Embed Size (px)
Citation preview

Rapid Publication
Urocortin Expression in Rat Brain:Evidence Against a Pervasive
Relationship of Urocortin-ContainingProjections With Targets Bearing
Type 2 CRF Receptors
JACKSON C. BITTENCOURT,1 JOAN VAUGHAN,2 CARLOS ARIAS,1
ROBERT A. RISSMAN,1 WYLIE W. VALE,2 AND PAUL E. SAWCHENKO1*1Laboratory of Neuronal Structure and Function, The Salk Institute for Biological Studies
and Foundation for Medical Research, La Jolla, California 920372Peptide Biology Laboratory, The Salk Institute for Biological Studies and Foundation
for Medical Research, La Jolla, California 92037
ABSTRACTHistochemical and axonal transport methods were used to clarify the central organization
of cells and fibers that express urocortin (UCN), a recently discovered corticotropin-releasingfactor (CRF)-related neuropeptide, which has been proposed as an endogenous ligand for type2 CRF receptors (CRF-R2). Neurons that display both UCN mRNA and peptide expressionwere found to be centered in the Edinger-Westphal (EW), lateral superior olivary (LSO), andsupraoptic nuclei; lower levels of expression are seen in certain cranial nerve and spinalmotoneurons and in small populations of neurons in the forebrain. Additional sites of UCNmRNA and peptide expression detected only in colchicine-treated rats are considered to beminor ones. UCN-immunoreactive projections in brain are predominantly descending andlargely consistent with central projections attributed to the EW and LSO, targetingprincipally accessory optic, precerebellar, and auditory structures, as well as the spinalintermediate gray. Although neither the EW nor LSO are known to project to the forebrain,UCN-ir neurons in the EW were identified that project to the lateral septal nucleus, whichhouses a prominent UCN-ir terminal field. Although substantial UCN-ir projections wereobserved to several brainstem cell groups that express CRF-R2, including the dorsal rapheand interpeduncular nuclei and the nucleus of the solitary tract (NTS), most prominent seatsof CRF-R2 expression were found to contain inputs immunopositive for piscine urotensin I,but not rat UCN. The results define a central UCN system whose organization suggests aprincipal involvement in motor control and sensorimotor integration; its participation instress-related mechanisms would appear to derive principally by virtue of projections to thespinal intermediolateral column, the NTS, and the paraventricular nucleus. Several observa-tions, including the lack of a pervasive relationship of UCN-ir projections with CRF-R2-
Grant sponsor: NIH; Grant number: DK-26741; Grant sponsor: CNPq;Grant numbers: 203301/87-0 and 300583/91-5.
*Correspondence to: Paul E. Sawchenko, Ph.D., The Salk Institute,10010 N. Torrey Pines Road, La Jolla, CA 92037.E-mail: [email protected].
Received 30 July 1999; Revised 9 September 1999; Accepted 14 Septem-ber 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 415:285–312 (1999)
r 1999 WILEY-LISS, INC.

expressing targets, support the existence of still additional CRF-related peptides in mamma-lian brain. J. Comp. Neurol. 415:285–312, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: autonomic nervous system; corticotropin-releasing factor; Edinger-Westphal
nucleus; neuropeptides; stress
Corticotropin-releasing factor (CRF), a peptide first iso-lated from mammalian brain (Vale et al., 1981), playscritical roles in regulating the pituitary-adrenal axis, andin other stress-related endocrine, autonomic, immune, andbehavioral mechanisms (for reviews, see Chadwick et al.,1993). Structurally related peptides identified in fish (uro-tensin I; Lederis et al., 1982) and frog (sauvagine; Mon-
tecucchi and Henschen, 1981) were originally consideredto be CRF homologs. The subsequent characterization ofpeptides closer in sequence to CRF in piscine and amphib-ian species (Okawara et al., 1988; Stenzel-Poore et al.,1992), however, raised the possibility that an additionalmember of this peptide family may also be represented inmammals. This was reinforced by observed differences in
Abbreviations
III oculomotor nucleusviii acoustico-vestibular nerveVII facial nucleusXII hypoglossal nucleusac anterior commissureAL anterior lobe (pituitary)Amb nucleus ambiguusAmyg amygdalaAP area postremaaq cerebral aqueductARH arcuate nucleus (hypothalamus)BST bed nucleus of the stria terminalisCBL cerebellumcc corpus callosumCLA claustrumcp cerebral peduncleCtx cerebral cortexda dorsal accessory nucleus (inferior olive)DBB nucleus of the diagonal banddc dorsal columnsDCo dorsal cochlear nucleusDH dorsal horn, spinal cordDMX dorsal motor nucleus of the vagus nerveDors. Col. dorsal column nucleiDR dorsal raphe nucleusEP endopiriform nucleusEW Edinger-Westphal nucleusExt. external lamina of the median eminenceFL flocculusfx fornixGP globus pallidusgr granule cell layer (cerebellum)Gr gracile nucleusHF hippocampal formationHYP hypothalamusIC inferior colliculusicp inferior cerebellar peduncleIGL intergeniculate leafletIL intermediate lobe (pituitary gland)IML intermediolateral column, spinal cordINC interstitial nucleus of CajalInt. internal lamina of the median eminenceIO inferior olivary complexIP interpeduncular nucleusL lumenLat. Sept. lateral septumlfu lateral funiculusLHA lateral hypothalamic areaLM longitudinal muscle layerLRN lateral reticular nucleusLSd dorsal lateral septal nucleus, dorsal regionLSi intermediate lateral septal nucleusLSO lateral superior olivary nucleusLSv ventral lateral septal nucleusma medial accessory nucleus (inferior olive)MB mammillary bodymcp middle cerebellar peduncle
MeA medial nucleus amygdalaMed medullaMid Thal midline thalamic nucleimlf medial longitudinal fascicleMM muscularis mucosamol molecular layer (cerebellum)MoV motor nucleus of the trigeminal nerveMR median raphe nucleusMS medial septal nucleusMSO medial superior olivary nucleusmtg mammillotegmental tractMuc mucosaND nucleus of DarkschewitschNTB nucleus of the trapezoid bodyNTS nucleus of the solitary tractoch optic chiasmOlf. Bulb olfactory cortex bulbot optic tractOVLT vascular organ of the lamina terminalisPAG periaqueductal grayPB parabrachial nucleusPG pontine grayPHA posterior hypothalamic areaPL posterior lobe (pituitary)POA preoptic areaPP peripeduncular nucleuspr principal nucleus (inferior olive)Pretect pretectal regionpur Purkinje cell layer (cerebellum)pv periventricular partPVH paraventricular nucleus (hypothalamus)py pyramidal tractRN red nucleusSC superior colliculusSCN suprachiasmatic nucleusSept septal regionSM smooth muscle layerSN substantia nigraSNc substantia nigra, compact partSNr substantia nigra, reticular partSNV spinal trigeminal nucleusSNVc spinal trigeminal nucleus, caudal partSO supraoptic nucleusSp. Cord spinal cordSpV spinal nucleus of the trigeminalstv spinal trigeminal tractSUM supramammillary nucleusSuV superior vestibular nucleusTRN tegmental reticular nucleusts solitary tractUCN urocortinV villiVest vestibular nucleiVH ventral horn, spinal cordVMH ventromedial nucleus (hypothalamus)VTA ventral tegmental area
286 J.C. BITTENCOURT ET AL.

the affinities of the known binding moieties for peptides inthis lineage (see, e.g., Turnbull and Rivier, 1997). A screenof rat brain based on the use of immunologic and molecularprobes for CRF and nonmammalian members of thispeptide family led to the cloning of cDNAs encodingurocortin (UCN), a 40-residue peptide expressed in ratbrain (Vaughan et al., 1995), and the second CRF-relatedpeptide to be identified in mammals.
UCN exhibits sequence homology with rat CRF (45%amino acid identity) and with carp urotensin I (63%) and isbound with high affinity by both type 1 and, particularly,type 2 CRF receptors (CRF-R1 and -R2, respectively), aswell as the CRF-binding protein (Vaughan et al., 1995). Ithas been found to be more potent than CRF itself instimulating adrenocorticotropic hormone (ACTH) releasefrom cultured anterior pituitary cells (Vaughan et al.,1995; Asaba et al., 1998). A good deal of the initial interestin UCN has derived from data to suggest that it maymediate some stress-related functions attributed previ-ously to CRF. For example, centrally administered UCN issubstantially more potent than CRF in suppressing appe-tite, but less so in producing anxiety-like effects andbehavioral activation (Spina et al., 1996). In view of thefact that a number of sites of action at which CRF elicitsstress-related autonomic and behavioral responses appearto lack either CRF-containing neural inputs, CRF recep-tors, or both (see e.g., Herkenham, 1987; Potter et al.,1994), the discovery of UCN and at least two major CRFreceptor subtypes in brain has offered promise for resolv-ing long-standing examples of ligand-receptor mismatchesin the central CRF system.
In the initial characterization of UCN (Vaughan et al.,1995), we described a restricted central distribution ofUCN mRNA that included a dominant seat of cellularexpression in the Edinger-Westphal nucleus (EW) andsecondary ones in the lateral superior olive (LSO) and thesupraoptic nucleus. Low-level expression was also noted incertain cranial nerve motor nuclei. Immunolabelingachieved using antisera raised against fish urotensin Ishowed similar patterns of cellular labeling and alsostained terminal fields in several areas enriched in CRF-R2expression. Coupled with the finding that central adminis-tration of UCN provoked cellular activational responses incell groups that were seen to both receive urotensinimmunoreactive (ir) projections and display CRF-R2 ex-pression, it was suggested that UCN may serve as apreferred endogenous ligand for type 2 CRF receptors(Vaughan et al., 1995). Subsequently, others have usedimmunohistochemical (Hara et al., 1997a,b; Kozicz et al.,1998; Yamamoto et al., 1998) and hybridization histochemi-cal methods (Wong et al., 1996) to confirm some, but notall, of these posited sites of cellular UCN expression and tosuggest a substantially broader (Wong et al., 1996) orsomewhat different distribution (Kozicz et al., 1998), whichincludes such pivotal components of neuroendocrine stress-related mechanisms as the paraventricular nucleus (Ko-zicz et al., 1998) and the anterior pituitary (Wong et al.,1996; Iino et al., 1997). Immunolabeling studies usingUCN-specific antisera have achieved only very limitedsuccess in labeling UCN-ir projections, and knowledge ofthese is critical to placing this peptide system in organiza-tional and functional context.
Here we report on the use of immunohistochemical,hybridization histochemical, and axonal transport meth-
ods to characterize the central UCN system. The majorobjectives were to use independent methods in an attemptto reconcile discrepancies in the reported central cellulardistribution of UCN expression, to delineate the organiza-tion of central UCN-ir projections, to provide a generalcontext for the UCN system as it relates to its capacity tointeract with CRF receptors, and to account for stress-related effects attributed to CRF. Portions of these findingshave been presented in abstract form (Bittencourt et al.,1995).
MATERIALS AND METHODS
Animals
Adult male and female Sprague-Dawley rats weighing250–350 g were used in this study and were housed twoper cage in a colony room maintained on a 12:12 hourlight:dark cycle. The animals had free access to food andwater at all times and were allowed a minimum of 7 days’adaptation to housing conditions prior to any manipula-tion or sacrifice or histology. Some rats received stereotaxi-cally guided intracerebroventricular injections of colchi-cine (50 µg in 25 µl saline) under ketamine/xylazine/acepromazine anesthesia (25:5:1 mg/kg, s.c.) 48–72 hoursprior to sacrifice. All procedures were approved by theInstitutional Animal Care and Use Committee of the SalkInstitute.
Perfusion and histology
Rats were anesthetized with 35% chloral hydrate forperfusion. Initial screens of UCN antisera were carried outon animals perfused via the ascending aorta with 0.9%saline, followed by 500–750 ml 4% paraformaldehyde in0.1 M borate buffer, pH 9.5, delivered over 20 minutes.Brains were postfixed for 0–6 hours and cryoprotectedovernight in 10% sucrose in 0.1 M phosphate-bufferedsaline (pH 7.4), prior to sectioning. Once the optimalanti-UCN serum was identified, a variety of fixationprotocols, postfixation times (0–18 hours), and immunoper-oxidase reaction conditions (see below) were evaluated inan effort to optimize staining for UCN-ir fibers. Thegreatest sensitivity in UCN-ir localization was achieved inanimals perfused as above, but using Zamboni’s fixative,pH 7.4, at 10°C (Zamboni and De Martino, 1967), withoutpostfixation. After cryoprotection, the forebrain, brain-stem, pituitary, and blocks from the cervical, thoracic,lumbar, and sacral regions of the spinal cord were frozenon dry ice and sectioned on a sliding microtome. Multiple1-in-5 or 1-in-10 series of 30-µm-thick frontal sectionsthroughout the brain were collected in ethylene glycol-based cryoprotectant solution, in which they were storedat 20°C until processing for immunoperoxidase demonstra-tion of UCN-ir, or stained with thionin for referencepurposes.
Tissue prepared using the Zamboni fixative was avail-able from 12 nonmanipulated rats and from 8 rats pre-treated with colchicine; additional material was obtainedfrom some 30 animals (12 of which were pretreated withcolchicine) perfused with 4% paraformaldehyde and sub-jected to varying postfixation schedules. Eight representa-tive levels of the spinal cord were sampled in five of theseanimals.
UROCORTIN EXPRESSION IN BRAIN 287

Immunohistochemistry
A conventional avidin-biotin immunoperoxidase method,utilizing reagents available in kit form (Vectastain Elite,Vector Labs, Burlingame, CA) was used. The standardprocedure involved tissue pretreatments with hydrogenperoxide (to quench endogenous peroxidase activity) andsodium borohydride (to reduce free aldehydes); the diami-nobenzidine-based reaction product was generated using anickel-enhanced diaminobenzidine reaction (Shu et al.,1988; see also Sawchenko et al., 1990). Empirically derivedmodifications found to enhance labeling of UCN-ir fibersincluded 1) substitution of 5 mg/ml heparin and 2% bovineserum albumin for normal (blocking) serum in all incuba-tions; and 2) elimination of the nickel enhancement steps.
Antisera and controls
Of six UCN antisera screened, optimal specific labelingwas achieved with one (code 5779), raised in rabbitsagainst a synthetic rat UCN conjugated to human a-globu-lin via glutaraldehyde (Turnbull et al., 1999). For purposesof comparison, similarly prepared antisera raised in rabbitagainst human a-globulin conjugates with rat CRF (coderC-70; Sawchenko, 1987), synthetic frog sauvagine (fromPhyllomedusa sauvagei; code 5678), and synthetic sucker-fish urotensin I (from Catostomus commersoni, code 5712;Vaughan et al., 1995) were used. All were adsorbed againstthe human a-globulin carrier (5 mg/ml serum for 24 hoursat 4°C) prior to use and were applied to tissue at 1:2,000dilutions. The rat CRF antiserum was adsorbed againsta-melanin-stimulating hormone (MSH) to eliminate cross-reactivity with a C-terminal epitope common to CRF,a-MSH and a peptide (neuropeptide EI) derived from themelanin-concentrating hormone precursor (Sawchenko,1987; Nahon et al., 1989). The anti-UCN serum wasaffinity purified against Tyr-Gly-UCN23–40, to eliminatecross-reactivity with N-terminal epitopes shared amongCRF family members (Fig. 1). To evaluate the specificity ofUCN-ir staining, competition studies were carried out thatinvolved comparison of staining patterns and intensitiesachieved using UCN antiserum that was preincubated(overnight at 4°C) with 0–300 µM synthetic UCN, CRF,sauvagine, or urotensin I, generously provided by Dr. JeanRivier (Salk Institute).
Combined retrogradetransport-immunohistochemical experiments
Because the EW is acknowledged to be a major locus ofcellular UCN expression, we sought to determine whetherUCN-ir neurons contribute to documented terminal fieldsof EW in the spinal cord (Saper et al., 1976; Loewy andSaper, 1978), as well as a suggested terminal field in the
lateral septal nucleus (Vaughan et al., 1995; Kozicz et al.,1998).
Under ketamine/xylazine/acepromazine anesthesia, onegroup of 10 animals received stereotaxically guided crystal-line implants of the retrogradely transported fluorescenttracer fast blue (Bentivoglio et al., 1980), aimed at thelateral septal nucleus, and delivered via prefilled glassmicropipettes (100–150 µm i.d.). A second group of 10 ratsreceived injections of 5% aqueous suspensions of fast blueand a second fluorescent tracer, diamidino yellow (Benti-voglio et al., 1980), in the spinal cord. For spinal injections,a hemilaminectomy was performed at T2, and two sepa-rate 0.5-µl injections of tracer were placed between T1 andT2 with the intent of involving an entire hemisection,including the white matter.
