Embed Size (px)
Citation preview

This article was downloaded by: [Artem Sinev]On: 07 December 2012, At: 02:09Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Extremalona timmsi gen. nov., sp.nov., a new cladoceran (Cladocera:Anomopoda: Chydoridae) from anacid saline lake in southwest WesternAustraliaArtem Y. Sinev a & Russell J. Shiel ba Biological Faculty, M.V. Lomonosov Moscow State University,Moscow, Russiab Ecology, Evolution and Landscape Sciences, BenhamLaboratories, University of Adelaide, AustraliaVersion of record first published: 07 Dec 2012.
To cite this article: Artem Y. Sinev & Russell J. Shiel (2012): Extremalona timmsi gen. nov., sp.nov., a new cladoceran (Cladocera: Anomopoda: Chydoridae) from an acid saline lake in southwestWestern Australia, Journal of Natural History, 46:45-46, 2845-2864
To link to this article: http://dx.doi.org/10.1080/00222933.2012.727215
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Journal of Natural HistoryVol. 46, Nos. 45–46, December 2012, 2845–2864
Extremalona timmsi gen. nov., sp. nov., a new cladoceran (Cladocera:Anomopoda: Chydoridae) from an acid saline lake in southwestWestern Australia
Artem Y. Sineva* and Russell J. Shielb
aBiological Faculty, M.V. Lomonosov Moscow State University, Moscow, Russia; bEcology,Evolution and Landscape Sciences, Benham Laboratories, University of Adelaide, Australia
(Received 8 February 2012; final version received 2 September 2012; printed 5 December 2012)
A new species of Cladocera, Extremalona timmsi gen. nov., sp. nov., was foundin acid saline lakes in the southwest of Western Australia. Extremalona gen. nov.belongs to the Coronatella-branch of Aloninae, but differs from all members ofthe group by the oval body with small high-set head (tip of rostrum located athalf-height of the body), the exopodite III with uniform, well-developed setae4–6 and male antennule with six lateral and nine terminal aesthetascs, and bynumerous other characters. In our opinion, Extremalona gen. nov. is one of theancestral genera of the Coronatella-branch of Aloninae, sharing numerous com-mon features with the elegans-group of Alona s.l. Our data confirm a high level ofendemism in Australian Chydoridae and Cladocera in general.
Keywords: Australia; cladocera; endemism; systematics; morphology
Introduction
The mesic southwest of Western Australia has long been recognized for its biodiversity,high levels of endemicity and conservation value (Halse et al. 2004). Much of the earlyresearch that led to the southwest being listed as one of the world’s 25 biodiversity“hotspots” was based on the extensive radiation of vascular plants (Myers et al. 2000).More recently, comparable endemism has begun to emerge for aquatic invertebrates,with a suite of endemic rotifers (Segers and Shiel 2003), copepods (Halse and McRae2001), cladocerans (Frey 1991b, 1998; Van Damme et al. 2007 a,b), ostracods (Halse2002) among others, described from the region.
The southwest is characterized by thousands of small saline lakes, often in theswales of old dunes, on carbonate-rich sands, i.e. alkaline, whereas those further inlandlie on granites or sandstones, and are subject to rising acidic groundwater (Timms2009). These “windows” on the water tables provide a heterogeneous range of habitatsin which the aquatic biota has evolved over millennia.
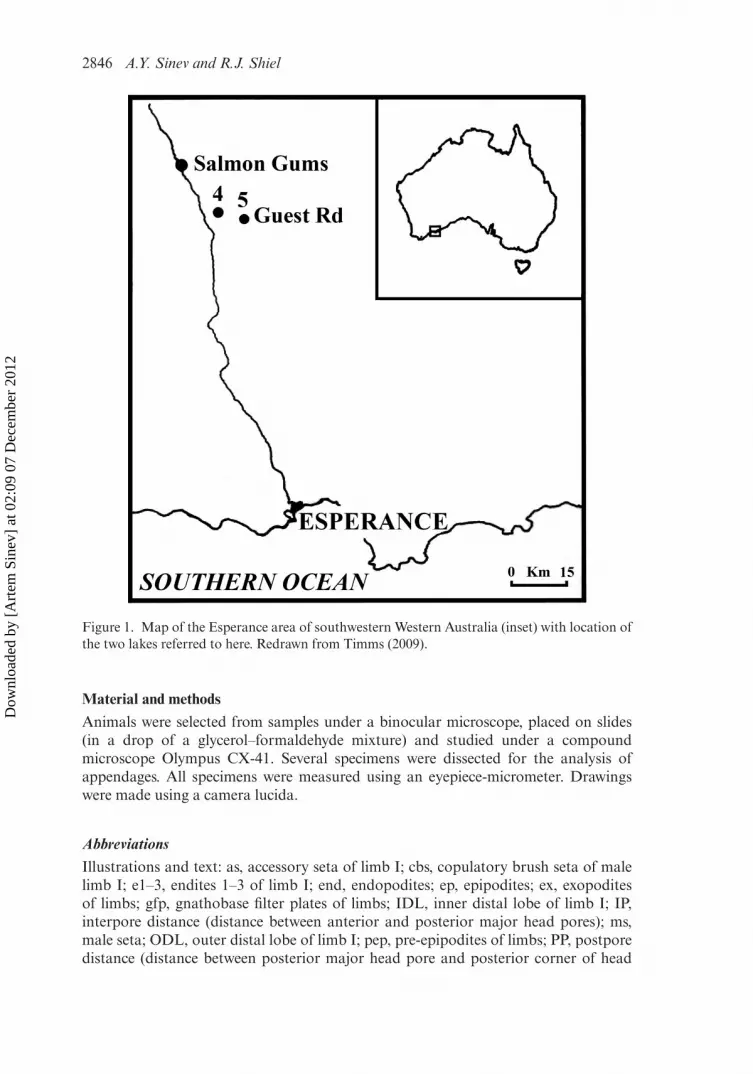
During a survey of more than 40 acid saline lakes in the Esperance region, Dr BrianTimms found two populations of alonine chydorids in adjacent lakes near SalmonGums, c.80 km north of Esperance (Figure 1), and forwarded individuals to us fortaxonomic determination. Collecting methods and site details from his study are givenin Timms (2009).
*Corresponding author. Email: [email protected]
ISSN 0022-2933 print/ISSN 1464-5262 online© 2012 Taylor & Francishttp://dx.doi.org/10.1080/00222933.2012.727215http://www.tandfonline.com
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012
2846 A.Y. Sinev and R.J. Shiel
Figure 1. Map of the Esperance area of southwestern Western Australia (inset) with location ofthe two lakes referred to here. Redrawn from Timms (2009).
