Embed Size (px)
Citation preview

Fetal FueIs. I. Utilization of Ketones by Isolated Tissues at Various Stages of Maturation and Maternal Nutrition During Late Gestation
George E. Shombaugh, III, Suzanne C. Mrozak, and Norbert Freinkel
The availability and utilization of B- hydroxybutyrate as an alternate oxidative fuel during fasting hypoglycemia has been examined in the rat conceptus at 18 and 20days gestation. A 48-hr maternal fast between days 16 and 18 or 18 and 20 re- sulted in a 50% fall in fetal glucose levels and a marked rise in B-hydroxybutyrate, i.e., 30-fold at 18 and 60-fold at 20 days. Tissue concentrations of 5hydroxybutymte or acetoacetate did not exceed extracellu- lar levels. Placenta, fetal brain, carcass, and liver all oxidized “C-labeled B- hydroxybutymte to “CO, when incubated in vitro in the presence of Bhydroxybuty- mte. Highest rates of oxidation were ap parent in the placenta, followed by bmin, liver, and carcass. The D isomer of Bhydroxybutyrate appeared to be oxi- dized preferentially by all tissues studied. Despite levels of 3-ketoacid CoA trans-
ferase and acetoacetyl CoA thiolase lower
at 18 than at 20 days, rates of oxidation in individual tissues incubated under identical concentrations of substrate were similar at both times. In liver and brain, increasing rates of “CO? generation pro- portionate to graded concentrations of Bhydroxybutyrate in vitro indicated that such rates were probably determined by substrate availability. B-hydroxybutyrate oxidation in extrahepatic fetal tissues was unaffected by maternal fasting. By con- trast, fetal liver derived from fasted mothers generated significantly less “CO, from B-hydroxybutyrate than livers from fed mothers. It has been suggested that capabilities for ketone utilization are widespread in tissues of the conceptus, and that such utilization may fulfill in part the oxidative demands for continued anabolic growth during fasting hypo- glycemia in the mother.
F ASTING DURING LATE GESTATION results in accelerated maternal
weight loss, heightened lipolysis,‘,2 ketogenesis,3-6 diminished levels of
plasma amino acids,‘s8 hypoglycemia,5~7~9-” and enhanced elaboration of cate- cholamines.12 In the face of these alterations in maternal fuels and the pattern
of “accelerated starvation,” l3 the mammalian fetus appears well adapted to survive a maternal fast4v5 That some of the fetal metabolic and growth demands under these conditions might be met by the utilization of ketones has been
inferred from the finding of ketone oxidizing enzymes in placenta and other
tissues of the neonatal rat’4Js and the manifest oxidation of ketones by perfused
human brain.16 However, the regulatory determinants have not been clarified. Low levels of enzyme activity measured under optimal substrate concentrations in fetal tissues have not provided information regarding their contribution to
From the Center for Endocrinology, Metabolism, and Nutrition, Department of Medicine, North-
western University Medical School, and Veterans Administration Lakeside Hospital, Chicago, IN. Received for publication July 8. 1976.
Supported in part by USPHS Research Grant A M-10699 and Training Grant A M-05071 from the
National Institute of Arthritis and Metabolic Diseases, Bethesda, Md.. from the National Institutes of Health, Bethesda, Maryland; The Kroc Foundation, a Schweppe Foundation Grant, The Chicago Wesley Memorial Hospital Cancer Research Fund, Chicago, III.. and Veterans Administration Lake- side Hospital.
Reprint requests should be addressed to Dr. George E. Shambaugh, Iii, General Medical Research. Veterans Administration Lakeside Hospital, 333 E. Huron St., Chicago, Ill. 6041 I.
Metoboiism, Vol. 26, No. 6 (June), 1977 623

624 SHAMBAUGH, MROZAK, AND FREINKEL
ketone oxidation under conditions of substrates that prevail in vivo. The rela- tive ease of manipulation of the maternal environment makes it potentially im-
portant to assess whether fetal rates of ketone utilization are controlled by levels of enzymes or by the availability of substrates derived from the mother. Since activities of B-hydroxybutyrate dehydrogenase and acetoacetyl CoA thiolase in fetal tissues have been reported to increase between the 18th and
20th days of gestation, ‘4.‘5 it seemed that this question could be approached by examining the impact of fuel concentrations and potential alterations in ketone
utilizing enzymes on ketone oxidation by tissues of the conceptus at sequential
stages of development. Herein are presented data which indicate that several
tissues of the conceptus oxidize ketones in vitro in the presence of physiologic
concentrations of B-hydroxybutyrate, and that substrate levels rather than
enzyme activities appear to regulate the utilization of ketones as alternate oxi-
dative fuels.
MATERIAL AND METHODS
Reagents. Acetoacetyl coenzyme A, coenzyme A, NAD. NADH+, DL B-hydroxybutyrate,
acetoacetate, and Trizma were obtained from Sigma Chemicals, St. Louis, MO. D 3-hydroxybutyric
acid-3 14C (26 mCi/mmole) and DL 3-hydroxybutyric acid-3 14C (4.7 mCi/mmole) were purchased
from Amersham/Searle and 3-hydroxybutyrate dehydrogenase (SA 12 units/mg) from Boehringer
Biochemicals, Mannheim, Germany. Aquasol and Hyamine OH were obtained from New England
Nuclear and Packard Instruments, respectively. Incubation flasks, stoppers, and center wells were
purchased from Kontes Glass, Vineland, N.J. All chemicals used were of reagent grade.
Test animals. Primiparous pregnant rats mated at 6 wk of age were given unrestricted access
to food and water. Gestational age was calculated from the breeding date and confirmed by fetal
weight at the time of sacrifice. “*I* Animals were deprived of food for 48 hr from the morning of
the 16th or 18th day to the morning of the 18th or 20th day, respectively. Fasted animals were
allowed continuous access to water. In the studies to be described, all tissues were obtained
between 8:00 a.m. and I I:00 a.m.
Preparation of samples. For analysis of maternal plasma, samples were obtained by aortic
puncture from rats anesthetized with pentobarbital. Fetuses and attached placentas were then
delivered and fetal blood obtained by heparinized micropipet from the neck following decapita-
tion. During bleeding, the placenta was gently compressed to prevent backflow through the um-
bilical vessels. Plasma specimens were deproteinized with Ba(OH)z-ZnS04 for glucose analysis
with glucose oxidase.” For ketone assays, aliquots of plasma and whole blood were precipitated
v/v with 8.45% perchloric acid and the 15.000 g supernatant neutralized with 307, KOH. Tissue
ketone concentrations were determined in portions of placenta and fetal brain, liver, and carcass
obtained from nonanesthetized mothers that had been sacrificed by decapitation with a guillotine.
“Carcass,” defined previously,” represented a preparation containing 40% skin and 60”< muscle and bone derived from pelvis and lower extremities. The total time lapse between sacrifice of the
mother and introduction of the fetal tissues into liquid nitrogen was less than 2 min. Frozen
tissues were homogenized in 3 ml 4.257; perchloric acid and centrifuged for IS min at 15,000 g,
and the supernatant was neutralized with 30% KOH. In vitro incubations were conducted with
freshly excised tissues from nonanesthetized mothers. The tissues were rinsed twice with ice-cold
Krebs bicarbonate buffer, pH 7.4, and then cut into minces, 0.5-l mm dimensions, utilizing a
McIlwain tissue chopper. A total of lOC-200 mg of minced tissue was added to incubation flasks
containing 2 ml Krebs Ringer buffer. The maximum time lapse between sacrifice of the mother
and delivery of tissues into the flasks was less than 5 min.
