Embed Size (px)
Citation preview

Molecular Microbiology (2004)
53
(2), 639–649 doi:10.1111/j.1365-2958.2004.04138.x
© 2004 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004
? 2004
53
2639649
Original Article
FbpA, a novel L. monocytogenes virulence factorS. Dramsi
et al
.
Accepted 19 March, 2004. *For correspondence. E-mail [email protected]; Tel. (
+
33) 1 45 68 88 41; Fax (
+
33) 1 45 68 87 06.
†
Presentaddress: SOREDAB SAS, La Tremblaye, 78125 La Boissière Ecole,France.
‡
These authors contributed equally to this work.
FbpA, a novel multifunctional
Listeria monocytogenes
virulence factor
S. Dramsi,
‡
F. Bourdichon,
†‡
D. Cabanes, M. Lecuit, H. Fsihi and P. Cossart*
Unité des Interactions Bactéries–Cellules, Institut Pasteur, 28 rue du Dr Roux, 75724 Paris, France.
Summary
Listeria monocytogenes
is a Gram-positive intracellu-lar bacterium responsible for severe opportunisticinfections in humans and animals. Signature-taggedmutagenesis (STM) was used to identify a genenamed
fbpA
, required for efficient liver colonizationof mice inoculated intravenously. FbpA was alsoshown to be required for intestinal and liver coloniza-tion after oral infection of transgenic mice expressinghuman E-cadherin.
fbpA
encodes a 570-amino-acidpolypeptide that has strong homologies to atypicalfibronectin-binding proteins. FbpA binds to immobi-lized human fibronectin in a dose-dependent and sat-urable manner and increases adherence of wild-type
L. monocytogenes
to HEp-2 cells in the presence ofexogenous fibronectin. Despite the lack of conven-tional secretion/anchoring signals, FbpA is detectedusing an antibody generated against the recombinantFbpA protein on the bacterial surface by immunoflu-orescence, and in the membrane compartment byWestern blot analysis of cell extracts. Strikingly, FbpAexpression affects the protein levels of two virulencefactors, listeriolysin O (LLO) and InlB, but not that ofInlA or ActA. FbpA co-immunoprecipitates with LLOand InlB, but not with InlA or ActA. Thus, FbpA, inaddition to being a fibronectin-binding protein,behaves as a chaperone or an escort protein for twoimportant virulence factors and appears as a novelmultifunctional virulence factor of
L. monocytogenes
.
Introduction
Listeria monocytogenes
is a ubiquitous Gram-positive,food-borne bacillus responsible for life-threatening infec-tions in humans and animals. It is a facultative intracellular
pathogen able to enter into a wide variety of host cellsincluding intestinal epithelial cells, hepatocytes, endothe-lial cells and fibroblasts. The cellular and molecular basisof its intracellular cycle have largely been elucidated (forrecent reviews, see Vazquez-Boland
et al
., 2001; Cossart,2002). Infection occurs in several successive steps, whichare entry of the bacterium into the host, lysis of the pha-gosomal vacuole, multiplication in the cytosol and directcell-to-cell spread using actin-based motility. Each steprequires expression of specific virulence factors. Themajor virulence genes are located in a cluster of genesencoding a regulatory protein PrfA, a phosphatidylinositol-specific phospholipase C (PlcA), a haemolysin calledlisteriolysin O (LLO), a metalloprotease (Mpl), an actin-recruiting protein (ActA) and a lecithinase (PlcB). On adifferent locus are encoded two proteins involved in inva-sion, InlA and InlB. Expression of virulence genes is co-ordinately and positively regulated by the transcriptionalregulator PrfA (Kreft and Vazquez-Boland, 2001; Milo-hanic
et al
., 2003). There is overwhelming evidence thatthe primary
L. monocytogenes
virulence determinant isLLO, which is responsible for escape from a vacuole andthus entrance in the cytosol (Dramsi and Cossart, 2002).Mutants lacking LLO are completely avirulent in themouse model of infection (Cossart
et al
., 1989).Adherence of
L. monocytogenes
to the host cell surfaceis a critical event during infection. It involves a number ofsurface proteins, including InlA (internalin), ActA, Ami andp104. InlA is the first member of the internalin multigenefamily characterized by the presence of a leucine-richrepeat region (Gaillard
et al
., 1991). InlA promotes adher-ence and entry into cell lines expressing its receptor, theadhesion protein E-cadherin (Mengaud
et al
., 1996a).ActA, the surface protein required for actin-based motility,may also promote attachment via host cell proteoglycans(Alvarez-Dominguez
et al
., 1997; Suarez
et al
., 2001).Ami is an autolysin, with an amidase activity present onthe surface of
L. monocytogenes
that contributes to bac-terial adherence to host cells (Milohanic
et al
., 2001).Finally, the involvement of a cell surface protein of104 kDa (p104) in the adhesion of
L. monocytogenes
tothe human intestinal cell line Caco-2 has been proposed(Pandiripally
et al
., 1999).Fibronectin is a dimeric glycoprotein (
ª
440 kDa) that ispresent in a soluble form in plasma and extracellular fluidsand in a fibrillar form of higher molecular weight on cell
640
S. Dramsi
et al.
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
53
, 639–649
surfaces. Although fibronectin has critical roles in eukary-otic cellular processes, such as adhesion, migration anddifferentiation, it is also a common substrate for theattachment of bacteria. The binding of pathogenic
Strep-tococcus pyogenes
and
Staphylococcus aureus
to epithe-lial cells via fibronectin facilitates their internalization(Fowler
et al
., 2000). The best characterized fibronectin-binding proteins from streptococci and staphylococcishare a similar structural organization and mechanism ofligand recognition (Joh
et al
., 1999). These surface pro-teins are cell wall-anchored proteins with a signal peptide,an LPXTG motif and a fibronectin-binding domain thatconsists of 45-amino-acid-long repeats within the C-termi-nal region. Streptococci also express several atypicalfibronectin-binding proteins. These proteins do not containany conventional secretion signal, anchorage motif or typ-ical fibronectin-binding sequences. Examples includeFbp54 in
S. pyogenes
(Courtney
et al
., 1994), PavA in
Streptococcus pneumoniae
(Holmes
et al
., 2001) andFbpA in
Streptococcus gordonii
(Christie
et al
., 2002).In this work, signature-tagged transposon mutagenesis
(STM) has allowed the identification of a new
L. monocy-togenes
gene,
fbpA
, required for efficient colonization ofhost tissues. The
fbpA
gene encodes a 570-amino-acidpolypeptide with strong homologies to the fibronectin-binding proteins PavA of
S. pneumoniae
, Fbp54 of
S.pyogenes
and FbpA of
S. gordonii
. We show here thatFbpA is a fibronectin-binding protein present on the liste-rial surface that can mediate adherence to host cells butalso modulates the protein levels of two virulence factors,LLO and InlB. These results point to the multiple contribu-tions of FbpA to
L. monocytogenes
virulence.
Results
Isolation of an attenuated mutant by STM
Signature-tagged mutagenesis of
L. monocytogenes
strain EGD was performed to identify new genes importantfor the development of listeriosis in the mouse model usingplasmid pID408t, which takes advantage of the transposi-tion properties of Tn
917
, as described previously for
S.aureus
(Mei
et al
., 1997). Ninety-six tagged plasmids,each carrying a different tag, were introduced individuallyby electroporation into the wild-type strain EGD andallowed to generate banks of
L. monocytogenes
insertionmutants. Pools of 96 differently tagged mutants were usedto infect 7- to 8-week-old Swiss mice intravenously. Weused a dose of 10
6
bacteria per mouse. Infected mice diedat day 4. In our protocol, mutants were recovered frominfected spleens and livers (output) 72 h after infection andcompared with the inoculated pool (input) by hybridization.Screening of
ª
2000 mutants resulted in obtaining 12attenuated mutants, among which one mutant (1D12) was
non-haemolytic as a result of transposon insertion in the
hly
gene, one of the main listerial virulence genes. Thismutant validated our approach. One attenuated mutant(3G11) was analysed further.