Material and methods
Animals were selected from samples under a binocular microscope, placed on slides(in a drop of a glycerol–formaldehyde mixture) and studied under a compoundmicroscope Olympus CX-41. Several specimens were dissected for the analysis ofappendages. All specimens were measured using an eyepiece-micrometer. Drawingswere made using a camera lucida.
AbbreviationsIllustrations and text: as, accessory seta of limb I; cbs, copulatory brush seta of malelimb I; e1–3, endites 1–3 of limb I; end, endopodites; ep, epipodites; ex, exopoditesof limbs; gfp, gnathobase filter plates of limbs; IDL, inner distal lobe of limb I; IP,interpore distance (distance between anterior and posterior major head pores); ms,male seta; ODL, outer distal lobe of limb I; pep, pre-epipodites of limbs; PP, postporedistance (distance between posterior major head pore and posterior corner of head
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2847
shield); s, sensillum. Numbering of limb setules is given from epipodite to gnathobase,without any implication of homology.
Results
The studied cladoceran belongs to a new species, which does not fit any of the existinggenera of Chydoridae, so a new genus, with a single species, is described.
Family CHYDORIDAE Dybowski and Grochowski, 1894 emended Dumont andSilva-Briano, 1998
Subfamily ALONINAE Dybowski and Grochowski, 1894 emended Frey, 1967Tribe ALONINI Dybowski and Grochowski, 1894 emended Kotov, 2000
Extremalona gen. nov.
Type species Extremalona timmsi sp. nov.
Etymology
The name reflects the ability of the genus to inhabit an extreme habitat for Cladocera –a highly acidic, saline lake.
Diagnosis
Female. Aloninae of moderate size. Body oval, valves and head shield without keel.Posteroventral angle low, being the posteriormost extremity of the body. Dorsal andventral margins of valves evenly curved, posterodorsal, and anterioventral angles ofvalves broadly rounded. Posteroventral angle of valves without denticles, with about25 small setules not organized into groups.
Head is set relatively high for Aloninae, tip of rostrum located at half height ofthe body. In lateral view head small, low triangular-round, rostrum short, directed for-ward and downward. Eye usually larger than ocellus. Head shield narrow, with short,broadly rounded rostrum, posterior margin of head shield broadly rounded. Threenarrowly connected major head pores of the same size. Lateral head pores elongated.Labrum with narrow oval keel, keel apex rounded, posterior margin of keel withoutclusters of setae.
Thorax twice as long as abdomen. Abdomen without abdominal joint.Postabdomen relatively short, slightly curved, narrowing distally, with broadlyrounded dorsodistal angle. Distal margin almost straight. Dorsal margin with dis-tal part about 1.3–1.5 times longer than preanal one, with postanal portion half thelength of the anal portion. Postanal portion of distal margin weakly convex, analportion concave. Preanal angle well-defined, prominent, postanal angle not defined.Preanal margin slightly concave. Provided with 10–12 clusters of marginal setules, evenin preanal portion these clusters are similar to lateral groups of setules. Lateral fasciclesof setules numerous, spaced irregularly; main row consists of 9–11 broad fascicles. Firstsetules in postanal fascicles thicker than others. Postabdominal claw curved, abouttwo-thirds length of preanal portion of postabdomen. Basal spine thin, about 0.3 timeslength of claw.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2848 A.Y. Sinev and R.J. Shiel
Antennule almost reaching the tip of rostrum. Antennular sensory seta slender,about two-thirds length of antennule, arising at two-thirds distance from the base.Nine terminal aesthetascs, three of them longer than others, as long as antennule.Antenna with antennal setae formula 0-0-3/1-1-3, spines 1-0-1/0-0-1. Basal seg-ment robust, branches short and stout. Seta of the basal endopodite segment thin,as long as endopodite itself. Seta of the middle endopodite segment and apicalsetae of same morphology. Spine on basal segment of exopodite about two-thirdslength of middle segment. Spines of apical segments significantly shorter than thesesegments.
Five pairs of thoracic limbs. Epipodites of all limbs indistinctly separated from themain body of the limb.
Limb I with accessory seta four times shorter than ODL seta. IDL with three setae,seta 3 longest, about two-thirds length of ODL seta, seta 1 quite large, about half-length of seta 2, setae 2 and 3 with thin setules in distal part. Endite 3 with four setae.Endite 2 with three setae. Endite 1 with two two-segmented setae, and a long setadirected to the base of the limb. No naked setae on anterior face of endites 1–2. Sixrows of long setules on ventral face of limb. Two ejector hooks, one a little larger thanthe other.
Limb II: exopodite narrow, elongated, with very small seta. Inner portion of limbwith eight scraping spines, scrapers 5 and 6 bear larger denticles than others. Smallnaked seta located near the base of scraper 1, small sensillum between bases of scrapers3 and 4. Distal armature of gnathobase with four elements. Filter plate II with sevensetae, the posteriormost seta considerably shorter than others.
Limb III: exopodite six setae, seta 3 being longest, setae 4–6 long, of similar sizeand shape, armed with thick setules in distal part. Distal endite with three setae,two distalmost members long, slender, sharp, with long denticles; basalmost setashorter, geniculate, with long setules. Basal endite with four setae. Four inner setaeincreasing in size basally. Distal armature of gnathobase with four elements –elongated, cylindrical sensillum, thin, geniculated seta, and two short acute spines.Filter plate III with seven setae.
Limb IV: exopodite broad oval, with six setae decreasing in size basally, setae5–6 plumose as others. Inner lobe of limb IV with four setae and ovoid sensillum.Scraping seta short and broad, flaming torch setae of similar morphology, decreas-ing in size basally. Three inner setae increasing in size basally. Gnathobase with shorttwo-segmented seta and a blunt process near it. Filter plate IV with five setae.
Limb V: exopodite oval, not separated into two lobes, with four plumose setaedecreasing in size basally. Inner limb portion as elongated lobe. Setae of inner face ofsame size. Filter plate as a single seta.
Male. Body low oval, maximum height before the middle of the body. Ocellus muchsmaller than eye. Postabdomen short, with almost parallel margins in distal portion,dorsodistal angle broadly rounded. Preanal angle not defined, postanal angle well-defined. Distal part of postabdomens 1.2–1.3 times longer than preanal. Sperm ductopenings located almost at the end of postabdomen. Armament of postabdomen sim-ilar to that of female. Postabdominal claw short, about two-thirds of preanal marginlength, basal spine same size as in female. Antennule much broader than in female,with nine terminal and six lateral aesthetascs. Male seta arising at one-third antennalength from tip, exceeding one-third of antennule length. Limb I with U-shapedcopulatory hook. Row of about 40 long, dense setules on ventral face of limb under
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2849
copulatory brush. IDL retains seta 1, setae 2 and 3 much thinner and shorter than infemale, male seta thick, curved, as long as seta 2.
