)introduces negative superhelical turns into covalently closed DNA by double strandedbreakage and rejoining of the phosphodiester bonds. As a result supercooled closedcircular duplex DNA and relaxed positively supercooled DNA is formed. T4 DNA Iigasecloses both, single and double strand breaks. Incubation of Iigase with OC or linear formproduces CC form.
Preparation of different plasmid forms
Positively superhelical CCC form was prepared by isolation of total plasmid DNAfollowed by separation of the most electromobile CCC form from the gel (figure la)using the method of Zhen and Swank (20). OC and linear forms used for the incubationstudies were prepared by the incubation of the CCC form with Eco RI under nonoptimalconditions (figure lb). The most relaxed CC form was prepared by the incubation of OCform with T4 DNA Iigase (17).

Vol. 62 No. 5, 1998
Occcccc
....
..-.
..-.
ThioredoxinControlsPlasmidDNA Coiling
1234567
401
Fig. la. Plasmid DNA preparation: 1%incubated with topoisomerase Il. (lane 1-3);plasmid purification kit (lane 4-7)
1 2
agarose gel with. pUCl 8pUC18 isolated by Quiagen
Fig. lb. The preparation of different forms of pUC 18 (1% agarose gel):1. CCC form incubated with *ECORI under non-optimal conditions givingrise to OC and linear forms. 2. CCC form modified with topoisomerase Igiving rise to OC and CC forms.
PCR experiments
For PCR verification of DNA nick removing activity of S.a. Trx a specially prepareddsDNA template as shown in figure 2 was used. The underlying principle is theexperimental formation of a nick between a dsDNA fragment and linkers. The

402 ThioredoxinControlsPlasmidDNA Coiling vol. 62, IWO.5, 1998
polymerization of complemented strands by PCR can take place only if linkers arecovalently connected with the DNA fragment, i.e. in the absence of nicks or when nicksare removed. The recombinant plasmid used for the preparation of dsDNA template inour study was described in a previous paper (21). For the preparation of the dsDNAtemplate an about 500 bp fragment was cut out from pUTrxBM12 by Droll withrecognition of the GACN4 1“N2GTC sequence as shown in figure 2.
0 puTrxBM12
I digestionwithDnlI.
+3 -S00 bpDNAfra@entbolateflfmm agamwegel
1 complemenla(tonofone 3’-overlappingend
1nick+wmn%rg
‘+-”-41DenatwdDNA template+ PCRprimerUI
e 111
*
I polymerszdionfmmprimer [E
1+ o
I PC’Rampli]cation
~. -320 bp p~ptillt?t
Fig. 2. Template preparation for PCR. Determination of nick-removingactivity of Trx S.a. Full circles - 5’ phosphoylated ends, empty circles - 5’-nonphosphorylated ends.