Animals were permitted to survive for 7–15 days aftersurgery, at which point they received intraventricularinjections of colchicine, as above. They were then perfused(2–3 days later) and prepared for concurrent fluorescencedemonstration of transported tracer and UCN-ir. Thisinvolved conventional indirect immunofluorescence local-ization of UCN-ir using affinity-purified fluorescein-labeledgoat anti-rabbit IgG (Tago, Burlingame, CA) to localize theprimary UCN antiserum (Sawchenko et al., 1990).
In each experiment, counts of the number of doublylabeled cells were obtained in a in series of 30-µm-thicksections collected at 150-µm intervals throughout therostrocaudal extent of the EW. Values were extrapolated toprovide estimates of population sizes and corrected for double-counting error using the method of Abercrombie (1946).
In situ hybridization
Hybridization histochemical localization of UCN mRNAwas carried out using 33P- or 35S-labeled antisense cRNAprobes. Perfusions (4% paraformaldehyde in 0.1 M sodiumborate buffer at 4°C, pH 9.5, with overnight postfixation)and histology were carried out as described above, and fullor partial series of sections through the brains, brain-stems, spinal cords, pituitaries, and selected peripheraltissues of over 20 animals were available for analysis.Techniques for probe synthesis, hybridization, and autora-diographic localization of mRNA signal were adapted fromSimmons et al. (1989). Briefly, tissue processed as abovewas mounted onto poly-L-lysine-coated slides and thendigested with 10 µg/ml of proteinase K for 30 minutes, at37°C. Antisense and sense (control) UCN probes weregenerated from a 550-bp UCN cDNA encompassing theentire coding sequence and ,60 bp of untranslated se-quence (Genbank accession number U33935), subclonedinto the pBluescript SK1 transcription vector (Strata-gene). Labeled antisense and sense were synthesized by
Fig. 1. Amino acid sequence of CRF family members. Deduced amino acid sequences of rat UCN(Vaughan et al., 1995) rat/human CRF (Vale et al., 1981), fish urotensin I (Lederis et al., 1982), and frogsauvagine (Monteucchi et al., 1981). Sequence identities are highlighted (gray). Boxes at the right denoteamidation sites.
288 J.C. BITTENCOURT ET AL.

linearizing the plasmid with XbaI and HindIII, respec-tively, and then using the appropriate RNA polymerases(T7 for antisense, T3 for sense) and 33P- or 35S-UTP and/orATP. In some experiments, the fragment length of probeswas adjusted to ,200 nucleotides by subjecting them tolimited alkaline hydrolysis, using the protocol of Cox et al.(1984). We have found this procedure to be effective indetecting multiple forms of mRNA that may be processedfrom a single primary transcript (unpublished observa-tions) and employed it here in an effort to minimize thepossibility that any splice variants of UCN mRNA mightescape detection.
The probes were used at concentrations of about 107
cpm/ml and applied to sections overnight at 56–58°C in asolution containing 50% formamide, 0.3 M NaCl, 10 mMTris (pH 8.0), 1 mM EDTA, 0.05% tRNA, 10 mM dithiothre-itol, 13 Denhardt’s solution, and 10% dextran sulfate,after which they were treated with 20 µg/ml of ribonucle-ase A for 30 minutes at 37°C and washed in 15 mMNaCl/1.5 mM sodium citrate at 55–60°C. Sections werethen dehydrated and exposed to X-ray films for 1–2 days.Sections were defatted in xylene, rinsed in absolute etha-nol, air dried, coated with Kodak NTB-2 liquid autoradio-graphic emulsion, and exposed at 4°C, dark and desic-cated, typically for 3–4 weeks, but occasionally longer.They were then developed with Kodak D-19 for 3.5 min-utes at 14°C, rinsed briefly in distilled water, fixed withfilm strength Kodak rapid fixer for 2 minutes at 14°C,rinsed again, and counterstained with thionin for refer-ence purposes.
Imaging
All images were captured on Ilford XP-2 or KodakEktachrome 160 (tungsten) film, imported into AdobePhotoshop (v. 5.02) using a Kodak RS-3570 film scanner,cropped and adjusted to balance brightness and contrast,exported to Canvas (v. 3.54) for assembly, and saved at 300dpi.
RESULTS
Antiserum specificity
Immunoperoxidase material from nonmanipulated ani-mals revealed a cellular distribution of UCN-ir that wascompatible with UCN mRNA expression patterns reportedpreviously (Vaughan et al., 1995, and see below). To gaininitial insight as to the specificity and selectivity of this, wecompared labeling achieved using molecular and/or immu-nologic probes for CRF, UCN, frog sauvagine, and fishurotensin I (Fig. 1) for their ability to label posited sites ofcellular UCN expression in the EW and LSO, as well asacknowledged loci of CRF expression, such as in theparaventricular nucleus of the hypothalamus and centralnucleus of the amygdala (Sawchenko and Swanson, 1989;Table 1a). CRF and UCN mRNA expression were, indeed,strictly segregated in this manner. Of the antisera tested,only the one raised against UCN supported staining ofcomparable selectivity, robustly labeling neurons in theEW and LSO, with no trace of cellular labeling in the PVHor central nucleus. Anti-CRF serum preferentially labeledcells and fibers in the PVH and amygdala, but also ones inthe EW and LSO with moderate intensity (see also Chunget al., 1987; Imaki et al., 1991). The reverse was seen usinganti-urotensin sera, and all four cell groups were labeled
with comparable (low-moderate) intensity using an anti-sauvagine serum.
The results of competition studies used to characterizethe UCN primary antiserum (code 5779) are given in Table1b. Staining for UCN-ir in cells and fibers throughout thebrain was completely abolished by preincubation of theantiserum with concentrations of the synthetic immuno-gen in the low micromolar range. The most closely relatedof the other family members tested, urotensin I (63%sequence homology), attenuated staining, although withsubstantially lesser potency than UCN. Preadsorptionwith sauvagine (33% homology) only moderately dimin-ished staining at higher concentrations, and rat CRF (45%homology) was completely inert in these experiments. It isnoteworthy that a-MSH, which shares an amidated C-terminal valine residue in common with UCN, was alsoineffective in interfering with immunostaining for UCN,even at milimolar concentrations. Bulky aliphatic aminoacids at this position appear to be highly immunogenic, acharacteristic that very likely accounts for spurious immu-nolabeling commonly reported using antisera against sev-eral peptides that share this motif (Nahon et al., 1989;Bittencourt et al., 1992).
Despite the apparent specificity of the anti-UCN serum,examination of cellular sites of UCN-ir expression incolchicine-treated animals presented initial difficulties ininterpretation. Thus, in addition to the EW, LSO, and thehandful of additional cell groups highlighted below, colchi-cine-pretreated rats displayed low-moderate intensity la-beling in a number of additional cell groups that werenever seen in normal material to express UCN mRNA,even using probes that were synthesized using two radiola-beled bases and/or were subjected to limited alkalinehydrolysis. These additional cell groups included the sub-stantia nigra and ventral tegmental area, sites at whichothers have also reported UCN-ir perikarya (Kozicz et al.,1998; Yamamoto et al., 1998). Staining at these additionalloci responded in an essentially similar manner as the EWor LSO in competition experiments with graded concentra-tions of a battery of CRF-related peptides. Repetition ofhybridization histochemical localization in tissue from
TABLE 1A. Cellular Labeling for CRF Family Membersin Rat Brain Regions1
Structure UCN mRNA UCN-ir CRF mRNA CRF-ir Uro-ir Svg-ir
EW 1111 1111 2 11 1111 11LSO 1111 1111 2 11 111 1PVH 2 2 1111 1111 11 11CeA 2 2 1111 1111 11 11
1For comparisons of hybridization histochemical material, the strength of CRF mRNAsignals in the PVH and CeA, and of UCN mRNA expression in the EW and LSO, wereassigned a rating of (1111). The same was done independently for immunostainedmaterial. Labeling intensity was rated blind using a 5-point scale, with (1111)denoting staining that was indistinguishable from that achieved using control (non-preadsorbed) antisera, and (2) indicating a complete absence of specific labeling.
TABLE 1B. Competition Characteristics of UCN-ir in Rat Brain1
Competingpeptide
Concentration (µM)
2.5 12.5 25 125 250
UCN 2 2 2 2 2CRF 1111 1111 1111 1111 1111Urotensin I 1111 1111 111 11 11Sauvagine 1111 1111 1111 111 11
1Staining intensity was rated blind using a 5-point scale, with (1111) denotinglabeling whose intensity was indistinguishable from that achieved using control(non-preadsorbed) antisera, and (2) indicating a complete absence of specific labeling.
UROCORTIN EXPRESSION IN BRAIN 289
colchicine-treated animals, however, revealed an ex-panded distribution, fully compatible with the immunola-beling results. Although the bases for the unexpectedcolchicine dependence of some UCN mRNA localizationsare unclear, the results do provide a coherent overview ofcellular UCN expression patterns using two independentmethods.
Cellular localization of UCN-ir and mRNA
Histochemical localization methods carried out in non-manipulated rats yielded consistent patterns of cellularUCN-ir and mRNA expression. Although the bulk of ourobservations were made on material derived from malerats, a smaller umber (n 5 6) of females showed similarlabeling patterns. These are described below in order oftheir prominence.
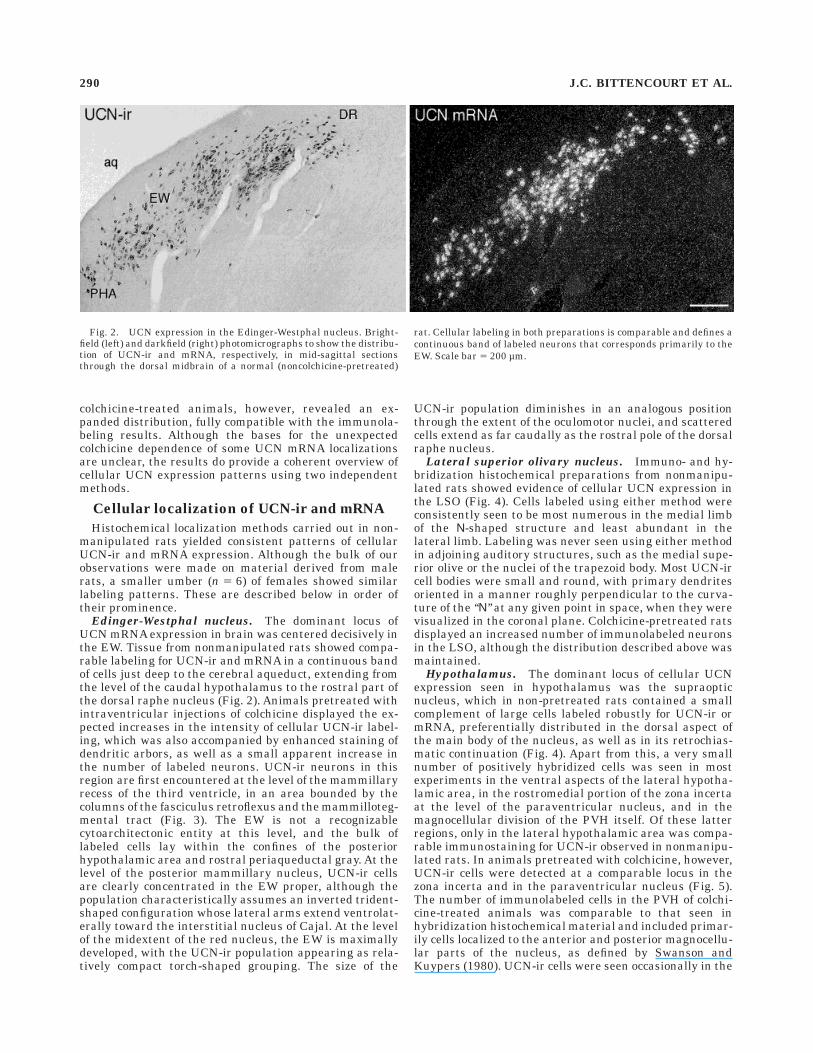
Edinger-Westphal nucleus. The dominant locus ofUCN mRNA expression in brain was centered decisively inthe EW. Tissue from nonmanipulated rats showed compa-rable labeling for UCN-ir and mRNA in a continuous bandof cells just deep to the cerebral aqueduct, extending fromthe level of the caudal hypothalamus to the rostral part ofthe dorsal raphe nucleus (Fig. 2). Animals pretreated withintraventricular injections of colchicine displayed the ex-pected increases in the intensity of cellular UCN-ir label-ing, which was also accompanied by enhanced staining ofdendritic arbors, as well as a small apparent increase inthe number of labeled neurons. UCN-ir neurons in thisregion are first encountered at the level of the mammillaryrecess of the third ventricle, in an area bounded by thecolumns of the fasciculus retroflexus and the mammilloteg-mental tract (Fig. 3). The EW is not a recognizablecytoarchitectonic entity at this level, and the bulk oflabeled cells lay within the confines of the posteriorhypothalamic area and rostral periaqueductal gray. At thelevel of the posterior mammillary nucleus, UCN-ir cellsare clearly concentrated in the EW proper, although thepopulation characteristically assumes an inverted trident-shaped configuration whose lateral arms extend ventrolat-erally toward the interstitial nucleus of Cajal. At the levelof the midextent of the red nucleus, the EW is maximallydeveloped, with the UCN-ir population appearing as rela-tively compact torch-shaped grouping. The size of the
UCN-ir population diminishes in an analogous positionthrough the extent of the oculomotor nuclei, and scatteredcells extend as far caudally as the rostral pole of the dorsalraphe nucleus.
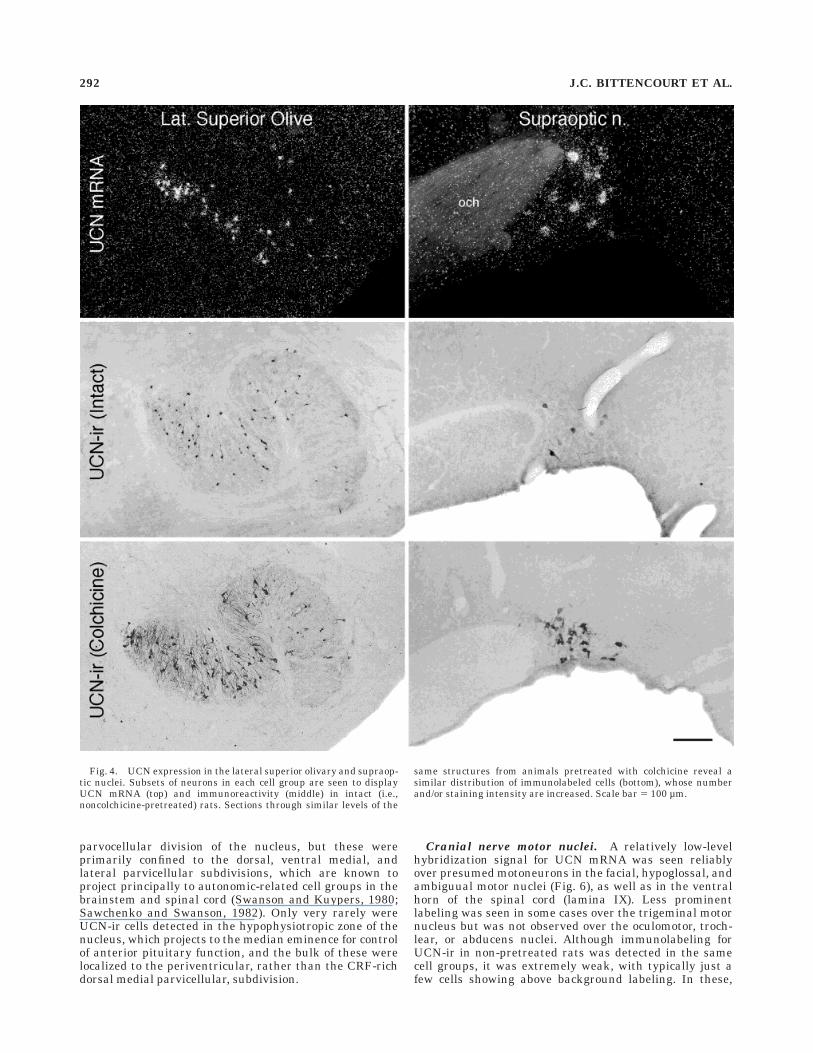
Lateral superior olivary nucleus. Immuno- and hy-bridization histochemical preparations from nonmanipu-lated rats showed evidence of cellular UCN expression inthe LSO (Fig. 4). Cells labeled using either method wereconsistently seen to be most numerous in the medial limbof the N-shaped structure and least abundant in thelateral limb. Labeling was never seen using either methodin adjoining auditory structures, such as the medial supe-rior olive or the nuclei of the trapezoid body. Most UCN-ircell bodies were small and round, with primary dendritesoriented in a manner roughly perpendicular to the curva-ture of the ‘‘N’’ at any given point in space, when they werevisualized in the coronal plane. Colchicine-pretreated ratsdisplayed an increased number of immunolabeled neuronsin the LSO, although the distribution described above wasmaintained.
