Embed Size (px)
Citation preview

Ž .Brain Research 855 2000 47–57www.elsevier.comrlocaterbres
Research report
Fos induction in selective hypothalamic neuroendocrine and medullary nucleiby intravenous injection of urocortin and corticotropin-releasing factor in rats
Lixin Wang a,b,), Vicente Martınez a,b, Wylie Vale c, Yvette Tache a,b´ ´a CURE: DigestiÕe Diseases Research Center, V.A. Medical Center, Bldg. 115, Rm. 203, 11301 Wilshire BlÕd., Los Angeles, CA 90073, USAb Department of Medicine, DiÕision of DigestiÕe Disease and Brain Research Institute, UniÕersity of California, Los Angeles, CA 90073, USA
c The Clayton Foundation Laboratories for Peptide Biology, The Salk Institute, La Jolla, CA 92037, USA
Accepted 5 October 1999
Abstract
CRF and urocortin, administrated systemically, exert peripheral biological actions which may be mediated by brain pathways. WeŽ .identified brain neuronal activation induced by intravenous i.v. injection of CRF and urocortin in conscious rats by monitoring Fos
Ž .expression 60 min later. Both peptides 850 pmolrkg, i.v. increased the number of Fos immunoreactive cells in the paraventricularnucleus of the hypothalamus, supraoptic nucleus, central amygdala, nucleus tractus solitarius and area postrema compared with vehicleinjection. Urocortin induced a 4-fold increase in the number of Fos-positive cells in the supraoptic nucleus and a 3.4-fold increase in thelateral magnocellular part of the paraventricular nucleus compared with CRF. Urocortin also elicited Fos expression in the accessoryhypothalamic neurosecretory nuclei, ependyma lining the ventricles and choroid plexus which was not observed after CRF. The intensity
Ž .and pattern of the Fos response were dose-related 85, 255 and 850 pmolrkg, i.v. and urocortin was more potent than CRF. Neither CRFnor urocortin induced Fos expression in the lateral septal nucleus, Edinger–Westphal nucleus, dorsal raphe nucleus, locus coeruleus, orhypoglossal nucleus. These results show that urocortin, and less potently CRF, injected into the circulation at picomolar doses activateselective brain nuclei involved in the modulation of autonomicrendocrine function; in addition, urocortin induces a distinct activation ofhypothalamic neuroendocrine neurons. q 2000 Elsevier Science B.V. All rights reserved.
Keywords: Paraventricular nucleus of the hypothalamus; Supraoptic nucleus; Central nucleus of the amygdala; Nucleus tractus solitarius; Area postrema
1. Introduction
Ž .Corticotropin-releasing factor CRF in the brain is animportant integrator of the endocrine, autonomic, immune,
w xbehavioral and visceral responses to stress 16,34,37,54,58 .Growing evidence indicates that peripheral administrationof CRF exerts specific biological actions that may alsohave physiological significance in the modulation of be-havior, pain, immune and gastrointestinal responses to
w xstress 2,7,22,24,27,50,60 . Urocortin is a newly identifiedpeptide cloned from the rat midbrain and a human ge-
w xnomic library which belongs to the CRF family 14,57 .Urocortin induces a similar pattern of biological actions as
Ž . w xCRF when injected intravenously i.v. 5,35,55,57 al-
) Corresponding author. Fax: q1-310-268-4963; e-mail:[email protected]
though its physiological endocrine role is still to be estab-w xlished 29,56 . CRF and urocortin mediate their actions
through activation of specific, seven transmembrane do-Ž .main receptors designed CRF receptor 1 CRF-R1 and 2
Ž .CRF-R2 which were identified through molecular cloningw xfrom distinct genes in rats and humans 9,26,30 . In rats,
CRF-R2 exists in two alternatively spliced forms, CRF-R2a and CRF-R2b which also differ in their amino acidN-terminal extracellular domains and tissue distributionw x13,39 . Existing reports indicate that CRF-R2b mRNA isexpressed mainly in the periphery including the rat heart
w xand gastrointestinal tract 39 , and also in non-neuronalbrain tissues such as the cerebral blood vessels and the
w xchoroid plexus 13 . Based on the greater affinity of uro-cortin for CRF-R2 compared with CRF, urocortin has beensuggested to be the endogenous ligand for this receptor
w xsubtype 57 .Peripheral administration of CRF or urocortin inhibits
w xgastric emptying in rodents and dogs 6,8,35,53,60 through
0006-8993r00r$ - see front matter q 2000 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 99 02200-3

( )L. Wang et al.rBrain Research 855 2000 47–5748
specific CRF receptor interactions which may involvew xCRF-R2 35 . The inhibitory influence of i.v. CRF action
on gastric motor function is partly mediated by hexametho-w xnium sensitive and vagal dependent pathways 8,43 . Taken
together, these observations suggest that peripheral CRFand urocortin may transmit information to the brain andactivate circuits which regulate autonomic and endocrinefunction. However, while there is a wealth of informationon brain neuronal circuitries activated by central injection
w xof CRF 3,4,42,57,59 , it is unknown whether peripheraladministration of CRF or urocortin activates brain path-ways. Therefore, in the present study we used immunohis-tochemical detection of the Fos protein as a measure of
w xneuronal activation 15 to identify brain areas activated byi.v. injection of CRF or urocortin in conscious rats. The
Ž .nucleus tractus solitarius NTS , which is involved in theintegration of somatovisceral and viscerovisceral reflex
w xresponses 61 , and the hypothalamic neurosecretory nucleiŽ .mainly the paraventricular and supraoptic nuclei , whichparticipate in stress-related endocrine and gastric responses
w xto stress 33,49 , were more specifically examined. Otherbrain sites being responsive to central administration ofCRF andror urocortin were also assessed, namely theEdinger–Westphal nucleus, hypoglossal nucleus, dorsal
w xraphe, central amygdala and locus coeruleus 3,4,42,57,59 .
2. Materials and methods
2.1. Animals
ŽAdult male Sprague–Dawley rats Harlan, San Diego,.CA weighing 230–270 g were housed in group cages with
Ž .free access to food Purina Rat Chow and tap water, andmaintained under a 12:12-h light–dark-cycle, and con-
Ž .trolled temperature 21–238C . Animals were fasted, buthad free access to water, for 18–20 h before experimentswhich were conducted under the VA Animal Componentof Research Protocol number 96-080-08.