Vol. 6%No. 5, 199S TbioredoxinControlsPlasmidDNA Coiling 403
The arising 3’ overlapping ends of the fragment were fully complemented on one endwith linkers I and II of the following oligonucleotide sequences:
I 5’ GGCGCGACGGTCAAGGTGACGAACGCGITC
II 3’ CCGCTCTGCCAGITCCACTGCITGCGCA
The 5’ends of primers I and II remained dephosphorylated whereas the 5’ end of the500 bp fragment was phosphorylated enabling ligation. As a result of this constellation(figure 2) a nick is being formed between the DNA fragment (hybrid molecule) andlinkers I / II (template, substrate used for nick removing activities).
This template (lOng each) was incubated with 100 ng of Trx S.a. at 30°C in 50 mM TrisHCI buffer pH 7.5 containing 2 mM EDTA for a period of one hour, with the rationale thatnick removing activity of Trx would result into PCR amplification of the template. After theincubation the reaction mixture was heated for 5 min at 95°C, cooled down to roomtemperature and primer Ill
Ill 5’ GAGCGACGCGTCCAGGAAGGCCGCGAGGCG
was added to anneal it to DNA template. Polymerization from primer Ill was performed at72°C for 30 seconds by Taq polymerase prior to PCR. 10 fg aliquots of the templatewere used for PCR with primers I and Ill, which was performed using the followingprogram: 94°C, 45 seconds were used for template denaturation, 65”C, 45 seconds forprimers-template annealing, 72°C, 30 sec for polymerization. 30 cycles were run (500pM of each primer, 200 UM dNTPs, 10 mM Tris HCI pH 8.8 at 25°C, 1.5 mM MgC12,50mM KCI, 0.1Y. Triton X -100 and 2.5 units of Taq polymerase in a total VOIof 50 vI), andPCR products were analyzed on a 1.5 9!0 agarose gel. All primers used weresynthesized by MWG, Germany.
Protein determinations were carried out with the commercially available kit ProteinAssayR (BIO-RAD).
Denistometry of films was performed using the Hirschmann elscript 400 densitometer(Germany).
Results and Discussion
Isolation of streptomyces aureofaciens thioredoxin
Peaks eluted from the ion exchange column (figure 3a-c) were separated andcharacterized on SDS-PAGE (figure 3d). A protein showing a Mr of approx. 10.000revealed thioredoxin activity in the specific insulin reduction assay given above.
DNA modification by thioredoxin and DNA modifying enzymes
As shown in figure 4 neither incubation of the plasmid DNA with ATP (lane 4) nor DIT(lane 3) changed the electrophoretic pattern of the plasmid DNA. The addition of TrxS.a. to the plasmid DNA without incubation failed to change the pattern as well (lane 5).Fraction 21 containing active Trx S.a. incubated with plasmid DNA showed a decrease

404 ThioredoxinControlsPlasmidDNA Coiling Vol. 62, No. 5, 199S
of CCC form with subsequent increase of CC and OC forms (lane 6). TM S,a. in thepresence of D1l’ revealed a decrease of CCC form along with a major CC product andthe OC form (lane 7). Incubation of Trx S.a. with plasmid DNA in the presence of ATPrevealed a decrease of CCC form along with a major CC form, a prominent OC form anda minor linear form (lane 8).
480
2
1.5
1
0.5
0
250uiml
200
150
100
50
01 21 41 61 81 n
FractionNumber
Fig. 3a. Elution profile of Trx S.a. on Sephacryl S-300 (80x1.5 cm), flowrate 10 ml/h, 1.8 ml fractions collected. Protein (-); activity (.);
0.1 ,,-”
h. u/ml,..,..
,.-O.m --s0 ““ 6,.-’
.,”,.-’ f+
J;
/,0.0s--w .-
----- .-” \-J’,..------ ,/- .n-------------- ,-”,.-
ao4 --43 -----><::.,.- ----,.- ----- p+,.. --...L#,.-
/
------,.-O.a?-a \‘--------,---
,.- --b....-.,., -----,.,” -------..-
o~1 21 41 sl 81 n
FmcUOM~
Fig. 3b. Elution profile of Trx S.a. on Phenyl Sepharose 4B (4.5x2.3 cm),flow rate 10 ml/h, 1.0 ml fractions collected. Protein (-); activity (.);decreasing gradient of ammonium sulfate (-.-); increasing gradient ofethylene glycol (---).

Vol. 62, No. 5, 1998 ThioredoxinControlsPlasmidDNA Coiling
IUWJIKXNp UO!JEJWWU03w to m
~ q q o0 0 0 0 0 0
~-----l-~
‘\ /’/’\ \.., .—.—...-A-—---4—~zu~+==-.---—~-—- ——:—~-=..—.~—.—.—, ———.\ .:
‘\‘\
\‘\
\.,, ,.
--.?,”—————,—.—.—---------”==----,-.-,-—--+.= i
—--.--.4>
\\
.-J,,../X’”
,---.—..—..——-—-——..———---—-—————__ <“—...—..—.—.—., ,, -...-.....-... U..,-.......J..--....—
No0
0