Differential diagnosis
Extremalona gen. nov. belongs to the Coronatella-branch of Alona-like Aloninae(genera Coronatella Dybowski and Grochowski, 1894, Karualona Dumont and Silva-Briano, 2000, Leberis Smirnov, 1989, Celsinotum Frey, 1991, Anthalona Van Damme,Sinev and Dumont, 2011, and elegans-group of Alona s.l.), sharing several main diag-nostic features of the group, including six setae on exopodite III, reduction of filterplate on limb VI and reduction of limb VI; these features clearly separate the genusfrom genera of the Hexalona-branch of Aloninae. Extremalona gen. nov. differs fromall members of the group by the oval body with small high-set head (tip of rostrumlocated at half-height of the body), the exopodite III with uniform, well-developedsetae 4–6 and male antennule with six lateral and nine terminal aesthetascs. Also,Extremalona gen. nov. clearly differs from Coronatella, Leberis, Celsinotum, Anthalonaand elegans-group by the presence of well-developed IDL seta 1; from Anthalona andKarualona by three major head pores; from Leberis and Celsinotum by the roundedrostrum and long apical spines of antenna. Other differences between these groups areseparated in Table 1.
Extremalona timmsi sp. nov.(Figures 2–5)
Type locality
A lake at Guest Road, Grass Patch, 79 km north of Esperance, Western Australia,Australia 33◦06′ S, 121◦44′ E (Figure 1). Samples collected by Brian Timms on12 March 2007.
Holotype
Parthenogenetic female from the type location, deposited in Western AustralianMuseum, slide C47938.
Paratypes
Ephippial female, male and ephippium from the type location, deposited in WesternAustralian Museum, slide C47938; parthenogenetic female, three ephippial females,two males, juvenile male of instar II from the type locality, presently in the first author’spersonal collection, AYS-AU-011.
Several ephippial females from the type location, two males and one juvenile maleof instar II were dissected and not kept afterward.
Etymology
The species name honours the species collector, the prominent Australian limnologistBrian Timms.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2850 A.Y. Sinev and R.J. Shiel
Tab
le1.
Diff
eren
ces
betw
een
gene
raof
Cor
onat
ella
-bra
nch
wit
hap
omor
phie
ssh
own
inbo
ldty
pe.
Ext
rem
alon
aE
lega
ns-g
roup
Leb
eris
,C
elsi
notu
mC
oron
atel
laK
arua
lona
Ant
halo
na
Max
imum
leng
thof
fem
ale
0.57
mm
0.5–
06m
m0.
53–1
.1m
m0.
25–0
.45
mm
,0.
6m
min
one
spec
ies
0.4–
0.45
mm
0.3–
0.4
mm
Bod
ysh
ape
Ova
lwit
hhi
gh-s
ethe
adO
valw
ith
low
-set
head
Rou
nded
Ova
lwit
hlo
w-s
ethe
adO
valw
ith
low
-set
head
Ova
lwit
hlo
w-s
ethe
adPo
ster
oven
tral
corn
erof
valv
esW
itho
utde
ntic
les
Wit
hout
dent
icle
sW
itho
utde
ntic
les
Wit
hout
dent
icle
sW
ith
dent
icle
sin
mos
tspe
cies
Wit
hout
dent
icle
s
Bod
yke
elA
bsen
tA
bsen
tP
rese
ntA
bsen
tA
bsen
tA
bsen
tR
ostr
umR
ound
edR
ound
edT
runc
ated
Rou
nded
Rou
nded
Rou
nded
Mai
nhe
adpo
res
Thr
eeT
hree
Thr
eeT
hree
Tw
oT
wo
Lat
eral
head
pore
sM
inut
eM
inut
eM
inut
eM
inut
eM
inut
eS
peci
aliz
ed,w
ith
cosm
aria
Seta
onba
sal
segm
ent
ofan
tenn
aen
dopo
dite
Lon
gL
ong
Lon
gS
hort
inm
ost
spec
ies
Sho
rtS
hort
Spin
eson
apic
alse
gmen
tsof
ante
nna
Lar
geL
arge
Sm
all
Lar
geL
arge
Lar
ge
Setu
les
ofID
Lse
tae
2an
d3
Thi
nT
hin
Thi
ck,b
utno
ttr
ansf
orm
edin
tode
ntic
les
Thi
ck,f
requ
entl
ytr
ansf
orm
edin
tode
ntic
les
Thi
nT
hick
,fre
quen
tly
tran
sfor
med
into
dent
icle
sF
irst
IDL
seta
Lar
geR
udim
enta
ryR
udim
enta
ryR
udim
enta
ryor
abse
ntO
fm
oder
ate
size
Abs
ent (C
onti
nued
)
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2851
Tab
le1.
(Con
tinu
ed).
Ext
rem
alon
aE
lega
ns-g
roup
Leb
eris
,C
elsi
notu
mC
oron
atel
laK
arua
lona
Ant
halo
na
Inne
rse
taof
limb
IIP
rese
ntP
rese
ntP
rese
tA
bsen
tA
bsen
tP
rese
nt,a
tlea
stin
som
esp
ecie
sE
xopo
dite
IIse
taV
ery
smal
lV
ery
smal
lA
bsen
tL
arge
Abs
ent
Abs
ent
Seta
e4–
6of
exop
odit
eII
IO
fsi
mila
rsi
zean
dar
mam
ent
Dif
fere
ntia
ted
Dif
fere
ntia
ted
Dif
fere
ntia
ted
Dif
fere
ntia
ted
Dif
fere
ntia
ted
Filt
erpl
ate
V1
seta
1se
ta1(
2?)
seta
e0–
1se
tae
Abs
ent
Abs
ent
Lat
eral
aest
heta
scs
onm
ale
ante
nnul
e
62
2in
Leb
eris
,2–
6in
Cel
sino
tum
Abs
ent
Abs
ent
Abs
ent
IDL
seta
1on
mal
elim
bI
Pre
sent
Abs
ent
Abs
ent
Abs
ent
Abs
ent
Abs
ent
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2852 A.Y. Sinev and R.J. Shiel
Diagnosis
As for genus.
Description
Parthenogenetic female. Preserved specimens of intense yellow-brown colour. In lat-eral view (Figure 2A), body irregular oval, moderately high, with maximum lengthat posterodorsal angle. Maximum height at the middle of the body in adults, height/length ratio about 0.7. Dorsal margin convex, posterodorsal and posteroventral anglesbroadly rounded. Posterior margin weakly curved. Anteroventral angle rounded.Sarapace covered by weakly defined longitudinal lines with dense fine striae betweenthem.
As only two parthenogenetic specimens were present in our material, detailed mor-phology was studied for ephippial females, which did not differ from parthenogeneticones in morphology of head pores, postabdomen and head and thoracic limbs in thefamily Chydoridae (Smirnov, 1971).