Hypothalamus. The dominant locus of cellular UCNexpression seen in hypothalamus was the supraopticnucleus, which in non-pretreated rats contained a smallcomplement of large cells labeled robustly for UCN-ir ormRNA, preferentially distributed in the dorsal aspect ofthe main body of the nucleus, as well as in its retrochias-matic continuation (Fig. 4). Apart from this, a very smallnumber of positively hybridized cells was seen in mostexperiments in the ventral aspects of the lateral hypotha-lamic area, in the rostromedial portion of the zona incertaat the level of the paraventricular nucleus, and in themagnocellular division of the PVH itself. Of these latterregions, only in the lateral hypothalamic area was compa-rable immunostaining for UCN-ir observed in nonmanipu-lated rats. In animals pretreated with colchicine, however,UCN-ir cells were detected at a comparable locus in thezona incerta and in the paraventricular nucleus (Fig. 5).The number of immunolabeled cells in the PVH of colchi-cine-treated animals was comparable to that seen inhybridization histochemical material and included primar-ily cells localized to the anterior and posterior magnocellu-lar parts of the nucleus, as defined by Swanson andKuypers (1980). UCN-ir cells were seen occasionally in the
Fig. 2. UCN expression in the Edinger-Westphal nucleus. Bright-field (left) and darkfield (right) photomicrographs to show the distribu-tion of UCN-ir and mRNA, respectively, in mid-sagittal sectionsthrough the dorsal midbrain of a normal (noncolchicine-pretreated)
rat. Cellular labeling in both preparations is comparable and defines acontinuous band of labeled neurons that corresponds primarily to theEW. Scale bar 5 200 µm.
290 J.C. BITTENCOURT ET AL.
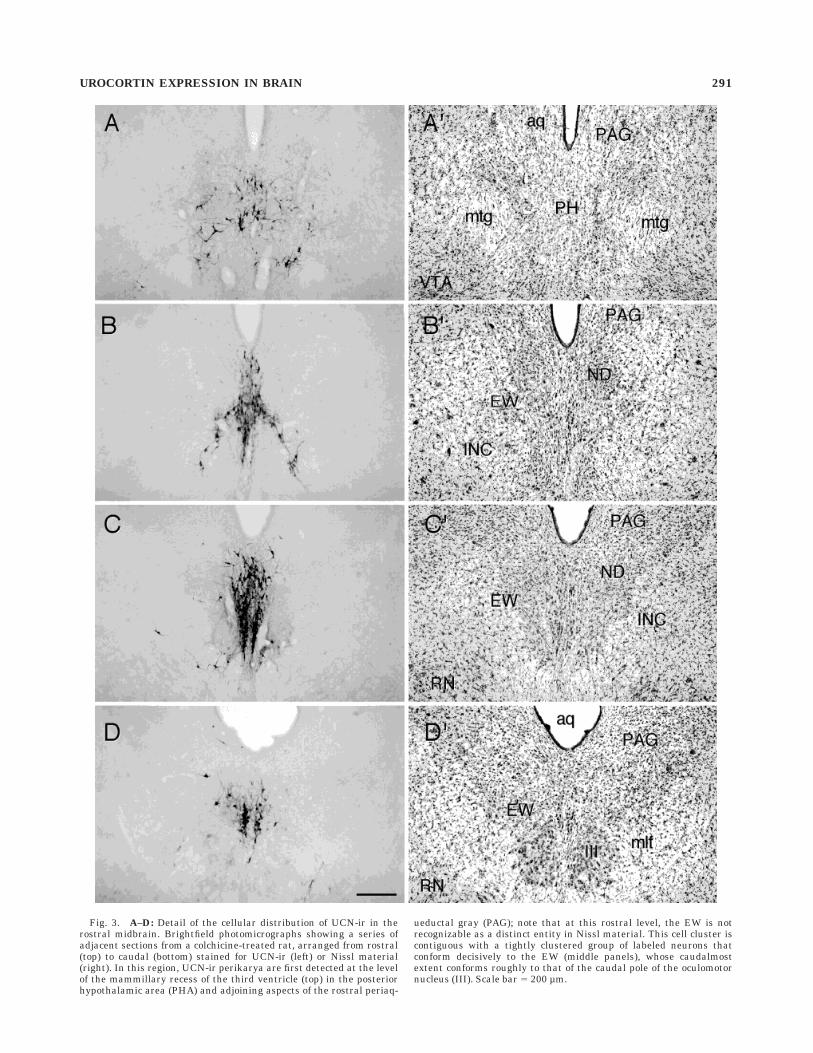
Fig. 3. A–D: Detail of the cellular distribution of UCN-ir in therostral midbrain. Brightfield photomicrographs showing a series ofadjacent sections from a colchicine-treated rat, arranged from rostral(top) to caudal (bottom) stained for UCN-ir (left) or Nissl material(right). In this region, UCN-ir perikarya are first detected at the levelof the mammillary recess of the third ventricle (top) in the posteriorhypothalamic area (PHA) and adjoining aspects of the rostral periaq-
ueductal gray (PAG); note that at this rostral level, the EW is notrecognizable as a distinct entity in Nissl material. This cell cluster iscontiguous with a tightly clustered group of labeled neurons thatconform decisively to the EW (middle panels), whose caudalmostextent conforms roughly to that of the caudal pole of the oculomotornucleus (III). Scale bar 5 200 µm.
UROCORTIN EXPRESSION IN BRAIN 291
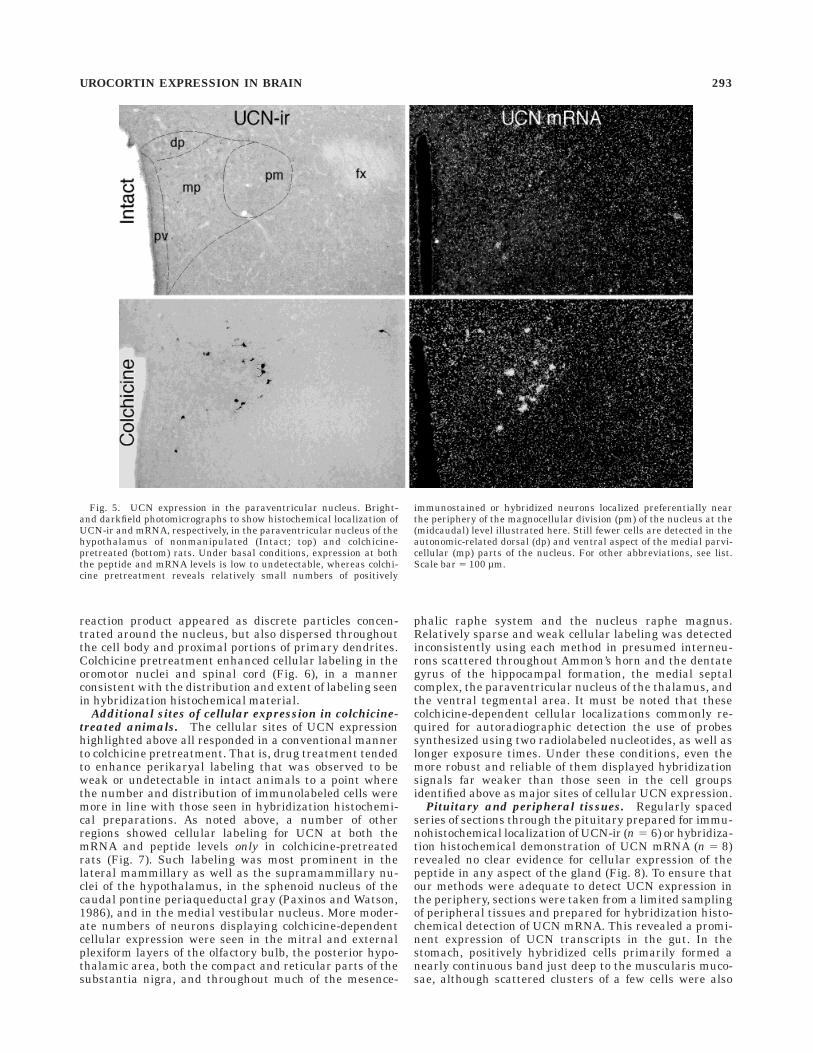
parvocellular division of the nucleus, but these wereprimarily confined to the dorsal, ventral medial, andlateral parvicellular subdivisions, which are known toproject principally to autonomic-related cell groups in thebrainstem and spinal cord (Swanson and Kuypers, 1980;Sawchenko and Swanson, 1982). Only very rarely wereUCN-ir cells detected in the hypophysiotropic zone of thenucleus, which projects to the median eminence for controlof anterior pituitary function, and the bulk of these werelocalized to the periventricular, rather than the CRF-richdorsal medial parvicellular, subdivision.
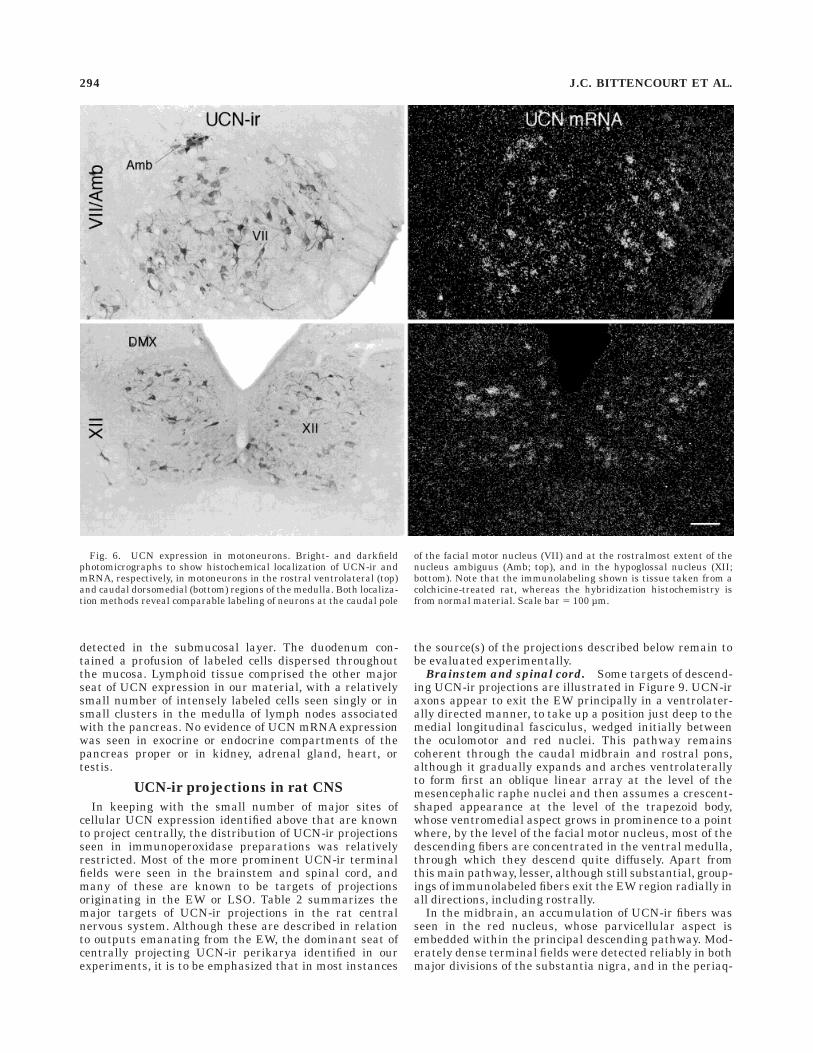
Cranial nerve motor nuclei. A relatively low-levelhybridization signal for UCN mRNA was seen reliablyover presumed motoneurons in the facial, hypoglossal, andambiguual motor nuclei (Fig. 6), as well as in the ventralhorn of the spinal cord (lamina IX). Less prominentlabeling was seen in some cases over the trigeminal motornucleus but was not observed over the oculomotor, troch-lear, or abducens nuclei. Although immunolabeling forUCN-ir in non-pretreated rats was detected in the samecell groups, it was extremely weak, with typically just afew cells showing above background labeling. In these,
Fig. 4. UCN expression in the lateral superior olivary and supraop-tic nuclei. Subsets of neurons in each cell group are seen to displayUCN mRNA (top) and immunoreactivity (middle) in intact (i.e.,noncolchicine-pretreated) rats. Sections through similar levels of the
same structures from animals pretreated with colchicine reveal asimilar distribution of immunolabeled cells (bottom), whose numberand/or staining intensity are increased. Scale bar 5 100 µm.
292 J.C. BITTENCOURT ET AL.
reaction product appeared as discrete particles concen-trated around the nucleus, but also dispersed throughoutthe cell body and proximal portions of primary dendrites.Colchicine pretreatment enhanced cellular labeling in theoromotor nuclei and spinal cord (Fig. 6), in a mannerconsistent with the distribution and extent of labeling seenin hybridization histochemical material.
Additional sites of cellular expression in colchicine-
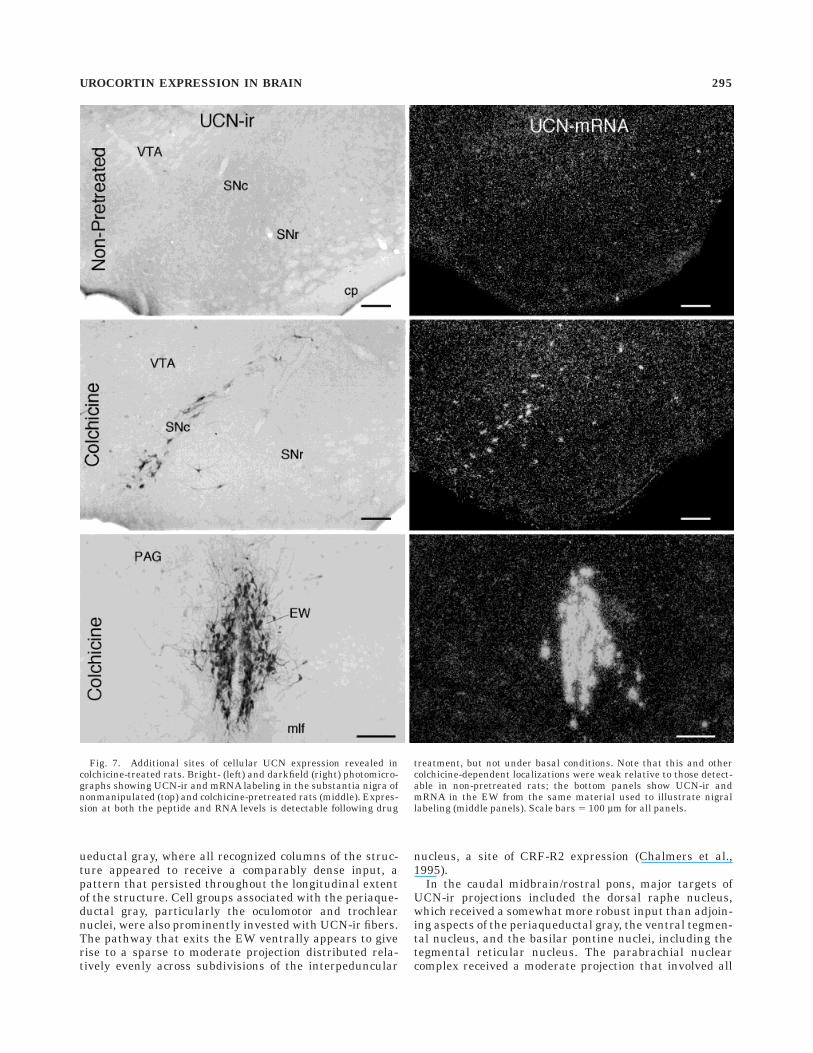
treated animals. The cellular sites of UCN expressionhighlighted above all responded in a conventional mannerto colchicine pretreatment. That is, drug treatment tendedto enhance perikaryal labeling that was observed to beweak or undetectable in intact animals to a point wherethe number and distribution of immunolabeled cells weremore in line with those seen in hybridization histochemi-cal preparations. As noted above, a number of otherregions showed cellular labeling for UCN at both themRNA and peptide levels only in colchicine-pretreatedrats (Fig. 7). Such labeling was most prominent in thelateral mammillary as well as the supramammillary nu-clei of the hypothalamus, in the sphenoid nucleus of thecaudal pontine periaqueductal gray (Paxinos and Watson,1986), and in the medial vestibular nucleus. More moder-ate numbers of neurons displaying colchicine-dependentcellular expression were seen in the mitral and externalplexiform layers of the olfactory bulb, the posterior hypo-thalamic area, both the compact and reticular parts of thesubstantia nigra, and throughout much of the mesence-
phalic raphe system and the nucleus raphe magnus.Relatively sparse and weak cellular labeling was detectedinconsistently using each method in presumed interneu-rons scattered throughout Ammon’s horn and the dentategyrus of the hippocampal formation, the medial septalcomplex, the paraventricular nucleus of the thalamus, andthe ventral tegmental area. It must be noted that thesecolchicine-dependent cellular localizations commonly re-quired for autoradiographic detection the use of probessynthesized using two radiolabeled nucleotides, as well aslonger exposure times. Under these conditions, even themore robust and reliable of them displayed hybridizationsignals far weaker than those seen in the cell groupsidentified above as major sites of cellular UCN expression.