406 ThkwdcminControlsPlasmidDNA Coiling Vol. 62, No. 5, 1998
Fig. 3d. SDS-PAGE of active Trx S.a. after purification on Mono Q HR 5/5(lane 4); protein markers from top 17200, 14660, 8240, 6380 (lane 1);Trx from spirulina sp. (lane 2); Trx from E.coli (lane 3 ).
Figure 4. Changes in the electrophoretic pattern of different plasmid DNAforms after incubation with Trx S.a. Results electrophoretic analysis in 1%agarose gel, 1- linear form of pUC 18 digested by Eco Rl; 2- pUC 18incubated in TE at 300C for 3 hrs; 3- pUC 18 in TE after incubation withlmM DTT at 30°C for 3 hrs; 4- pUC 18 in TE after incubation with 1mMATP at 30°C for 3 hrs; 5- pUC 18 in TE, lmM ATP and 1 microgram TrxS.a. without incubation; 6- pUC 18 in TE and 1 microgram Trx S.a. afterincubation at 30°C for 3 hrs; 7 - pUC 18 in TE, lmM DIT and 1microgram Trx S.a. after incubation at 30°C for 3 hrs; 8- pUC 18 in TE,lmM ATP and 1 microgram of Trx S.a. after incubation at 30°C for 3 hrs,
Figure 5 demonstrates the results from DNA modifications by Trx and individual DNAmodifying enzymes.
Figure 5a :The first lane shows the modifications of CCC form following the incubationwith Eco RI under nonoptimal conditions: all described forms were shown in adensitometry pattern of OC:CC:linear:CCC = 491:109:57:308. A single linear form isshown after the incubation of CCC with Eco RI under optimal conditions (lane 3). Lane5 shows the pattern of all forms of plasmid DNA after the incubation with DIT formingthe basis for the comparison with incubation products with the densitometry pattern

Vol. 62, No. 5, 1998 ThioredoxinControlsPlasmidDNA Coiling 4Q7
OC:CC:linear:CCC =291:100:86:208. In lane 6 (Trx inactive) fraction 20 from Trx S.a.purification and in lane 7 (Trx inactive) fraction 22 after incubation with all present formsis shown and an unchanged pattern as compared to lane 5 was found.
Trx S.a. incubated with all present forms of plasmid DNA (lane 8) showed the reductionof the linear form along with an increased CC- and OC form, verified by thedensitometrical pattern of OC:CC:linear:CCC=663:267:33:l 77. In lane 2 the patternfollowing the incubation of CCC form with topoisomerase I is given. The CCC-,OC- asmajor bands and a band representing a CC form are demonstrated (Densitometricalpattern OC:CC:linear:OC= 441:213:21:180). In lane 4 a major band representing theisolated OC form and a minor (lower) band standing for the CC form.
In lane 1 of figure 5b the incubation product of plasmid DNA with Trx S.a. (also shown infig.5a, lane 8) pattern is given and compared to the pattern resulting from DNAse Idigestion of the Trx.S.a. - plasmid DNA complex: the Trx S.a.- plasmid DNA -complexwas comparable to lanes 8 in figure 5a and lane 1 of 5b indicating a protective effect ofTrx S.a. against DNase I cleavage.
Figure 5c: The effect of DNAse I treatment (of all plasmid forms ) in the absence of activeTrx S.a. (fractions 20 in lane 13 and fraction 22 in lane 14 ) consisted of total digestionof all plasmid DNA forms.
OC and CC forms prepared as given above and shown in figure 5a lane 4 wereincubated with T4 DNA Iigase resulting into an increase of a CC form (lane 2) in contrastto incubation of OC and CC with inactive fractions 20 (lane 1) and fraction 22 (lane 3). Inlanes 4-6 incubation of OC and CC with Trx S.a. (no DTT - DTT - with following DNAseI digestion ) showed an increase of CC and protection against DNAse I digestion by TtxS.a. In the absence of Trx S.a. OC and CC forms were degraded by DNase I (lane 12).Topoisomerase II (gyrase) activity showed relaxing activity as well, turning CCC formsinto CC but unlike for topoisomerase 11,no negative supercooling by Trx S.a. wasobserved (data not shown).
PCR - Experiments
The amplification of specially prepared DNA template after incubation with TRX resultedin the formation of a 320 bp fragment, whereas this fragment was not amplified withoutpreincubation of the template with Trx S.a. (figure 6, lane 2). This fragment was also notamplified in the absence of complementation between the DNA fragment with the linker1/11(lane 4). These two negative controls confirm that the amplification of the target 320bp fragment was indeed possible after nick-removal by the incubation with Trx S.a.(lane 3 ) or T4 DNA Iigase (lane 5).
The involvement of TRX in the processes of DNA repair as well as its multifunctional rolein replication has been proposed previously (22). No experimental data supporting thishypothesis, however, have been reported so far. As shown in the Results we found thatTrx S.a. bound to all forms of plasmid DNA independent of reducing agent or ATP byelectrophoretic patterns and by protection against DNase 1, with the biochemicalconsequence of relaxing the supercooled closed circular form, i.e. CCC and nickremoval.