For enzyme assays freshly excised tissues were homogenized 10% w/v in 0.25 M sucrose con- taining 0.3% sodium deoxycholate.‘4 Homogenates were centrifuged for I5 min at 20,000 g and the supernatant utilized for assay of acetoacetyl CoA thiolase and 3-ketoacid CoA transferase.

FETAL FUELS 625
Procedures
Assays of ketones. Acetoacetate was measured spectrophotometrically utilizing the reduction
of NAD at 340 pm, as described by Mellanby and Williamson.zo B-hydroxybutyrate was measured
similarly, utilizing the oxidation of NADH + as described by Williamson and Mellanby.” Dupli-
cate assays varied by less than 5%.
Enzyme assays. Acetoacetyl CoA thiolase was assayed spectrophotometrically by a moditica-
tion of the method of Williamson et al.,” wherein the decomposition of acetoacetyl CoA in the
presence of thiolase is measured at 313 pm. 3-ketoacid CoA transferase was assayed by the
spectrophotometric method of Williamson et al.z3 Both enzymes were assayed at 37°C. Duplicate
assays varied by less than 5%.
In vifro studies. Ketone utilization was estimated by measuring evolution of 14C02 from
hydroxybutyrate-3 14C in vitro.24 Minces of placenta and fetal liver, brain, or carcass in IO&
l50-mg aliquots were incubated at 37°C in 2 ml Krebs Ringer bicarbonate buffer (KRB), pH 7.4.
Preliminary measurements of glucose and B-hydroxybutyrate in fetal plasma from 20-day pregnant
rats which had been fasted for 48 hr disclosed average concentrations of glucose and B-hydroxy-
butyrate of I.1 and 5.4 mM, respectively. The KRB media were supplemented with these same
concentrations of glucose and B-hydroxybutyrate to simulate in vitro the availability of fuels in
vivo. Each tissue was incubated in quadruplicate. The contents of one pair of flasks were labeled
with 0.2 $Zi DL B-hydroxybutyrate-3 14C, and contents of the second pair with 0.2 PCi D
B-hydroxybutyrate-3 14C. For experiments utilizing the labeled D isomer, the concentration of
the nonradioactive B-hydroxybutyrate carrier containing equal quantities of both D and L isomers
was assumed to be halved. In experiments where graded concentrations of B-hydroxybutyrate
were utilized, the medium was modified to contain glucose I. I mM and lactate 8 mM.
Flasks were gased for 5 min at 37°C with 95% 02/5% CO, prior to incubation for 40 min.
Reactions were stopped by placing flasks on ice, adding 0.2 ml hyamine hydroxide to the center
well and 0.2 ml IO N H,SO, to the medium and continuing to shake flasks for 30 min at 37°C
thereafter to evolve 14C02. Center wells containing hyamine were then transferred to counting
vials containing I5 ml Aquasol. The radioactive assays were converted to nmoles of CO, evolved
from B-hydroxybutyrate on the basis of the radioactivity in an aliquot of the incubation medium
containing known quantities of this ketone body. Control experiments indicated that no detect-
able spontaneous decarboxylation occurred under the above conditions with either preparation
of labeled B-hydroxybutyrate and that evolution of 14C02 was completed 30 min following addition
of the acid. For all tissues, the reactions were linear during 40 min of incubation. Duplicate
assays varied by less than 10%. No 14C02 was evolved when tissues were boiled prior to incuba-
tion.
Histologic studies. Portions of 20-day fetal liver from fed and fasted mothers were fixed in
Bouin’s solution, embedded in the same block, and stained with hematoxylin and eosin. In each
section, four to six photomicrographs of fed and fasted livers were obtained at high dry magnifica-
tion. At least 100 nucleated cells were counted in identical grids laid over each photomicrograph,
in order to identify hematopoietic cells and hepatocytes. Results were then averaged for a single
section. A total of I2 sections were studied.
Other analyses. Segments of fetal liver and placenta were frozen in liquid nitrogen and anal-
yzed for glycogen as described elsewhere.‘2’25 Protein and DNA were estimated by the methods
of Lowry et al.26 and Martin,27 respectively. Student’s r test was utilized for statistical analyses.
RESULTS
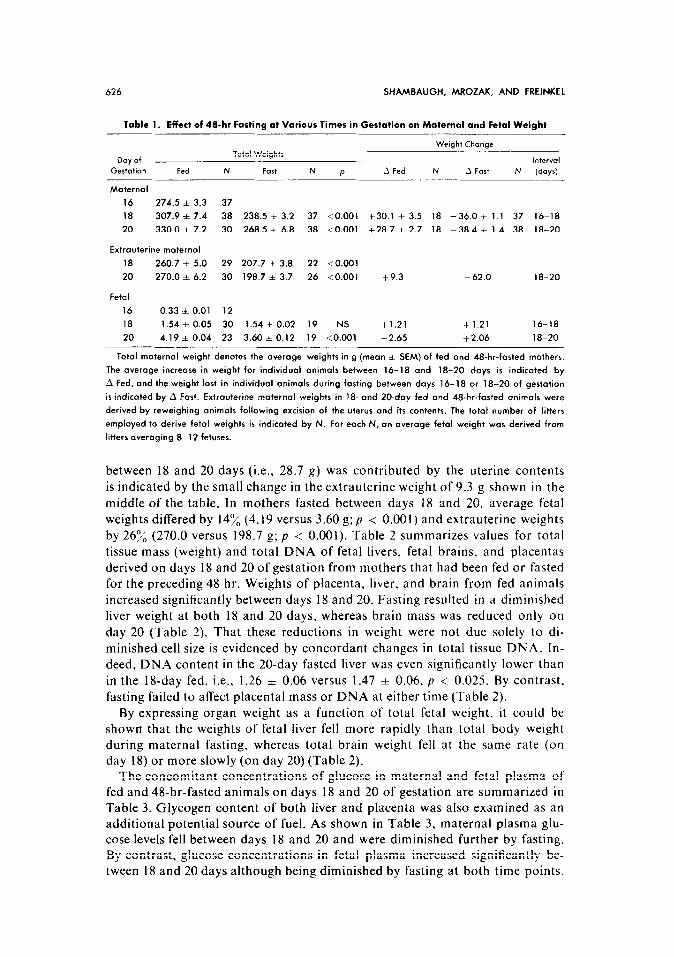
The effects of 48-hr fasting during days 16-18 or 18-20 of gestation upon body weights are summarized in Table 1. During both intervals, the changes in total body weight were similar, i.e., 30.1 f 3.5 and 28.7 A 2.7 g in the fed and -36.0 f 1.1 and -38.4 * 1.4 g in the fasted groups. The litter size in all groups ranged from 8 to 12 fetuses, and was similar in both fed and fasted ani- mals, i.e., 10.7 f 0.3 and 9.9 f 0.3 (mean + SEM) at 18 days and 10.5 f 0.5 and 9.9 + 0.3 at 20 days. That the majority of weight gain in the fed animals
626 SHAMBAUGH, MROZAK, AND FREINKEL
Table 1. Effect of 48-hr Fasting at Various Times in Gestation on Maternal and Fetal Weight
Weight Change
Day of Total Weights
lntervol
Gestation Fed N Fort N P .A Fed N A Fart N (days)
Maternal
16 274.5 f 3.3 37
18 307.9 * 7.4 38 238.5 f 3.2 37 <O.OOl +30.1 f 3.5 18 -36.0 zt 1.1 37 16-18
20 330.0 f 7.2 30 268.5 f 6.8 38 <O.OOl i-28.7 f 2.7 18 -38.4 i 1.4 38 18-20
Extrauterine maternal
18 260.7 f 5.0 29 207.7 f 3.8 22 <O.OOl
20 270.0 f 6.2 30 198.7 * 3.7 26 <O.OOl +9.3 - 62.0 18-20
Fetal
16 0.33 i 0.01 12
18 1.54 f 0.05 30 1.54 f 0.02 19 NS +1.21 +1.21 16-18
20 4.19 zt 0.04 23 3.60 zk 0.12 19 <O.OOl +2.65 +2.06 18-20
Total maternal weight denotes the average weights in g (mean * SEM) of fed and 48-hr-fasted mothers.