2.2. Peptides
ŽHumanrrat CRF and rat urocortin Salk Inst., ClaytonFoundation Laboratories for Peptide Biology, La Jolla,
.CA were synthesized and purified as previously describedw x57 . Peptides were kept in lyophilized powder at y708Cand dissolved in sterile saline immediately before theexperiments.
2.3. Treatments
Two to three days before treatment, rats were anes-Žthetized with a mixture of ketamine hydrochloride 75
mgrkg, i.p.; Ketasetw , Fort Dodge Laboratories, Fort
. Ž wDodge, IA and xylazine 5 mgrkg, i.p.; Rompun ,. ŽMobay, Shawnee, KS , and a catheter PE-50 polyethylene
w .tubing, I.D. 0.58 mm, O.D. 0.965 mm, Intramedic waschronically implanted into the right jugular vein. Thecatheter was tunneled subcutaneously, exteriorized in theinterscapular space, and maintained capped. After recoveryfrom anesthesia, rats were housed in individual cages withdirect bedding.
On the day of the experiments, the exteriorized catheterwas connected to the 23-gauge needle of a tuberculinsyringe in animals lightly restrained by hands. Then, vehi-
Ž . Žcle saline, 0.1 mlrrat, ns4 , CRF 85, 255 or 850pmolrkg, i.e., 0.4, 1.2 and 4 mgrkg, ns3–4 in each
. Žgroup or urocortin 85, 255 or 850 pmolrkg, ns4–5 in.each group was injected followed by 0.1 ml of saline to
flush the catheter. Thereafter, animals were placed in theirhome cages for 1 h. The i.v. doses of peptides wereselected based on the demonstration that CRF and uro-
w xcortin at 850 pmolrkg inhibit gastric emptying 35,43 andw xstimulate ACTH release 5,56 with a significant higher
response induced by urocortin compared with CRF. Ineach experiment, vehicle-, CRF- or urocortin-treated ani-mals, at different doses, were included and the experimentwas repeated on multiple days to reach 3–5 ratsrgroup.
2.4. Fos immunohistochemistry
Fos-like immunoreactivity was monitored 1 h after thei.v. administration of vehicle or peptide, essentially as
w xpreviously reported 59 . Animals were deeply anesthetizedŽ .with sodium pentobarbital 0.2–0.3 mlrrat, i.v.
Ž w .Nembutal , Abbott Laboratories, Chicago, IL, USA andŽperfused transcardially with 50 ml of isotonic saline 0.9%
.NaCl followed by 400–500 ml of 4% paraformaldehydeand 14% saturated picric acid in 0.1 M phosphate bufferŽ . Ž .PB pH 7.4 . The brain was then removed, postfixed for4–6 h at 48C in the same fixative and then cryoprotectedovernight in 20% sucrose in 0.1 M PB. Transverse frozen
Ž .sections 30 mm thick were obtained and processed forFos-like immunohistochemistry using the biotin–avidin–peroxidase complex. Free-floating sections were incubated,overnight at 48C, with rabbit polyclonal anti-Fos antibodyŽ .1:4000; Santa Cruz Biotechnology, Santa Cruz, CA as aprimary antibody. Then sections were incubated with bio-
Žtinylated secondary goat anti-rabbit IgG 1:1000; Jackson.ImmunoResearch, West Grove, PA for 1 h at room tem-
perature, followed by the incubation with avidin–biotin–Ž .peroxidase complex 1:100; Vector, Burlingame, CA for 1
h at room temperature. The chromogen was 3,3X-amino-Ž .benzidine tetrachloride 0.025% with hydrogen peroxide
Ž .0.01% . After the staining, sections were mounted, air-dried, dehydrated in ethanol, cleared in xylene and coverslipped. The presence of Fos immunoreactivity was re-vealed as a dark brown precipitate located in the neuronalnuclei.

( )L. Wang et al.rBrain Research 855 2000 47–57 49
Specificity of the antibody was assessed by preabsorp-tion of the Fos antibody with an excess of Fos proteinŽSanta Cruz Biotechnology, 25 mgr1 ml of Fos antibodyat a working dilution of 1:4000; ratio antigen:antibody:
.100:1 . The antigen-antibody mixture was incubated for 24h at 48C and then used as a working primary antibody, asdescribed above.
2.5. Cell counting and statistics
The number of cells containing Fos immunoreactivityŽwas counted in selected nuclei unilaterally except for the
.area postrema in consecutive sections based on Paxinosw xand Watson’s atlas 38 : 10 for the paraventricular nucleusŽ . Žof the hypothalamus PVN around the segment at bregma
.y1.80 and y1.88 mm , 15 for the supraoptic nucleusŽ . ŽSON in the segment between bregma y0.92 and y1.40
.mm , 18 for the area postrema and 20 for the NTScorresponding to the area postrema level and the caudal
Žpart of the opening of the fourth ventricle approximately.from bregma y13.30 to y14.08 mm . Due to different
distribution patterns induced by urocortin and CRF, furthercell counting was performed in three subdivisions of thePVN, namely, the medial and lateral magnocellular andmedial parvocellular parts. Cells were counted in an areaŽ .200=200 mm by a grid placed in the center of eachsubdivision, since the boundaries among these subdivisionsare hardly defined. No correction was done for doublecounting.
Data are expressed as mean"S.E.M., and represent themean number of Fos-positive cellsrsection determinedunilaterally from all sections per rat in all animals pertreatment group. Peptide and dose effects were analyzedby a two-way ANOVA. Differences among groups weredetermined by Kruskal–Wallis nonparametric analysis andDunn’s multiple comparison test. Data were consideredsignificantly different when P-0.05.
3. Results
3.1. Fos immunoreactiÕity in neuroendocrine hypothalamicnuclei
Rats with a chronic intrajugular catheter injected intra-venously with vehicle had a few Fos-positive cells in the
Ž .PVN and SON Figs. 1 and 3A and D . Intravenous CRFŽ .and urocortin 850 pmolrkg increased the number of
Fos-immunoreactive cells in these hypothalamic nuclei, asŽ .determined 60 min after peptide administration Fig. 1 .
Preabsorption of the Fos antibody with an excess of Foscontrol peptide completely abolished Fos immunoreactiv-ity, thus confirming the specificity of the staining.