Ephippial female (Figures 2B, C, 3A, B) with body as high as in parthenogeneticfemale, dorsal outline highly arched, posterodorsal angle of valves obtuse, posteriormargin of valves straight, ventral margin of valves less convex than in parthenogeneticfemale. Ephippium very dark brown to black, non-transparent, without distinctivesculpture. Posterodorsal angle with about 20–25 short setules of similar size, pass-ing into a row of about 100 setules along the posterior margin (Figure 2D), at somedistance from the margin on inner side of carapace. Ventral margin almost straight,with about 50 short setae. Anteroventral angle rounded. Sarapace covered by weaklydefined longitudinal lines with dense fine striae between them.
Head is set relatively high for Aloninae, tip of rostrum located at half-height of thebody. In lateral view head small, low triangular-round, rostrum short, directed forwardand downward. Eye larger or equal to ocellus. Distance from tip of rostrum to ocellusabout 1.5 times that between ocellus and eye.
Head shield relatively narrow, with maximum width behind mandibular articula-tion, covered by same striae as valves (Figure 2E). Rostrum short, broadly rounded.Posterior part of head shield protruding, posterior margin of head shield broadlyrounded. Three narrowly connected major head pores of same size (Figures 2F, 3C)evenly spaced. PP about 1.0–1.2 times IP. Lateral head pores elongated, located about1.1–1.2 times IP distance from midline, at the level of the middle major head pore.
Labrum (Figure 2G–I) with elongated, narrow oval keel (height/width ratio about2.5), with rounded apex. Anterior margin of keel convex, posterior margin convexor straight without clusters of setules. Several specimens in the studied material hadabnormal labral keels (Figure 2J, K).
Thorax twice as long as abdomen, dorsal surface of abdominal segments notsaddle-shaped. No abdominal joint.
Postabdomen (Figures 3D, 4A, B) slightly curved, narrowing distally, with broadlyrounded dorsodistal angle. Length about 3–3.5 times height. Ventral margin weaklyconvex. Basis of claws bordered from distal margin by clear incision. Distal marginalmost straight. Dorsal margin with distal part about 1.3–1.5 times longer thanpreanal one, with postanal portion half length of anal portion. Postanal portionof distal margin weakly convex, anal portion concave. Preanal angle well defined,prominent, postanal angle not defined. Preanal margin slightly concave. Postabdomen
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2853
Figure 2. Extremalona timmsi sp. nov. from the type location. (A) Parthenogenetic female,holotype;, Western Australian Museum, slide C47938; (B–K) ephippial females (B, C paratypesfrom the personal collection of the first author, AYS-AU-011, other drawings from dissectedspecimens): (B, C) lateral view, (D) posteroventral angle of valves, (E) head shield, (F) headpores, (G–I) labrums, (J, K) labrums with abnormal keels; (L) juvenile male of instar II;(M) adult male, Western Australian Museum, slide C47938. Scale bars A–C, E, L–M, 0.1 mm;D–F and G–K, 0.05 mm.
with 10–12 clusters of marginal setules, decreasing in size basally, along preanaland anal margin, even in preanal portion these clusters are similar to lateral groupsof setules. In preanal portion each cluster consists of two to seven setules, lengthof longest setules in postanal clusters little less than width of base of postbdomi-nal claw. Lateral fascicles of setules numerous, spaced irregularly; main row consistsof 9–11 broad fascicles, additional smaller fascicles located above the main row inpostanal portion and basal part of anal portion. Several rows of setules in preanal por-tion of postabdomen close to preanal angle. First setules in postanal fascicles thickerthan others. Postabdominal claw curved, of moderate length, about two-thirds the
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2854 A.Y. Sinev and R.J. Shiel
Figure 3. Extremalona timmsi sp. nov. from the type location, paratypes from the personal col-lection of the first author, AYS-AU-011. (A–D) ephippial females: (A, B) lateral view, (C) headpores, (D) postabdomen; (E, F) adult male: (F) lateral view, (E) postabdomen.
length of preanal portion of postabdomen. Basal spine thin, about 0.3 times lengthof claw, a cluster of long setules located near its base.
Antennule (Figure 4C) comparatively large and broad, reaching the tip of rostrum,without clusters of setules at anterior face. Length/width ratio about 2. Antennularsensory seta slender, about two-thirds length of antennule, arising at two-thirds dis-tance from the base. Nine aesthetascs, three of them 1.3–1.4 times longer than others,as long as of antennule, others of similar size. All aesthetascs projecting beyondanterior margin of head shield.
Antenna short (Figure 4D). Antennal formula, setae 0-0-3/1-1-3, spines 1-0-1/0-0-1. Basal segment robust, branches short and stout. Basal segment of endopodite1.5 times and two times longer than middle and apical parts of same branch, respec-tively; basal segment of exopodite three times longer. Seta arising from basal segmentof endopodite thin, as long as endopodite itself. Seta arising from middle segment ofendopodite slightly smaller than apical setae. Apical setae of similar size and thickness.Spine on basal segment of exopodite about two-thirds length of middle segment. Spinefrom apical segment of exopodite about two-thirds length of this segment. Spine fromapical segment of exopodite about half length of this segment.
Thoracic limbs: five pairs. Epipodites of all limbs not clearly separated from themain body of the limb, without any projections.
Limb I (Figure 4E, F) of moderate size. ODL with exceptionally long seta, morethan three-quarters length of limb body. Accessory seta about one-quarter length of
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2855
Figure 4. Extremalona timmsi sp. nov. from the type location, drawings from dissected speci-mens. (A–F) ephippial females: (A, B) postabdomen, (C) antennule, (D) antenna, (E) thoraciclimb I, (F) inner and outer distal lobes of limb I; (G–J) juvenile male of instar II, slide in thefirst author’s personal collection: (G) postabdomen, (H) antennule, (I, J) thoracic limb I; (K–N)adult male: (K) postabdomen, (L) antennule, (M, N) thoracic limb I. Scale bar 0.05 mm.
ODL seta. IDL with three setae, seta 1 well-developed, sharp and slender, about one-third length of ODL seta. Setae 2 and 3 of similar shape, armed with thin setulesin distal part, about two-thirds and three-quarters length of ODL seta, respectively.Endite 3 with four setae of similar length. Endite 2 with three setae of different length,
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2856 A.Y. Sinev and R.J. Shiel
longest of them (seta e) as long as IDL seta 3, two others much shorter. Endite 1 withtwo distally setulated two-segmented setae (g, h) and a long flat seta (i). No nakedinner setae and sensillae found on endites 1 and 2. Six rows of thin long setules onventral face of limb. Two ejector hooks of similar size. Maxillar process not found.
Limb II (Figure 5A). Exopodite elongated, with minute seta; eight scraping spines(Figure 4J), evenly increasing in length distally, scrapers 5 and 6 armed with morerobust setules than others. A small inner seta is located near the base of scraper 1.Distal armature of gnathobase with four elements. Filter plate with seven setae, twoposteriormost considerably shorter than others.