The average increase in weight for individual animals between 16-18 and 18-20 days is indicated by
A Fed, and the weight lost in individual animals during fosting between days 16-18 or 18-20 of gestation
is indicated by A Fast. Extrauterine maternal weights in 18. and ZO-day fed and 48-hr-fasted animals were
derived by reweighing animals following excision of the uterus and its contents. The total number of litters
employed to derive fetal weights is indicated by N. For each N, an average fetal weight was derived from
litters averaging 8-12 fetuses.
between 18 and 20 days (i.e., 28.7 g) was contributed by the uterine contents
is indicated by the small change in the extrauterine weight of 9.3 g shown in the
middle of the table. In mothers fasted between days 18 and 20, average fetal
weights differed by 14% (4.19 versus 3.60 g; p < 0.001) and extrauterine weights by 26:& (270.0 versus 198.7 g; p < 0.001). Table 2 summarizes values for total
tissue mass (weight) and total DNA of fetal livers, fetal brains, and placentas derived on days 18 and 20 of gestation from mothers that had been fed or fasted for the preceding 48 hr. Weights of placenta, liver, and brain from fed animals increased significantly between days 18 and 20. Fasting resulted in a diminished
liver weight at both 18 and 20 days, whereas brain mass was reduced only on
day 20 (Table 2). That these reductions in weight were not due solely to di-
minished cell size is evidenced by concordant changes in total tissue DNA. In- deed, DNA content in the 20-day fasted liver was even significantly lower than
in the 18-day fed, i.e., 1.26 f 0.06 versus 1.47 & 0.06, p < 0.025. By contrast. fasting failed to affect placental mass or DNA at either time (Table 2).
By expressing organ weight as a function of total fetal weight, it could be shown that the weights of fetal liver fell more rapidly than total body weight during maternal fasting, whereas total brain weight fell at the same rate (on day 18) or more slowly (on day 20) (Table 2).
The concomitant concentrations of glucose in maternal and fetal plasma of fed and 48-hr-fasted animals on days 18 and 20 of gestation are summarized in Table 3. Glycogen content of both liver and placenta was also examined as an additional potential source of fuel. As shown in Table 3, maternal plasma glu- cose levels fell between days 18 and 20 and were diminished further by fasting. By contrast, glucose concentrations in fetal plasma increased significantly be- tween 18 and 20 days although being diminished by fasting at both time points.

Tab
le
2.
Eff
ects
of
Fas
ting
on
Mas
s an
d D
NA
Co
nte
nt
of T
issu
es in
the
Con
cept
us
Day
o
f T
ota
l M
oss
(g)
To
tal
DN
A
(mg
) P
er C
ent
of
Tota
l Fe
tal
We
igh
t
Ge
sta
tion
Fe
d
N
Fort
N
P
Fed
N
Fo
rt
N
P Fe
d
N
Fart
N
P
Pla
ce
nta
18
0.45
9 f
0.01
9 16
0.
429
f 0.
011
15
NS
1.
16
f 0.
05
16
1.05
~1~
0.
03
15
NS
30
.42
f 2.
13
7 26
.80
f 1.
36
7 N
S
20
0.53
5 zt
0.
022$
14
0.
526
f 0.
0161
: 14
N
S
1.13
zt
0.
06
14
l.Ob
zt
0.06
14
N
S
13.0
4 f
0.53
$ 7
14.1
9 f
0.47
1 7
NS
liver
18
0.12
2 f
0.00
2 10
0.
106&
0.
004
12
<O.O
l 1.
47
f 0.
06
10
l.lO
zt
0.06
12
<O
.OO
l 7.
95
f 0.
12
7 7.
38
f 0.
19
7 co
.05
20
0.31
9 +
O.O
OS
t.
11
0.20
7 f
O.O
lOt
14
<O.O
Ol
1.63
&
0.
10
11
1.26
zt
0.06
14
<O
.OO
l 7.
72
f 0.
18
7 6.
93
f 0.
18
7 <O
.Ol
18
0.13
4 f
0.00
6 12
0.
119~
0.
004
14
NS
0.
83
* 0.
02
12
0.74
~
0.06
14
N
S
7.19
+
0.16
7
6.99
+
0.26
7
NS
20
0.19
9 *
0.00
s:
14
0.18
1 f
0.00
61
14
<O.O
Ol
0.94
&
0.02
* 14
0.
87zt
0.
02
14
<0.
05
4.05
f
0.09
1 7
4.74
f
0.15
t 7
<0.
005
Wei
gh
ts
of
tiss
ues
fr
om
fe
d
and
48
-hr-
fast
ed
anim
als
wer
e m
easu
red
at
18
an
d
20
day
s g
esta
tio
n,
and
to
tal
DN
A
con
ten
t es
tim
ated
O
S d
escr
ibed
in
M
ater
ials
an
d
Met
ho
ds.
p
den
ote
s co
mp
aris
on
s o
f fe
d
ve
rsu
s
fast
ed
at
each
ti
me
po
int;
su
per
scri
pts
d
eno
te
sig
nif
ican
ce
of
dif
fere
nce
s b
etw
een
va
lues
o
bta
ined
o
n
day
18
v
ers
us
d
ay
20
(*
40.0
5;
t 10
.01;
1
<O
.OO
l).
At
leas
t si
x p
lace
nta
s o
r ti
ssu
es
fro
m
six
fetu
ses
in
the
som
e lit
ter
wer
e u
tiliz
ed
to
der
ive
a si
ng
le
valu
e.
N
den
ote
s th
e to
tal
nu
mb
er
of
litte
rs
wh
ich
w
ere
sam
ple
d.
Ind
ivid
ual
ti
ssu
e w
eig
hts
h
av
e
also
b
een
d
ivid
ed
by
tota
l fe
tal
bo
dy
wei
gh
t to
o
bto
in
an
esti
mat
e o
f th
e p
erce
nta
ge
con
trib
uti
on
o
f th
at
tiss
ue
to
tota
l fe
tal
mas
s.
All
resu
lts
ore
sh
ow
n
as t
he
mea
n
f S
EM
.