40s ThioredoxinControlsPlasmidDNA Coiling Vol. 62, No. 5, 1998
a b c
Fig. 5. Changes of individual forms of pUC 18 after the incubation withDNA modifying enzymes and Trxs. Electrophoretic analysis in 1YOagarose gel (all incubations performed at 37°C for 1 hr).
5a) see densitometry in the Results section1- pUC 18 treated with *ECORI under nonoptimal conditions; 2- CCCform treated with topoisomerase 1;3 - linear form of plasmid PUC 18digested with Eco RI under optimal conditions; 4- Separated OC and CCforms ;5 - all plasmid forms in Trx buffer with lmM DIT; 6- all plasmidforms incubated with a TM S.a. non-active fraction 20 (figure 3c); 7- allplasmid forms incubated with a Trx S.a. non-active fraction 22 (figure 3c);8- all plasmid forms incubated with 1 microgram of Trx Sa.
5b) 1- all plasmid forms incubated with 1 microgram of Trx S.a.; 2- aslane 1 but followed by the incubation with 10 micrograms of DNAse 1;
5c) 1- OC and CC forms incubated with the (Trx inactive) fraction 20; 2- OC and CC forms incubated with T4 DNA Iigase showing an increase ofthe CC form; 3- OC and CC forms incubated with the (Trx inactive )fraction 22; 4- OC and CC forms incubated with isolated Trx S.a.; 5- as 4but with the addition of lmM DIT; 6 -as 4 but followed by the incubationwith DNAse I indicating protection against clevage; 7-11 not discussed12- as 27 followed by incubation with DNAse 1;13- all plasmid forrrsincubated with (Trx inactive) fraction 20 and subsequent incubation withDNAse 1;14- all plasmid forms incubated with the (Trx inactive ) fraction22 with subsequent incubation with DNAse 1.