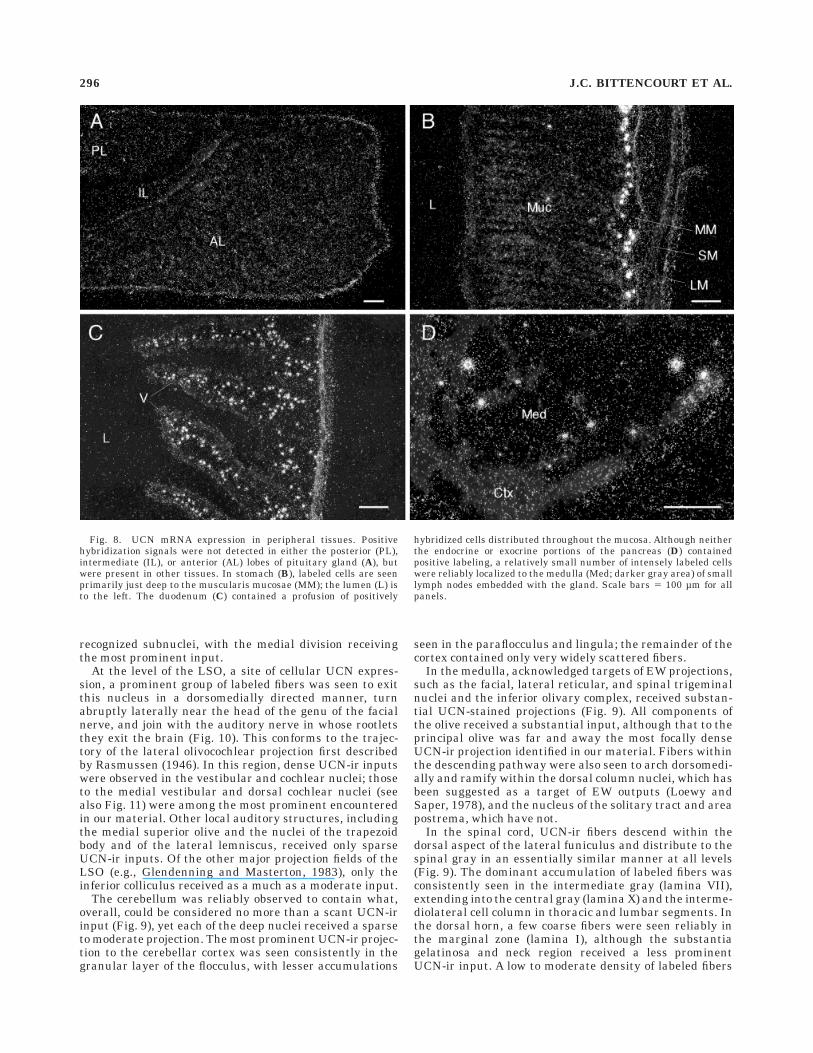
Pituitary and peripheral tissues. Regularly spacedseries of sections through the pituitary prepared for immu-nohistochemical localization of UCN-ir (n 5 6) or hybridiza-tion histochemical demonstration of UCN mRNA (n 5 8)revealed no clear evidence for cellular expression of thepeptide in any aspect of the gland (Fig. 8). To ensure thatour methods were adequate to detect UCN expression inthe periphery, sections were taken from a limited samplingof peripheral tissues and prepared for hybridization histo-chemical detection of UCN mRNA. This revealed a promi-nent expression of UCN transcripts in the gut. In thestomach, positively hybridized cells primarily formed anearly continuous band just deep to the muscularis muco-sae, although scattered clusters of a few cells were also
Fig. 5. UCN expression in the paraventricular nucleus. Bright-and darkfield photomicrographs to show histochemical localization ofUCN-ir and mRNA, respectively, in the paraventricular nucleus of thehypothalamus of nonmanipulated (Intact; top) and colchicine-pretreated (bottom) rats. Under basal conditions, expression at boththe peptide and mRNA levels is low to undetectable, whereas colchi-cine pretreatment reveals relatively small numbers of positively
immunostained or hybridized neurons localized preferentially nearthe periphery of the magnocellular division (pm) of the nucleus at the(midcaudal) level illustrated here. Still fewer cells are detected in theautonomic-related dorsal (dp) and ventral aspect of the medial parvi-cellular (mp) parts of the nucleus. For other abbreviations, see list.Scale bar 5 100 µm.
UROCORTIN EXPRESSION IN BRAIN 293
detected in the submucosal layer. The duodenum con-tained a profusion of labeled cells dispersed throughoutthe mucosa. Lymphoid tissue comprised the other majorseat of UCN expression in our material, with a relativelysmall number of intensely labeled cells seen singly or insmall clusters in the medulla of lymph nodes associatedwith the pancreas. No evidence of UCN mRNA expressionwas seen in exocrine or endocrine compartments of thepancreas proper or in kidney, adrenal gland, heart, ortestis.
UCN-ir projections in rat CNS
In keeping with the small number of major sites ofcellular UCN expression identified above that are knownto project centrally, the distribution of UCN-ir projectionsseen in immunoperoxidase preparations was relativelyrestricted. Most of the more prominent UCN-ir terminalfields were seen in the brainstem and spinal cord, andmany of these are known to be targets of projectionsoriginating in the EW or LSO. Table 2 summarizes themajor targets of UCN-ir projections in the rat centralnervous system. Although these are described in relationto outputs emanating from the EW, the dominant seat ofcentrally projecting UCN-ir perikarya identified in ourexperiments, it is to be emphasized that in most instances
the source(s) of the projections described below remain tobe evaluated experimentally.
Brainstem and spinal cord. Some targets of descend-ing UCN-ir projections are illustrated in Figure 9. UCN-iraxons appear to exit the EW principally in a ventrolater-ally directed manner, to take up a position just deep to themedial longitudinal fasciculus, wedged initially betweenthe oculomotor and red nuclei. This pathway remainscoherent through the caudal midbrain and rostral pons,although it gradually expands and arches ventrolaterallyto form first an oblique linear array at the level of themesencephalic raphe nuclei and then assumes a crescent-shaped appearance at the level of the trapezoid body,whose ventromedial aspect grows in prominence to a pointwhere, by the level of the facial motor nucleus, most of thedescending fibers are concentrated in the ventral medulla,through which they descend quite diffusely. Apart fromthis main pathway, lesser, although still substantial, group-ings of immunolabeled fibers exit the EW region radially inall directions, including rostrally.
In the midbrain, an accumulation of UCN-ir fibers wasseen in the red nucleus, whose parvicellular aspect isembedded within the principal descending pathway. Mod-erately dense terminal fields were detected reliably in bothmajor divisions of the substantia nigra, and in the periaq-
Fig. 6. UCN expression in motoneurons. Bright- and darkfieldphotomicrographs to show histochemical localization of UCN-ir andmRNA, respectively, in motoneurons in the rostral ventrolateral (top)and caudal dorsomedial (bottom) regions of the medulla. Both localiza-tion methods reveal comparable labeling of neurons at the caudal pole
of the facial motor nucleus (VII) and at the rostralmost extent of thenucleus ambiguus (Amb; top), and in the hypoglossal nucleus (XII;bottom). Note that the immunolabeling shown is tissue taken from acolchicine-treated rat, whereas the hybridization histochemistry isfrom normal material. Scale bar 5 100 µm.
294 J.C. BITTENCOURT ET AL.
ueductal gray, where all recognized columns of the struc-ture appeared to receive a comparably dense input, apattern that persisted throughout the longitudinal extentof the structure. Cell groups associated with the periaque-ductal gray, particularly the oculomotor and trochlearnuclei, were also prominently invested with UCN-ir fibers.The pathway that exits the EW ventrally appears to giverise to a sparse to moderate projection distributed rela-tively evenly across subdivisions of the interpeduncular
nucleus, a site of CRF-R2 expression (Chalmers et al.,1995).
In the caudal midbrain/rostral pons, major targets ofUCN-ir projections included the dorsal raphe nucleus,which received a somewhat more robust input than adjoin-ing aspects of the periaqueductal gray, the ventral tegmen-tal nucleus, and the basilar pontine nuclei, including thetegmental reticular nucleus. The parabrachial nuclearcomplex received a moderate projection that involved all
Fig. 7. Additional sites of cellular UCN expression revealed incolchicine-treated rats. Bright- (left) and darkfield (right) photomicro-graphs showing UCN-ir and mRNA labeling in the substantia nigra ofnonmanipulated (top) and colchicine-pretreated rats (middle). Expres-sion at both the peptide and RNA levels is detectable following drug
treatment, but not under basal conditions. Note that this and othercolchicine-dependent localizations were weak relative to those detect-able in non-pretreated rats; the bottom panels show UCN-ir andmRNA in the EW from the same material used to illustrate nigrallabeling (middle panels). Scale bars 5 100 µm for all panels.
UROCORTIN EXPRESSION IN BRAIN 295
recognized subnuclei, with the medial division receivingthe most prominent input.
At the level of the LSO, a site of cellular UCN expres-sion, a prominent group of labeled fibers was seen to exitthis nucleus in a dorsomedially directed manner, turnabruptly laterally near the head of the genu of the facialnerve, and join with the auditory nerve in whose rootletsthey exit the brain (Fig. 10). This conforms to the trajec-tory of the lateral olivocochlear projection first describedby Rasmussen (1946). In this region, dense UCN-ir inputswere observed in the vestibular and cochlear nuclei; thoseto the medial vestibular and dorsal cochlear nuclei (seealso Fig. 11) were among the most prominent encounteredin our material. Other local auditory structures, includingthe medial superior olive and the nuclei of the trapezoidbody and of the lateral lemniscus, received only sparseUCN-ir inputs. Of the other major projection fields of theLSO (e.g., Glendenning and Masterton, 1983), only theinferior colliculus received as a much as a moderate input.
The cerebellum was reliably observed to contain what,overall, could be considered no more than a scant UCN-irinput (Fig. 9), yet each of the deep nuclei received a sparseto moderate projection. The most prominent UCN-ir projec-tion to the cerebellar cortex was seen consistently in thegranular layer of the flocculus, with lesser accumulations
seen in the paraflocculus and lingula; the remainder of thecortex contained only very widely scattered fibers.
In the medulla, acknowledged targets of EW projections,such as the facial, lateral reticular, and spinal trigeminalnuclei and the inferior olivary complex, received substan-tial UCN-stained projections (Fig. 9). All components ofthe olive received a substantial input, although that to theprincipal olive was far and away the most focally denseUCN-ir projection identified in our material. Fibers withinthe descending pathway were also seen to arch dorsomedi-ally and ramify within the dorsal column nuclei, which hasbeen suggested as a target of EW outputs (Loewy andSaper, 1978), and the nucleus of the solitary tract and areapostrema, which have not.
In the spinal cord, UCN-ir fibers descend within thedorsal aspect of the lateral funiculus and distribute to thespinal gray in an essentially similar manner at all levels(Fig. 9). The dominant accumulation of labeled fibers wasconsistently seen in the intermediate gray (lamina VII),extending into the central gray (lamina X) and the interme-diolateral cell column in thoracic and lumbar segments. Inthe dorsal horn, a few coarse fibers were seen reliably inthe marginal zone (lamina I), although the substantiagelatinosa and neck region received a less prominentUCN-ir input. A low to moderate density of labeled fibers
Fig. 8. UCN mRNA expression in peripheral tissues. Positivehybridization signals were not detected in either the posterior (PL),intermediate (IL), or anterior (AL) lobes of pituitary gland (A), butwere present in other tissues. In stomach (B), labeled cells are seenprimarily just deep to the muscularis mucosae (MM); the lumen (L) isto the left. The duodenum (C) contained a profusion of positively
hybridized cells distributed throughout the mucosa. Although neitherthe endocrine or exocrine portions of the pancreas (D) containedpositive labeling, a relatively small number of intensely labeled cellswere reliably localized to the medulla (Med; darker gray area) of smalllymph nodes embedded with the gland. Scale bars 5 100 µm for allpanels.
296 J.C. BITTENCOURT ET AL.

was detected in the ventral horn, including a small num-ber of fibers directly apposed to motoneurons.
Forebrain projections. Although ascending projec-tions arising from the EW nucleus have not been docu-mented, UCN-ir fibers were reliably seen to exit thenucleus in a rostralward direction and ascend through(and ramify within) the rostral periaqueductal gray, poste-rior hypothalamic area, and particularly the nucleus reu-niens of the thalamus. The extent to which these ascend-ing projections might be reinforced, or even dominated, byfibers arising from nearby cell groups (posterior hypotha-lamic area, supramammillary nucleus, rostral periaqueduc-tal gray) in which cellular UCN expression was detectedonly in colchicine-treated animals could not be ascertainedwith confidence in our material. In the course of theirascent, labeled axons appeared to peel off laterally to enterthe zona incerta and ventrolaterally to join the medialforebrain bundle. With a few prominent exceptions, UCN-irprojections to the forebrain were sparse and diffuselyorganized. The strongest inputs were seen to aspects of thehypothalamus and limbic forebrain (Table 2).
Through the course of their ascent through the ventralmidline thalamus, labeled fibers splay out in such amanner that most major thalamic and epithalamic (lateralhabenula) nuclei contained at least a few scattered fibers,with nearby midline and intralaminar nuclei tending toreceive somewhat stronger inputs. The dominant UCN-irterminal field in the dorsal thalamus, however, was local-ized to the intergeniculate leaflet and secondarily to theventral lateral geniculate nucleus. Fibers traversing thecaudal zona incerta and reticular nucleus of the thalamusappear to contribute to this projection, although the pres-ence of UCN-ir fibers in the optic tract may be indicative ofa retinal contribution.
Several major territories of the forebrain, including thecerebral cortex, basal ganglia, and amygdala were verysparsely invested with UCN-ir axons. A small contingenttraveling in the medial forebrain bundle appears to followthe ventral amygdalofugal pathway to give rise to focal,although still no more than moderately dense, terminalfields in the medial part of the central nucleus, and in theposterior cortical nucleus, of the amygdala. Most of thelabeled fibers that reach the cerebral cortex appear to do sovia this route, as temporal isocortical fields contained thegreatest (although still very small) number of UCN-irfibers, which were preferentially distributed to deeperlayers. Similarly, only isolated UCN-ir fibers were seen inthe hippocampal formation.
In the course of their ascent through the medial fore-brain bundle, UCN-ir fibers ramified and gave rise toterminal bouton-like specializations diffusely in the lateralhypothalamic area.All recognized hypothalamic cell groupsreceived contained fibers, with only a few standing out asreceiving anything more than a sparse input. These in-cluded the dorsomedial nucleus, the parvicellular divisionof the paraventricular nucleus, the central cell condensa-tion of the anterior hypothalamic area, the medial preopticnucleus, and, particularly, the suprachiasmatic nucleus,which was seen to receive a relatively dense input concen-trated in is ventromedial aspect.
The principal seat of UCN-ir perikarya in the forebrain,the supraoptic nucleus, is known as a pure magnocellularneurosecretory cell group whose only established projec-tion is via the internal lamina of the median eminence tothe posterior pituitary. In keeping with this, a small
number of varicose UCN-ir fibers visualized in the hypo-thalamohypophyseal tract could be traced to the medianeminence, where labeled axons and varicosities cut incross section were quite strictly confined to the internallamina (Fig. 12). These were joined by fibers originating inthe paraventricular nucleus, that arch laterally over thefornix and then ventrally and medially over the optictracts toward the median eminence. In the posteriorpituitary, terminal arbors of axons distributed preferen-tially toward the periphery of the gland; the density ofthese was moderate, relative to other UCN-labeled projec-tions in brain, but quite modest, relative to that commonlyreported using antisera against oxytocin or vasopressin(e.g., van Leeuwen et al., 1979), the dominant secretedproducts of the magnocellular neurosecretory system. La-beled fibers were not detected in the intermediate oranterior lobes.