628 SHAMBAUGH, MROZAK, AND FREINKEL
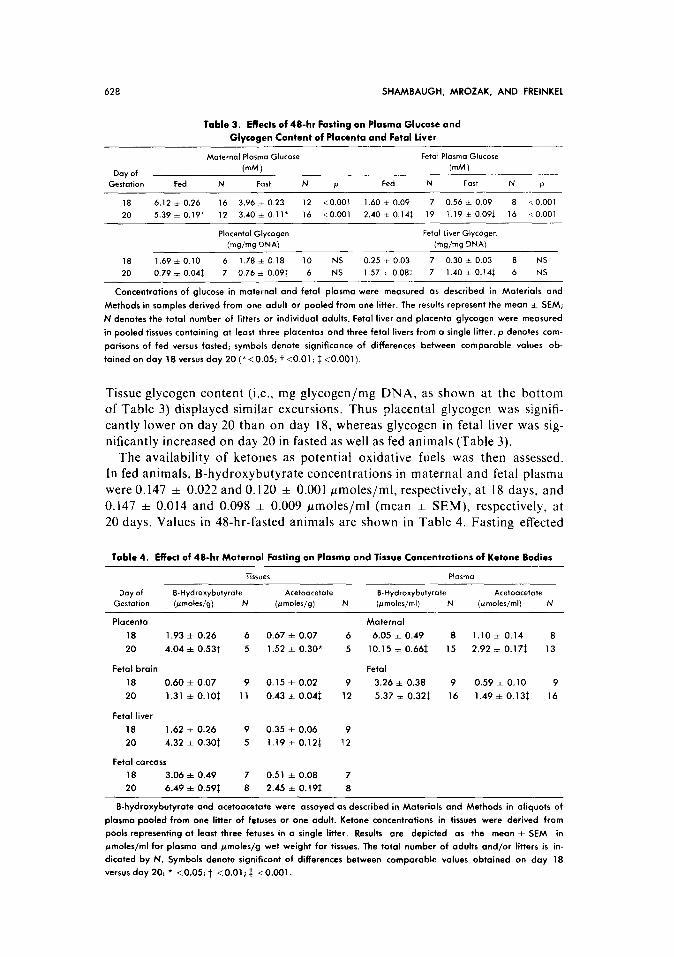
Table 3. Effeclr of 48-hr Fasting on Plasma Glucose and
Glvcogen Content of Placenta and Fetal liver
Maternal Plasma Glucose Fetal Plasma Glucose
Day of (mM) (mM 1 Gestation Fed N Fart N P Fed N Fast N P
18 6.12 i 0.26 16 3.96 zt 0.23 12 <O.OOl 1.60 zt 0.09 7 0.56 i 0.09 8 co.001
20 5.39 * 0.19* 12 3.40 * 0.11* 16 <O.OOl 2.40 zk 0.141 19 1.19 i 0.091 16 <O.OOl
Placental Glvcoaen Fetal Liver Glyco~en
(mgjmg D’NA) -
(w/w DNA)
18 1.69 zt 0.10 6 1.78 zk 0.18 10 NS 0.25 k 0.03 7 0.30 i 0.03 8 NS
20 0.79 * 0.041 7 0.76 + 0.091 6 NS 1.57 k 0.081 7 1.40 i 0.14$ 6 NS
Concentrations of glucose in maternal and fetal plasma were measured OS described in Materials and
Methods in samples derived from one adult or pooled from one litter. The results represent the meon f SEM;
N denotes the total number of litters or individual adults. Fetal liver ond placenta glycogen were measured
in pooled tissues containing at least three placentas and three fetal livers from o single litter. p denotes com-
parisons of fed versus fosted; symbols denote significance of differences between comparable values ob-
tained on day 18 versus day 20 (*<0.05; t <O.Ol; f <O.OOl).
Tissue glycogen content (i.e., mg glycogen/mg DNA. as shown at the bottom of Table 3) displayed similar excursions. Thus placental glycogen was signifi-
cantly lower on day 20 than on day 18, whereas glycogen in fetal liver was sig-
nificantly increased on day 20 in fasted as well as fed animals (Table 3). The availability of ketones as potential oxidative fuels was then assessed.
In fed animals, B-hydroxybutyrate concentrations in maternal and fetal plasma
were 0.147 & 0.022 and 0.120 f 0.001 ~moles/ml, respectively, at 18 days, and 0.147 f 0.014 and 0.098 =t 0.009 ~moles/ml (mean * SEM), respectively, at 20 days. Values in 48-hr-fasted animals are shown in Table 4. Fasting effected
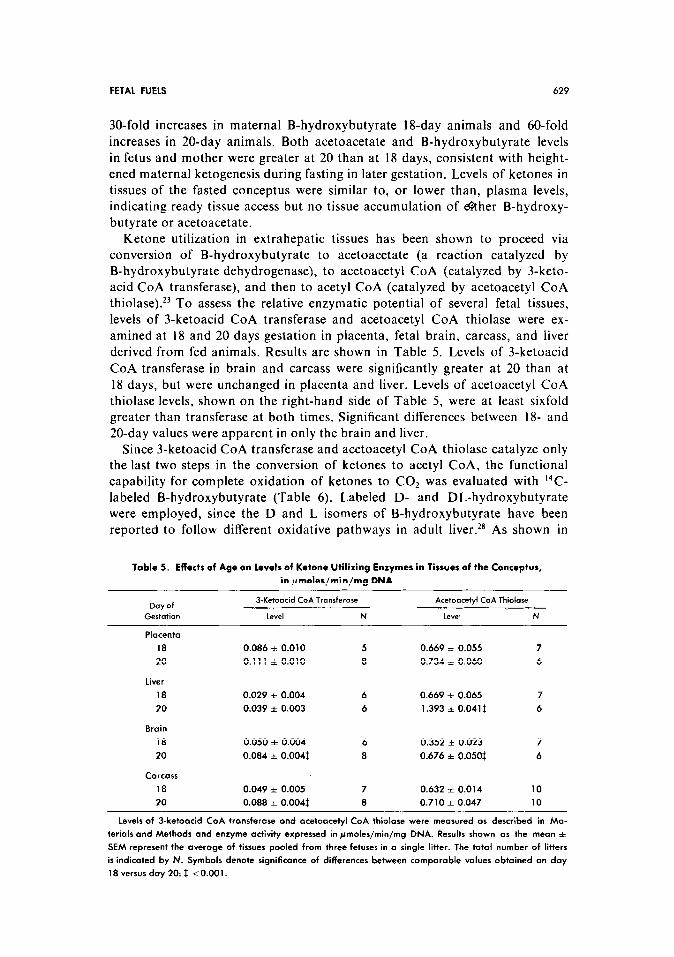
Table 4. Effect of 48-hr Maternal Fasting on Plasma and Tissue Concentrations of Ketone Bodies
Tissues Plasma
Day of B-Hydroxybutyrote Acetoacetate B-Hydroxybutyrote Acetoacetate
Gestation (ymoWg) N (PmoWg) N (~maler/ml) N (~molesjml) N
Placenta Maternal
18 1.93 f 0.26 6 0.67 zt 0.07 6 6.05 f 0.49 8 1.10 * 0.14 8
20 4.04 f 0.53t 5 1.52 * 0.30* 5 10.15 * 0.661 15 2.92 i 0.17$ 13
Fetal brain Fetal
18 0.60 f 0.07 9 0.15 i 0.02 9 3.26 f 0.38 9 0.59 i 0.10 9
20 1.31 f O.lO$ 11 0.43 f 0.041 12 5.37 i 0.321 16 1.49 f 0.13f 16
Fetal liver
18 1.62 i 0.26 9 0.35 + 0.06 9
20 4.32 f 0.30t 5 1.19 l 0.12$ 12
Fetal carcass
18 3.06 f 0.49 7 0.51 f 0.08 7
20 6.49 f 0.59$ 8 2.45 f 0.19i 8
B-hydroxybutyrote and acetoocetote were assayed (11s described in Materials and Methods in oliquots of
plasma pooled from one litter of fetuses or one adult. Ketone concentrations in tissues were derived from
pools representing ot least three fetuses in o single litter. Results ore depicted as the mean f SEM in
pmoles/ml for plasma and pmoles/g wet weight for tissues. The total number of adults and/or litters is in-
dicated by N. Symbols denote significant of differences between comporoble values obtained on day 18
versus day 20: * <0.05; t <O.Ol; $ <O.OOl.