Virulence analysis of the 3G11 mutant
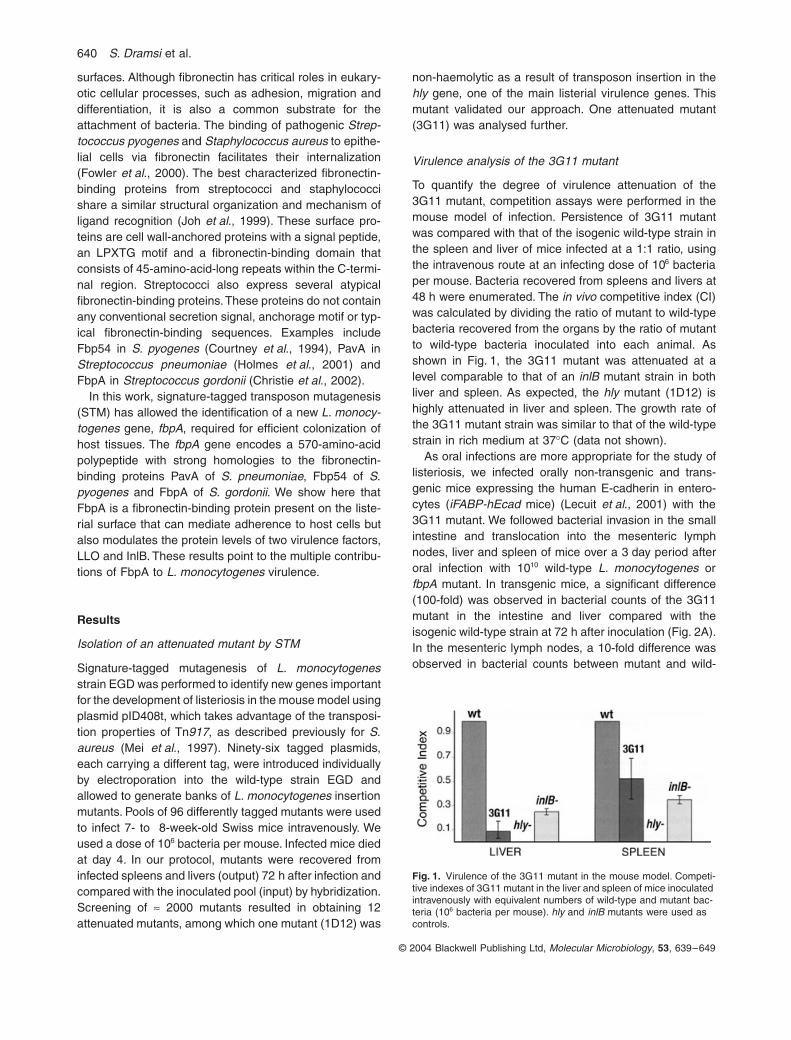
To quantify the degree of virulence attenuation of the3G11 mutant, competition assays were performed in themouse model of infection. Persistence of 3G11 mutantwas compared with that of the isogenic wild-type strain inthe spleen and liver of mice infected at a 1:1 ratio, usingthe intravenous route at an infecting dose of 10
6
bacteriaper mouse. Bacteria recovered from spleens and livers at48 h were enumerated. The
in vivo
competitive index (CI)was calculated by dividing the ratio of mutant to wild-typebacteria recovered from the organs by the ratio of mutantto wild-type bacteria inoculated into each animal. Asshown in Fig. 1, the 3G11 mutant was attenuated at alevel comparable to that of an
inlB
mutant strain in bothliver and spleen. As expected, the
hly
mutant (1D12) ishighly attenuated in liver and spleen. The growth rate ofthe 3G11 mutant strain was similar to that of the wild-typestrain in rich medium at 37
∞
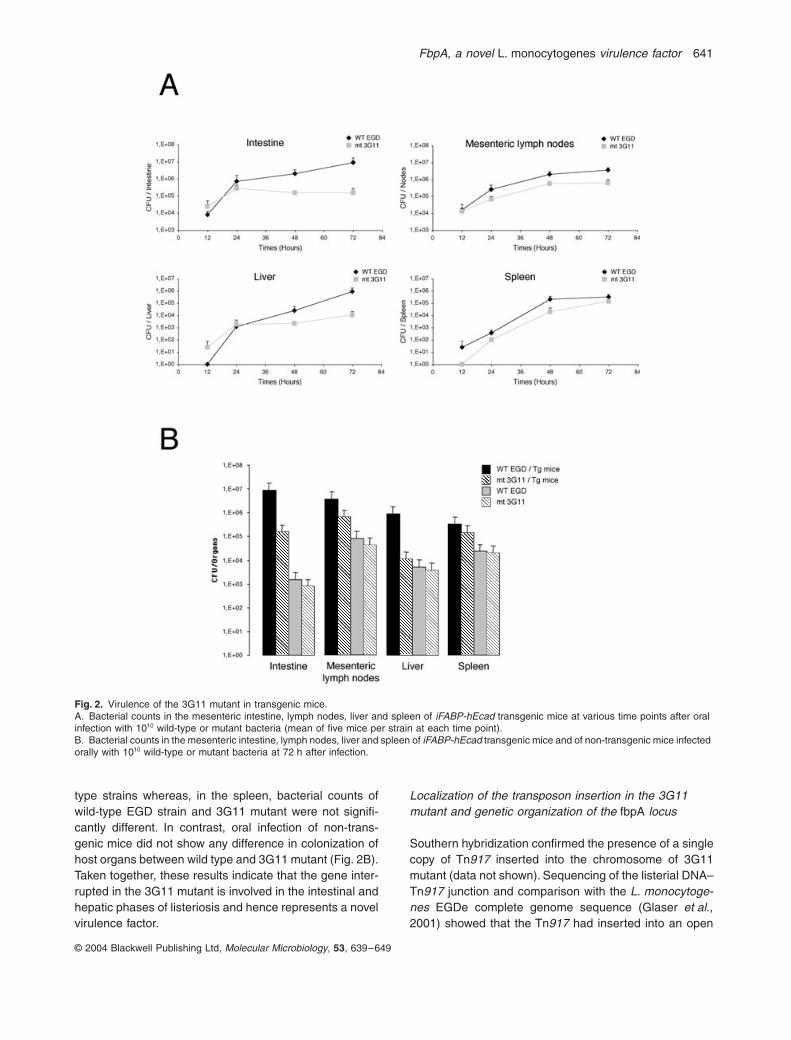
C (data not shown).As oral infections are more appropriate for the study of
listeriosis, we infected orally non-transgenic and trans-genic mice expressing the human E-cadherin in entero-cytes (
iFABP-hEcad
mice) (Lecuit
et al
., 2001) with the3G11 mutant. We followed bacterial invasion in the smallintestine and translocation into the mesenteric lymphnodes, liver and spleen of mice over a 3 day period afteroral infection with 10
10
wild-type
L. monocytogenes
or
fbpA
mutant. In transgenic mice, a significant difference(100-fold) was observed in bacterial counts of the 3G11mutant in the intestine and liver compared with theisogenic wild-type strain at 72 h after inoculation (Fig. 2A).In the mesenteric lymph nodes, a 10-fold difference wasobserved in bacterial counts between mutant and wild-
Fig. 1.
Virulence of the 3G11 mutant in the mouse model. Competi-tive indexes of 3G11 mutant in the liver and spleen of mice inoculated intravenously with equivalent numbers of wild-type and mutant bac-teria (10
6
bacteria per mouse).
hly
and
inlB
mutants were used as controls.
FbpA, a novel
L. monocytogenes
virulence factor
641
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
53
, 639–649
type strains whereas, in the spleen, bacterial counts ofwild-type EGD strain and 3G11 mutant were not signifi-cantly different. In contrast, oral infection of non-trans-genic mice did not show any difference in colonization ofhost organs between wild type and 3G11 mutant (Fig. 2B).Taken together, these results indicate that the gene inter-rupted in the 3G11 mutant is involved in the intestinal andhepatic phases of listeriosis and hence represents a novelvirulence factor.
Localization of the transposon insertion in the 3G11 mutant and genetic organization of the
fbpA
locus
Southern hybridization confirmed the presence of a singlecopy of Tn
917
inserted into the chromosome of 3G11mutant (data not shown). Sequencing of the listerial DNA–Tn
917
junction and comparison with the
L. monocytoge-nes
EGDe complete genome sequence (Glaser
et al
.,2001) showed that the Tn
917
had inserted into an open
Fig. 2.