The septal/preoptic region contained the most robustUCN-ir projections to the forebrain. Thus the bed nucleusof the stria terminalis, substantia innominata, and medialseptal/diagonal band complex all contained focally moder-ate projections. However, far and away the most promi-nent and distinctive UCN-ir input to forebrain was one todiscrete aspects of the intermediate lateral septal nucleus,which ended in pericellular basket-like arrays (see Figs. 11and 13 and below). Beyond the level of the septum, no morethan sparse to moderate immunolabeled projections werelabeled to olfactory structures and frontal cortex, althoughthe infra- and prelimbic areas did receive a somewhatmore prominent input than other medial cortical fields.
Urocortin- versus urotensin-ir projections to the fore-
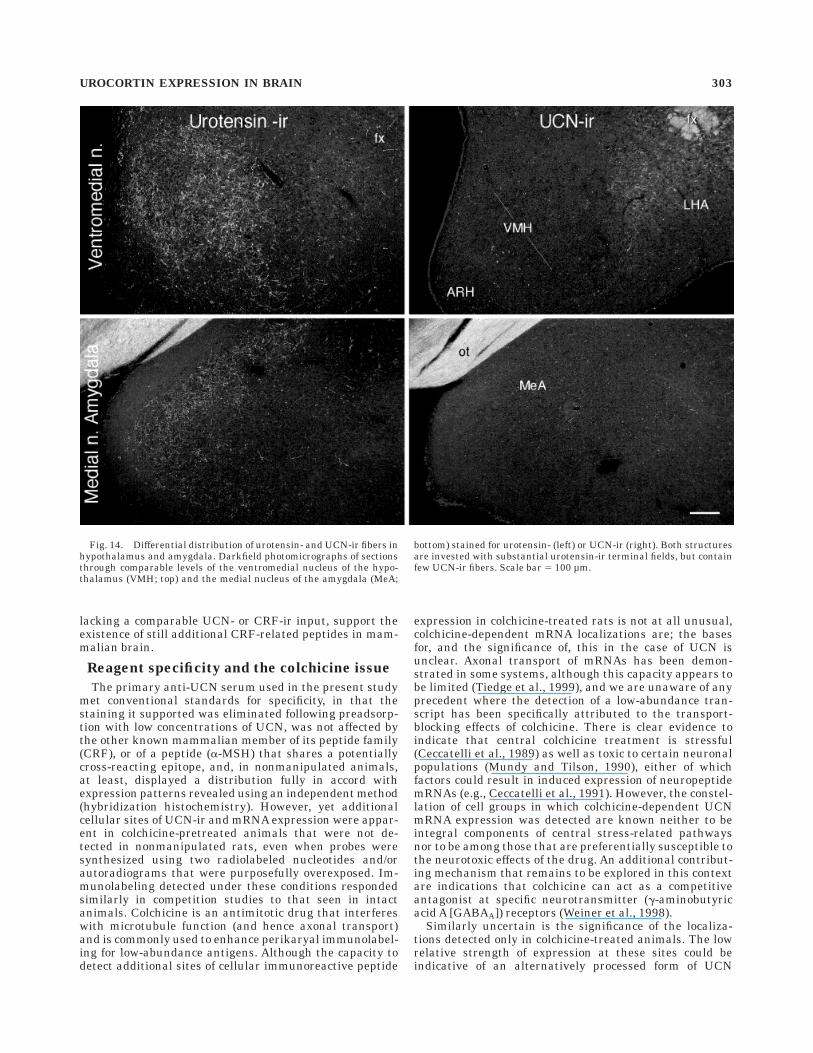
brain. In previous work, the presence of urotensin-ir incellular sites of UCN mRNA expression prompted the useof urotensin antisera to glean initial insight into what waspresumed to be the distribution of UCN-ir projections(Vaughan et al., 1995; see also Table 1a); this revealedprominent urotensin-ir projections that overlapped withthe distribution of CRF-R2 expression in a number of loci,prominently including the lateral septal and dorsal raphenuclei. Here, we have compared directly the distribution ofUCN- and urotensin-ir projections and have found a ratherstriking lack of congruence in the forebrain. Although bothantisera do indeed label strong projections to the lateralseptal nucleus, their distributions are distinct, with uroten-sin-ir fibers occupying the ventrolateral aspect of theintermediate lateral septal nucleus and UCN-ir ones termi-nating more dorsally and medially in the intermediatepart and in the ventral nucleus (Fig. 13). In addition, weconsistently failed to observed UCN-ir counterparts ofrobust urotensin-stained inputs to the ventromedialnucleus of the hypothalamus and to the medial nucleus ofthe amygdala (Fig. 14).
Some UCN-containing projections of the EW. Com-bined retrograde transport-immunohistochemical meth-ods were used to evaluate the extent to which UCN-irmight be contained in cells that project to documentedtargets of the EW, namely, the spinal cord and in somespecies the ciliary ganglion. In addition, and in view of thelack of evidence that the EW gives rise to ascendingprojections, or that any of the major cellular sites of UCNexpression highlighted above project to the forebrain,similar methods were used to determine whether UCN-ircells could be identified that contribute to the prominentterminal field in the septal region (Figs. 15, 16).
UROCORTIN EXPRESSION IN BRAIN 297
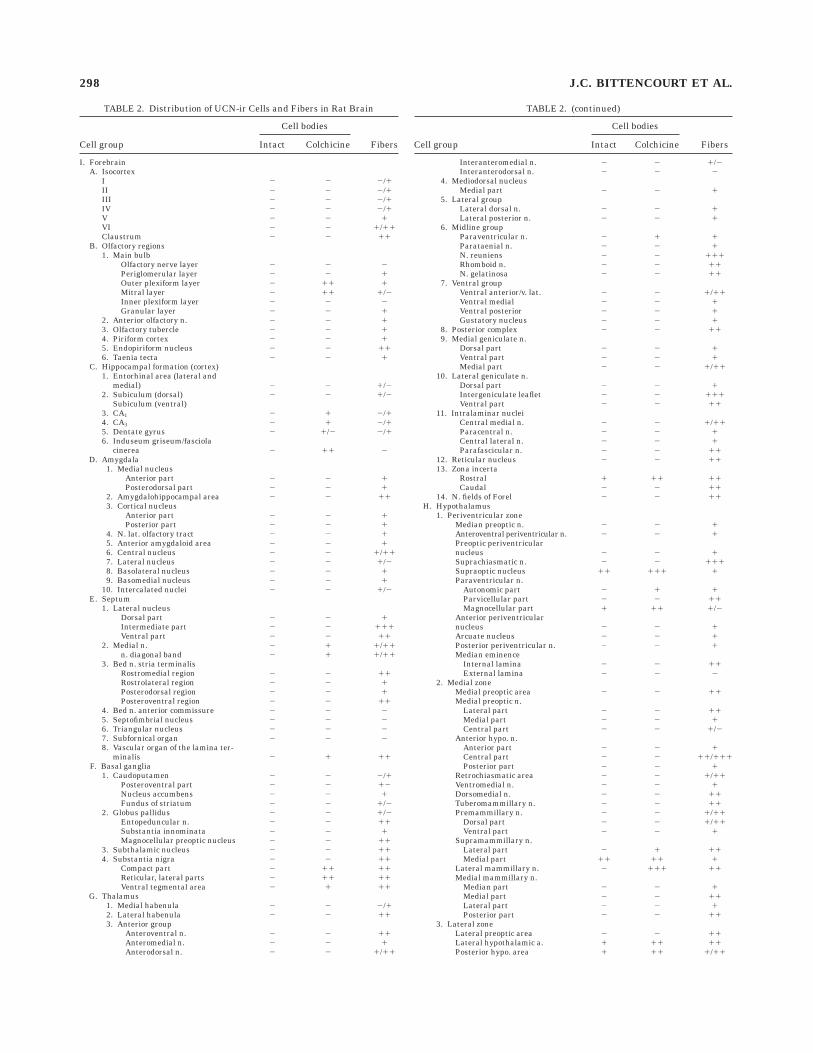
TABLE 2. Distribution of UCN-ir Cells and Fibers in Rat Brain
Cell group
Cell bodies
FibersIntact Colchicine
I. ForebrainA. Isocortex
I 2 2 2/1II 2 2 2/1III 2 2 2/1IV 2 2 2/1V 2 2 1VI 2 2 1/11Claustrum 2 2 11
B. Olfactory regions1. Main bulb
Olfactory nerve layer 2 2 2Periglomerular layer 2 2 1Outer plexiform layer 2 11 1Mitral layer 2 11 1/2Inner plexiform layer 2 2 2Granular layer 2 2 1
2. Anterior olfactory n. 2 2 13. Olfactory tubercle 2 2 14. Piriform cortex 2 2 15. Endopiriform nucleus 2 2 116. Taenia tecta 2 2 1
C. Hippocampal formation (cortex)1. Entorhinal area (lateral and
medial) 2 2 1/22. Subiculum (dorsal) 2 2 1/2
Subiculum (ventral)3. CA1 2 1 2/14. CA3 2 1 2/15. Dentate gyrus 2 1/2 2/16. Induseum griseum/fasciola
cinerea 2 11 2D. Amygdala
1. Medial nucleusAnterior part 2 2 1Posterodorsal part 2 2 1
2. Amygdalohippocampal area 2 2 113. Cortical nucleus
Anterior part 2 2 1Posterior part 2 2 1
4. N. lat. olfactory tract 2 2 15. Anterior amygdaloid area 2 2 16. Central nucleus 2 2 1/117. Lateral nucleus 2 2 1/28. Basolateral nucleus 2 2 19. Basomedial nucleus 2 2 1
10. Intercalated nuclei 2 2 1/2E. Septum
1. Lateral nucleusDorsal part 2 2 1Intermediate part 2 2 111Ventral part 2 2 11
2. Medial n. 2 1 1/11n. diagonal band 2 1 1/11
3. Bed n. stria terminalisRostromedial region 2 2 11Rostrolateral region 2 2 1Posterodorsal region 2 2 1Posteroventral region 2 2 11
4. Bed n. anterior commissure 2 2 25. Septofimbrial nucleus 2 2 26. Triangular nucleus 2 2 27. Subfornical organ 2 2 28. Vascular organ of the lamina ter-
minalis 2 1 11F. Basal ganglia
1. Caudoputamen 2 2 2/1Posteroventral part 2 2 12Nucleus accumbens 2 2 1Fundus of striatum 2 2 1/2
2. Globus pallidus 2 2 1/2Entopeduncular n. 2 2 11Substantia innominata 2 2 1Magnocellular preoptic nucleus 2 2 11
3. Subthalamic nucleus 2 2 114. Substantia nigra 2 2 11
Compact part 2 11 11Reticular, lateral parts 2 11 11Ventral tegmental area 2 1 11
G. Thalamus1. Medial habenula 2 2 2/12. Lateral habenula 2 2 113. Anterior group
Anteroventral n. 2 2 11Anteromedial n. 2 2 1Anterodorsal n. 2 2 1/11
TABLE 2. (continued)
Cell group
Cell bodies
FibersIntact Colchicine
Interanteromedial n. 2 2 1/2Interanterodorsal n. 2 2 2
4. Mediodorsal nucleusMedial part 2 2 1
5. Lateral groupLateral dorsal n. 2 2 1Lateral posterior n. 2 2 1
6. Midline groupParaventricular n. 2 1 1Parataenial n. 2 2 1N. reuniens 2 2 111Rhomboid n. 2 2 11N. gelatinosa 2 2 11
7. Ventral groupVentral anterior/v. lat. 2 2 1/11Ventral medial 2 2 1Ventral posterior 2 2 1Gustatory nucleus 2 2 1
8. Posterior complex 2 2 119. Medial geniculate n.
Dorsal part 2 2 1Ventral part 2 2 1Medial part 2 2 1/11
10. Lateral geniculate n.Dorsal part 2 2 1Intergeniculate leaflet 2 2 111Ventral part 2 2 11
11. Intralaminar nucleiCentral medial n. 2 2 1/11Paracentral n. 2 2 1Central lateral n. 2 2 1Parafascicular n. 2 2 11
12. Reticular nucleus 2 2 1113. Zona incerta
Rostral 1 11 11Caudal 2 2 11
14. N. fields of Forel 2 2 11H. Hypothalamus
1. Periventricular zoneMedian preoptic n. 2 2 1Anteroventral periventricular n. 2 2 1Preoptic periventricularnucleus 2 2 1Suprachiasmatic n. 2 2 111Supraoptic nucleus 11 111 1Paraventricular n.
Autonomic part 2 1 1Parvicellular part 2 2 11Magnocellular part 1 11 1/2
Anterior periventricularnucleus 2 2 1Arcuate nucleus 2 2 1Posterior periventricular n. 2 2 1Median eminence
Internal lamina 2 2 11External lamina 2 2 2
2. Medial zoneMedial preoptic area 2 2 11Medial preoptic n.
Lateral part 2 2 11Medial part 2 2 1Central part 2 2 1/2
Anterior hypo. n.Anterior part 2 2 1Central part 2 2 11/111Posterior part 2 2 1
Retrochiasmatic area 2 2 1/11Ventromedial n. 2 2 1Dorsomedial n. 2 2 11Tuberomammillary n. 2 2 11Premammillary n. 2 2 1/11
Dorsal part 2 2 1/11Ventral part 2 2 1
Supramammillary n.Lateral part 2 1 11Medial part 11 11 1
Lateral mammillary n. 2 111 11Medial mammillary n.
Median part 2 2 1Medial part 2 2 11Lateral part 2 2 1Posterior part 2 2 11
3. Lateral zoneLateral preoptic area 2 2 11Lateral hypothalamic a. 1 11 11Posterior hypo. area 1 11 1/11
298 J.C. BITTENCOURT ET AL.

Six experiments were obtained in which spinal injec-tions of fast blue or diamidino yellow effectively encom-passed a hemisection of the cord at upper thoracic levels.In these experiments, injections into the spinal cord werefound to result in labeling of as many as 25 cells per sectionin the EW, the overwhelming majority of which were foundto colabel for UCN-ir (Fig. 16). Corrected counts of thenumber of doubly labeled neurons in these experimentsestimated the size of the spinally projecting UCN-ir popu-lation to be 172 6 18 (mean 6 SEM; n 5 6).
Five separate animals received crystalline deposits offast blue in the forebrain that were centered in the lateralseptal nuclei (Fig. 15). These commonly impinged upon themedial septal nucleus to varying extents and occasionallyspread ventrally to include small portions of the bednucleus of the stria terminals or dorsally into overlyingcortex. In these cases, retrogradely labeled neurons werereliably detected in the EW (Fig. 16), but these were
TABLE 2. (continued)
Cell group
Cell bodies
FibersIntact Colchicine
I. Pituitary gland1. Anterior lobe 2 2 22. Intermediate lobe 2 2 23. Posterior lobe 2 2 11
II. BrainstemA. Sensory
1. VisualSuperior colliculus
Zonal layer 2 2 1Superficial gray 2 2 1/11Optic layer 2 2 1Intermediate gray 2 2 11/111Deep gray 2 2 1/11
Parabigeminal n. 2 2 1Pretectal region
Olivary n. 2 2 1N. optic tract 2 2 1Anterior n. 2 2 11/111Posterior n. 2 2 1Medial pretectal a. 2 2 1N. posterior commissure 2 2 11
Medial terminal n. 2 2 22. Somatosensory
Mesencephalic n. V 2 2 11Principal sensory n. V 2 2 1/11Spinal n. V
Oral part 2 2 11Interpolar part 2 2 11Caudal part 2 2 11
Dorsal column n. 2 2 11External cuneate n. 2 2 1/11
3. AuditoryCochlear nuclei
Dorsal 2 2 1111Ventral 2 2 11
N. trapezoid body 2 2 1Lateral superior olive 111 1111 111Medial superior olive 2 2 1N. lateral lemniscus
Ventral 2 2 1Dorsal 2 2 1
Inferior colliculusExternal 2 2 11Dorsal 2 2 11Central 2 2 11
N. brachium inf. coll. 2 2 1N. saguluum 2 2 1
4. VestibularMedial n. 2 111 1111Lateral n. 2 2 111Superior n. 2 2 111Spinal n. 2 2 11
5. GustatoryN. solitary tract, ant. 2 2 1
6. VisceralN. solitary tract
Medial part 2 2 11Commissural part 2 2 11Lateral part 2 2 11
Area postrema 2 2 11Parabrachial n.