FETAL FUELS 629
30-fold increases in maternal B-hydroxybutyrate 18-day animals and 60-fold
increases in 20-day animals. Both acetoacetate and B-hydroxybutyrate levels in fetus and mother were greater at 20 than at 18 days, consistent with height-
ened maternal ketogenesis during fasting in later gestation. Levels of ketones in
tissues of the fasted conceptus were similar to, or lower than, plasma levels, indicating ready tissue access but no tissue accumulation of &her B-hydroxy-
butyrate or acetoacetate. Ketone utilization in extrahepatic tissues has been shown to proceed via
conversion of B-hydroxybutyrate to acetoacetate (a reaction catalyzed by
B-hydroxybutyrate dehydrogenase), to acetoacetyl CoA (catalyzed by 3-keto- acid CoA transferase), and then to acetyl CoA (catalyzed by acetoacetyl CoA
thiolase). 23 To assess the relative enzymatic potential of several fetal tissues, levels of 3-ketoacid CoA transferase and acetoacetyl CoA thiolase were ex-
amined at 18 and 20 days gestation in placenta, fetal brain, carcass, and liver
derived from fed animals. Results are shown in Table 5. Levels of 3-ketoacid
CoA transferase in brain and carcass were significantly greater at 20 than at
18 days, but were unchanged in placenta and liver. Levels of acetoacetyl CoA
thiolase levels, shown on the right-hand side of Table 5, were at least sixfold greater than transferase at both times. Significant differences between 18- and 20-day values were apparent in only the brain and liver.
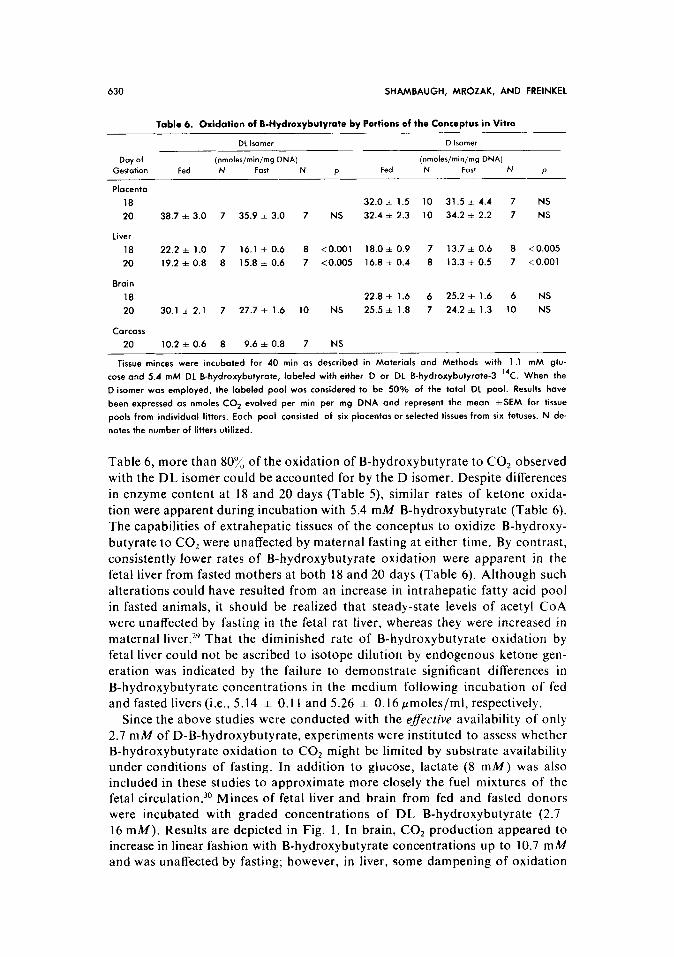
Since 3-ketoacid CoA transferase and acetoacetyl CoA thiolase catalyze only the last two steps in the conversion of ketones to acetyl CoA, the functional
capability for complete oxidation of ketones to CO2 was evaluated with 14C-
labeled B-hydroxybutyrate (Table 6). Labeled D- and DL-hydroxybutyrate were employed, since the D and L isomers of B-hydroxybutyrate have been
reported to follow different oxidative pathways in adult liver.28 As shown in
Table 5. Effects of Age on levels of Ketone Utilizing Enzymes in Tissues of the Conceptus,
in pmoles/min/mg DNA
Day of 3.Ketoocid CoA Transferare Acetoocetyl CoA Thialase
Gestation Level N Level N
Placenta
18 0.086 * 0.010 5 0.669 f 0.055 7
20 0.111 f 0.010 8 0.734 f 0.060 6
Liver
18 0.029 f 0.004 6 0.669 f 0.065 7
20 0.039 f 0.003 6 1.393 * 0.041t 6
Brain
18 0.050 f 0.004 6 0.352 f 0.023 7
20 0.084 f 0.004$ 8 0.676 + 0.0501 6
COWXS
18 0.049 f 0.005 7 0.632 zt 0.014 10
20 0.088 f 0.004$ 8 0.710 * 0.047 10
Levels of 3-ketoacid CoA transferose and ocetoocetyl CoA thiolose were measured os described in Ma-
terials and Methods and enzyme activity expressed in pmoles/min/mg DNA. Results shown as the mean *
SEM represent the avercge of tissues pooled from three fetuses in a single litter. The total number of litters
is indicated by N. Symbols denote significance of differences between comparable values obtained on day
18 versus day 20: 1 < 0.001.
630 SHAMBAUGH, MROZAK, AND FREINKEL
Table 6. Oxidation of B-Hydroxybutyrate by Portions of the Conceptus in Vitro
DL Isomer D Isomer
Day of (nmoler/min/mg DNA) (nmoles/min/mg DNA)
Gestation Fed N Fast N P Fed N Fast N P
PlOCWliO
18 32.0+ 1.5 10 31.5 + 4.4 7 NS
20 38.7 f 3.0 7 35.9 f 3.0 7 NS 32.4 f 2.3 10 34.2 f 2.2 7 NS
Liver
18 22.2 f 1.0 7 16.1 + 0.6 8 <O.OOl 18.Ozk 0.9 7 13.7 * 0.6 8 <0.005
20 19.2~1~0.8 8 15.8 f 0.6 7 <0.005 16.8 f 0.4 8 13.3hO.5 7 <O.OOl
Brain
18 22.8 f 1.6 6 25.2 f 1.6 6 NS
20 30.1 f 2.1 7 27.7 f 1.6 10 NS 25.5 zk 1.8 7 24.2 f 1.3 10 NS
CWCCtSS
20 10.2 f 0.6 8 9.6 f 0.8 7 NS
Tissue minces were incubated for 40 min os described in Materials and Methods with 1 .l mM glu-
cose and 5.4 mM DL B-hydroxybutymte, labeled with either D or DL B-hydroxybutyrote-3 14C. When the
D isomer was employed, the labeled pool was considered to be 50% of the total DL pool. Results have
been expressed OS nmoles CO, evolved per min per mg DNA and represent the mean &SEM for tissue
pools from individual litters. Each pool consisted of six placentas or selected tissues from six fetuses. N de-
notes the number of litters utilized.
Table 6, more than 803/, of the oxidation of B-hydroxybutyrate to CO, observed with the DL isomer could be accounted for by the D isomer. Despite differences
in enzyme content at 18 and 20 days (Table 5) similar rates of ketone oxida-
tion were apparent during incubation with 5.4 mM B-hydroxybutyrate (Table 6). The capabilities of extrahepatic tissues of the conceptus to oxidize B-hydroxy-
butyrate to CO2 were unaffected by maternal fasting at either time. By contrast, consistently lower rates of B-hydroxybutyrate oxidation were apparent in the fetal liver from fasted mothers at both 18 and 20 days (Table 6). Although such
alterations could have resulted from an increase in intrahepatic fatty acid pool in fasted animals, it should be realized that steady-state levels of acetyl CoA were unaffected by fasting in the fetal rat liver, whereas they were increased in
maternal liver.29 That the diminished rate of B-hydroxybutyrate oxidation by fetal liver could not be ascribed to isotope dilution by endogenous ketone gen- eration was indicated by the failure to demonstrate significant differences in
B-hydroxybutyrate concentrations in the medium following incubation of fed and fasted livers (i.e., 5.14 * 0.11 and 5.26 + 0.16 pmoles/ml, respectively.