Virulence of the 3G11 mutant in transgenic mice.A. Bacterial counts in the mesenteric intestine, lymph nodes, liver and spleen of
iFABP-hEcad
transgenic mice at various time points after oral infection with 10
10
wild-type or mutant bacteria (mean of five mice per strain at each time point).B. Bacterial counts in the mesenteric intestine, lymph nodes, liver and spleen of
iFABP-hEcad
transgenic mice and of non-transgenic mice infected orally with 10
10
wild-type or mutant bacteria at 72 h after infection.

642
S. Dramsi
et al.
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
53
, 639–649
reading frame (ORF) encoding a protein exhibiting stronghomologies (
ª
60% similarity and 40% identity over
ª
550amino acids) to the fibronectin-binding proteins FbpA of
S. gordonii
, PavA of
S. pneumoniae
, Fbp54 of
S. pyo-genes
and YloA of
Bacillus subtilis
(
Supplementary mate-rial
, Fig. S1). This gene, listed as
lmo1829
in the EGDegenome (http://genolist.pasteur.fr/ListiList), was desig-nated
fbp
A for fibronectin-binding protein A. It encodes aprotein of 570 amino acids with a predicted molecularmass of 65161 Da and a pI of 6.68. Figure 3 depicts thegenetic organization of the
fbpA
-containing region in
L.monocytogenes
strain EGDe (Glaser
et al
., 2001). Flank-ing
fbpA
(
lmo1829
) are two genes (
lmo1828
and
lmo1830
) transcribed in opposite directions and encodingconserved proteins of unknown functions. Lmo 1828 ishomologous to a putative stress-induced protein of theYicC family, and Lmo 1830 shows homology to a dehy-drogenase. A putative transcription terminator is presentdownstream from
fbpA
making unlikely a polar effect ofthe Tn
917
insertion on the downstream
lmo1830
gene.The transposon insertion in mutant 3G11 was locatedbetween nucleotides 1097 and 1098 of the ORF. Interest-ingly, the gene encoding
fbpA
is also present in the closelyrelated non-pathogenic
Listeria
species,
L. innocua
(des-ignated
lin1943
in L. innocua CLIP 11262 genome). Anal-ysis of the primary and secondary structures of the proteindid not reveal any putative signal peptide in the N-termi-nus or motifs (LPXTG signature, hydrophobic region, GWrepeats) characteristic of surface proteins in Gram-positive bacteria.
FbpA binds to immobilized fibronectin and mediates adherence to HEp-2 cells
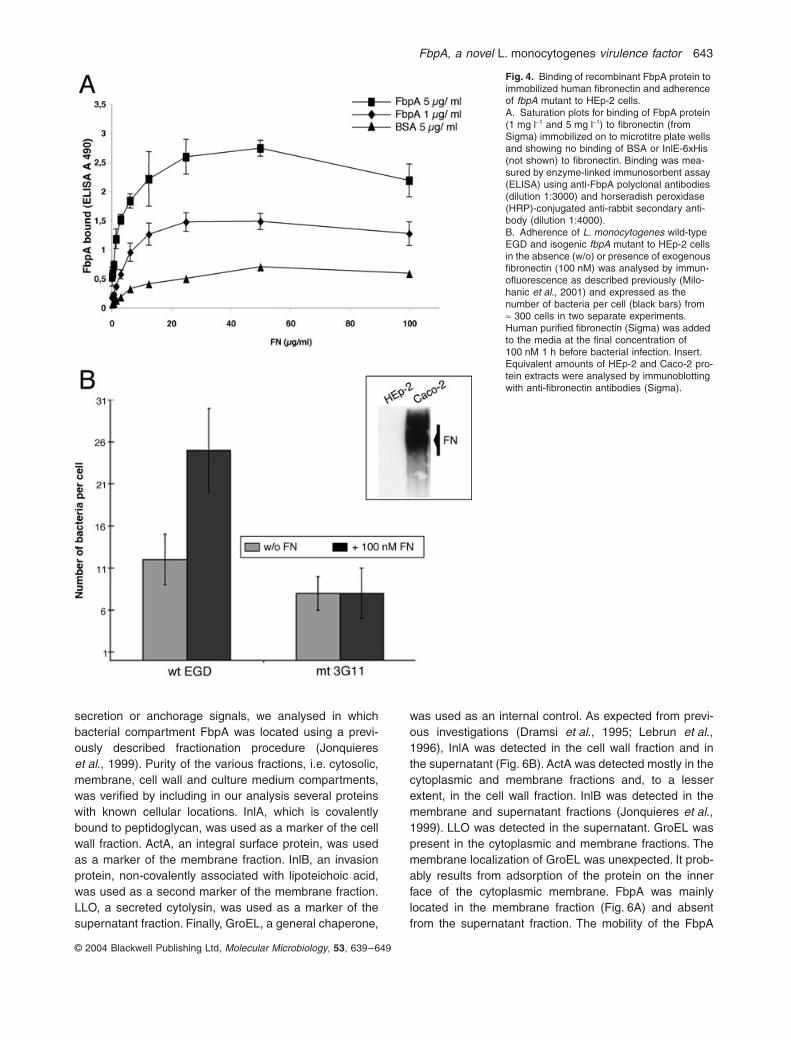
A full-length recombinant C-tagged FbpA-6xHis proteinwas purified from Escherichia coli and shown to bind tohuman fibronectin immobilized onto microtitre plate wells(Fig. 4A). Binding of FbpA to fibronectin was saturable,and maximal binding levels were dependent upon theamounts of immobilized fibronectin. Note that bovineserum albumin (BSA) used as a control did not bind tofibronectin, nor did an unrelated His-tagged protein InlE-6xHis. FbpA did not bind to fibrinogen (data not shown).
Most fibronectin-binding proteins are known to promoteadherence to eukaryotic cells through their affinity forfibronectin acting as a bridge between bacteria and host
cells (Fowler et al., 2000). The role of L. monocytogenesFbpA in bacterial adherence to human epithelial HEp-2cell line was investigated. HEp-2 cells were chosenbecause they are deficient in fibronectin synthesis com-pared with the human intestinal epithelial cell line Caco-2(see insert in Fig. 4B). As shown in Fig. 4B, adherencewas reduced by ªtwofold in the EGD:fbpA Tn917 mutantcompared with the isogenic wild-type strain. Addition ofexogenous fibronectin (100 nM) to HEp-2 cells increasedthe adherence of wild-type strain EGD, but not that of thefbpA mutant strain, resulting in a greater difference inadherence between wild type and the isogenic fbpAmutant strain. These results reinforce the idea that FbpAis a fibronectin-binding protein that contributes to L. mono-cytogenes adherence to the human epithelial HEp-2 cells.
FbpA is a listerial cell surface protein
To detect expression of FbpA protein by L. monocytoge-nes, FbpA-specific polyclonal antibodies were generatedagainst the purified recombinant FbpA-6xHis. In addition,complementation of the fbpA mutant was carried out bytransformation with the plasmid pFB1.5 carrying fbpAunder the control of its own promoter.