Limb III. Exopodite (Figure 5B) of irregular shape, with six setae. Seta 3 beinglongest, armed with thin setules. Setae 4–6 of same morphology and similar size,plumose in distal part, evenly decreasing in size basally. Seta 4 about two-thirds lengthof seta 3, seta 6 about half length of seta 3. Seta 1 about one-quarter length of seta3, seta 2 very short. Distal endite (Figure 5C) with three setae and two small sensillaebetween their bases, two distalmost setae (1–2) slender, with distal parts unilaterally
Figure 5. Extremalona timmsi sp. nov. from the type location, thoracic limbs of ephippialfemales, drawings from dissected specimens: (A) limb II, (B) exopodite of limb III, (C, D) innerportion of limb III, (E) exopodite of limb IV, (F, G) inner portion of limb IV and its inner setae,(H) limb V. Scale bar 0.05 mm.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2857
armed with denticles; basalmost seta (3) with shortened distal portion, bilaterallyarmed with long setules. Basal endite with four setae, increasing in size in basal direc-tion. Gnathobase not clearly separated from basal endite. Four inner setae of similarsize (Figure 5D), a small sensillum near the base of distalmost seta. Distal armature ofgnathobase with four elements. The first one elongated, cylindrical sensillum, secondthin, bent seta, others two short spines. Filter plate III with seven setae.
Limb IV. Exopodite (Figure 5E) as broad oval lobe with six plumose setae. Seta1 very broad, longest, setae 2 and 3 about three-quarters length of seta 1, setae 5 and6 about two-thirds length of seta 1, seta 6 about half length of seta 1. Inner lobe of limbIV with four setae (Figure 5F, G). Distalmost scraping seta (1) short, with broad base,three flaming-torch setae (2–4) strongly decreasing in size basally. Sensillum elongated.Three inner setae increasing in size basally. Gnathobase with two-segmented seta, anda small hillock distally. Filter plate with five setae.
Limb V (Figure 5H). Exopodite low oval, not divided into two lobes, with fourplumose seta. Setae 2, 3 and 4 about two-thirds, one-half and one-quarter length ofseta 1, respectively. Inner limb portion an elongated lobe, length about twice width,with setulated inner margin. At inner face, two setae of same size, as long as exopoditeseta 3. Filter plate as a single seta, no sensillum-like structure near it.
Male. Juvenile males of instar II (Figure 2L) and adult males (Figures 2M, 3F) similarin shape and size, small in comparison with females. Body low oval, height/lengthratio about 0.6, maximum height before the middle of the body. Ocellus much smallerthan eye.
Postabdomen. In juvenile males of instar II (Figure 4G), lower than that of female,with almost parallel margins, with clear step on ventral margin in region of gonopores.Gonopores large, round, located close to ventral margin at three-quarters distancefrom its base. Armament of postabdomen similar to that of female, but setules inboth marginal clusters and lateral fascicles are less numerous. Postabdominal clawsame as in female. In adult male, postabdomen short, with almost parallel marginsin distal portion, dorsodistal angle broadly rounded (Figures 3G, 4K). Preanal anglenot defined, postanal angle well-defined. Distal part of postabdomens 1.2–1.3 timeslonger than preanal. Sperm duct openings located almost at the end of postabdomen.Armament of postabdomen similar to that of female, but setules in postanal clus-ters are somewhat shorter. Postabdominal claw shorter than that of female, abouttwo-thirds of preanal margin length, basal spine of same size as in female.
Antennule. In instar II male antennule slightly broader than in female, with anlageof male seta located opposite the base of antennal seta, aesthetascs same as in female(Figure 4H). In adult male antennule much broader than in female (Figure 4L), withnine terminal and six lateral aesthetascs as long as terminal ones. Male seta arising atone-third antenna length from tip, exceeding one-third of antennule length.
Thoracic limb I. In instar II male, copulatory hook curved (Figure 4I, J). Ventralface of limb with anlage of copulatory brush seta and a peculiar hillock above it,not present in any other instar. IDL with anlage of male seta, other setae same as infemale. In adult male, copulatory hook U-shaped (Figure 3M, N). Copulatory brushand copulatory brush seta present. Row of about 40 long, dense setules on ventralface of limb under copulatory brush, endite 3 with several clusters of very long setules.IDL seta 1 thinner than in previous instar, moved to the inner margin of IDL, setae
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2858 A.Y. Sinev and R.J. Shiel
2 and 3 subequal in length, thinner and shorter than in previous instar, male seta thick,curved, as long as seta 2. Endites with setae same as in female.
Size. In the studied population, length of two adult parthenogenetic females was0.49 and 0.51 mm, height 0.35 and 0.36 mm, respectively; in ephippial females length0.44–0.57 mm, height 0.29–0.40 mm; in single instar II male length 0.31 mm, height0.20 mm; in adult males length 0.33–0.34 mm, height 0.19–0.21 mm.
Ecology
Extremalona timmsi sp. nov. is known to date only from the type locality, B.V. Timms’field sites 4 [Guest Rd (west)] and 5 [Guest Rd (southeast)], without official name, butnamed from the nearest road, 33◦06′ S, 121◦44′ E and 33◦08′ S, 121◦49′ E, respectively(Timms, 2009). Guest Rd west was c.3.2 ha when full, 22 cm mean depth, c.50 cm max-imum, pH 3.7–5.8, salinity 11–95 g/l (on two occasions on which Extremalona gen.nov. was collected: 33–37 g/l), turbidity10–250 NTU. Guest Rd southeast was. 2.6 hawhen full, 49 cm mean depth, c.150 cm maximum, pH 3.7–7.8, salinity 4–182 g/l,turbidity10–130 NTU). Co-occuring acidophile microinvertebrates and macroinverte-brates are tabled in Timms (2009). Extremalona timmsi sp. nov. is the only species ofchydorid recorded from such an environment to date. Co-occurring cladocerans wereDaphnia carinata s.l. in site 5, and Moina cf. australiensis in both sites 4 and 5 (seetable 2 in Timms 2009).
Discussion
Why the new genus?