Ž .In the PVN, CRF 255 and 850 pmolrkg increased FosŽexpression by 1.7 and 2.6 folds 116"17 and 223"12
.Fos-positive cellsrsection respectively over that of vehi-
Fig. 1. Cell counting of Fos-immunoreactive cells induced by i.v. CRFŽ .and urocortin in the hypothalamic nuclei: paraventricular nucleus PVN
Ž .and supraoptic nucleus SON , and in the medulla: nucleus of the solitaryŽ . Ž .tract NTS and area postrema AP . Each column represents the mean"
Ž .S.E.M. of Fos-positive cells per section, unilaterally except AP . Num-bers of animals per treatment are indicated in the columns of the bottomplot. U : P -0.05 vs. vehicle-treated animals; a: P -0.05 vs. the samepeptide at the dose of 85 pmolrkg; q: P -0.05 vs. CRF at the samedose.
Žcle-treated animals 62"11 Fos-positive cellsrsection,. Ž .P-0.001 vs. CRF . The lowest dose of CRF 85 pmolrkg
did not modify significantly the number of cells containingŽFos immunoreactivity in the PVN 85"7 Fos-positive. Ž .cellsrsection, P)0.05 vs. vehicle Fig. 1 . Urocortin
Ž .85, 225 or 850 pmolrkg, i.v. resulted in a 1.5-, 1.6- and4.1-fold increase in the number of Fos-positive cells in the
ŽPVN respectively 157"18, 162"25 and 316"12 Fos-positive cellsrsection; P-0.01 all doses vs. vehicle; and
. Ž .P-0.01 850 pmolrkg vs. the lower doses Fig. 1 .ŽFos-positive cells induced by i.v. injection of CRF 850
.pmolrkg were localized mainly in the medial parvocellu-lar subdivision of the PVN and to a lesser extent also in
Žthe magnocellular part. After urocortin 850 pmolrkg,.i.v. , in addition to the parvocellular part, a robust increase
in Fos expression was found in the medial and lateralŽ .magnocellular subdivisions Figs. 2 and 3E–G . Urocortin
at 85 pmolrkg increased Fos expression in the parvocellu-

( )L. Wang et al.rBrain Research 855 2000 47–5750
Fig. 2. Cell counting of Fos-immunoreactive cells induced by i.v. CRFŽ .and urocortin in some subdivisions of the paraventricular nucleus PVN
in the hypothalamus. Each column represents the mean"S.E.M. ofŽ .Fos-positive cells in a squared counting area as described in Section 2 .
Numbers of animals per treatment are indicated in the columns of the topplot. U : P -0.05 vs. vehicle-treated animals; a: P -0.05 vs. the samepeptide at the dose of 85 pmolrkg; q: P -0.05 vs. CRF at the samedose.
lar, but not significantly in the medial and lateral magno-Ž .cellular subdivisions Fig. 2 . Between CRF and urocortin
at 850 pmolrkg there was no significant difference in theFos expression in the medial parvocellular part, whereas inthe medial and lateral magnocellular parts, the magnitudeof increases was significantly different and more promi-
Ž .nent in the lateral part Fig. 2 .Ž .In the SON, CRF and urocortin 850 pmolrkg, i.v.
Želicited a 4.3-fold 37"3 Fos-positive cellsrsection, P-.0.001 vs. vehicle and a 20-fold increase in Fos immuno-
Žreactivity 147"4 Fos-positive cellsrsection, P-0.001. Ž .vs. vehicle or CRF respectively Figs. 1 and 3A–C
whereas the 85 or 255 pmolrkg dose did not significantlymodify Fos expression, although a trend to increase wasobserved at 255 pmolrkg.
In the accessory hypothalamic neurosecretory nucleiŽnucleus circularis, fornical nuclei and scattered neurons in
.the lateral preoptic area and lateral hypothalamus , numer-ous Fos-containing cells were observed after urocortin
Ž .injected at 850 pmolrkg Fig. 3G–I . By contrast, veryfew cells containing Fos immunoreactivity were found
Ž .after injection of vehicle, CRF at all three doses orŽ .urocortin at 85 and 225 pmolrkg data not shown .
3.2. Fos immunoreactiÕity in the dorsal Õagal complex andarea postrema
Control animals, receiving intravenous saline, showed afew Fos-positive cells in the NTS and almost no labellingwas present in the dorsal motor nucleus of the vagusŽ .DMN . After injection of either CRF or urocortin, Fosimmunoreactivity was localized in the NTS, while none ofthe treatments increase Fos expression in the DMN. Thelowest dose of CRF and urocortin did not significantly
Žincrease Fos expression in the NTS CRF: 33"4; uro-.cortin: 25"1 Fos-positive cellsrsection compared with
Žvehicle 24"2 Fos-positive cellsrsection; P)0.05; Fig.. Ž .1 . CRF and urocortin 850 pmolrkg, i.v. increased the
Žnumber of Fos positive cells in the NTS by 1.5 57"3.Fos-positive cellsrsection; P-0.001 vs. vehicle and 2.3
Žfold 80"2 Fos-positive cellsrsection; P-0.001 vs. ve-. Ž .hicle respectively Figs. 1 and 4A–C . At 225 pmolrkg,
urocortin, unlike CRF, significantly increased by 1.0 foldŽ .Fos expression Fig. 1 . Neuronal activation in the NTS,
after either CRF or urocortin injection, was localizedmainly in the medial subnucleus, scattered cells were alsolocated in the dorsomedial and commissural subnucleus
w xaccording to the nomenclature of Altschuler et al. 1 .Fos immunoreactivity was also increased in the area
Žpostrema in response to CRF 85 and 850 pmolrkg:35"2 and 55"4 Fos-positive cellsrsection respectively
.P-0.05 vs. vehicle, 22"2 Fos-positive cellsrsection orŽurocortin 85, 255 and 850 pmolrkg: 32"2, 35"2, and
109"7 Fos-positive cellsrsection; P-0.05 vs. vehicle;.Figs. 1 and 4A–C . The increase in Fos immunoreactivity
in the area postrema was specially striking after urocortininjection at 850 pmolrkg.