Since the above studies were conducted with the efective availability of only 2.7 mM of D-B-hydroxybutyrate, experiments were instituted to assess whether B-hydroxybutyrate oxidation to CO2 might be limited by substrate availability under conditions of fasting. In addition to glucose, lactate (8 mM) was also included in these studies to approximate more closely the fuel mixtures of the fetal circulation.30 Minces of fetal liver and brain from fed and fasted donors were incubated with graded concentrations of DL B-hydroxybutyrate (2.7 16 mM). Results are depicted in Fig. 1. In brain, CO2 production appeared to increase in linear fashion with B-hydroxybutyrate concentrations up to 10.7 mA4 and was unaffected by fasting; however, in liver, some dampening of oxidation
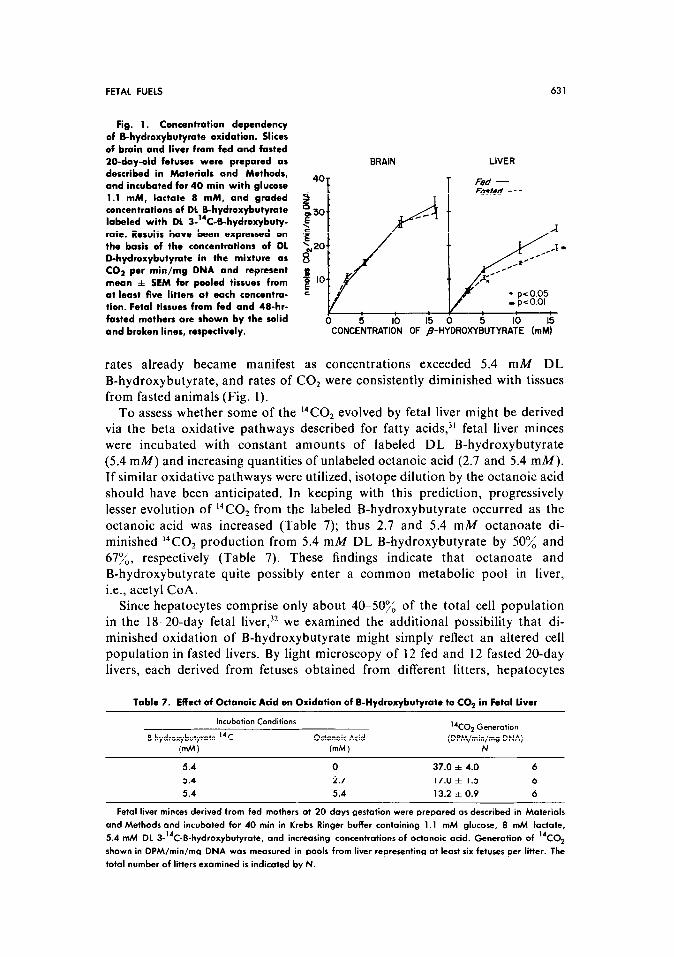
FETAL FUELS 631
Fig. 1. Concentration dependency of B-hydroxybutyrate oxidation. Slices of brain and liver from fed and fasted 20-day-old fetuses were prepared as described in Materials and Methods, and incubated for 40 min with glucose 1.1 mM, lactate 8 mM, and graded concentrations of DL B-hydroxybutyrate labeled with DL 3-“C-B-hydroxybuty- rate. Results have been expressed on the basis of the concentmtions of DL D-hydroxybutyrate in the mixture as CO? per min/mg DNA and represent mean f SEM for pooled tissues from at least five litters at each concentra- tion. Fetal tissues from fed and 4B-hr- fasted mothers are shown by the solid and broken lines, respectively.
rates already became manifest as concentrations exceeded 5.4 mM DL
B-hydroxybutyrate, and rates of CO, were consistently diminished with tissues
from fasted animals (Fig. 1).
BRAIN LIVER
40 T
F& - 1 Fmteo+ ---
,pco.o1
0 5 IO I5 0 5 IO 15 -CONCENTRATION OF $~YDR~XYBUTYRATE (mt$
To assess whether some of the *‘CO2 evolved by fetal liver might be derived via the beta oxidative pathways described for fatty acids,)’ fetal liver minces
were incubated with constant amounts of labeled DL B-hydroxybutyrate
(5.4 mM) and increasing quantities of unlabeled octanoic acid (2.7 and 5.4 mM). If similar oxidative pathways were utilized, isotope dilution by the octanoic acid
should have been anticipated. In keeping with this prediction, progressively lesser evolution of 14C0, from the labeled B-hydroxybutyrate occurred as the
octanoic acid was increased (Table 7); thus 2.7 and 5.4 mM octanoate di-
minished 14C0, production from 5.4 mM DL B-hydroxybutyrate by 50% and 67x, respectively (Table 7). These findings indicate that octanoate and B-hydroxybutyrate quite possibly enter a common metabolic pool in liver,
i.e., acetyl CoA. Since hepatocytes comprise only about 40-50x of the total cell population
in the 18-20-day fetal liver,‘* we examined the additional possibility that di-
minished oxidation of B-hydroxybutyrate might simply reflect an altered cell population in fasted livers. By light microscopy of 12 fed and 12 fasted 20-day livers, each derived from fetuses obtained from different litters, hepatocytes
Table 7. Effect of Octanoic Acid on Oxidation of B-Hydroxybutyrate to CO2 in Fetal liver
Incubation Conditions “CO2 Generation
B-hydroxybutyrate “C Octanoic Acid (DPM/min/mg DNA)
(mm ) (mM) N
5.4 0 37.0 f 4.0 6 5.4 2.7 17.0 f 1.5 6 5.4 5.4 13.2 f 0.9 6
Fetal liver minces derived from fed mothers ot 20 days gestation were prepored OS described in Moteriols
and Methods and incubated for 40 min in Krebs Ringer buffer containing 1 .l mM glucose, 8 mM lactate,
5.4 mh4 DL 3-“C-8-hydroxybutyrate, and increasing concentrations of octanoic acid. Generation of “CO2
shown in DPM/min/mg DNA was measured in pools from liver representing at least six fetuses per litter. The
total number of litters examined is indicated by N.

632 SHAMBAUGH, MROZAK, AND FREINKEL
constituted 41.0:/, & 1.4% of the nucleated cell population in the fed and 41.7% f 1.8% in the fasted groups, a nonsignificant difference.