Vol. 63 No. 5,1998 ThioredoxinControlsPlasmidDNA Coiling 409
1 2345
320bp
Fig. 6. Analysis of PCR products in 1.5 % agarose gel: 1 - DNA standard(Boehringer Mannheim, No Vi); 2- Negative control of PCR: the templatewas prepared as described in figure 3 and used for PCR withoutincubation with Trx; 3 - PCR performed with 10 fg aliquot of a Trx -modified template after the incubation of 1 ng of the template (figure 3)with 1 microgram of Trx S.a. at 30”C for 1 h; 4- Negative control of PCR:the template was prepared as described in figure 3 but without thecomdementation of the 3’ - overlapping end of the DNA fragment withlinker MI; 5 - Positive control of PCR:-the template wasdescribed in figure 3 and used in the PCR after incubationpolymerase at 370C for 1 h.
pr&pared aswith T4 DNA
Trx binding to plasmid DNA led to the relaxation of circular plasmid forms: Incubation incontrast to simply addition of Trx to plasmid DNA revealed a decrease of the tense CCCform represented by a single band on agarose electrophoresis on the expense of theCC forms with one major prominent band.
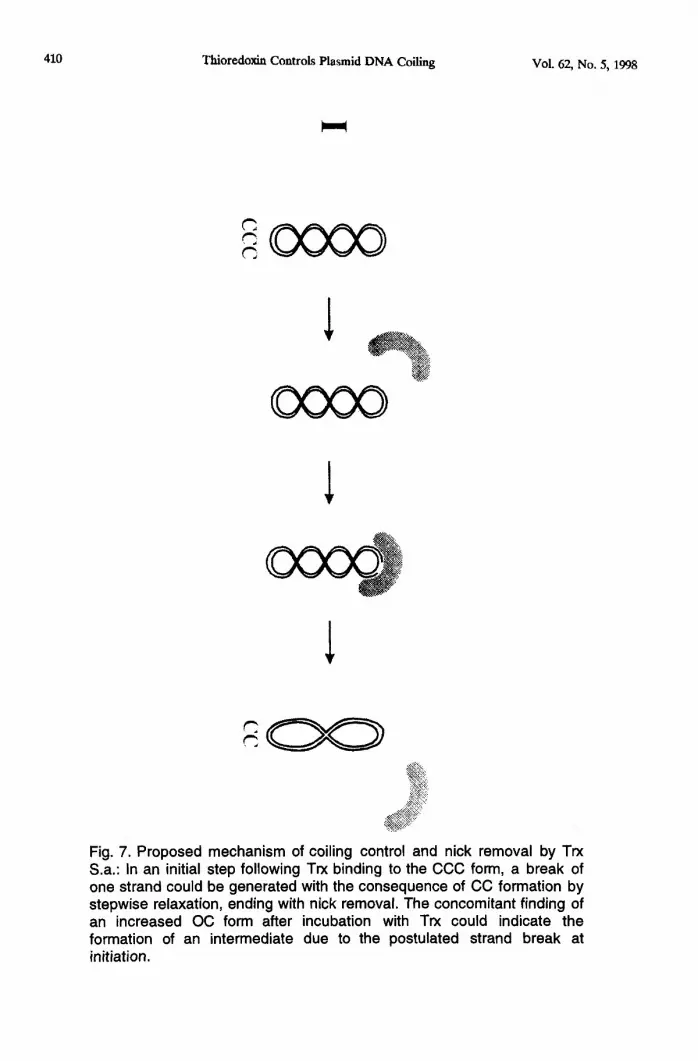
The proposed mechanism, shown in figure 7, could be the following: In a first stepfollowing Trx binding to the CCC form, a break of one strand could be produced with theconsequence of CC formation by stepwise relaxation ending with nick removal. Theconcomitant finding of an increased OC form after incubation with Trx could indicate theformation of an intermediate due to the postulated strand break at initiation.
To test the hypothesis on the proposed mechanism of action, we performed parallelstudies using DNA modifying enzymes with well - defined effects on DNA.Topoisomerase I relaxed the CCC form resulting into CC formation with OC as anintermediate thus resembling Trx S.a. activity. Topoisomerase II showed relaxing activityas well, turning CCC forms into CC, topoisomerase II activity was, however, to bediscriminated from Trx s.a. activity as no negative supercooling by Trx S,a. wasobserved.
410 ThioredoxinControlsPlasmidDNA Coiling
. ..,..,>.>*,:},e:.+ &
$%’
1
1
vol.62,No. 5, 1998
Fig. 7. Proposed mechanism of coiling control and nick removal by TrxS.a.: In an initial step following Trx binding to the CCC form, a break ofone strand could be generated with the consequence of CC formation bystepwise relaxation, ending with nick removal. The concomitant finding ofan increased OC form after incubation with Trx could indicate theformation of an intermediate due to the postulated strand break atinitiation.
—