3.3. Fos immunoreactiÕity in other brain nuclei
Ž .Intravenous CRF and urocortin 850 pmolrkg alsoinduced Fos expression, compared with vehicle, in the
Ž .central nucleus of the amygdala CeA and in medullary
Fig. 3. Photomicrographs of representative sections of Fos immunoreactivity in the hypothalamus induced by i.v. injection of vehicle, CRF and urocortinŽ . Ž . Ž .850 pmolrkg . Treatments are indicated in the upright corner H and I are the same as F . Panels A–C show the supraoptic nucleus SON and D–F
Ž . Ž .the paraventricular nucleus PVN . Panels G–I show the accessory neurosecretory nuclei, G and H: the nuclei circularis H is the framed area in G ; I:neurons in the lateral hypothalamus. Abbreviations in D and G indicate the subdivisions of the PVN: DC: dorsal cap; LM: lateral magnocellular;MM: medial parvocellular; MP: medial parvocellular and V: ventral part. Scale barss100 mm. The magnification is the same in A–C, same in D–F andsame in H and I.

( )L. Wang et al.rBrain Research 855 2000 47–57 51

( )L. Wang et al.rBrain Research 855 2000 47–5752
Ž .Fig. 4. Photomicrographs of representative sections of Fos immunoreactivity in the medulla and central nucleus in the amygdala CeA induced by i.v.Ž . Ž . Ž .injection of vehicle, CRF and urocortin 850 pmolrkg . Panels A–C show the nucleus of the solitary tract NTS and area postrema AP , and D–F the
CeA. Treatments are indicated in the upright corner of each panel. Abbreviations: com: commissural, dm: dorsomedial, or med: medial subnucleus of theNTS; st: solitary tract; DMN: dorsal motor nucleus of the vagus; CeAM and CeAL: medial and lateral subnuclei of the CeA. Scale bars100 mm.
catecholaminergic cell groups. Few Fos-positive cells wereobserved in CeA after vehicle injection, while CRF andurocortin induced Fos labeling in this area, with urocortinbeing consistently more effective than CRF. Fos expres-sion in the CeA was observed mainly in the lateral subnu-cleus, only few scattered cells were observed the medial
Ž .subnucleus Fig. 4E, F . In catecholaminergic neurons,
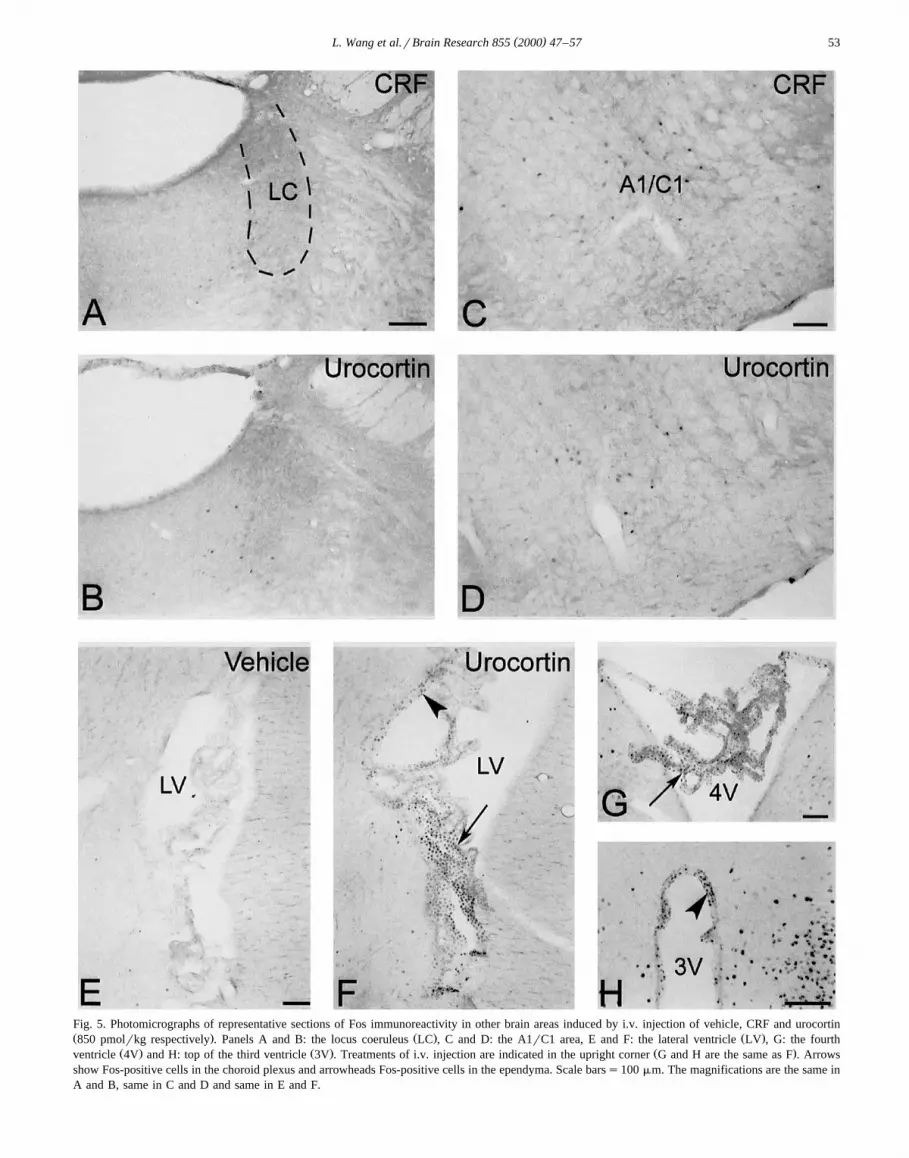
CRF and urocortin induced a similar amount of Fos ex-Ž .pression in the A1rC1 cell group Fig. 5C, D , however,
the response was not robust and there was no activation inŽ . Žthe A6 catecholaminergic group locus coeruleus Fig.
.5A, B .No changes in Fos expression, compared with vehicle
injected animals, were observed in the lateral septal nu-
( )L. Wang et al.rBrain Research 855 2000 47–57 53
Fig. 5. Photomicrographs of representative sections of Fos immunoreactivity in other brain areas induced by i.v. injection of vehicle, CRF and urocortinŽ . Ž . Ž .850 pmolrkg respectively . Panels A and B: the locus coeruleus LC , C and D: the A1rC1 area, E and F: the lateral ventricle LV , G: the fourth
Ž . Ž . Ž .ventricle 4V and H: top of the third ventricle 3V . Treatments of i.v. injection are indicated in the upright corner G and H are the same as F . Arrowsshow Fos-positive cells in the choroid plexus and arrowheads Fos-positive cells in the ependyma. Scale barss100 mm. The magnifications are the same inA and B, same in C and D and same in E and F.