DISCUSSION
The above investigations have focused on fetal fuel economy. Like others,15s33 we have shown that the reductions in maternal plasma glucose which occur during fasting in late gestation are attended by even greater decrements
in fetal plasma glucose. Neither fetal hepatic nor placental content of glycogen are affected by maternal fasting, although the former appears to increase be-
tween days 18 and 20 of gestation, whereas the latter diminishes. Thus tissue
glycogen stores may be affected by tissue maturation but are seemingly un-
responsive to prevailing energy needs. The failure of these stores to respond to
maternal fasting are consistent with earlier observations34 and indicate that
the fetus must depend on other endogenous reserves when challenges of starva- tion supervene. Although strong evidence for some gluconeogenesis by fetal rat liver has been adduced,35 the manifest hypoglycemia in fasted fetal rats indicates
that this potentiality may be limited. Thus to sustain energy requirements during
fasting the fetus may be required to rely on fuels other than glucose. In late gestation, 48-hr fasting decreases concentrations of both lactate and amino
acids in the mother,’ and despite increases in maternal FFA, transplacental
passage may be limited36 and steady-state concentrations of fatty acids in the
fetal circulation remain unchanged. I5 By contrast, high levels of B-hydroxy-
butyrate and acetoacetate are observed in fasted mothers and fetuses at both
18 and 20 days. The maternal rat late in gestation develops progressive mobili- zation of fat stores,’ resulting in a threefold rise in plasma free fatty acids even in fed animals between days 16 and 21. 37~3x Heightened lipolysis is even more
pronounced in the fasted state,’ and this lipolytic propensity in late gestation
has now been confirmed with isolated human adipose tissue.? Since rates of ketogenesis in the adult are dependent on substrate concentrations, the levels
of B-hydroxybutyrate greater at 20 than at 18 days may reflect greater rates of recall of stored maternal fatty acids on day 20, although the possibility of con-
currently reinforcing changes in the intrahepatic ketogenic “set”39 remains to
be investigated. In any event, dietary deprivation in late pregnancy is attended by supranormal increases in circulating ketones in the rat: comparable findings
during late gestation have been encountered in some6 but not alla” studies of human gravidas. These ketones have been shown to cross the placenta readily,‘,” thereby providing abundant substrates for potential fetal utilization as alterna-
tive fuels during maternal fasting. The present studies have indicated that this potentially may be fulfilled.
In extrahepatic tissues of the conceptus, the conversion of B-hydroxybuty- rate to acetyl CoA, a utilizable fuel, proceeds by three enzymatically controlled
steps. 23 Initially, B-hydroxybutyrate is converted to acetoacetate, a reaction catalyzed by B-hydroxybutyrate dehydrogenase. The acetoacetate generated then combines with succinyl CoA in the presence of 3-ketoacid CoA transferase to form acetoacetyl CoA, and this compound is converted to acetyl CoA via the thiolase reaction. Our findings of abundant 3-ketoacid CoA transferase and acetoacetyl CoA thiolase activities at 18 and 20 days gestation in placenta,

FETAL FUELS 633
brain, liver, and carcass parallel reported developmental changes in B-hydroxy-
butyrate dehydrogenase levels in fetal brain’5541 and acetoacetyl CoA thiolase
levels in fetal brain and liver. I4 Thus enzymes catalyzing all steps of ketone oxidation are present in meaningful amounts during the period of greatest
growth of the conceptus. Indeed, in some instances, the activities expressed per
unit cell mass (i.e., DNA), as in the present studies, actually appear to be
greater on day 20 than on day 18 of gestation (see Table 5). In this regard, it should be noted that the previously reported fall of placental acetoacetyl CoA
thiolase with advancing placental agel does not appear to be significant when measurements are expressed on the basis of DNA and thereby corrected for the
increase in size of the placental cells that occurs between days 18 and 20.42
However, that enzyme activity per se need not be the major determinant of ketone utilization has been readily demonstrated by our experiences with
B-hydroxybutyrate oxidation in cellular systems. Despite activities for some of
the enzymes lower on day 18 than on day 20, we have observed that the evolu-
tion of 14C02 from 5.4 mM DL-3-‘4C-B-hydroxybutyrate during incubation
of minces of placenta, liver, brain, and carcass is not significantly different with tissues from 18- versus 20-day fetuses. We have also observed that the evolu- tion of 14C02 appears to depend upon the concentration of extracellular ketones during additional studies with fetal brain and liver in which the extracellular
concentration of B-hydroxybutyrate was varied from 2.7 to 16 mM. Thus al- though minces preclude rigorous estimations of tissue permeability, it would
appear that ketone oxidation during late fetal life, as in the neonate43 and the
adult,23 may be delineated more by the availability of substrate than by the
absolute enzymatic potential. Our finding of ketones in tissue extracts also sug- gest that penetrance (even for brain) may not be rate limiting, although ad-
herence of extracellular fluid cannot be excluded. The liver has been recognized traditionally as an exporter of ketones, and
the absence of intrahepatic 3-ketoacid CoA transferase (required to convert
acetoacetate to acetoacetyl CoA) in adult liver has been viewed as consistent with the inability of liver to utilize ketones as oxidative fuels. However, in the present studies with fetal liver, we observed not only 3-ketoacid CoA trans- ferase but also unequivocal formation of 14C02 from 3-C’4-B-hydroxybutyrate.
This variance with findings reported for the adult may be related in part to the large contribution of hematopoietic tissue (i.e., 60%) in the 20-day fetal liver
and the presence of 3-ketoacid CoA transferase activity in this cell population. Additional possibilities warrant consideration. More than two decades ago it
was shown that D-B-hydroxybutyrate, unlike acetoacetate, could be oxidized to citrate by adult liver if an ATP-generating system was present.28 Others had demonstrated that the enzyme converting fatty acids to CoA derivatives was
capable of activating fatty acids containing 4414 carbons.44 The subsequent reports that either D or L B-hydroxybutyryl CoA can be oxidized to aceto- acetyl COAST and that the fatty acid converting enzyme can convert B-hydroxy- butyrate to B-hydroxybutyryl CoA3’s4’j could provide an additional biochemical framework for our observations. It remains to be demonstrated whether all these adult potentialities for ketone utilization are operative in fetal liver: appropriate studies to explore these potentialities in fetal liver are presently underway.

634 SHAMBAUGH, MROZAK, AND FREINKEL
Nonetheless, it is conceivable that some of the increase in fetal liver glycogen
between days 18 and 20, which in our hands and from previous reports’” ap- pears to be unaffected by maternal fasting, may reflect some “sparing” of fetal
glucose” via the increased availability of oxidizable ketones rather than activa- tion of fetal gluconeogenesis alone. On the other hand, the lesser ability of fetal
liver, as compared to other fetal structures, to sustain near-normal development during maternal dietary deprivation could reflect, at least in part, a lower ca-
pacity for sustaining oxidative needs via the substitution of maternal ketones as
alternate fuels.
ACKNOWLEDGMENT
The authors gratefully acknowledge the assistance of Dr. Hector Battifora and his staff in the
preparation and interpretation of fetal liver sections. the technical support of Ronald Koehler
and Elizabeth Jackson, and the patient secretarial support of Pat Sopcich.
REFERENCES
I. Knopp RH, Herrera E, Freinkel N: Car-
bohydrate metabolism in pregnancy. VIII.
Metabolism of adipose tissue isolated from fed
and fasted pregnant rats during late gestation.
J Clin Invest 49:143881446, 1970
2. Elliott JA: The effect of pregnancy on the
control of lipolysis in fat cells isolated from
human adipose tissue. Eur J Clin Invest 5:159-
163, 1975
3. Mackay EM, Barnes RH: Fasting ketosis
in the pregnant rat as influenced by adrenal-
ectomy. Proc Sot Exp Biol Med 34:682-683.
1936
4. Scow RO, Chernick SS, Brinley MS: Hyperlipemia and ketosis in the pregnant rat.
Am J Physiol206:796-804, 1964
5. Herrera E, Knopp RH, Freinkel N:
Carbohydrate metabolism in pregnancy. VI.