Extremalona timmsi sp. nov. shares numerous common features with the Coronatella-branch of Alona s.l., defined by Van Damme and Dumont (2008). This group, in itswidest interpretation, includes five genera: Coronatella (former rectangula-group ofAlona s.l.), Karualona, Leberis, Celsinotum and Anthalona, as well as the elegans-groupof Alona s.l. The latter group, presently consisting of five species, A. elegans Kurz, 1875,A. salina Alonso, 1996, A. orellanai Alonso, 1996, A. irinae Sinev, Alonso et Sheveleva,2009 and A. floessneri Sinev, Alonso et Sheveleva, 2009, also have generic rank (VanDamme and Dumont 2008; Sinev et al., 2009) but is not yet formally recognized asa genus. Extremalona gen. nov. shares most important features of the Coronatella-group – numerous reductions in trunk limb structures, including absence of limb VI,reduced filter plate V and six setae on exopodite III. Extremalona timmsi sp. nov.clearly did not fit any of the five established genera of Coronatella-group (see Dumontand Silva-Briano 2000; Sinev et al. 2005; Van Damme and Dumont 2008; Van Dammeet al. 2011; Sinev and Kotov 2012 for emended diagnoses of the genera).
In general appearance and morphology of appendages, Extremalona gen. nov.shares numerous similarities with the species of the elegans-group. They include: sim-ilar body shape and size; head shield with three connected main head pores; similararmament of postabdomen (one species of the elegans-group, Alona orellanai, evenhas a second row of lateral groups of setae); same morphology of antennula; similarmorphology of antenna, with well-developed seta on exopodite basal segment; samemorphology of setae on endites 1–3 of limb I; limb II with rudimentary exopodite setaand inner seta located near base of scraper 1; presence of lateral aesthetascs on male
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2859
antennule. Data on the elegans-group morphology are taken from Alonso (1996) andSinev et al. (2009).
Species of the elegans-group are distributed mostly in the arid belt of thePalaearctic, inhabiting mostly temporary and saline water bodies, and are not presentin large permanent freshwater lakes. One of the species, Alona salina, can toleratesalinities > 100 mg/l (Boronat et al. 2001), so E. timmsi sp. nov. is also close to theelegans-group ecologically.
However, differences between the elegans-group (the sixth would-be genus of theCoronatella-branch) and E. timmsi sp. nov. are too numerous to merge the species withthe group (for full list of differences see Table 2) and fully justify the description ofa new genus. Uniform morphology of trunk limbs is always observed in all recentlyestablished genera within Alona s.l. as well as in well-defined species groups, like affi-nis, costata, quadrangularis-group. Even in large assemblages, consisting of 10 or morespecies, like Coronatella, Anthalona or pulchella-group (see Van Damme and Dumont2008; Van Damme et al. 2011; Sinev et al. 2012) differences between species lie mostlyin armament and proportions of setae. In the elegans-group, limb morphology is veryuniform, species differ from each other mostly by proportions of exopodite setae (seeAlonso 1996; Sinev et al. 2009) But in Extremalona gen. nov., each limb is quite differ-ent from those of the elegans-group. Epipodites of the limbs in Extremalona gen. nov.are merged with the limb body, unlike in the elegans-group. Limb I of Extremalona gen.nov. bears well-developed IDL seta 1, this feature is especially important in Aloninaesystematics, both recently established genera and all species-groups uniformly haveeither well-developed or reduced IDL seta 1, and both states of this character are neverpresent together. In the elegans-group, this seta is rudimentary. Also, accessory seta oflimb I in Extremalona gen. nov. is much shorter than these in the elegans-group. LimbII of Extremalona gen. nov. have strongly differentiated scraping setae, whereas in allspecies of the elegans-group scrapers are uniform. Limb III of Extremalona gen. nov.have uniform, well-developed setae 4–6, this feature is unique for the genus. In theelegans-group these setae are of different morphology – seta 4 is flat, plumose, of vari-able size, seta 5 is of similar morphology with the same seta in Extremalona gen. nov.and seta 6 is small, twice as short and as thin as seta 5. Exopodites of limbs IV andV in Extremalona gen. nov. differ in shape from these of the elegans-group, and haverelatively short setae. Limb V of Extremalona gen. nov. have shortened inner setae,both of them are shorter than inner lobe of the limb, whereas in the elegans-group thedistal inner seta is much longer, 1.5 times longer than basal one.
Also, E. timmsi sp. nov. strongly differs from species of the elegans-group by thebody shape. Species of elegans-group have typical “Alona-like” body, with large low-set head, tip of rostrum located almost at the level of ventral margin of valves. Also,species of the elegans-group uniformly have moderately wide, subrectangular femalepostabdomen with parallel margins and with well-defined, but not protruding preanalangle. Males of Extremalona gen. nov. have unusual morphology of antennule, withsix lateral and nine terminal aesthetascs, while species of the elegans-group uniformlyhave two lateral and 10 terminal aesthetascs. The situation observed in Extremalonagen. nov. is rare for the subfamily in general, where most genera either lack lateralaesthetascs on male antennule, or, like the elegans-group, have only two terminalaesthetascs. Additional differences between the elegans-group and Extremalona gen.nov. observed in the morphology of antennule, antenna, labrum and postabdomenof female, and in the morphology of limb I of male (see Table 2) although not ofgenus-level themselves, also confirm the independent status of Extremalona gen. nov.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2860 A.Y. Sinev and R.J. Shiel
Table 2. Main differences between Extremalona gen. nov. and species of the elegans-group ofAlona s. l.
Extremalona gen. nov. Alona of elegans-group
Female charactersBody shape Irregular oval, with
maximum length atposterodorsal angle
Regular oval, withmaximum length at themiddle of dorsal margin
Head Small, high-set, tip ofrostrum located almost atthe middle of body height
Large, low-set, tip ofrostrum located almost atthe level of ventral valvesmargin
Posterior margin of valves Convex Almost straightPosteroventral angle of
valvesWith 25 setules With 50 to 100 setules in
different speciesLabral keel Very narrow, height/width
ratio about 2.5Broad, height/width ratio
about 1.5 in all speciesPostabdomen Narrowing distally, with
prominent preanal angleSubrectangular, preanal
angle well-defined, but notprominent
Aesthetascs of antennule Longest three as long asantennule itself; othersabout two-thirds length ofantennule
Longest three abouttwo-thirds length ofantennule; others abouthalf length of antennule
Spines of antennae Significantly shorter thanapical segments
Longer than apical segments
Epipodites of trunk limbs Merged with the limb body Of typical morphology,separated from the limbbody
Accessory seta of limb I Short, about one-quarterlength of ODL seta
Long, about two-thirds tothree-quarters length ofODL seta
IDL seta 1 Well-developed Very small, rudimentaryScrapers of limb II Differentiated Not differentiatedSetae 4–6 of exopodite III Long, of similar
morphology and sizeOf different size and
morphologyExopodite IV Broad oval Round or subquadrangularInner setae of limb V Short, of similar length,
shorter than inner lobeDistal setae 1.5 times of
more longer than basal,much longer than innerlobe
Male charactersAntennule With six lateral and nine
terminal aesthetascsWith two lateral and ten
terminal aesthetascsIDL seta 1 Present AbsentSetules on ventral face of
limb I below copulatorybrush
Long and thin, numerous(about 40)
Short, thick, not numerous(less than 20)
IDL, inner distal lobe of limb I; ODL, outer distal lobe of limb I.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2861
In comparison with other groups of the Coronatella-branch, Extremalona gen. nov.retains two important ancestral features – unique for the genus non-differentiatedsetae 4–6 of exopodite III and well-developed seta on IDL of limb I (this featureis shared by Karualona). Other important features of Extremalona gen. nov., likeadditional groups of lateral setules on postabdomen, thickened scrapers 5 and 6 oflimb II, unusual shapes of exopodites IV and V, numerous lateral aesthetascs in maleantennule, are obviously autoapomorphies of the genus. Like species of the elegans-group, Extremalona gen. nov. lack numerous advanced features characteristic for suchgenera as Coronatella, Karualona, Leberis and Anthalona (see Table 1).