( )L. Wang et al.rBrain Research 855 2000 47–5754
cleus, dorsal raphe nucleus, hypoglossal nucleus, orŽ .Edinger–Westphal nucleus data not shown .
3.4. Fos immunohistochemistry in the Õentricular ependymaand choroid plexus
Cells in the ependyma of brain ventricles and choroidplexus had Fos induction after intravenous injection of
Ž .urocortin at 850 pmolrkg Fig. 5F–H but not after uro-Ž . Žcortin at 85 pmolrkg, CRF all doses and vehicle Fig.
.5E . Cells in the ependyma of the third ventricle wereŽ .mostly observed in the top area Figs. 3 and 5H .
4. Discussion
The present study demonstrates for the first time thatsystemic injection of CRF and urocortin in conscious ratsinduced neuronal activation in discrete brain nuclei asrevealed by the induction of Fos expression 1 h afterpeptide administration. Intravenous injection of CRF orurocortin at 850 pmolrkg increased the number of Fosimmunoreactive cells in the PVN, SON, NTS and areapostrema compared with vehicle injected under the sameconditions. Fos immunoreactivity was also expressed inthe CeA and medullary catecholaminergic cell groups. Theactivation appears to be selective to these nuclei as noincrease in Fos expression was detected in the locuscoeruleus, lateral septal nucleus, raphe nuclei, hypoglossalnucleus and Edingler–Westphal nucleus. This contrast withintracerebroventricular injection of CRF or urocortin at asimilar dose which elicited a robust Fos expression at thesebrain sites in addition to the PVN, NTS, SON and CeAw x3,4,57,59 .
The threshold doses at which urocortin significantlyincreased Fos expression in the PVN and NTS were lowerŽ . Ž85 and 255 pmolrkg respectively than CRF 255 and
.850 pmolrkg respectively . In addition, at the highest dosetested, urocortin induced a significant greater number ofFos immunoreactive cells than CRF in all hypothalamicand medullary sites investigated. These data indicate thaturocortin injected i.v. is more potent than CRF to activateselective brain nuclei. Likewise, urocortin injected i.v. atsimilar doses was reported to be more potent than CRF tostimulate ACTH release, inhibit gastric emptying, and to
w xdecrease edema due to thermal injury in rats 35,55–57 . InCHO cells stably expressing CRF-R1, CRF-R2a or CRF-R2b, the binding affinity of urocortin was about 6, 22 and
w x41 fold higher respectively than rrhCRF 40 which mayaccount for the greater potency of urocortin compared with
w xCRF when injected peripherally 35,55–57 .Whereas the overall distribution of brain nuclei showing
an increase in Fos expression in response to i.v. CRF andurocortin at the highest dose was similar, differences be-tween the peptides were observed with respect to the
delineations of Fos expression within the hypothalamicnuclei. Intravenous urocortin elicits Fos expression in themagnocellular neurons of the PVN, SON and accessorynuclei which constitute the hypothalamic-neurohypo-
w xphyseal system 21 . The localization of Fos-positive neu-Žrons is indicative of staining both classes oxytocin and
. w xvasopressin of magnocellular neurons 52 . By contrast,the numbers of Fos-positive cells were not increased in thelateral magnocellular neurons in the PVN or in the scat-tered populations of cells forming the accessory neurose-cretory nuclei after intravenous CRF at all doses tested. Inaddition, although CRF increases Fos in the SON, thedifference in the magnitude of the Fos induction after
Ž . Žintravenous CRF 4-fold increase and urocortin 20-fold.increase was numerically dominant in this nucleus. The
preferential activation of magnocellular neurosecretoryw xneurons containing vasopressin and oxytocin 21 in re-
sponse to i.v. urocortin, suggests an important role ofperipheral urocortin in the maintenance of fluid balanceand diverse functions regulated by these cell groups.
The parvocellular part of the PVN is activated by bothCRF and urocortin, although there is a lower threshold ofactivation by urocortin as shown by the lack of response toCRF at 85 pmolrkg while urocortin increased significantlyFos expression. The characterization of efferent projectionsfrom activated neurons in the parvocellular part of PVN tobrainstem and spinal cord autonomic nuclei andror the
w xexternal layer of the median eminence 41,48,52 may haveimplications in the mechanisms through which these pep-tides injected peripherally exert endocrine and visceralactions. Of interest is the possibility that the greater stimu-lation of ACTH release induced by intravenous urocortin
w xcompared with CRF 5,56,57 may be related, in additionto its higher affinity to CRF-R1 on pituitary corti-
w xcotrophins than CRF 5,57 , to the enhanced activation oftranscription factors regulating gene expression of peptidesin PVN cells stimulating ACTH secretion.
Another interesting observation was the activation ofthe CeA in response to peripheral administration of CRFand urocortin. This nucleus is involved in several efferentcomponents of the stress response including the endocrine
w xand autonomic components 19,47 . In particular, the acti-vation of the CeA exerts a facilitory role on the hypothala-
w xmic pituitary axis 20,23 and can decrease parasympa-thetic outflow to the heart and stomach through projections
w xto the NTS 19,41 . This suggests possible implications ofthe CeA as part of the circuitry able to modulate theendocrine and visceral responses to CRF and urocortinexerted at the periphery.
Both CRF and, more potently, urocortin, increased Fosexpression in the NTS, mainly in the medial subnucleuswhereas no increase in Fos expression was observed in theDMN. An important component of the visceral informationis carried by vagal afferent pathways that project to the
w xNTS 31,45 . In support for such a peripheral neuralpathways, CRF binding sites are present on the cervical

( )L. Wang et al.rBrain Research 855 2000 47–57 55
and subdiaphragmatic vagus and undergo axonal transportw xsimilar to that observed for CCK receptors 32 . In addi-
tion, electrophysiological studies showed that i.v. adminis-tration of CRF increases gastric vagal afferent discharges
w xin rats 25 . Functional and electrophysiological studiesestablished an inverse relationship between activation ofcommissural and medial NTS neurons and DMN neurons
w xprojecting to the stomach 36,46 . The present resultsprovide neuroanatomical support for the activation of avagovagal inhibitory reflex which may contribute to thedelayed gastric emptying induced by i.v. CRF and uro-cortin at 850 pmolrkg and the greater potency of urocortin
w xto induce gastric stasis 35 .With one exception, all the doses tested of both pep-
tides injected i.v. significantly increased Fos in the areapostrema. These data suggest that peripherally adminis-tered CRF and urocortin may also influence neuronalactivity through humoral mechanisms. Again, urocortinwas more potent than CRF, especially at the dose of 850
w xpmolrkg. The CRF-R2b is present in brain arterioles 13and the area postrema is a circumventricular organ withspecialized vasculature outside of the blood–brain barrierw x12 . This allows the blood-borne signals to be relayed tothe brain via neural efferents projecting from the area
w xpostrema to the NTS 11,51 . The main targets of theseprojections are the commissural, medial and dorsal subnu-
w xclei of the NTS 11 which showed increased Fos expres-sion in response to i.v. injection of CRF and urocortin.Therefore, the area postrema-NTS pathways may be aswell implicated in Fos activation in these subnuclei.