Lateral 2 2 1/11Medial 2 2 11Kolliker-Fuse n. 2 2 1
B. Motor1. Eye
Oculomotor (III) 2 2 11/111Trochlear (IV) 2 2 1Abducens (VI) 2 2 1/11
2. JawMotor n. V 11 11 1/2
3. FaceFacial n. (VII) 111 111 11
4. Pharynx/larynxN. ambiguus 111 111 1
5. TongueHypoglossal n. (XII) 111 111 11
6. VisceraDorsal motor n. X 2 2 1
C. Reticular core (inc. central gray andraphe)1. Periaqueductal gray/assoc.
w/PAGPeriaqueductal gray 1 11 11Interstitial nucleus of Cajal 2 2 111N. Darkschewitsch 2 2 11Dorsal tegmental n. 2 2 11
TABLE 2. (continued)
Cell group
Cell bodies
FibersIntact Colchicine
Edinger-Westphal n. 1111 1111 11Ventral tegmental n. 2 2 11N. incertus 2 2 11Laterodorsal teg. n. 2 2 1Barrington’s n. 2 2 1Locus coeruleus 2 2 1/11Sphenoid n. 2 111 11
2. RapheInterfascicular n. 2 11 1Rostral linear n. 2 1 1Dorsal raphe 2 11 11/111Median raphe 2 111 1/11N. raphe pontis 2 2 1N. raphe magnus 2 11 1/11N. raphe obscurus 2 2 1N. raphe pallidus 2 2 1
3. Interpeduncular n.Rostral subnucleus 2 2 1Apical subnucleus 2 1 1Dorsomedial subnucleus 2 1 1/11Lateral subnucleus 2 2 1Intermediate subnucleus 2 11 1Central subnucleus 2 11 1/11
4. Reticular formationCentral teg. field
Subcuneiform part 2 2 1/11Retrorubral part 2 2 11
Peripeduncular n. 2 2 1Pedunculopontine n. 2 2 11Cuneiform n. 2 2 1/11Pontine reticular 2 2 1Linear n. medulla 2 2 1/11Parvicellular ret. field 2 2 1/11Gigantocellular ret. field 2 2 1/11Lateral paragigantocellular 2 2 111Paramedian reticular n. 2 2 1Magnocellular reticular n. 2 11 11
D. Pre- and postcerebellar1. Pontine gray 2 2 1112. Tegmental reticular n. 2 2 11/1113. Inferior olive 2 2 1114. Lateral reticular n. 2 2 115. Red nucleus 2 2 1116. N. Roller 2 2 17. N. prepositus 2 2 11
III. CerebellumA. Deep nuclei 2 2 1/11B. Cortex
Molecular layer 2 2 1Purkinje layer 2 2 1Granular layer 2 2 1
IV. Spinal cordA. Marginal zone 2 2 1B. Substantia gelatinosa 2 2 1C. Intermediate gray 2 2 11/111D. Intermediolateral column 2 2 11E. Ventral horn 2 11 11F. Central gray 2 2 11
UROCORTIN EXPRESSION IN BRAIN 299
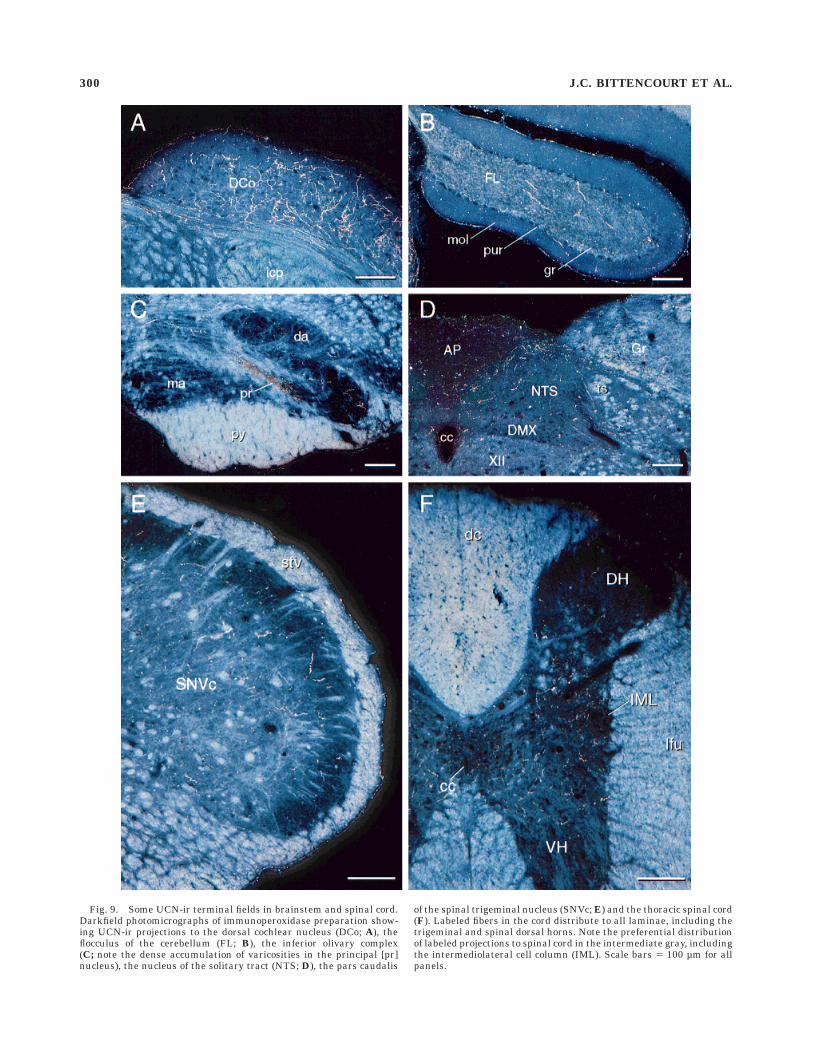
Fig. 9. Some UCN-ir terminal fields in brainstem and spinal cord.Darkfield photomicrographs of immunoperoxidase preparation show-ing UCN-ir projections to the dorsal cochlear nucleus (DCo; A), theflocculus of the cerebellum (FL; B), the inferior olivary complex(C; note the dense accumulation of varicosities in the principal [pr]nucleus), the nucleus of the solitary tract (NTS; D), the pars caudalis
of the spinal trigeminal nucleus (SNVc; E) and the thoracic spinal cord(F). Labeled fibers in the cord distribute to all laminae, including thetrigeminal and spinal dorsal horns. Note the preferential distributionof labeled projections to spinal cord in the intermediate gray, includingthe intermediolateral cell column (IML). Scale bars 5 100 µm for allpanels.
300 J.C. BITTENCOURT ET AL.
substantially less numerous (1–5 per section) than wasseen following spinal injections. Very nearly all of thesestained positively for UCN-ir, and the estimated size of thetotal UCN-ir population identified as projecting to theseptal region was 43 6 6 neurons.
DISCUSSION
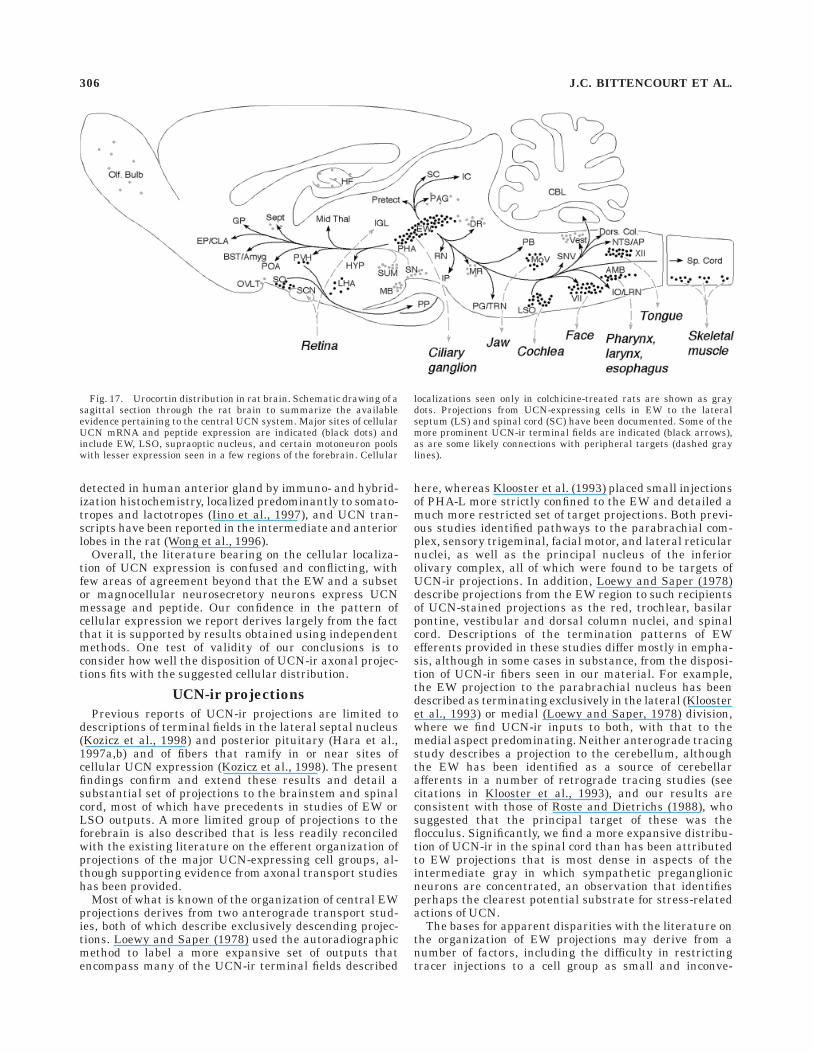
An overview of the organization of the central UCNsystem suggested by the present findings is provided asFigure 17. The results suggest that the principal sites ofcellular UCN expression in rat brain are ones whose majorconnections are thought or known to be with structuresthat lay outside the central nervous system. These (andtheir targets) include the EW (ciliary ganglion), the LSO(cochlea), the supraoptic nucleus (posterior pituitary), andmotoneurons of the caudal brainstem and spinal cord(striated muscle). UCN-ir projections within the CNS arepredominantly descending and are consistent with knownoutputs of the EW and LSO, targeting principally acces-
sory optic, precerebellar, and auditory cell groups, as wellas the spinal intermediate gray. Whereas neither the EWnor LSO are known to project to the forebrain, UCN-irneurons in the EW were identified that project to thelateral septal nucleus, which houses a prominent UCN-irterminal field. Substantial UCN-ir projections were ob-served to several cell groups in the brainstem that expressCRF-R2, including the dorsal raphe and interpeduncularnuclei and the nucleus of the solitary tract; prominentseats of CRF-R2 expression in the forebrain were found tocontain inputs that stained positively using antisera raisedagainst piscine urotensin I, but not rat UCN. The resultsdefine a central UCN system that is involved principally inmotor control and sensorimotor integration; its participa-tion in stress-related mechanisms probably derives princi-pally from projections to the spinal intermediolateralcolumn, the NTS, and the paraventricular nucleus. Sev-eral observations, including the lack of a pervasive relation-ship of UCN-ir projections with CRF-R2-expressing tar-gets and the presence of urotensin-ir projections in areas
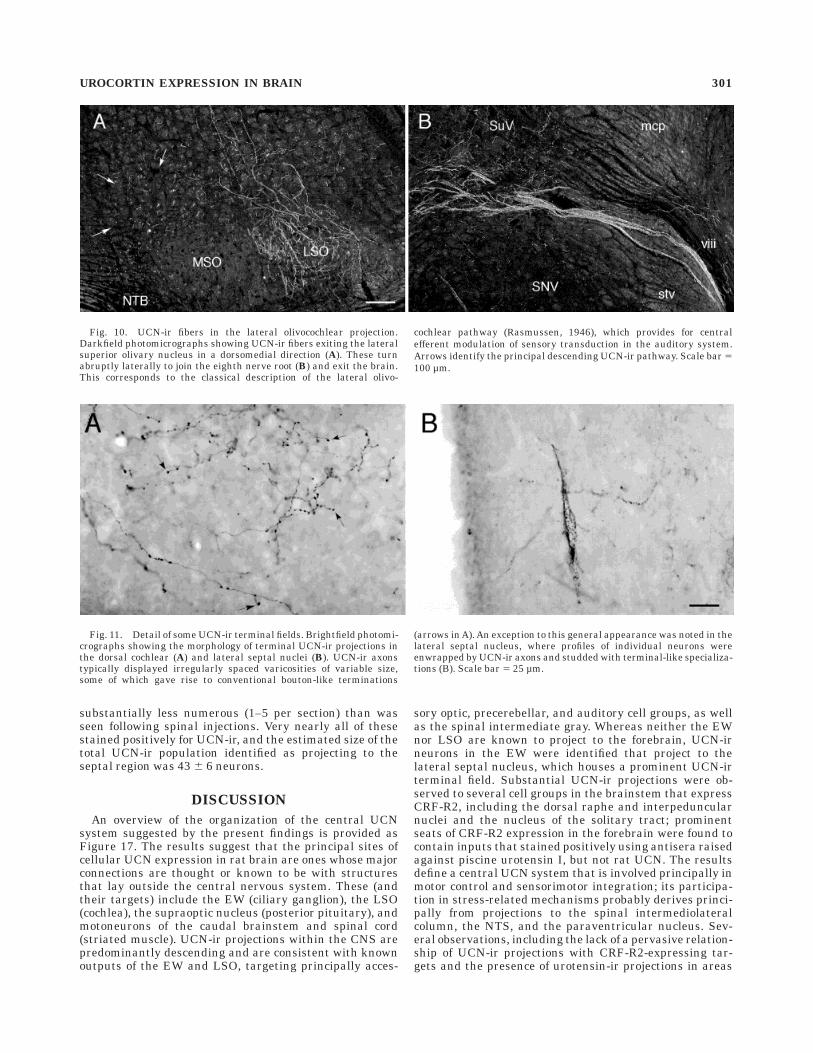
Fig. 10. UCN-ir fibers in the lateral olivocochlear projection.Darkfield photomicrographs showing UCN-ir fibers exiting the lateralsuperior olivary nucleus in a dorsomedial direction (A). These turnabruptly laterally to join the eighth nerve root (B) and exit the brain.This corresponds to the classical description of the lateral olivo-
cochlear pathway (Rasmussen, 1946), which provides for centralefferent modulation of sensory transduction in the auditory system.Arrows identify the principal descending UCN-ir pathway. Scale bar 5100 µm.
Fig. 11. Detail of some UCN-ir terminal fields. Brightfield photomi-crographs showing the morphology of terminal UCN-ir projections inthe dorsal cochlear (A) and lateral septal nuclei (B). UCN-ir axonstypically displayed irregularly spaced varicosities of variable size,some of which gave rise to conventional bouton-like terminations
(arrows in A). An exception to this general appearance was noted in thelateral septal nucleus, where profiles of individual neurons wereenwrapped by UCN-ir axons and studded with terminal-like specializa-tions (B). Scale bar 5 25 µm.