Endemism in the Australia ChydoridaeTo date, 36 genera/120 species of Chydoridae and Sayciidae have been recordedfrom Australia. Twelve genera (31%) are endemic, with Saycia also known from NewZealand. In comparison, there are just three endemic genera in Eurasia (Brancelia VanDamme and Sinev, 2011, Nedorhynchotalona Kotov and Sinev, 2011 and PhreatalonaVan Damme et al., 2009), one in North America (Spinalona Ciros-Perez et Elias-Gutierrez, 1997), one in South America (Geoffreya Kotov et al., 2010), and one inAfrica (Dumontiellus Smirnov, 2006) (see Ciros-Perez and Elias-Gutierrez 1997; Kotovet al. 2010, 2012; Kotov and Sinev 2011; Smirnov 2006, 2008; Van Damme andSinev 2011; Van Damme et al. 2009). Three genera previously considered Australianendemics (Smirnov and Timms 1983; Frey 1991a; Sinev 2004a) have since been foundon other continents: Celsinotum in South America (Smirnov and Santos Silva 1995;Sinev and Elmoor-Loureiro 2010), Armatalona in Thailand (Sinev et al. 2007) anda species of Rak in South Africa (Smirnov 2008). But during the last decade stud-ies of Australian Aloninae revealed several new genera, three of them endemic (Sinev2004b; Sinev and Shiel 2008; Sinev and Kobayashi 2012), and more new genera canpossibly be revealed after revision of Australian species of Alona s.l. (Van Dammeet al. 2010). In toto approximately 50% of Australian chydorid species are endemic,many members of more widely distributed genera are known only from that con-tinent – globally the most diverse endemic chydorid fauna (see also Forró et al.2008).
Evidence is beginning to emerge of significant endemicity in other cladoceranfamilies, e.g. Macrotrichidae and Neotrichidae (Smirnov 1992), with an endemicfamily, Gondwanotrichidae, recently described (Van Damme et al. 2007a, b) from thesouthwest of Western Australia.
This apparent cladoceran radiation in the west of the continent, and particularlyin the southwest, was recognized by Frey (1991b: 358), who predicted that “more,possibly many more, species of Pleuroxus and related genera will be distinguished, par-ticularly in Western Australia, as the weakly saline, minute and ephemeral waterbodiesare studied more closely”. These observations apply equally to the northwest ofWestern Australia, which has hardly begun to be investigated. The region is subject tocyclonic rain events at the margins of arid zones, with a profusion of small, ephemeralwater bodies of widely differing water quality. It is likely that a suite of species remainto be described in genera such as Anthalona, recently reviewed (Van Damme et al.2011) but lacking Australian records. Species of Anthalona are known across the northof the continent, but their taxonomic position is as yet unresolved (Van Damme et al.2011).
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2862 A.Y. Sinev and R.J. Shiel
Drivers for the high level of endemicity in Australian Cladocera, and particu-larly the littoral Chydoridae, relate to the post-Gondwanan separation of eastern andwestern Australia by an epicontinental sea, continued isolation over an extremely longevolutionary time span, heterogeneity of both habitat and climate, including long peri-ods of aridity, such that the microcrustacea adapted to ephemerality. Habitats whichFrey (1991a,b) recognized as requiring particular attention are the shallow epidisodicpools of varying pH and salinity, which dry over summer. These ephemeral poolsrepresent “special” habitats to which some chydorids have adapted, and endemic, spe-cialized taxa can be expected in such areas. Our results confirm this opinion. Suchhabitats have barely begun to be sampled in Western Australia, and given the currentimpacts of mining on surface and subsurface waters in that State, the need for study ofthem cannot be overemphasized.
Acknowledgements
Our thanks to Brian Timms for providing the specimens of Extremalona gen. nov., and subse-quently site details and chemistry from the Guest Rd lakes, and for Nikolai Smirnov and AlexeyKotov for constructive critique and suggestions.
References
Alonso M. 1996. Crustacea Branchiopoda. Serie Fauna Ibérica, Vol. 7. Madrid: CSIC.Boronat L, Miracle MR, Armengol X. 2001. Cladoceran assemblages in a mineralization
gradient. Hydrobiologia 442:75–88.Ciros-Perez J, Elias-Gutierrez M. 1997. Spinalona anophtalma, n. gen. n. sp. (Anomopoda,
Chydoridae) a blind epigean cladoceran from the Neovolcanic Province of Mexico.Hydrobiologia 353:19–28.
Dumont HJ, Silva-Briano M. 2000 Karualona n. gen. (Anomopoda: Chydoridae), with adescription of two new species, and a key to all known species. Hydrobiologia 435:61–82.
Forró L, Korovchinsky NM, Kotov AA, Petrusek A. 2008. Global diversity of cladocerans(Cladocera: Crustacea) in freshwater. Hydrobiologia 595:177–184.
Frey DG. 1991a. A new genus of alonine chydorid cladocerans from athalassic saline waters ofNew South Wales, Australia. Hydrobiologia 224:11–48.
Frey DG. 1991b. The species of Pleuroxus and three related genera (Anomopoda, Chydoridae)in southern Australia and New Zealand. Rec Aust Mus. 434:291–372.
Frey DG. 1998. Expanded description of Leberis aenigmatosa Smirnov (Anomopoda:Chydoridae): a further indication of the biological isolation between western and easternAustralia. Hydrobiologia 367:31–42.
Halse SA. 2002. Diversity of Ostracoda (Crustacea) in inland waters of Western Australia. VerhInternat Verein Limnol. 28:914–918.
Halse SA, McRae JM. 2001. Calamoecia trilobata n. sp. (Copepoda: Calanoida) from salt lakesin south-western Australia. J R Soc Western Aust. 84:5–11.
Halse SA, Lyons MN, Pinder AM, Shiel RJ. 2004. Biodiversity patterns and their conservationin wetlands of the Western Australian wheatbelt. In: Keighery GJ, Halse SA, Harvey MS,McKenzie NL, editors. A biodiversity survey of the Western Australian agricultural zone.Rec WA Mus Suppl. 67:337–364.