Vol. 62 No. 5, 1998 ThioredoxinControlsPlasmidDNA Coiling 411
T4 DNA Iigase activity was shown by the incubation of OC with T4 DNA Iigaseproducing CC form. Trx S.a. did not show Iigase activity as revealed on agarose gelelectrophoresis when incubated with OC. Using the PCR design for the assay of nickremoval, i.e. Iigase activity, Trx S.a. showed this effect. This phenomenon would becompatible with the findings on electrophoresis (figure 5) that Trx S.a. significantlyreduced the linear form reflecting Iigase activity. Trx activities as e.g. topoisomerase Iand Iigase activity could be detected for Trx from other species as E.coli and spirulina aswell (Data not shown). It has been shown that Trx is an essential subunit of the T7polymerase complex (23) but no specific role has been assigned to Trx. Based upon ourobservation on relaxation activity on the CCC form we suggest a biological role of Trxfor replication; the relaxation activity may provide CC forms, forming a readily availablesubstrate for DNA polymerases. It is known from literature that topoisomerase Igenerates the CC form as the readily available substrate for DNA replication (24). Thisfinding would also explain the more effective processivity of replication in the presenceof Trx (25).
Another role for Trx in the replication process could be proposed by the nick removingactivity during synthesis of daughter strand from mother lagging strand.
Apart from the proposed multifunctional role of Trx in replication, the possibleinvolvement of Trx in DNA repair can be suggested, particularly as it could be repairingall kinds of breaks due to Trx’ s nonspecific DNA binding. In addition, it was shownpreviously that Trx either enhances or reduces fidelity of DNA synthesis depending onthe type of error considered (25).
1.2.3.4.5.
6.
7.8.
9.
10.
11.
12.13.14.15.
16.
References
P. REICHARD, Ann Rev Biochem. 57349-374 (1988)E.G.D. MULLER, J Biol Chem. 26924466-24471 (1994)M.RUSSEL, P. MODL and A. HOLMGREN, J Bacteriol. 1721923-1929 (1990)E.G.D. MULLER, J Biol Chem. 2669194-9202 (1991)H.M. HARTMAN, M. WU, B.B. BUCHANANand J.C. GERHART, Proc Natl AcadSci (USA). 902271-2275 (1993).H.K. SALZ, T.W. FLICKINGER,E.Mlll_ENDORF, A.PELICENA-PALLE,J.P.PETSCHEK and E.B. ALBRECHT, Genetics. 1361075-1086 (1994)P. MODRICH and C.C. RICHARDSON, J Biol Chem. 2505508-5522 (1975)D.F. MARK and C.C. RICHARDSON, Proc Natl Acad Sci (USA). 73780-784(1976)J.S. HIMAWAN and C.C. RICHARDSON, Proc Natl Acad Sci (USA). 899774-9778 (1992)S.TABOR, H.E. HUBER and C.C. RICHARDSON, J Biol Chem. 26216212-16223(1987)H.E. HUBER, S. TABOR and C.C. RICHARDSON, J Biol Chem. 26216224-16232 (1987)S.S.PATEL, 1.WONG and K.A. JOHNSON, Biochemistry. 30511-525 (1991)I.WONG, S.S. PATEL and K.A. JOHNSON, Biochemistry 30,526-537 (1991)M.J.DONLIN, S.S.PATEL and K.A.JOHNSON, Biochemistry 30538-546 (1991)T.A.KUNKEL, S.S. PATEL and K.A. JOHNSON, Proc Natl Acad Sci (USA) 916830-6834 (1994)J.G. NIEDERCORN, U.S. Patent 2609,329 (1952)

412 ThioredoxinControlsPlasmidDNA Coiling Vol. 62 No. 5, 199S
17. J.SAMBROOK, E.F.FRITSCH and T.A. MANIATIS, Mo/ecu/arC/oning:ALaboratory Marrua/, 2nd Ed., Cold Spring Harbor Laboratory, Cold SpringHarbor, NY (1989)
18. U.K.LAEMMLI, Nature. 227680-682 (1970 )19. A. HOLMGREN, J. Biol. Chem. 254 9627-9635 (1979).20. L. ZHEN and R.K. SWANK, BioTechniques. 14894-898 (1993)21. O.LABUDOVA, M. NEMETHOVA, J. TURNA and M. KOLLAROVA. Gene 13863-
264 (1994)22. C.A. LUNN and V.P. PIGIET, Int J Radiat Biol. 5129-38 (1987)23. Z. DEBYSER, S. TABOR and C.C. RICHARDSON, Cell 77157-166 (1994)24. J.D. RAWN, Biochemistry, lnt.Edition, Neil Patterson Publishers, Burlington, NC
(1989)25. T.A.KUNKEL, S.S. PATEL and K.A. JOHNSON, Proc Natl Acad Sci (USA). 91
6830-6834 (1994).
——





