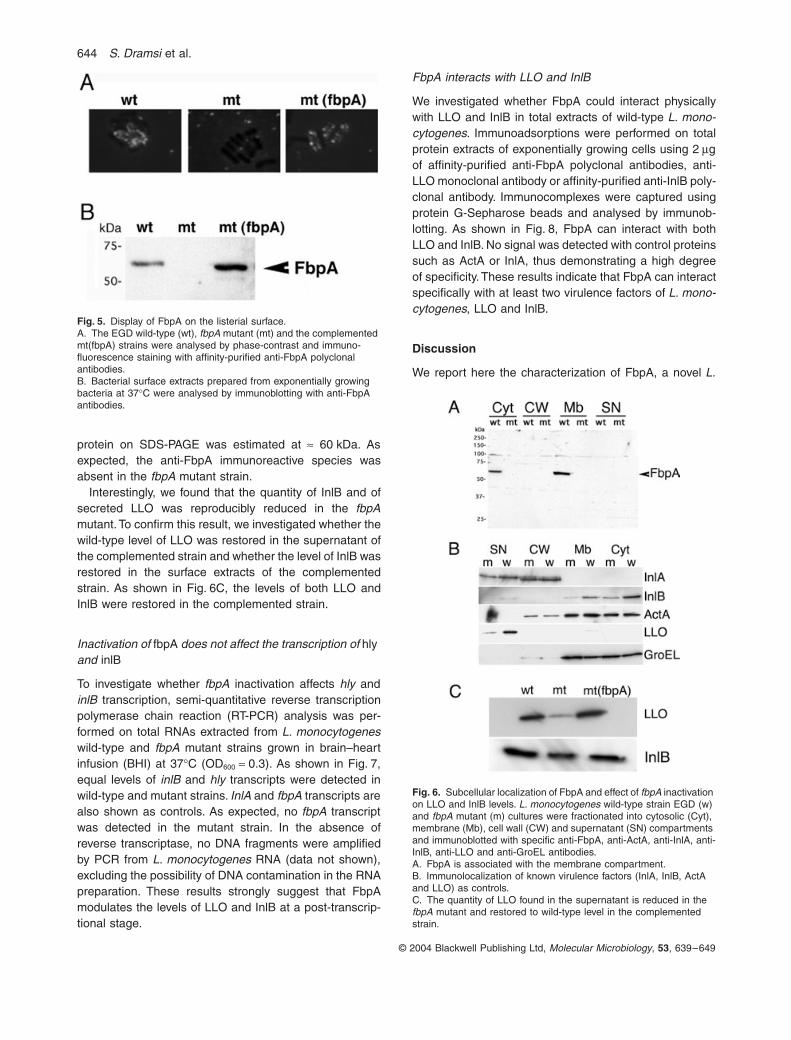
Immunofluorescence microscopy with affinity-purifiedFbpA antibodies demonstrated the surface localization ofthe FbpA protein on both wild-type and complemented L.monocytogenes strains. As expected, the anti-FbpAimmunoreactive species was absent from the surface ofthe fbpA mutant strain (Fig. 5A).
The surface proteins from wild-type, fbpA mutant andcomplemented strains were prepared by solubilizationwith 1% SDS as described previously (Kocks et al., 1992)and subjected to Western blotting analysis using affinity-purified FbpA polyclonal antibodies. The antiserum recog-nized a band of ª 60 kDa in both wild-type and comple-mented strains (Fig. 5B). The 60 kDa band was absentin the fbpA mutant. Altogether, these results indicatethat FbpA is expressed as a cell surface protein in L.monocytogenes.
FbpA associates with the bacterial cytoplasmic membrane and regulates the levels of LLO and InlB proteins
As the primary sequence of FbpA lacks the conventional
Fig. 3. Genetic organization of the fbpA locus on the L. monocytogenes chromosome. The transposon (Tn917) insertion in mutant 3G11 is shown by a black arrow. ORFs are indicated according to the L. monocytogenes genome nomenclature (Glaser et al., 2001). The fbpA gene corresponds to lmo1829. The putative transcription terminator is indicated.
FbpA, a novel L. monocytogenes virulence factor 643
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
secretion or anchorage signals, we analysed in whichbacterial compartment FbpA was located using a previ-ously described fractionation procedure (Jonquiereset al., 1999). Purity of the various fractions, i.e. cytosolic,membrane, cell wall and culture medium compartments,was verified by including in our analysis several proteinswith known cellular locations. InlA, which is covalentlybound to peptidoglycan, was used as a marker of the cellwall fraction. ActA, an integral surface protein, was usedas a marker of the membrane fraction. InlB, an invasionprotein, non-covalently associated with lipoteichoic acid,was used as a second marker of the membrane fraction.LLO, a secreted cytolysin, was used as a marker of thesupernatant fraction. Finally, GroEL, a general chaperone,
was used as an internal control. As expected from previ-ous investigations (Dramsi et al., 1995; Lebrun et al.,1996), InlA was detected in the cell wall fraction and inthe supernatant (Fig. 6B). ActA was detected mostly in thecytoplasmic and membrane fractions and, to a lesserextent, in the cell wall fraction. InlB was detected in themembrane and supernatant fractions (Jonquieres et al.,1999). LLO was detected in the supernatant. GroEL waspresent in the cytoplasmic and membrane fractions. Themembrane localization of GroEL was unexpected. It prob-ably results from adsorption of the protein on the innerface of the cytoplasmic membrane. FbpA was mainlylocated in the membrane fraction (Fig. 6A) and absentfrom the supernatant fraction. The mobility of the FbpA
Fig. 4. Binding of recombinant FbpA protein to immobilized human fibronectin and adherence of fbpA mutant to HEp-2 cells.A. Saturation plots for binding of FbpA protein (1 mg l-1 and 5 mg l-1) to fibronectin (from Sigma) immobilized on to microtitre plate wells and showing no binding of BSA or InlE-6xHis (not shown) to fibronectin. Binding was mea-sured by enzyme-linked immunosorbent assay (ELISA) using anti-FbpA polyclonal antibodies (dilution 1:3000) and horseradish peroxidase (HRP)-conjugated anti-rabbit secondary anti-body (dilution 1:4000).B. Adherence of L. monocytogenes wild-type EGD and isogenic fbpA mutant to HEp-2 cells in the absence (w/o) or presence of exogenous fibronectin (100 nM) was analysed by immun-ofluorescence as described previously (Milo-hanic et al., 2001) and expressed as the number of bacteria per cell (black bars) from ª 300 cells in two separate experiments. Human purified fibronectin (Sigma) was added to the media at the final concentration of 100 nM 1 h before bacterial infection. Insert. Equivalent amounts of HEp-2 and Caco-2 pro-tein extracts were analysed by immunoblotting with anti-fibronectin antibodies (Sigma).
644 S. Dramsi et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
protein on SDS-PAGE was estimated at ª 60 kDa. Asexpected, the anti-FbpA immunoreactive species wasabsent in the fbpA mutant strain.
Interestingly, we found that the quantity of InlB and ofsecreted LLO was reproducibly reduced in the fbpAmutant. To confirm this result, we investigated whether thewild-type level of LLO was restored in the supernatant ofthe complemented strain and whether the level of InlB wasrestored in the surface extracts of the complementedstrain. As shown in Fig. 6C, the levels of both LLO andInlB were restored in the complemented strain.
Inactivation of fbpA does not affect the transcription of hly and inlB
To investigate whether fbpA inactivation affects hly andinlB transcription, semi-quantitative reverse transcriptionpolymerase chain reaction (RT-PCR) analysis was per-formed on total RNAs extracted from L. monocytogeneswild-type and fbpA mutant strains grown in brain–heartinfusion (BHI) at 37∞C (OD600 = 0.3). As shown in Fig. 7,equal levels of inlB and hly transcripts were detected inwild-type and mutant strains. InlA and fbpA transcripts arealso shown as controls. As expected, no fbpA transcriptwas detected in the mutant strain. In the absence ofreverse transcriptase, no DNA fragments were amplifiedby PCR from L. monocytogenes RNA (data not shown),excluding the possibility of DNA contamination in the RNApreparation. These results strongly suggest that FbpAmodulates the levels of LLO and InlB at a post-transcrip-tional stage.