Kotov AA, Jeong HG, Lee W. 2012. Cladocera (Crustacea: Branchiopoda) of the south-east ofthe Korean Peninsula, with twenty new records for Korea. Zootaxa 3368:50–90.
Kotov AA, Sinev AY. 2011 Cladocera (Crustacea, Branchiopoda) of the Zeya basin (AmurskayaArea, Russian Federation). 2. Descriptions of new taxa. Zool Zh. 90:272–284.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

Journal of Natural History 2863
Kotov AA, Sinev AY, Berrios VL. 2010. The Cladocera (Crustacea: Branchiopoda) of six highaltitude water bodies in the North Chilean Andes, with discussion of Andean endemism.Zootaxa 2430:1–66.
Myers N, Mittermeier RA, Mittermeier CG, da Foncesca GAB, Kents J. 2000. Biodiversityhotspots for conservation priorities. Nature 403:853–858.
Segers H, Shiel RJ. 2003. Microfaunal diversity in a biodiversity hotspot: new rotifers fromsouthwestern Australia. Zool Stud. 42:516–521.
Sinev AY. 2004a. Armatalona gen. n. – a new genus of subfamily Aloninae (Anomopoda,Chydoridae), separated from genus Alona. Hydrobiologia 520:29–47.
Sinev AY. 2004b. Miralona gen. n. – a new genus of the subfamily Aloninae (Anomopoda,Chydoridae) from Australia. Hydrobiologia 526:3–14.
Sinev AY, Alonso M, Miracle MR, Sahuquillo M. 2012. The West Mediterranean Alona azoricaFrenzel & Alonso, 1988 (Cladocera: Anomopoda: Chydoridae) is composed of two species.Zootaxa 3276:51–68.
Sinev AY, Alonso M, Sheveleva NG. 2009. New species of Alona from South-East Russia andMongolia related to Alona salina Alonso, 1996 (Cladocera: Anomopoda: Chydoridae).Zootaxa 2326:1–23.
Sinev AY, Elmoor-Loureiro LMA. 2010. Three new species of chydrorid cladocerans ofsubfamily Aloninae (Brachiopoda: Anomopoda: Chydoridae) from Brazil. Zootaxa2390:1–25.
Sinev AY, Kotov AA. 2012. New and rare Aloninae (Cladocera: Anomopoda: Chydoridae)from Indochina. Zootaxa 3334:1–28.
Sinev AY, Kobayashi T. 2012. Redescription of the endemic Australian cladoceran Alonawillisi (Smirnov, 1989) and its assignment to Acanthalona gen. n. (Cladocera: Anomopoda:Chydoridae). Zootaxa 3390:43–55.
Sinev AY, Nachai S, Sanoamuang L. 2007. Occurrence of the Australian cladoceran Armatalonamacrocopa (Sars, 1894) (Cladocera: Anomopoda: Chydoridae) in Thailand. Invert Zool.4:25–29.
Sinev AY, Shiel RJ 2008. Redescription of Alona macracantha Smirnov & Timms, 1983 andits reassignment to Maraura gen. nov. (Cladocera: Anomopoda: Chydoridae). J Nat Hist.42:2809–2824.
Sinev AY, Van Damme K, Kotov AA. 2005. Redescription of tropical-temperate cladocer-ans Alona diaphana King, 1853 and Alona davidi Richard, 1895 and their translocationto Leberis Smirnov, 1989 (Branchiopoda: Anomopoda: Chydoridae). Arthropoda Selecta.14:183–205.
Smirnov NN. 1971. Chydoridae of the world. Fauna SSSR. Rakoobraznie, 1(2). Leningrad:Nauka.
Smirnov NN. 1992. The Macrothricidae of the world. Guides to the Identification of theMicroinvertebrates of the Continental Waters of the World. The Hague: SPB AcadenicPublishing.
Smirnov NN. 2006 Pleuroxus-like chydorids (Crustacea: Anomopoda) from South Africa, withthe description of Dumontiellus africanus gen. n., sp. n. Hydrobiologia 575:433–439.
Smirnov NN. 2008 Check-List of the South-African Cladocera (Crustacea: Branchiopoda).Zootaxa 1788:47–56.
Smirnov NN, Santos-Silva EN. 1995. Some littoral anomopods (Crustacea) from CentralAmazonia. Hydrobiologia 315:227–230.
Smirnov NN, Timms BV. 1983. A revision of the Australian Cladocera (Crustacea). Rec AustMus Suppl. 1:1–132.
Timms BV. 2009. Study of the saline lakes of the Esperance hinterland, Western Australia, withspecial reference to the roles of acidity and episodicity. Nat Res Envt Issues. 15, article44:215–225.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012

2864 A.Y. Sinev and R.J. Shiel
Van Damme K, Brancelj A, Dumont HJ. 2009. Adaptations to the hyporheic in Aloninae(Crustacea: Cladocera): allocation of Alona protzi Hartwig, 1900 and related species toPhreatalona gen. nov. Hydrobiologia 618:1–34.
Van Damme K, Dumont HJ. 2008. Further division of Alona Baird, 1843: separation and posi-tion of Coronatella Dybowski & Grochowski and Ovalona gen. n. (Crustacea: Cladocera).Zootaxa 1960:1–44.
Van Damme K, Kotov AA, Dumont HJ. 2010. A checklist of names in Alona Baird, 1843(Crustacea: Cladocera: Chydoridae) and their current status: an analysis of the taxonomyof a lump genus. Zootaxa 2330:1–63.
Van Damme K, Shiel RJ, Dumont HJ. 2007a. Notothrix halsei gen. n., sp. n., representative ofa new family of freshwater cladocerans (Branchiopoda, Anomopoda) from SW Australia,with a discussion of ancestral traits and a preliminary molecular phylogeny of the order.Zool Scripta. 36:465–487.
Van Damme K, Shiel RJ, Dumont HJ. 2007b. Corrigendum – Gondwanotrichidae nom. nov.pro Nototrichidae Van Damme, Shiel & Dumont, 2007. Zool Scripta. 36:623.
Van Damme K, Sinev AY. 2011. A new genus of cave-dwelling microcrustaceans from theDinaric Region (south-east Europe): adaptations of true stygobitic Cladocera (Crustacea:Branchiopoda). Zool J Linn Soc. 161:31–52.
Van Damme K, Sinev AY, Dumont HG. 2011. Separation of Anthalona gen.n. from AlonaBaird, 1843 (Branchiopoda: Cladocera: Anomopoda): morphology and evolution ofscraping stenothermic alonines. Zootaxa 2875:1–64.
Dow
nloa
ded
by [
Art
em S
inev
] at
02:
09 0
7 D
ecem
ber
2012





