UROCORTIN EXPRESSION IN BRAIN 301
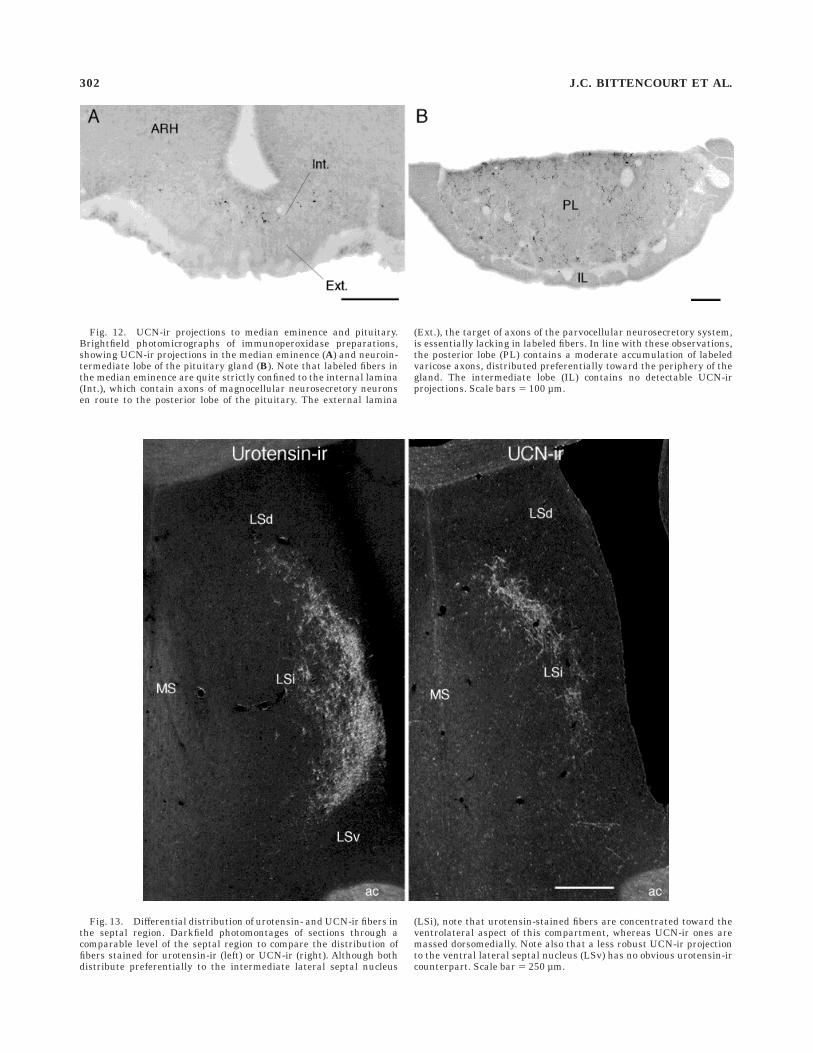
Fig. 12. UCN-ir projections to median eminence and pituitary.Brightfield photomicrographs of immunoperoxidase preparations,showing UCN-ir projections in the median eminence (A) and neuroin-termediate lobe of the pituitary gland (B). Note that labeled fibers inthe median eminence are quite strictly confined to the internal lamina(Int.), which contain axons of magnocellular neurosecretory neuronsen route to the posterior lobe of the pituitary. The external lamina
(Ext.), the target of axons of the parvocellular neurosecretory system,is essentially lacking in labeled fibers. In line with these observations,the posterior lobe (PL) contains a moderate accumulation of labeledvaricose axons, distributed preferentially toward the periphery of thegland. The intermediate lobe (IL) contains no detectable UCN-irprojections. Scale bars 5 100 µm.
Fig. 13. Differential distribution of urotensin- and UCN-ir fibers inthe septal region. Darkfield photomontages of sections through acomparable level of the septal region to compare the distribution offibers stained for urotensin-ir (left) or UCN-ir (right). Although bothdistribute preferentially to the intermediate lateral septal nucleus
(LSi), note that urotensin-stained fibers are concentrated toward theventrolateral aspect of this compartment, whereas UCN-ir ones aremassed dorsomedially. Note also that a less robust UCN-ir projectionto the ventral lateral septal nucleus (LSv) has no obvious urotensin-ircounterpart. Scale bar 5 250 µm.
302 J.C. BITTENCOURT ET AL.
lacking a comparable UCN- or CRF-ir input, support theexistence of still additional CRF-related peptides in mam-malian brain.
Reagent specificity and the colchicine issue
The primary anti-UCN serum used in the present studymet conventional standards for specificity, in that thestaining it supported was eliminated following preadsorp-tion with low concentrations of UCN, was not affected bythe other known mammalian member of its peptide family(CRF), or of a peptide (a-MSH) that shares a potentiallycross-reacting epitope, and, in nonmanipulated animals,at least, displayed a distribution fully in accord withexpression patterns revealed using an independent method(hybridization histochemistry). However, yet additionalcellular sites of UCN-ir and mRNA expression were appar-ent in colchicine-pretreated animals that were not de-tected in nonmanipulated rats, even when probes weresynthesized using two radiolabeled nucleotides and/orautoradiograms that were purposefully overexposed. Im-munolabeling detected under these conditions respondedsimilarly in competition studies to that seen in intactanimals. Colchicine is an antimitotic drug that interfereswith microtubule function (and hence axonal transport)and is commonly used to enhance perikaryal immunolabel-ing for low-abundance antigens. Although the capacity todetect additional sites of cellular immunoreactive peptide
expression in colchicine-treated rats is not at all unusual,colchicine-dependent mRNA localizations are; the basesfor, and the significance of, this in the case of UCN isunclear. Axonal transport of mRNAs has been demon-strated in some systems, although this capacity appears tobe limited (Tiedge et al., 1999), and we are unaware of anyprecedent where the detection of a low-abundance tran-script has been specifically attributed to the transport-blocking effects of colchicine. There is clear evidence toindicate that central colchicine treatment is stressful(Ceccatelli et al., 1989) as well as toxic to certain neuronalpopulations (Mundy and Tilson, 1990), either of whichfactors could result in induced expression of neuropeptidemRNAs (e.g., Ceccatelli et al., 1991). However, the constel-lation of cell groups in which colchicine-dependent UCNmRNA expression was detected are known neither to beintegral components of central stress-related pathwaysnor to be among those that are preferentially susceptible tothe neurotoxic effects of the drug. An additional contribut-ing mechanism that remains to be explored in this contextare indications that colchicine can act as a competitiveantagonist at specific neurotransmitter (g-aminobutyricacid A [GABAA]) receptors (Weiner et al., 1998).
Similarly uncertain is the significance of the localiza-tions detected only in colchicine-treated animals. The lowrelative strength of expression at these sites could beindicative of an alternatively processed form of UCN
Fig. 14. Differential distribution of urotensin- and UCN-ir fibers inhypothalamus and amygdala. Darkfield photomicrographs of sectionsthrough comparable levels of the ventromedial nucleus of the hypo-thalamus (VMH; top) and the medial nucleus of the amygdala (MeA;
bottom) stained for urotensin- (left) or UCN-ir (right). Both structuresare invested with substantial urotensin-ir terminal fields, but containfew UCN-ir fibers. Scale bar 5 100 µm.
UROCORTIN EXPRESSION IN BRAIN 303

mRNA and peptide, although splice variants of UCNtranscripts have not been described, and the use of probessubjected to limited alkaline hydrolysis (Cox et al., 1984), aprocedure that has proved effective in localizing multipleRNA species that may be processed from a single primarytranscript, failed to yield any more robust labeling inintact or colchicine-treated rats. The fact that hybridiza-tion histochemistry was carried out under high stringencyconditions make it seem unlikely that both antiserum andprobe may be recognizing the products of a novel gene,unless the degree of structural relatedness is much greaterthan that exhibited among known CRF family members.We consider it most likely that these represent sites thatexpress authentic UCN at very low levels and that can beactivated by as yet unknown sequelae of intracerebroven-tricular colchicine pretreatment.
Our findings suggest that these additional sites ofcellular expression very likely contribute to the distribu-tion of UCN-ir projections detectable in the brains ofnonmanipulated rats. This supposition is based partly onthe lack of evidence (see below) that the principal sites ofUCN expression project to the forebrain, where a low-moderate density of UCN-ir projections was seen reliably.For example, it seems more reasonable to expect that themoderate density of UCN-ir projections to the anteriorthalamic nuclear group arises from the lateral mammil-lary nucleus than from the EW or LSO. That thesepresumed secondary sites of cellular expression are notdominant sources of UCN-ir fibers is supported by suchobservations as the paucity of labeled fibers detected in thebasal ganglia, despite perikaryal staining in monominer-gic cell groups (substantia nigra, ventral tegmental area,mesencephalic raphe nuclei) that are major sources ofafferent input to the striatum.
Cellular expression
The present findings confirm previously reported(Vaughan et al., 1995) major sites of UCN mRNA expres-sion in the EW and LSO and less intense but pervasiveones in the caudal cranial nerve and spinal motor nucleiand in small populations in the supraoptic nucleus, lateralhypothalamic area, and olfactory bulb. Newly generatedmaterial adds to this only very small populations in therostromedial zona incerta and the paraventricular nucleusof the hypothalamus, as well as the ostensibly colchicine-dependent sites of expression considered above. Each ofthese principal localizations was also identified in thepresent study as a seat of cellular UCN-ir expression, withall but the small groupings in the paraventricular nucleusand zona incerta being consistently demonstrable in non-colchicine-pretreated animals. The only other study thathas used similar methods to examine UCN mRNA expres-sion in rat brain reports, in addition to a dominant signalover EW, widespread expression in such major territoriesof brain as the olfactory system, basal ganglia, amygdala,hippocampal formation, and cerebellar cortex (Wong et al.,1996). We are unable to account for the discrepanciesbetween our results and these, and it must be said thatreverse transcriptase polymerase chain reaction data areavailable from human brain to support such a broadpattern of cellular localization (Takahashi et al., 1998). Wewould note only that none of the immunolabeling worksupports so broad a distribution, and a recent study insheep brain has revealed an even more restricted distribu-tion of UCN mRNA than we describe here in rat (Cepoi etal., 1999).
Only a handful of previous reports have examinedUCN-ir expression in brain by immunohistochemicalmeans. In an analysis restricted to the endocrine hypothala-mus, Hara and colleagues (1997a,b) described a subset ofUCN-ir neurons in the supraoptic nucleus of nonmanipu-lated rats and found that exposure to certain challengeparadigms increased their number and provoked the ap-pearance of UCN-ir in a small number of cells in themagnocellular division of the PVH. These findings arefully compatible with the view advanced here, that thesupraoptic nucleus is the dominant seat of UCN-ir expres-sion in the endocrine hypothalamus, with the capacity forUCN expression in the PVH not being readily demon-strable under resting conditions. A study focused on themidbrain has recently found prominent UCN-ir labeling ofEW neurons, but also of dopaminergic cells in the substan-tia nigra and ventral tegmental area in nonmanipulatedrats (Yamamoto et al., 1998).
The most comprehensive study available (Kozicz et al.,1998) describes prominent labeling of UCN-ir neurons inthe EW and adjoining regions in and around the periaque-ductal gray, extending caudally well into the dorsal raphe,dorsal tegmental, and laterodorsal tegmental nuclei. Inaddition, cellular labeling was seen in magnocellularelements of the supraoptic, and to a lesser extent, theparaventricular nuclei. More substantial numbers of la-beled neurons were reported in the parvicellular divisionof the PVH, in the ventromedial and anterior periventricu-lar nuclei of the hypothalamus, and in the substantianigra. Labeled neurons were not observed in the pons,medulla, or spinal cord.
It is noteworthy that we adduced no evidence for cellularUCN expression in the pituitary gland. UCN has been
Fig. 15. Tracer injection site in the septal region. Combinedfluorescence/dim brightfield (A) and brightfield (B) photomicrographsof a section through the septal region to show the position and extentof a deposit of the retrogradely transported fluorochrome, fast blue.The injection clearly involves the intermediate lateral septal nucleus(LSi), which receives a prominent UCN-ir innervation. Scale bar 50.5 mm.
304 J.C. BITTENCOURT ET AL.
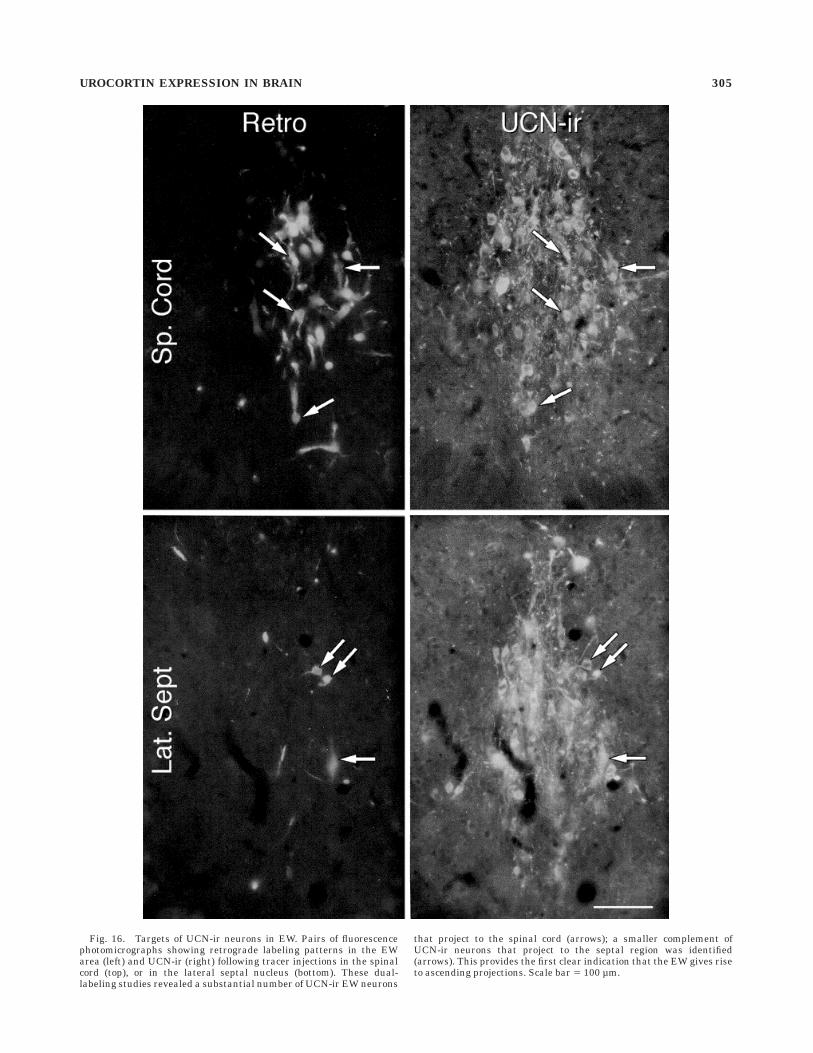
Fig. 16. Targets of UCN-ir neurons in EW. Pairs of fluorescencephotomicrographs showing retrograde labeling patterns in the EWarea (left) and UCN-ir (right) following tracer injections in the spinalcord (top), or in the lateral septal nucleus (bottom). These dual-labeling studies revealed a substantial number of UCN-ir EW neurons
that project to the spinal cord (arrows); a smaller complement ofUCN-ir neurons that project to the septal region was identified(arrows). This provides the first clear indication that the EW gives riseto ascending projections. Scale bar 5 100 µm.
UROCORTIN EXPRESSION IN BRAIN 305
detected in human anterior gland by immuno- and hybrid-ization histochemistry, localized predominantly to somato-tropes and lactotropes (Iino et al., 1997), and UCN tran-scripts have been reported in the intermediate and anteriorlobes in the rat (Wong et al., 1996).
Overall, the literature bearing on the cellular localiza-tion of UCN expression is confused and conflicting, withfew areas of agreement beyond that the EW and a subsetor magnocellular neurosecretory neurons express UCNmessage and peptide. Our confidence in the pattern ofcellular expression we report derives largely from the factthat it is supported by results obtained using independentmethods. One test of validity of our conclusions is toconsider how well the disposition of UCN-ir axonal projec-tions fits with the suggested cellular distribution.
UCN-ir projections
Previous reports of UCN-ir projections are limited todescriptions of terminal fields in the lateral septal nucleus(Kozicz et al., 1998) and posterior pituitary (Hara et al.,1997a,b) and of fibers that ramify in or near sites ofcellular UCN expression (Kozicz et al., 1998). The presentfindings confirm and extend these results and detail asubstantial set of projections to the brainstem and spinalcord, most of which have precedents in studies of EW orLSO outputs. A more limited group of projections to theforebrain is also described that is less readily reconciledwith the existing literature on the efferent organization ofprojections of the major UCN-expressing cell groups, al-though supporting evidence from axonal transport studieshas been provided.
Most of what is known of the organization of central EWprojections derives from two anterograde transport stud-ies, both of which describe exclusively descending projec-tions. Loewy and Saper (1978) used the autoradiographicmethod to label a more expansive set of outputs thatencompass many of the UCN-ir terminal fields described
here, whereas Klooster et al. (1993) placed small injectionsof PHA-L more strictly confined to the EW and detailed amuch more restricted set of target projections. Both previ-ous studies identified pathways to the parabrachial com-plex, sensory trigeminal, facial motor, and lateral reticularnuclei, as well as the principal nucleus of the inferiorolivary complex, all of which were found to be targets ofUCN-ir projections. In addition, Loewy and Saper (1978)describe projections from the EW region to such recipientsof UCN-stained projections as the red, trochlear, basilarpontine, vestibular and dorsal column nuclei, and spinalcord. Descriptions of the termination patterns of EWefferents provided in these studies differ mostly in empha-sis, although in some cases in substance, from the disposi-tion of UCN-ir fibers seen in our material. For example,the EW projection to the parabrachial nucleus has beendescribed as terminating exclusively in the lateral (Kloosteret al., 1993) or medial (Loewy and Saper, 1978) division,where we find UCN-ir inputs to both, with that to themedial aspect predominating. Neither anterograde tracingstudy describes a projection to the cerebellum, althoughthe EW has been identified as a source of cerebellarafferents in a number of retrograde tracing studies (seecitations in Klooster et al., 1993), and our results areconsistent with those of Roste and Dietrichs (1988), whosuggested that the principal target of these was theflocculus. Significantly, we find a more expansive distribu-tion of UCN-ir in the spinal cord than has been attributedto EW projections that is most dense in aspects of theintermediate gray in which sympathetic preganglionicneurons are concentrated, an observation that identifiesperhaps the clearest potential substrate for stress-relatedactions of UCN.