Of notice was the observation that i.v. urocortin inducedFos expression in ependymal cells lining the brain ventri-cles and choroid plexus cells where CRF-R2b are locatedw x13 . No Fos-positive cells were observed after CRF ad-ministration, consistent with CRF low affinity to CRF-R2b
w xin rats 57 . In addition, studies in mice showed that CRFw xis not transported from the blood to the brain 28 which
do not support a direct action of the peptide on the brain,except in areas outside of the blood–brain barrier such asthe area postrema.
Changes in visceral function have been shown to inducew xFos expression in the brain 10,18 . CRF, at the doses
w xused, does not alter blood pressure 17,44 while urocortinw xelicits a hypotensive response 57 . Although normov-
olemic hypotension induces Fos expression in the brain,the pattern of changes induced by i.v. urocortin is notoverlapping with the one induced by hypotensive drugsparticularly as it relates to Fos expression in the AP, locus
w xcoeruleus and lateral hypothalamus 10,18 . Whether theincrease in hypothalamic magnocellular activity after intra-venous urocortin at the highest dose may result frombaroreceptor input associated to cardiovascular changesdeserves further investigation.
In summary, urocortin injected i.v. at picomolar dosesinduced, via neural andror humoral mechanisms, yet to beestablished, a robust activation of magnocellular neurose-
cretory neurons in the PVN, SON, and accessory neurose-cretory nuclei of the hypothalamus. Peripheral urocortinalso activates sites known to be enriched in CRF-R2b suchas the choroid plexus, as well as in other brain sitesinvolved in autonomicrendocrine regulation namely thearea postrema, NTS, CeA and parvocellular PVN. Intra-venous CRF at the same doses yielded a pattern of Fosinduction similar to urocortin, except in accessory neurose-cretory neurons and choroid plexus where the responsewas lacking. Moreover, CRF was less potent than uro-cortin. These activated selective brain nuclei are in posi-tion to participate in neuroendocrine and autonomic adap-tive reflex mechanisms and may be implicated in themodulation of the peripheral actions of systemic urocortinand CRF.
Acknowledgements
This work was supported by the National Institute ofDiabetes and Digestive and Kidney Diseases, grants DK
Ž .33061 and DK 41301 Center Grant, Animal Models Core .Dr. V. Martınez’s current address: CEU-San Pablo, Veteri-´nary School, Department of Physiology. 46113-Moncada,Valencia, Spain.
References
w x1 S.M. Altschuler, D.A. Ferenci, R.B. Lynn, R.R. Miselis, Representa-tion of the cecum in the lateral dorsal motor nucleus of the vagusnerve and commissural subnucleus of the nucleus tractus solitarii in
Ž .rat, J. Comp. Neurol. 304 1991 261–274.w x2 T. Ando, J. Rivier, H. Yanaihara, A. Arimura, Peripheral corti-
cotropin-releasing factor mediates the elevation of plasma IL-6 byŽ .immobilization stress in rats, Am. J. Physiol. 275 1998 R1461–
R1467.w x3 L.C. Andreae, J. Herbert, Expression of c-fos in restricted areas of
the basal forebrain and brainstem following single or combinedintraventricular infusions of vasopressin and corticotropin-releasing
Ž .factor, Neuroscience 53 1993 735–748.w x4 F.J.L. Arnold, M. De Lucas Bueno, H. Shiers, D.C. Hancock, G.I.
Evan, J. Herbert, Expression of c-fos in regions of the basal limbicforebrain following intracerebroventricular corticotropin-releasing
Ž .factor in unstressed or stressed male rats, Neuroscience 51 1992377–390.
w x5 K. Asaba, S. Makino, K. Hashimoto, Effect of urocortin on ACTHsecretion from rat anterior pituitary in vitro and in vivo: comparison
Ž .with corticotropin-releasing hormone, Brain Res. 806 1998 95–103.w x6 A. Asakawa, A. Inui, N. Ueno, S. Makino, M.A. Fujino, M. Kasuga,
Urocortin reduces food intake and gastric emptying in lean andŽ .obrob obese mice, Gastroenterology 116 1999 1287–1292.
w x7 E. Barquist, M. Zinner, J. Rivier, Y. Tache, Abdominal surgery-in-´duced delayed gastric emptying in rats: role of CRF and sensory
Ž .neurons, Am. J. Physiol. 262 1992 G616–G620.w x8 M. Broccardo, G. Improta, Pituitary–adrenal and vagus modulation
of sauvagine- and CRF-induced inhibition of gastric emptying inŽ .rats, Eur. J. Pharmacol. 182 1990 357–362.
w x9 D.T. Chalmers, T.W. Lovenberg, D.E. Grigoriadis, D.P. Behan, E.B.De Souza, Corticotropin-releasing factor receptors: from molecular
Ž .biology to drug design, Trends Pharmacol. Sci. 17 1996 166–172.

( )L. Wang et al.rBrain Research 855 2000 47–5756
w x10 R.K. Chan, P.E. Sawchenko, Spatially and temporally differentiatedpatterns of c-fos expression in brainstem catecholaminergic cellgroups induced by cardiovascular challenges in the rat, J. Comp.