FbpA interacts with LLO and InlB
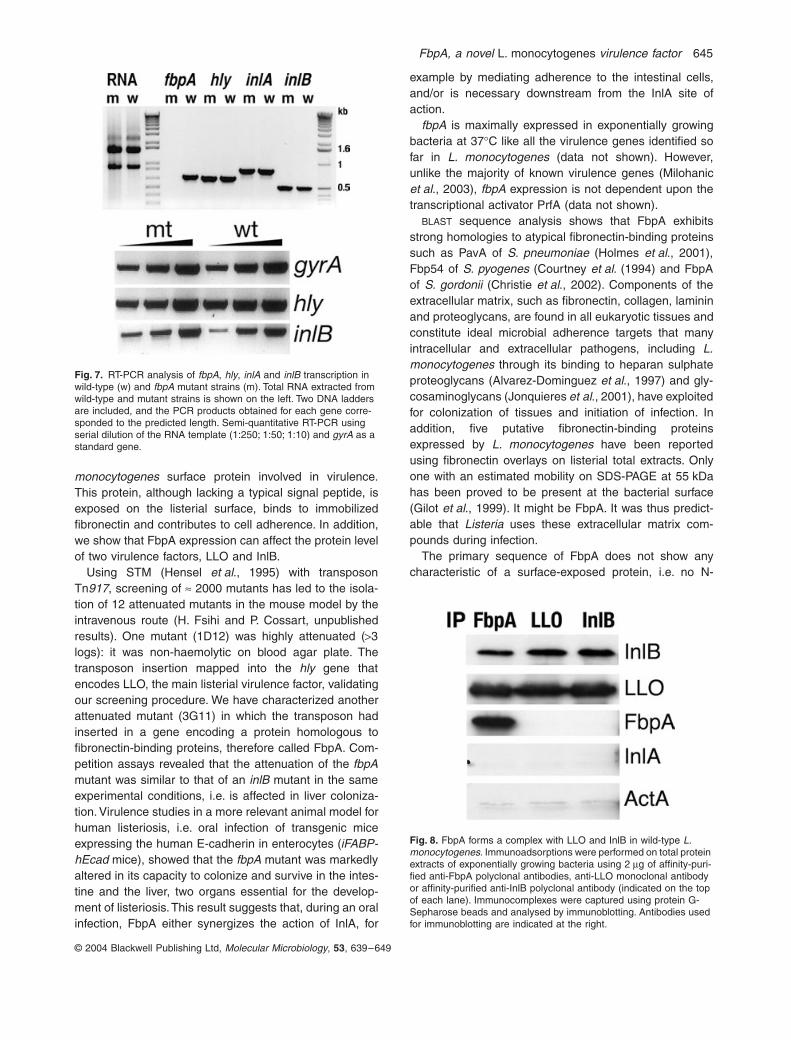
We investigated whether FbpA could interact physicallywith LLO and InlB in total extracts of wild-type L. mono-cytogenes. Immunoadsorptions were performed on totalprotein extracts of exponentially growing cells using 2 mgof affinity-purified anti-FbpA polyclonal antibodies, anti-LLO monoclonal antibody or affinity-purified anti-InlB poly-clonal antibody. Immunocomplexes were captured usingprotein G-Sepharose beads and analysed by immunob-lotting. As shown in Fig. 8, FbpA can interact with bothLLO and InlB. No signal was detected with control proteinssuch as ActA or InlA, thus demonstrating a high degreeof specificity. These results indicate that FbpA can interactspecifically with at least two virulence factors of L. mono-cytogenes, LLO and InlB.
Discussion
We report here the characterization of FbpA, a novel L.
Fig. 5. Display of FbpA on the listerial surface.A. The EGD wild-type (wt), fbpA mutant (mt) and the complemented mt(fbpA) strains were analysed by phase-contrast and immuno-fluorescence staining with affinity-purified anti-FbpA polyclonal antibodies.B. Bacterial surface extracts prepared from exponentially growing bacteria at 37∞C were analysed by immunoblotting with anti-FbpA antibodies.
Fig. 6. Subcellular localization of FbpA and effect of fbpA inactivation on LLO and InlB levels. L. monocytogenes wild-type strain EGD (w) and fbpA mutant (m) cultures were fractionated into cytosolic (Cyt), membrane (Mb), cell wall (CW) and supernatant (SN) compartments and immunoblotted with specific anti-FbpA, anti-ActA, anti-InlA, anti-InlB, anti-LLO and anti-GroEL antibodies.A. FbpA is associated with the membrane compartment.B. Immunolocalization of known virulence factors (InlA, InlB, ActA and LLO) as controls.C. The quantity of LLO found in the supernatant is reduced in the fbpA mutant and restored to wild-type level in the complemented strain.
FbpA, a novel L. monocytogenes virulence factor 645
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
monocytogenes surface protein involved in virulence.This protein, although lacking a typical signal peptide, isexposed on the listerial surface, binds to immobilizedfibronectin and contributes to cell adherence. In addition,we show that FbpA expression can affect the protein levelof two virulence factors, LLO and InlB.
Using STM (Hensel et al., 1995) with transposonTn917, screening of ª 2000 mutants has led to the isola-tion of 12 attenuated mutants in the mouse model by theintravenous route (H. Fsihi and P. Cossart, unpublishedresults). One mutant (1D12) was highly attenuated (>3logs): it was non-haemolytic on blood agar plate. Thetransposon insertion mapped into the hly gene thatencodes LLO, the main listerial virulence factor, validatingour screening procedure. We have characterized anotherattenuated mutant (3G11) in which the transposon hadinserted in a gene encoding a protein homologous tofibronectin-binding proteins, therefore called FbpA. Com-petition assays revealed that the attenuation of the fbpAmutant was similar to that of an inlB mutant in the sameexperimental conditions, i.e. is affected in liver coloniza-tion. Virulence studies in a more relevant animal model forhuman listeriosis, i.e. oral infection of transgenic miceexpressing the human E-cadherin in enterocytes (iFABP-hEcad mice), showed that the fbpA mutant was markedlyaltered in its capacity to colonize and survive in the intes-tine and the liver, two organs essential for the develop-ment of listeriosis. This result suggests that, during an oralinfection, FbpA either synergizes the action of InlA, for
example by mediating adherence to the intestinal cells,and/or is necessary downstream from the InlA site ofaction.
fbpA is maximally expressed in exponentially growingbacteria at 37∞C like all the virulence genes identified sofar in L. monocytogenes (data not shown). However,unlike the majority of known virulence genes (Milohanicet al., 2003), fbpA expression is not dependent upon thetranscriptional activator PrfA (data not shown).
BLAST sequence analysis shows that FbpA exhibitsstrong homologies to atypical fibronectin-binding proteinssuch as PavA of S. pneumoniae (Holmes et al., 2001),Fbp54 of S. pyogenes (Courtney et al. (1994) and FbpAof S. gordonii (Christie et al., 2002). Components of theextracellular matrix, such as fibronectin, collagen, lamininand proteoglycans, are found in all eukaryotic tissues andconstitute ideal microbial adherence targets that manyintracellular and extracellular pathogens, including L.monocytogenes through its binding to heparan sulphateproteoglycans (Alvarez-Dominguez et al., 1997) and gly-cosaminoglycans (Jonquieres et al., 2001), have exploitedfor colonization of tissues and initiation of infection. Inaddition, five putative fibronectin-binding proteinsexpressed by L. monocytogenes have been reportedusing fibronectin overlays on listerial total extracts. Onlyone with an estimated mobility on SDS-PAGE at 55 kDahas been proved to be present at the bacterial surface(Gilot et al., 1999). It might be FbpA. It was thus predict-able that Listeria uses these extracellular matrix com-pounds during infection.
The primary sequence of FbpA does not show anycharacteristic of a surface-exposed protein, i.e. no N-
Fig. 7. RT-PCR analysis of fbpA, hly, inlA and inlB transcription in wild-type (w) and fbpA mutant strains (m). Total RNA extracted from wild-type and mutant strains is shown on the left. Two DNA ladders are included, and the PCR products obtained for each gene corre-sponded to the predicted length. Semi-quantitative RT-PCR using serial dilution of the RNA template (1:250; 1:50; 1:10) and gyrA as a standard gene.
Fig. 8. FbpA forms a complex with LLO and InlB in wild-type L. monocytogenes. Immunoadsorptions were performed on total protein extracts of exponentially growing bacteria using 2 mg of affinity-puri-fied anti-FbpA polyclonal antibodies, anti-LLO monoclonal antibody or affinity-purified anti-InlB polyclonal antibody (indicated on the top of each lane). Immunocomplexes were captured using protein G-Sepharose beads and analysed by immunoblotting. Antibodies used for immunoblotting are indicated at the right.

646 S. Dramsi et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
terminal signal peptide that might direct the export ofthe protein via the general secretion pathway, no trans-membrane domain, no typical signature sequence thatcould explain cell surface association in L. monocytoge-nes (Cossart and Jonquieres, 2000). However, ourimmunoblotting and immunofluorescence data demon-strate unambiguously that FbpA is localized on the cellsurface of L. monocytogenes. Orthologues of FbpAincluding PavA of S. pneumoniae and Fbp54 of S. pyo-genes were also shown to be cell surface proteins(Courtney et al., 1996; Holmes et al., 2001). Moreover,immunizations with Fbp54 induce protective immuneresponses against S. pyogenes challenge in mice, indi-cating that Fbp54 is expressed in vivo and, in physiolog-ical conditions, behaves as a cell surface protein(Kawabata et al., 2001). Whether it is also the case forFbpA is unknown.