Plasma fuels, insulin, liver composition, gluco-
neogenesis, and nitrogen metabolism during
late gestation in the fed and fasted rat. J Clin
Invest 48:2260-2272, 1969
6. Williamson DH: Regulation of the utiliza-
tion of glucose and ketone bodies by brain in
the perinatal period in Camerini-Davalos RA,
Cole HS (eds): Early Diab Early Life. New
York, Academic, 1975, pp 195.-202
7. Metzger BE, Hare JW. Freinkel N: Carbohydrate metabolism in pregnancy. IX.
Plasma levels of gluconeogenic fuels during fasting in the rat. J Clin Endocrinol 33:869-
873. 1971
8. Felig P, Kim YJ, Lynch V, et al: Amino acid metabolism during starvation in human
pregnancy. J Clin Invest 51:1195-1202, 1972
9. Scow RO, Chernick SS, Smith BB: Ketosis
in the rat fetus. Proc Sot Exp Med 98:833-835,
1958
IO. Felig P, Lynch V: Starvation in human
pregnancy: hypoglycaemia, hypoinsulinaemia,
and hyperketonaemia. Science 170:990-992.
1970
II. Girard JR, Cruendet GS, Marliss EB,
et al: Fuels, hormone, and liver metabolism at
term and during the early postnatal period in
the rat. J Clin Invest 52:3190-3200, 1973
12. Herrera E, Knopp RH, Freinkel N:
Urinary excretion of epinephrine and nor-
epinephrine during fasting in late pregnancy in
the rat. Endocrinology 84:447 450, 1969
13. Freinkel N: Effects of the conceptus on
maternal metabolism during pregnancy, in
Liebel BS. Wrenshall GA (eds): On the Nature
and Treatment of Diabetes. Amsterdam, Ex-
cerpta Medica. 1965, pp 679-691
14. Dierkes-Ventling C, Cone AL: Aceto-
acetyl-coenzyme A thiolase in brain, liver, and
kidney during maturation of the rat. Science
172:380 382. 1971
15. Dahlquist G, Persson U, Persson B: The
activity of D-B-hydroxybutyrate dehydrogenase
in fetal, infant, and adult rat brain and the
influence of starvation. Biol Neonate 20:40 50,
1972
16. Adam PAJ, Raiha N, Rahhala EL. et al:
Oxidation of glucose and D-B-OH-butyrate by the early human foetal brain. Acta Paediatr
Stand 64: 17-24, 1975 17. Shambaugh GE III, Mrozak SC. Metr-
ger BE. et al: Glutamine-dependent carbaml
phosphate synthetase during fetal and neo-
natal life in the rat. Dev Biol 37: 17 I 185, I974
18. Stotsenburg JM: The growth of the fetus
of the albino rat from the thirteenth to the
twenty-second day of gestation. Anat Ret
9:667~682. 1915

FETAL FUELS 635
19. Huggett ASG, Nixon DA: Use of glucose
oxidase peroxidase and 0-dianisidine in deter-
mination of blood and urinary glucose. Lancet
2:368-370, 1957
20. Mellanby J, Williamson DH: Substances
involved in the metabolism of fatty acids,
lipids, and steroids: Acetoacetate, in Berg-
meyer HU (ed): Methods of Enzymatic Anal-
ysis. New York, Academic, 1965. p 454
21. Williamson DH, Mellanby J: Substances
involved in the metabolism of fatty acids,
lipids, and steroids: D-(-)-B-Hydroxybutyrate,
in Bergman HU (ed): Methods of Enzymatic
Analysis. New York, Academic, 1965, p 459
22. Williamson DH. Bates MW, Krebs HA:
Activity and intracellular distribution of en-
zymes of ketone-body metabolism in rat liver.
Biochem J 108353-361, 1968
23. Williamson DH, Bates MW, Page MA,
et al: Activities of enzymes involved in aceto-
acetate utilization in adult mammalian tissues.
Biochem J 121:41l47, 1971
24. Ide T, Steinke J, Cahill GF Jr: Metabolic
interactions of glucose, lactate, and B-hydroxy-
butyrate in rat brain slices. Am J Physiol 217:
784-792. 1969
25. Good CA, Kramer H, Somogyi M: The determination of glycogen. J Biol Chem 110:
485.491,1933
26. Lowry OH, Rosebrough NJ, Farr AL,
et al: Protein measurement with the folin
phenol reagent. J Biol Chem 193:2655275,
1951
27. Martin RF: Estimation of DNA on tis-
sue. Proc Austr Biochem Sot 3:89, 1970
28. Lehninger AL, Greville CD: The en- zymatic oxidation of d- and I-B-hydroxybuty-
rate. Biochim Biophys Acta 12:188-202, 1953
29. Herrera E, Freinkel N: Metabolites in
the liver, brain, and placenta of fed or fasted
mothers and fetal rats. Horm Metab Res
7~247-249, 1975
30. Shambaugh GE III, Freinkel N: Effects of ketonemia during pregnancy upon fuel homeostasis in the conceptus. Clin Res 24:
504A. 1976
31. McCann WP: The oxidation of ketone
bodies by mitochondria from liver and periph-
eral tissues. J Biol Chem 226: 15-22. 1957
32. Finck W, Theil S: Die embryonale
erythropoeses und der nukleinsauregeha in der
embryonalen leber bei ganzkorperbestrahlten
tarren. Acta Biol Med Ger 12:354-364, 1964
33. Girard JR, Ferre P, Gilbert M, et al:
Stimulation of fetal gluconeogenesis in utero
by maternal fasting near term in the rat. Clin
Res 24:36lA, 1976
34. Goodner CJ, Conway MJ, Werrbach JH:
Relation between plasma glucose levels of
mother and fetus during maternal hypergly-
cemia, hypoglycemia, and fasting in the Rat.
Pediatr Res 3:121-127, 1969
35. Goodner CJ, Thompson DJ: Glucose
metabolism in the fetus in utero: The effect of
maternal fasting and glucose loading in the rat.
Pediatr Res 1:443-451, 1967
36. Koren Z, Shafrin E: Placental transfer of
free fatty acids in the pregnant rat. Proc Sot
Exp Biol Med 116:41 I-416, 1964
37. Knopp RH, Saubek CD, Arky RA, et al:
Two phases of adipose tissue metabolism in
pregnancy: Maternal adaptations for fetal
growth. Endocrinology. 92:894-988, 1973
38. Costrini NV, Kalhoff RK: Relative ef-
fects of pregnancy, estradiol and progesterone
on plasma insulin and pancreatic islet insulin
secretion. J Clin Invest 50:992-999, 1971
39. McGarry JD, Wright PH, Foster DW:
Hormonal control of ketogenesis: Rapid ac-
tivation of hepatic ketogenic capacity in fed
rats by anti-insulin serum and glucagon. J Clin
Invest 55: 1202- 1209, 1975
40. Lunell NO, Persson B, Ohquist G: The
effects of an oral combined contraceptive on
plasma levels of glucose, free fatty acids,
glycerol, D-B-hydroxybutyrate, and triglycer-
ides. Acta Obstet Gynecol Stand 52:23-24,
1973
41. Thaler MM: Effects of starvation on
normal development of B-hydroxybutyrate
dehydrogenase activity in foetal and newborn
rat brain. Nature (New Biol) 236:140-141, 1972
42. Winick M, Noble A: Quantitative changes in ribonucleic acids and protein during
normal growth of rat placenta. Nature 212:34-
35, 1966
43. Bailey E, Lockwood EA: Some aspects of
fatty acid oxidation and ketone body formation
and utilization during development of the rat.
Enzyme 15:239-253, 1973
44. Mahler HR. Wakil SJ: Studies on fatty
oxidation. J Biol Chem 204:453-468, 1953
45. Wakil SJ: D(-)-B-hydroxybutyryl CoA
dehydrogenase. Biochim Biophys Acta 18:314-
315.1955
46. Wakil SJ, Green DE, Mii S, et al: Studies
on the fatty acid oxidizing system of animal
tissues. J Biol Chem 207:631-638, 1954





