Ž .Neurol. 348 1994 433–460.w x11 E.T. Cunningham, R.R. Miselis, P.E. Sawchenko, The relationship
of efferent projections from the area postrema to vagal motor andbrain stem catecholamine-containing cell groups: an axonal transport
Ž .and immunohistochemical study in the rat, Neuroscience 58 1994635–648.
w x12 E.W. Dempsey, Neural and vascular ultrastructure of the areaŽ .postrema in the rat, J. Comp. Neurol. 150 1973 177–199.
w x13 K.D. Dieterich, H. Lehnert, E.B. De Souza, Corticotropin-releasingfactor receptors: an overview, Exp. Clin. Endocrinol. Diabetes 105Ž .1997 65–82.
w x14 C.J. Donaldson, S.W. Sutton, M.H. Perrin, A.Z. Corrigan, K.A.Lewis, J.E. Rivier, J.M. Vaughan, W.W. Vale, Cloning and charac-
Ž .terization of human urocortin, Endocrinology 137 1996 2167–2170.w x15 M. Dragunow, R. Faull, The use of c-fos as a metabolic marker in
Ž .neuronal pathways tracing, J. Neurosci. Methods 29 1989 261–265.w x16 A.J. Dunn, C.W. Berridge, Physiological and behavioral response to
corticotropin-releasing factor administration: is CRF a mediator ofŽ .anxiety or stress responses?, Brain Res. Rev. 15 1990 71–100.
w x17 L. Fisher, C. Rivier, J. Rivier, M. Brown, Differential antagonistactivity of a-helical CRF in three bioassay systems, Endocrinol-9-41
Ž .ogy 129 1991 1312–1316.w x18 J.C. Graham, G.E. Hoffman, A.F. Sved, c-Fos expression in brain in
response to hypotension and hypertension in conscious rats, J.Ž .Auton. Nerv. Syst. 55 1995 92–104.
w x19 T.S. Gray, E.W. Bingaman, The amygdala: corticotropin-releasingŽ .factor, steroids, and stress, Crit. Rev. Neurobiol. 10 1996 155–168.
w x20 T.S. Gray, M.E. Carney, D.J. Magnuson, Direct projections from thecentral amygdaloid nucleus to the hypothalamic paraventricular nu-cleus: possible role in stress-induced adrenocorticotropin release,
Ž .Neuroendocrinology 50 1989 433–446.w x21 G.I. Hatton, Emerging concepts of structure-function dynamics in
adult brain: the hypothalamo-neurohypophysial system, Prog. Neu-Ž .robiol. 34 1990 437–504.
w x22 M.B. Hennessy, S.M. McInturf, S.J. Mazzei, Evidence that endoge-nous corticotropin-releasing factor suppresses behavioral responsesof guinea pig pups to brief isolation in novel surroundings, Dev.
Ž .Psychobiol. 31 1997 39–47.w x23 J.P. Herman, W.E. Cullinan, Neurocircuitry of stress: central control
of the hypothalamo-pituitary-adrenocortical axis, Trends Neurosci.Ž .20 1997 78–84.
w x24 A. Kita, K. Imano, H. Nakamura, Involvement of corticotropin-re-leasing factor in the antinociception produced by interleukin-1 in
Ž .mice, Eur. J. Pharmacol. 237 1993 317–322.w x25 H.P. Kosoyan, J.Y. Wei, Y. Tache, Corticotropin-releasing factor´
Ž .CRF and modulation of vagal activity, Neurogastroenterology andŽ . Ž .motility 10 1998 365, Abstract .
w x26 T.W. Lovenberg, C.W. Liaw, D.E. Grigoriadis, W. Clevenger, D.T.Chalmers, E.B. De Souza, T. Oltersdorf, Cloning and characteriza-tion of a functionally distinct corticotropin-releasing factor receptor
Ž .subtype from rat brain, Proc. Natl. Acad. Sci. U.S.A. 92 1995836–840.
w x27 V. Martinez, C.L. Rivier, Y. Tache, Peripheral injection of a new´Ž .corticotropin-releasing factor CRF antagonist, astressin, blocks
peripheral CRF- and abdominal surgery-induced delayed gastricŽ .emptying in rats, J. Pharmacol. Exp. Ther. 290 1999 629–634.
w x28 J.M. Martins, A.J. Kastin, W.A. Banks, Unidirectional specific andmodulated brain to blood transport of corticotropin-releasing hor-
Ž .mone, Neuroendocrinology 63 1996 338–348.w x29 M. Masuzawa, Y. Oki, M. Ozawa, F. Watanabe, T. Yoshimi,
Corticotropin-releasing factor but not urocortin is involved inadrenalectomy-induced adrenocorticotropin release, J. Neuroen-
Ž .docrinol. 11 1999 71–74.w x30 J.R. McCarthy, Recent advances with the CRF1 receptor: design of
small molecule inhibitors, receptor subtypes and clinical indications,Ž .Curr. Pharm. Des. 5 1999 289–315.
w x31 D. Menetrey, A.I. Basbaum, Spinal and trigeminal projections to the´nucleus of the solitary tract: a possible substrate for somatovisceral
Ž .reflex activation, J. Comp. Neurol. 255 1987 439–450.w x32 J.G. Mercer, C.B. Lawrence, P.A. Copeland, Corticotropin-releasing
factor binding sites undergo axonal transport in the rat vagus nerve,Ž .J. Neuroendocrinol. 4 1992 281–285.
w x33 H. Monnikes, B.G. Schmidt, H.E. Raybould, Y. Tache, CRF in theparaventricular nucleus mediates gastric and colonic motor response
Ž .to restraint stress, Am. J. Physiol. 262 1992 G137–G143.w x34 A. Morimoto, T. Nakamori, K. Morimoto, N. Tan, N. Murakami,
Ž .The role of corticotrophin-releasing factor CRF-41 in psychologi-Ž . Ž .cal stress in rats, J. Physiol. London 460 1993 221–229.
w x35 T. Nozu, V. Martinez, J. Rivier, Y. Tache, Peripheral urocortindelays gastric emptying: role of CRF receptor 2, Am. J. Physiol. 276Ž .1999 867–875.
w x36 B.R. Olson, M. Freilino, G.E. Hoffman, E.M. Stricker, A.F. Sved,J.G. Verbalis, c-Fos expression in rat brain and brainstem nuclei inresponse to treatments that alter food intake and gastric motility,
Ž .Mol. Cell. Neurosci. 4 1993 93–106.w x37 M.J. Owens, C.B. Nemeroff, Physiology and pharmacology of corti-
Ž .cotropin-releasing factor, Pharmacol. Rev. 43 1991 425–473.w x38 G. Paxinos, C. Watson, The rat brain in stereotaxic coordinates,
Academic Press, Orlando, 1998, pp. 1–116.w x39 M. Perrin, C. Donaldson, R. Chen, A. Blount, T. Berggren, L.