Fractionation studies showed that FbpA is associatedwith the bacterial cytoplasmic membrane. Moreover, FbpAcould be extracted from listerial surface with 1% SDS,reinforcing the finding of its association with the mem-brane. It is the first time, to our knowledge, that the preciselocalization of this type of atypical surface protein fromGram-positive bacteria has been determined. To testwhether mislocalization of LPXTG-containing proteins inthe cell wall of L. monocytogenes could alter the subcel-lular localization of FbpA, we analysed the localization ofFbpA in L. monocytogenes sortase A and B single anddouble mutants (Bierne et al., 2002; 2004). Expressionand membrane localization of FbpA in the sortasemutants were identical to that in isogenic wild-type strainEGDe (data not shown).
Recently, a second secA machinery has been discov-ered in L. monocytogenes (Lenz et al., 2003). This path-way allows secretion of 17 listerial proteins amongst which10 do not have a signal peptide. We therefore suspectedthat FbpA might be exported through this non-essentialsecretion pathway. It is indeed the case (S. Dramsi and P.Cossart, unpublished). Whether other FbpA-like proteinsof Gram-positive bacteria also use this pathway is undercurrent investigation.
The inactivation of fbpA correlated with a significantreduction in the protein levels of two virulence factors, LLOand InlB. Interestingly, the protein levels of InlA and ActA,two other virulence factors, were not affected by the fbpAmutation, suggesting that the effect of FbpA is specific andindependent of PrfA, the central regulator of virulencegenes in L. monocytogenes. Semi-quantitative RT-PCRanalysis indicated that the regulation of hly and inlB byfbpA occurs at a post-transcriptional level. It is worth not-ing that inactivation of fbpA in S. gordonii led to a signifi-cant reduction in the expression of the major cell wall-anchored fibronectin-binding protein, CshA, albeit at thetranscriptional level (Christie et al., 2002).
In view of these results, we propose that FbpA, in addi-tion to being a fibronectin-binding protein, also acts as a‘chaperone’ that can prevent degradation and/or increasethe secretion mechanism of specific virulence proteinssuch as LLO and InlB. These findings raise the possibilityof an indirect role for FbpA and the other homologousfibronectin-binding proteins of this family in virulence.Indeed, in S. pneumoniae, pavA mutants were stronglyimpaired (104-fold) in virulence in the mouse sepsismodel, which was somewhat surprising as mutants inadhesins are expected to be moderately attenuated (10-to 100-fold). We hypothesize that, as in L. monocytoge-nes, PavA can modulate the expression of key virulencefactors in S. pneumoniae. It is important to note that, inanother STM study of type 3 S. pneumoniae, Lau et al.(2001) also isolated an attenuated mutant in pavA.
Thus, although the role of these FbpA proteins is notfully understood, they are all clearly important for infection.
Experimental procedures
Strains, plasmids and growth conditions
Brain–heart infusion (BHI; Difco Laboratories) and Luria–Bertani (LB; Difco Laboratories) broth and agar were used togrow Listeria and Escherichia coli strains respectively. Thewild-type virulent strain of L. monocytogenes EGD (BUG600) belongs to the serovar 1/2a. The EGD:fbpA Tn917mutant (3G11) was put in the collection as BUG 1898. Strainsharbouring plasmids were grown in the presence of thefollowing antibiotics: pAT28 derivatives, 60 mg l-1 spe-ctinomycin, pET22b+ derivatives, 100 mg l-1 ampicillin. L.monocytogenes strains carrying the Tn917 transposon weregrown in the presence of 5 mg l-1 erythromycin.
General DNA techniques
Plasmid DNA from E. coli was prepared by rapid alkaline lysisusing the QIAprep spin prep kit (Qiagen), and PCR productswere purified using the QIAquick kit (Qiagen) according tothe manufacturer’s instructions. Genomic DNA from L. mono-cytogenes was prepared using the Boehringer–Roche highpurification kit. Standard techniques were used for DNA frag-ment isolation, DNA cloning and restriction analysis (Sam-brook et al., 1989). Restriction enzymes and ExpandTM High-fidelity Taq polymerase were purchased from Boehringer–Roche. Nucleotide sequencing was carried out by GenomeExpress.
Trans-complementation with fbpA
A 1987 bp fbpA fragment was amplified by PCR from EGDgenomic DNA using the primers starting 256 bp before thestart codon (5¢-GGGGTACCCCAATTCGGAGTAGCGGTGG-3¢) and 20 bp downstream from the stop codon (5¢-ACATGCATGCATGTACGGCTCCACGAAGGAAC-3¢), which containKpnI and SphI sites respectively (underlined). After KpnI andSphI digestion, the PCR product was cloned into the shuttlevector pAT28 (Trieu-Cuot et al., 1990). The resulting plas-

FbpA, a novel L. monocytogenes virulence factor 647
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
mid, pFB1.5 in E. coli, BUG 2122, was introduced into L.monocytogenes mutant strain 3G11 by conjugation asdescribed previously (Poyart and Trieu-Cuot, 1997) givingrise to BUG 1900. The construction was verified bysequencing the insert on both strands.
RNA isolation and RT-PCR assays
Bacteria (10 ml) were grown in BHI broth until OD600 reached0.3. Bacteria were centrifuged, washed twice with 1 ml of 1¥PBS and resuspended in 1 ml of Trizol (Invitrogen). Bacteriawere broken using a FastPROTEIN BLUE tube (Bio101 Sys-tems) and homogenized using a FastPrep instrument at aspeed of 6.5 for 30 s. RNA was extracted with 300 ml ofchloroform–isoamyl alcohol 24:1 (Sigma). After 10 min ofcentrifugation at 13 000 g, the aqueous phase was trans-ferred to a tube containing 270 ml of isopropanol. Total RNAwas then precipitated overnight at 4∞C and washed with 1 mlof a 75% ethanol solution before suspension in RNase-freewater. Contaminating DNA was removed by digestion withRNA-free DNase I according to the manufacturer’s instruc-tions (Roche), and RNAs were cleaned up using RNeasyprotocol (Qiagen). RT-PCR analysis was carried out using theSuperscriptTM one-step RT-PCR system according to themanufacturer’s instructions (Life Technologies). Sequenceswere as follows: inlB primers, forward AGCAGGGACGCGGATAACTGC and reverse CCTTGGTAGACCGATAGCTTATTC; hly primers, forward CCACGGAGATGCAGTGACAAATG and reverse GACGGCCATACGCCACACTTGAG; inlAprimers, forward CAACATTTAGTGGAACCGTGACGC andreverse GGGTCATAAGCGTTCATTGTACTTG; fbpA primers,forward CGCCAACTTGAGTACTCTTCTAG and reverseCGATTGAATCACCACATGCGAAC; gyrA primers, forwardGTTGCCCGTGCCCTACCTG and reverse GCGCGACGGATTCCGCTAC.
Expression and purification of recombinant FbpA-6xHis
A 1730 bp PCR fragment was produced using genomic DNAfrom EGD as template and the primers 5¢-(GGAATTC)CATATGGCGTTTGATGCAATG-3¢ and 5¢-(CCG)CTCGAGATTTTTTAACTCTAAAACGAGC-3¢, which contain NdeI andXhoI sites respectively (underlined). The resulting fragmentwas digested with NdeI and XhoI and inserted in frameupstream from the His tag sequence in the expression vectorpET22b+ (Novagen). The resulting plasmid, pFB2.6, was ver-ified by sequencing the insert from both junctions. It was usedto transform E. coli DH5a giving rise to BUG 1958 and E. coliBL21(DE3) (Novagen) as recommended by the manufac-turer. Recombinant FbpA-6xHis was purified using a two-stepchromatographic procedure. The first step of purification bymetal affinity chromatography (Novagen) has been detailedelsewhere (Braun et al., 1998). The fractions containingFbpA-6xHis were pooled and subjected to anion exchangechromatography on a POROS HQ20 column (PerSeptive Bio-systems) using a Biocad Sprint HPLC (PE Biosystem). Theloading buffer was 50 mM Tris (pH 8) and 150 mM NaCl.Protein was then eluted by an NaCl gradient up to 1 M in thesame buffer, and the UV detector at 280 nm monitored theprotein presence. The protein fractions were pooled and con-centrated on Centriprep 30 devices (Amicon). Protein con-centrations were determined with the BCA system (Pierce).