Bilezikjian, P. Sawchenko, W. Vale, Identification of a secondcorticotropin-releasing factor receptor gene and characterization of a
Ž .cDNA expressed in heart, Proc. Natl. Acad. Sci. U.S.A. 92 19952969–2973.
w x40 M.H. Perrin, S.W. Sutton, L.A. Cervini, J.E. Rivier, W.W. Vale,Comparison of an agonist, urocortin, and an antagonist, astressin, asradioligands for characterization of CRF receptors, J. Pharmacol.
Ž .Exp. Ther. 288 1999 729–734.w x41 T. Petrov, T.L. Krukoff, J.H. Jhamandas, Convergent influence of
the central nucleus of the amygdala and the paraventricular hypotha-lamic nucleus upon brainstem autonomic neurons as revealed by
Ž .c-fos expression and anatomical tracing, J. Neurosci. Res. 42 1995835–845.
w x42 S. Rassnick, G.E. Hoffman, B.S. Rabin, A.F. Sved, Injection ofcorticotropin-releasing hormone into the locus coeruleus or foot
Ž .shock increases neuronal Fos expression, Neuroscience 85 1998259–268.
w x43 H.E. Raybould, C.B. Koelbel, E.A. Mayer, Y. Tache, Inhibition of´gastric motor function by circulating corticotropin-releasing factor in
Ž .anesthetized rats, J. Gastrointest. Motil. 2 1990 265–272.w x44 R.M. Richter, M.J. Mulvany, Comparison of hCRF and oCRF
effects on cardiovascular responses after central, peripheral, and inŽ .vitro application, Peptides 16 1995 843–849.
w x45 L. Rinaman, J.P. Card, J.S. Schwaber, R.R. Miselis, Ultrastructuraldemonstration of a gastric monosynaptic vagal circuit in the nucleus
Ž .of the solitary tract in rat, J. Neurosci. 9 1989 1985–1996.w x46 R.C. Rogers, M.J. McCann, Intramedullary connections of the gas-
tric region in the solitary nucleus: a biocytin histochemical tracingŽ .study in the rat, J. Auton. Nerv. Syst. 42 1993 119–130.
w x47 B. Roozendaal, J.M. Koolhaas, B. Bohus, The role of the centralamygdala in stress and adaption, Acta Physiol. Scand. Suppl. 640Ž .1997 51–54.
w x48 P.E. Sawchenko, Evidence for differential regulation of corti-cotropin-releasing factor and vasopressin immunoreactivities in par-vocellular neurosecretory and autonomic-related projections of the
Ž .paraventricular nucleus, Brain Res. 437 1987 253–263.w x49 P.E. Sawchenko, E.R. Brown, R.K. Chan, A. Ericsson, H.Y. Li,
B.L. Roland, K.J. Kovacs, The paraventricular nucleus of the hypo-thalamus and the functional neuroanatomy of visceromotor re-
Ž .sponses to stress, Prog. Brain Res. 107 1996 201–222.w x50 M. Schafer, S.A. Mousa, Q. Zhang, L. Carter, C. Stein, Expression

( )L. Wang et al.rBrain Research 855 2000 47–57 57
of corticotropin-releasing factor in inflamed tissue is required forintrinsic peripheral opioid analgesia, Proc. Natl. Acad. Sci. U.S.A.
Ž .93 1996 6096–6100.w x51 R.E. Shapiro, R.R. Miselis, The central neural connections of the
Ž .area postrema of the rat, J. Comp. Neurol. 234 1985 344–364.w x52 L.W. Swanson, P.E. Sawchenko, Hypothalamic integration: organi-
zation of the paraventricular and supraoptic nuclei, Ann. Rev.Ž .Neurosci. 6 1983 269–324.
w x53 Y. Tache, M. Maeda-Hagiwara, C.M. Turkelson, Central nervoussystem action of corticotropin-releasing factor to inhibit gastric
Ž .emptying in rats, Am. J. Physiol. 253 1987 G241–G245.w x Ž .54 A.V. Turnbull, C. Rivier, Corticotropin-releasing factor CRF and
endocrine response to stress: CRF receptors, binding protein, andŽ .related peptides, Proc. Soc. Exp. Biol. Med. 215 1997 1–10.
w x55 A.V. Turnbull, W. Vale, C. Rivier, Urocortin, a corticotropin-releas-ing factor-related mammalian peptide, inhibits edema due to thermal
Ž .injury in rats, Eur. J. Pharmacol. 303 1996 213–216.w x56 A.V. Turnbull, J. Vaughan, J.E. Rivier, W.W. Vale, C. Rivier,
Urocortin is not a significant regulator of intermittent electrofoot-
shock-induced adrenocorticotropin secretion in the intact male rat,Ž .Endocrinology 140 1999 71–78.
w x57 J. Vaughan, C. Donaldson, J. Bittencourt, M.H. Perrin, K. Lewis, S.Sutton, R. Chan, A.V. Turnbull, D. Lovejoy, C. Rivier, J. Rivier,P.E. Sawchenko, W. Vale, Urocortin, a mammalian neuropeptiderelated to fish urotensin I and to corticotropin-releasing factor,
Ž .Nature 378 1996 287–292.w x58 M. Venihaki, J.A. Majzoub, Animal models of CRH deficiency,
Ž .Front Neuroendocrinol. 20 1999 122–145.w x59 L. Wang, S. Cardin, V. Martinez, Y. Tache, Intracerebroventricular´
CRF inhibits cold restraint-induced c-fos expression in the dorsalmotor nucleus of the vagus and gastric erosions in rats, Brain Res.
Ž .736 1996 44–53.w x60 C.L. Williams, J.M. Peterson, R.G. Villar, T.F. Burks, Corti-
cotropin-releasing factor directly mediates colonic responses to stress,Ž .Am. J. Physiol. 253 1987 G582–G586.
w x61 B.J. Yates, S.D. Stocker, Integration of somatic and visceral inputsby the brainstem: functional considerations, Exp. Brain Res. 119Ž .1998 269–275.





