Cell culture, adherence assays
The human HEp-2 epithelial cell line was cultured in Dul-becco’s modified Eagle medium (DMEM with Glutamax,25 mM glucose; Gibco Laboratories) supplemented with 10%fetal bovine serum (FBS; Bio West) and incubated in a 10%CO2 atmosphere at 37∞C. For the assays, cells were seededat 5 ¥ 104 cells ml-1 on to 22 mm square glass coverslips in24-well plates (adherence assays). Adherence assays wereperformed as described previously (Milohanic et al., 2001)with exponentially growing bacteria (0D600 ª0.8). Human puri-fied fibronectin (Sigma) was added to the media 1 h beforebacterial infection.
Microscopy, fractionation and immunoblots
Generation of polyclonal antibodies against recombinantFbpA-6xHis and subsequent affinity purification were per-formed as described previously (Friederich et al., 1995).Immunofluorescence staining of FbpA was performedaccording to procedures described elsewhere (Lebrun et al.,1996) using polyclonal antibody (this work) and a fluoresceinisothiocyanate (FITC)-conjugated secondary antibody (Bio-sys). For analysis of FbpA expression, bacterial proteinswere separated into cytoplasm, membrane, cell wall andculture medium, according to a previously described protocol(Jonquieres et al., 1999). Bacteria were grown in richmedium (BHI) at 37∞C and harvested for protein analysisduring the exponential phase of culture. Equivalent amountsof all fractions, representing 100 or 200 ml of bacterial culture,were analysed by immunoblotting. Alternatively, cell surfaceextracts were prepared as described previously (Kocks et al.,1992). Proteins were boiled in Laemmli sample buffer,resolved on a 10% SDS-PAGE gel and transferred to nitro-cellulose membrane. FbpA was detected using polyclonalantibodies and horseradish peroxidase (HRP)-coupled anti-rabbit secondary antibodies (Biosys) and the ECL kit (Amer-sham). The other primary antibodies used in this study werepolyclonal antibodies raised against ActA (Cabanes et al.,2003), InlB (Braun et al., 1997) and GroEL (Sigma), andmonoclonal antibodies raised against InlA (L7.7; Mengaudet al., 1996b) and LLO (Nato et al., 1991). Secondary HRP-conjugated goat anti-mouse antibodies were from Biosys.
Immunoblotting to detect fibronectin synthesis in HEp-2and Caco-2 cell extracts was performed using polyclonalantibodies raised against fibronectin (Sigma).
Binding of FbpA to immobilized fibronectin
Binding of C-terminal 6xHis-tagged FbpA protein to fibronec-tin immobilized on to microtitre plate wells was measuredessentially as described previously (Braun et al., 2000).Briefly, wells were coated overnight at 4∞C with fibronectin(or BSA as a control) in the range 0.1–10 mg per well in PBS.After blocking with 1% BSA in PBS, the plates were incubatedwith 100 ml of various concentrations of FbpA-6xHis, BSA orInlE-6xHis diluted in PBS. Bound FbpA was detected usingan anti-FbpA polyclonal antibody, and InlE-6xHis wasrevealed using the penta-His monoclonal antibody (Qiagen).Antibodies were detected using HRP-conjugated anti-rabbitor anti-mouse antibody and the chromogenic substrate 1,2-phenylenediamine dihydrochloride (Dako). The absorbance

648 S. Dramsi et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
of the resulting colour development was measured at492 nm.
Infection of mice
Competition experiments were performed by co-infecting thesame animal (four mice) with equal amounts of wild-type andmutant strains (106 bacteria final) and sampling each strainat day 3 in both liver and spleen by plating on selective platesand counting the bacteria recovered from each organ. Deter-mination of the competitive index is as follows:
where wt represents the number of wild-type bacteria and mtrepresents the number of mutant bacteria.
Oral infections were performed by infecting 8-week-oldiFABP-hEcad transgenic mice with 1010 wild-type or mutantbacteria diluted in 0.5 ml of PBS containing 50 mg of CaCO3
(Sigma) and injected intragastrically with a needle (0.9 mmby 38 mm, ref 20GX1; 1/2¢¢ Popper) as described previously(Lecuit et al., 2001). Four animals for each time point werekilled and dissected for determination of bacterial counts inground organs.
Acknowledgements
We are extremely grateful to Edith Gouin for invaluable helpin antibody preparation and purification, Renaud Jonquièresfor expertise in running the HPLC, Javier Pizzaro and ElianeMilohanic for assistance in the cellular assays, and PierreDehoux for bioinformatic analysis. We thank Ji-Min Mei forsupplying the Tn917 tags, and David Holden for helpful dis-cussions on STM. This work received financial support fromthe Pasteur Institute, the Ministère de l’Education Nationale,de la Recherche et de la Technologie (Programme de Micro-biologie Fondamentale et Appliquée, Maladies Infectieuses,Environnement et Bioterrorisme) and the European Commis-sion (contract QLG2-CT 1999-00932). P.C. is an internationalscholar from the Howard Hughes Medical Institute.
Supplementary material
The following material is available from http://www.blackwellpublishing.com/products/journals/suppmat/mmi/mmi4138/mmi4138sm.htmFig. S1. Multiple sequence alignment of FbpA-like proteinsfrom different species.
References
Alvarez-Dominguez, C., Vazquez-Boland, J.A., Carrasco-Marin, E., Lopez-Mato, P., and Leyva-Cobian, F. (1997)Host cell heparan sulfate proteoglycans mediate attach-ment and entry of Listeria monocytogenes, and the listerialsurface protein ActA is involved in heparan sulfate receptorrecognition. Infect Immun 65: 78–88.
Bierne, H., Mazmanian, S.K., Trost, M., Pucciarelli, M.G.,Liu, G., Dehoux, P., et al. (2002) Inactivation of the srtAgene in Listeria monocytogenes inhibits anchoring of
Competition Index Formula
Ci
Output mt wtInput mt wt
=( / )
( / )
surface proteins and affects virulence. Mol Microbiol 43:869–881.
Bierne, H., Garandeau, C., Pucciarelli, M.G., Sabet, C., New-ton, S., Garcia-del Portillo, F., et al. (2004) Sortase B, anew class of sortase in Listeria monocytogenes. J Bacteriol186: 1972–1982.
Braun, L., Dramsi, S., Dehoux, P., Bierne, H., Lindahl, G.,and Cossart, P. (1997) InlB: an invasion protein of Listeriamonocytogenes with a novel type of surface association.Mol Microbiol 25: 285–294.
Braun, L., Ohayon, H., and Cossart, P. (1998) The InIBprotein of Listeria monocytogenes is sufficient to promoteentry into mammalian cells. Mol Microbiol 27: 1077–1087.
Braun, L., Ghebrehiwet, B., and Cossart, P. (2000) gC1q-R/p32, a C1q-binding protein, is a receptor for the InlB inva-sion protein of Listeria monocytogenes. EMBO J 19: 1458–1466.
Cabanes, D., Dussurget, O., Dehoux, P., and Cossart, P.(2003) Auto, a surface associated autolysin of Listeriamonocytogenes required for entry into eukaryotic cells andvirulence. Mol Microbiol 51: 1601–1614.
Christie, J., McNab, R., and Jenkinson, H.F. (2002) Expres-sion of fibronectin-binding protein FbpA modulates adhe-sion in Streptococcus gordonii. Microbiology 148: 1615–1625.
Cossart, P. (2002) Molecular and cellular basis of the infec-tion by Listeria monocytogenes: an overview. Int J MedMicrobiol 291: 401–409.
Cossart, P., and Jonquieres, R. (2000) Sortase, a universaltarget for therapeutic agents against gram-positive bacte-ria? Proc Natl Acad Sci USA 97: 5013–5015.
Cossart, P., Vicente, M.F., Mengaud, J., Baquero, F., Perez-Diaz, J.C., and Berche, P. (1989) Listeriolysin O is essen-tial for virulence of Listeria monocytogenes: direct evi-dence obtained by gene complementation. Infect Immun57: 3629–3636.
Courtney, H.S., Li, Y., Dale, J.B., and Hasty, D.L. (1994)Cloning, sequencing, and expression of a fibronectin/fibrin-ogen-binding protein from group A streptococci. InfectImmun 62: 3937–3946.
Courtney, H.S., Dale, J.B., and Hasty, D.I. (1996) Differentialeffects of the streptococcal fibronectin-binding protein,FBP54, on adhesion of group A streptococci to humanbuccal cells and HEp-2 tissue culture cells. Infect Immun64: 2415–2419.
Dramsi, S., Biswas, I., Maguin, E., Braun, L., Mastroeni, P.,and Cossart, P. (1995) Entry of Listeria monocytogenesinto hepatocytes requires expression of inIB, a surfaceprotein of the internalin multigene family. Mol Microbiol 16:251–261.
Dramsi, S., and Cossart, P. (2002) Listeriolysin O: a genuinecytolysin optimized for an intracellular parasite. J Cell Biol-ogy 156: 943–946.
Fowler, T., Wann, E.R., Joh, D., Johansson, S., Foster, T.J.,and Hook, M. (2000) Cellular invasion by Staphylococcusaureus involves a fibronectin bridge between the bacterialfibronectin-binding MSCRAMMs and host cell beta1 inte-grins. Eur J Cell Biol 79: 672–679.
Friederich, E., Gouin, E., Hellio, R., Kocks, C., Cossart, P.,and Louvard, D. (1995) Targeting of Listeria monocytoge-nes ActA protein to the plasma membrane as a tool to

FbpA, a novel L. monocytogenes virulence factor 649
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 53, 639–649
dissect both actin-based cell morphogenesis and ActAfunction. EMBO J 14: 2731–2744.
Gaillard, J.L., Berche, P., Frehel, C., Gouin, E., and Cos-sart, P. (1991) Entry of L. monocytogenes into cells ismediated by internalin, a repeat protein reminiscent ofsurface antigens from gram-positive cocci. Cell 65:1127–1141.
Gilot, P., Andre, P., and Content, J. (1999) Listeria monocy-togenes possesses adhesins for fibronectin. Infect Immun67: 6698–6701.
Glaser, P., Frangeul, L., Buchrieser, C., Rusniok, C., Amend,A., Baquero, F., et al. (2001) Comparative genomics ofListeria species. Science 294: 849–852.
Hensel, M., Shea, J.E., Gleeson, C., Jones, M.D., Dalton, E.,and Holden, D.W. (1995) Simultaneous identification ofbacterial virulence genes by negative selection. Science269: 400–403.
Holmes, A.R., McNab, R., Millsap, K.W., Rohde, M.,Hammerschmidt, S., Mawdsley, J.L., and Jenkinson, H.F.(2001) The pavA gene of Streptococcus pneumoniaeencodes a fibronectin-binding protein that is essential forvirulence. Mol Microbiol 41: 1395–1408.
Joh, D., Wann, E.R., Kreikemeyer, B., Speziale, P., andHook, M. (1999) Role of fibronectin-binding MSCRAMMsin bacterial adherence and entry into mammalian cells.Matrix Biol 18: 211–223.
Jonquieres, R., Bierne, H., Fiedler, F., Gounon, P., and Cos-sart, P. (1999) Interaction between the protein InlB ofListeria monocytogenes and lipoteichoic acid: a novelmechanism of protein association at the surface of gram-positive bacteria. Mol Microbiol 34: 902–914.
Jonquieres, R., Pizarro-Cerda, J., and Cossart, P. (2001)Synergy between the N- and C-terminal domains of InlBfor efficient invasion of non-phagocytic cells by Listeriamonocytogenes. Mol Microbiol 42: 955–965.
Kawabata, S., Kunitomo, E., Terao, Y., Nakagawa, I., Kikuchi,K., Totsuka, K., and Hamada, S. (2001) Systemic andmucosal immunizations with fibronectin-binding proteinFBP54 induce protective immune responses against Strep-tococcus pyogenes challenge in mice. Infect Immun 69:924–930.
Kocks, C., Gouin, E., Tabouret, M., Berche, P., Ohayon, H.,and Cossart, P. (1992) L. monocytogenes-induced actinassembly requires the actA gene product, a surface pro-tein. Cell 68: 521–531.
Kreft, J., and Vazquez-Boland, J.A. (2001) Regulation of vir-ulence genes in Listeria. Int J Med Microbiol 291: 145–157.
Lau, G.W., Haataja, S., Lonetto, M., Kensit, S.E., Marra, A.,Bryant, A.P., et al. (2001) A functional genomic analysis oftype 3 Streptococcus pneumoniae virulence. Mol Microbiol40: 555–571.
Lebrun, M., Mengaud, J., Ohayon, H., Nato, F., and Cossart,P. (1996) Internalin must be on the bacterial surface tomediate entry of Listeria monocytogenes into epithelialcells. Mol Microbiol 21: 579–592.
Lecuit, M., Vandormael-Pournin, S., Lefort, J., Huerre, M.,Gounon, P., Dupuy, C., et al. (2001) A transgenic model
for listeriosis: role of internalin in crossing the intestinalbarrier. Science 292: 1722–1725.
Lenz, L.L., Mohammadi, S., Geissler, A., and Portnoy,D.A. (2003) SecA2-dependent secretion of autolyticenzymes promotes Listeria monocytogenes pathogene-sis. Proc Natl Acad Sci USA 100: 12432–12437.
Mei, J.M., Nourbakhsh, F., Ford, C.W., and Holden, D.W.(1997) Identification of Staphylococcus aureus virulencegenes in a murine model of bacteraemia using signature-tagged mutagenesis. Mol Microbiol 26: 399–407.
Mengaud, J., Ohayon, H., Gounon, P., Mege, R.M., andCossart, P. (1996a) E-cadherin is the receptor for interna-lin, a surface protein required for entry of L. monocytoge-nes into epithelial cells. Cell 84: 923–932.
Mengaud, J., Lecuit, M., Lebrun, M., Nato, F., Mazie, J.C.,and Cossart, P. (1996b) Antibodies to the leucine-richrepeat region of internalin block entry of Listeria monocy-togenes into cells expressing E-cadherin. Infect Immun 64:5430–5433.
Milohanic, E., Jonquieres, R., Cossart, P., Berche, P., andGaillard, J.L. (2001) The autolysin Ami contributes to theadhesion of Listeria monocytogenes to eukaryotic cells viaits cell wall anchor. Mol Microbiol 39: 1212–1224.
Milohanic, E., Glaser, P., Coppee, J.Y., Frangeul, L., Vega,Y., Vazquez-Boland, J.A., et al. (2003) Transcriptome anal-ysis of Listeria monocytogenes identifies three groups ofgenes differently regulated by PrfA. Mol Microbiol 47:1613–1625.
Nato, F., Reich, K., Lhopital, S., Rouyre, S., Geoffroy, C.,Mazie, J.C., and Cossart, P. (1991) Production and char-acterization of neutralizing and nonneutralizing monoclonalantibodies against listeriolysin O. Infect Immun 59: 4641–4646.
Pandiripally, V.K., Westbrook, D.G., Sunki, G.R., and Bhunia,A.K. (1999) Surface protein p104 is involved in adhesionof Listeria monocytogenes to human intestinal cell line,Caco-2. J Med Microbiol 48: 117–124.
Poyart, C., and Trieu-Cuot, P. (1997) A broad-host-rangemobilizable shuttle vector for the construction of transcrip-tional fusions to beta-galactosidase in gram-positive bac-teria. FEMS Microbiol Lett 156: 193–198.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: a Laboratory Manual. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press.
Suarez, M., Gonzalez-Zorn, B., Vega, Y., Chico-Calero, I.,and Vazquez-Boland, J.A. (2001) A role for ActA in epithe-lial cell invasion by Listeria monocytogenes. Cell Microbiol3: 853–864.
Trieu-Cuot, P., Carlier, C., Poyart-Salmeron, C., and Courva-lin, P. (1990) A pair of mobilizable shuttle vectors confer-ring resistance to spectinomycin for molecular cloning inEscherichia coli and in gram-positive bacteria. NucleicAcids Res 18: 4296.
Vazquez-Boland, J.A., Kuhn, M., Berche, P., Chakraborty, T.,Dominguez-Bernal, G., Goebel, W., et al. (2001) Listeriapathogenesis and molecular virulence determinants. ClinMicrobiol Rev 14: 584–640.





