The bases for apparent disparities with the literature onthe organization of EW projections may derive from anumber of factors, including the difficulty in restrictingtracer injections to a cell group as small and inconve-
Fig. 17. Urocortin distribution in rat brain. Schematic drawing of asagittal section through the rat brain to summarize the availableevidence pertaining to the central UCN system. Major sites of cellularUCN mRNA and peptide expression are indicated (black dots) andinclude EW, LSO, supraoptic nucleus, and certain motoneuron poolswith lesser expression seen in a few regions of the forebrain. Cellular
localizations seen only in colchicine-treated rats are shown as graydots. Projections from UCN-expressing cells in EW to the lateralseptum (LS) and spinal cord (SC) have been documented. Some of themore prominent UCN-ir terminal fields are indicated (black arrows),as are some likely connections with peripheral targets (dashed graylines).
306 J.C. BITTENCOURT ET AL.

niently organized as the EW, which may be surmounted, insome respects, by the availability of an endogenous markerfor what is obviously a substantial fraction of EW neurons.In addition, however, projections from other UCN-ir cellgroups may account for portions of the observed labelingpatterns. The outputs of the LSO have been studied indetail, mainly in the cat (e.g., Elverland, 1978; Glenden-ning and Masterton, 1983; see also Webster, 1995), and itsmajor targets are known to include the nuclei or the laterallemniscus, the inferior colliculus, and the organ of Corti,where terminations on spiral ganglion fibers associatedwith inner hair cells provide a basis for central efferentmodulation of auditory sensory transduction (Warr et al.,1986). The distribution and morphology of UCN-ir neuronsin the LSO conformed to descriptions of lateral olivoco-chlear neurons identified in retrograde transport studiesin rat (Vetter and Mugnaini, 1992), and we were readilyable to follow a substantial complement of UCN-ir axonsinto the eighth nerve root. UCN-ir cells seem apt tocontribute in much less prominent ways to LSO projec-tions to the nuclei of the lateral lemniscus and inferiorcolliculus, which are known to arise from cells distinctfrom olivocochlear neurons (e.g., Aschoff and Ostwald,1988). Attributing a source to the prominent UCN-stainedinput to the cochlear nuclei is more problematic. Themedial olivocochlear projection, which arises from UCN-negative cells in the medial superior olive, is known toissue collateral projections to the cochlear and vestibularnuclei, but the lateral pathway is not (Brown et al., 1988).
The UCN-ir input to the NTS is among the moreintriguing projections identified in our material, as itconforms to a stress-related site of CRF action (Brown,1986), and one that has recently been found to express type2 CRF receptors (van Pett et al., unpublished observa-tions). This holds potential for resolving one of the moreglaring instances of ligand-receptor mismatch in the cen-tral CRF system, but to our knowledge neither the EW norLSO have been implicated as projecting to the NTS. Onepotential contributor to this pathway is a subset of theoxytocinergic neurons of the paraventricular nucleus ofthe hypothalamus that were identified as expressingUCN-ir. Although UCN-ir neurons in the paraventricularnucleus appeared to conform principally to magnocellularneurosecretory neurons, a small number of labeled cellswere found reliably in aspects of the parvicellular divisionthat project to central autonomic-related targets, includ-ing the NTS, and oxytocin is among the dominant knownneurochemical constituents of this projection (Sawchenkoand Swanson, 1982).
Because neither the EW nor the LSO are known toproject to the forebrain, UCN-ir projections to this regionare the most difficult to reconcile with the existing litera-ture. Several considerations are germane. First, UCN-ircells in the EW were reliably labeled following retrogradetracer injections into the lateral septal nucleus, whichhouses the most prominent UCN-stained terminal field inthe forebrain. The size of this population was small,relative to the spinally projecting contingent, but theresult provides strong evidence to support the existence ofascending projections from EW. Second, it must be borne inmind that UCN-ir cells in the EW region may not comprisean homogeneous population. Thus, UCN-ir neurons werefound outside the morphological confines of the EW, in theperiaqueductal gray, posterior hypothalamic area, and (incolchicine-treated rats) supramammillary nucleus, and
some of these were of a size and morphology that clearlydistinguished them from the main population of immuno-labeled cells in EW. Thus, for example, the standardreference (Herkenham, 1978) on the connections of thenucleus reuniens of the thalamus, which we found tocontain a major complement of ascending UCN-ir fibers,did not identify the EW as an afferent source; cellsprojecting to the nucleus reuniens were detected in thevery regions highlighted above as containing ‘‘ectopic’’UCN-ir neurons in the EW region. The same may well betrue of cells that project to the paraventricular nucleus (cf.,Sawchenko and Swanson, 1983). Third, minor sites ofcellular UCN expression were identified in the lateralhypothalamic area, zona incerta, and olfactory bulb thatmay contribute to, or independently account for, someUCN-ir projections in the forebrain. Finally, some of theUCN-ir fibers in forebrain may be indicative of connectionswith regions outside the brain. Magnocellular neurosecre-tory neurons provide a clear case in point, and prominentUCN-ir inputs to the suprachiasmatic nucleus and theintergeniculate leaflet may well originate, at least in part,in the retina; individual retinal ganglion cells that sendcollateral projections to both of these targets have beenidentified (Pickard, 1985).
Relationship to sites of CRFreceptor expression
Based on the finding that type 2 CRF receptors bindUCN with substantially higher affinity than they do CRF,and the observations that some major sites of CRF-R2expression display activational responses following cen-tral administration of UCN and contain substantial uroten-sin-stained terminal fields, we suggested that UCN may bea preferred endogenous ligand for this receptor subtype(Vaughan et al., 1995), and this hypothesis has served toframe a good deal of the ensuing functional and anatomicalwork on the new peptide. Although we have identifiedinstances in which UCN-ir projections are in a position tointeract with CRF-R2-expressing neurons, such a relation-ship appears not to be a pervasive, or defining, one. Areasof ligand-receptor congruence include the interpeduncularand dorsal raphe nuclei (cf. Chalmers et al., 1995) and theNTS/area postrema (van Pett et al., personal communica-tion. It is noteworthy that other major targets of descend-ing UCN-ir projections, including the red, basilar pontine,vestibular, inferior olivary, lateral reticular, dorsal col-umn, spinal trigeminal, and cochlear nuclei have beenfound to be sites of substantial CRF-R1 mRNA expression(Potter et al., 1994) and completely lacking in CRF-R2transcripts (Chalmers et al., 1995; van Pett et al., personalcommunication.
Major forebrain sites of CRF-R2 expression in the ventro-medial nucleus of the hypothalamus and medial nucleus ofthe amygdala were found to be sparsely invested withUCN-ir fibers but contained more substantial urotensin-irinputs. While it could be argued that this may reflect a lowtiter of our UCN antiserum or inadequate method sensitiv-ity, the lateral septum contained equally prominent termi-nal fields stained for UCN-ir, as well as urotensin-ir, butthese displayed distinctive topographies, with that of theurotensin-ir input having been shown to mirror the pat-tern of CRF-R2 expression (Vaughan et al., 1995). Collec-tively, these results provide indirect evidence for theexistence of one or more additional CRF-related peptidesin mammalian brain.
UROCORTIN EXPRESSION IN BRAIN 307

Functional considerations
General. The limited central expression of UCN,coupled with the fact that it is strongly expressed in thegut (present observations), immune cells (lymphocytes;Bamberger et al., 1998 and present observations), myocar-dium (Okosi et al., 1998), and placenta (Petraglia et al.,1996) and can act peripherally to exert diverse and power-ful effects on the cardiovascular system (Parkes et al.,1997; Ikeda et al., 1998; Leitch et al., 1998; Schilling et al.,1998) and on immune and inflammatory functions (Turn-bull et al., 1996; Poliak et al., 1997; Agnello et al., 1998;Ando et al., 1998; Singh et al., 1999), UCN appears likelyto prove a considerably more important peripheral peptidethan a central one. The delineation of the central UCNsystem provided here is more one of a peptide involved inmotor function and sensorimotor integration than with thestress-related endocrine, autonomic, and behavioral ac-tions that its relatedness to CRF might suggest.
Although the EW comprised the dominant seat of UCN-irperikarya in rat brain, it should be noted that this is not aunique or defining marker for this cell group. Otherneuropeptides, including cholecystokinin and substance P,have been localized to EW perikarya (Skirboll et al., 1982;Innis and Aghajanian, 1986), including cells identified asprojecting to the spinal cord or the spinal trigeminalnucleus (Phipps et al., 1983; Maciewicz et al., 1984). TheEW has also been reported to house CRF-ir neurons(Chung et al., 1987), but the absence of correspondingtranscripts (Vaughan et al., 1995) makes it likely that thisis attributable to cross-reactivity with UCN. The EW isknown classically as the source of parasympathetic pregan-glionic projections to the ciliary ganglion, which suppliesthe anterior chamber of the eye to regulate the size of thepupil and accommodation of the lens (Loewy, 1990). Insome species, including rat (Martin and Dolivo, 1983) andcat (Loewy et al., 1978), evidence has been adduced toindicate that the EW contributes in only a minor way tothe oculomotor parasympathetic preganglionic projection,with the bulk of the pathway originating from cells distrib-uted more ventrally in the midbrain (see also discussion inLoewy and Saper, 1978).
As noted above, the patterns of descending UCN-irprojections conform to descriptions of EW efferents and tothe extent that they are representative of such would beindicative of modulatory, and perhaps integrative, roles inoculomotor reflexes and associated cerebellar functions.For example, the cerebellum is known to play a role inaccommodation (Hosoba et al., 1978), via projections toEW originating in the deep nuclei. Our findings indicatethat UCN-ir projections, probably of EW origin, are in aposition to reciprocate this, both directly and via wide-spread inputs to precerebellar nuclei. Second, the pupil-lary light reflex is thought to be mediated by retinalafferents routed to the EW by way of the olivary pretectalnuclei (Loewy, 1990). UCN-ir projections are seen to thisand other pretectal structures, as well as the oculomotornuclei. Finally, the spinal trigeminal complex receivesprimary afferents from the anterior chamber of the eye(Panneton and Burton, 1981) and projects to the facialmotor nucleus (Loewy and Saper, 1978; Klooster et al.,1993), aspects of which supply the eyelid musculature(Hinrichsen and Watson, 1984), defining substrates for acomponent of the corneal blink reflex. As noted (Klooster etal., 1993) EW, as well as UCN-ir, projections are in a
position to modulate both central components of thisreflex.
In terms of sheer numbers of neurons, UCN was found tobe most widely expressed, albeit at low-moderate levels, insomatic motor neurons of the caudal brainstem and spinalcord. The significance of this is not clear. Central adminis-tration of UCN has been reported to increase locomotoractivity and, interestingly, orofacial movements, but CRFappears to be at least equally potent in this regard (Joneset al., 1998). It is noteworthy that the distribution of UCNin motoneurons, and, indeed, elsewhere in the brain,shares some striking similarities with that of calcitoningene-related peptide (CGRP; Rosenfeld et al., 1983), whosemultiple proposed actions at the neuromuscular junctionwill doubtless provide a context for evaluating UCN ef-fects. Thus, CGRP has been shown to be capable ofinfluencing neuromuscular transmission by modulatingthe release of acetylcholine (Kimura et al., 1997), theproduction of cholinesterase (Choi et al., 1996), the forma-tion and maintenance of the neuromuscular junction itself(Sala et al., 1995), and as a trophic factor that increasesacetylcholine receptor synthesis (New and Mudge, 1986).
The distribution and morphology of UCN-expressingcells in the lateral superior olive, as well as immunola-beled fibers in the auditory brainstem, suggest that thepeptide is contained principally in neurons that originatethe lateral olivocochlear projection. This rather uniquepathway provides a central efferent innervation of theauditory transduction apparatus and, by analogy with thebetter studied medial olivocochlear path, is generallythought to serve a protective function through a capacityto decrease the sensitivity of hair cells to acoustic stimuli(Warr et al., 1986). Lateral olivocochlear neurons areknown to synthesize acetylcholine and GABA (Vetter et al.,1991), and markers for these phenotypes coexist exten-sively with one another and with ones for dopamine, opiatepeptides, and, again, CGRP in the lateral superior olive(Safieddine et al., 1997). Unfortunately, there is a lack ofinformation regarding the role of any neuropeptide in thisprojection.
Magnocellular neurosecretory neurons are character-ized by their capacity to synthesize the nonapeptide hor-mones oxytocin and vasopressin and release them fromterminals in the posterior pituitary with access to thegeneral circulation. By virtue of its pressor, antidiuretic,and natriuretic effects, vasopressin plays important rolesin the regulation of the pressure, volume, and ioniccomposition of the blood. Oxytocin’s best known functionsare in promoting milk letdown in lactating dams anduterine contraction as occurs during parturition, but italso synergizes with vasopressin at the kidney to promotesodium excretion (for review, see Cunningham and Saw-chenko, 1991). Magnocellular oxytocin and vasopressinneurons are well known to produce a plethora of coexistingneuropeptides, expressed at concentrations typically 3–4orders of magnitude lower than those of the nonapeptides,making it unlikely that their concentrations in plasmaattributable to magnocellular origin achieve biologicallysignificant (i.e., hormone-like) levels (Brownstein andMezey, 1986; Sawchenko et al., 1992). Instead, such coex-isting peptides are thought to exert modulatory (autocrineor paracrine) effects at the level of the posterior pituitaryon the release of oxytocin and/or vasopressin. In view of itslimited expression in the magnocellular system, UCNappears most likely to fulfill such a role. Observations that
308 J.C. BITTENCOURT ET AL.

its expression in the magnocellular system is upregulatedin response to salt loading or water deprivation (Hara etal., 1997a,b) provide clues as to the nature of its involve-ment, but the targets of its presumed influence(s) on theneurohypophysis have not yet been evaluated.
Stress-related. The discovery of UCN has spawnedevidence to suggest that this peptide may participate instress-related neuroendocrine, autonomic, and behavioralfunctions originally thought to be the province of CRF,offering some potential for resolving long-standing in-stances of ligand-receptor mismatch at stress-related sitesof peptide action in the central CRF system. An involve-ment in pituitary-adrenal regulation is suggested by thefindings that UCN is more potent than CRF in promotingACTH secretion (Vaughan et al., 1995; Asaba et al., 1998),along with indications that it may be expressed in parvocel-lular neurons of the PVH (Kozicz et al., 1998) and strongevidence for expression in the human anterior pituitary(Iino et al., 1997). However, passive immunoneutralizationof UCN has been found to affect neither resting plasmaACTH levels nor the increased titers seen in response tofootshock stress (Turnbull et al., 1999) or adrenalectomy(Masuzawa et al., 1999). This is compatible with thepresent findings, which suggest that any direct involve-ment of UCN in regulating the hypothalamo-pituitary-adrenal axis is apt to be achieved by way of UCN-ir inputsto the paraventricular nucleus, which would not be ex-pected to be accessed by systemically administered immu-noglobulins in immunoneutralization experiments.
Central CRF administration is well known to elicitbehavioral and autonomic (sympathoadrenal) responsesas are commonly seen in response to stress (e.g., Brown etal., 1985; Brown and Fisher, 1985; Fisher, 1993), and sucheffects have provided a cornerstone of support for the viewthat this peptide system serves to integrate diverse physi-ological and behavioral response avenues that may becalled into play under threatening circumstances. Intraven-tricular injection of UCN has been shown to produce a mildpressor response (Spina et al., 1996) and to inhibit mitogen-stimulated splenic lymphocyte proliferation (Okamoto etal., 1998), an effect also seen in response to CRF (Straus-baugh and Irwin, 1992), which is mediated by the sympa-thetic nervous system (Okamoto et al., 1998). UCN-irprojections that ramify within the intermediolateral col-umn of thoracic and lumbar spinal cord present them-selves as likely substrates for such responses, although thefact that the NTS receives a UCN-containing input and isknown to project to a core group of structures that arepivotally involved in various facets of central autonomiccontrol (e.g., Sawchenko, 1983; Saper, 1995) defines sev-eral indirect pathways through which autonomic outflowmay be subject to modulation by this peptide. Althoughthere remains a question as to whether hypothalamic cellgroups may contribute to the spinal UCN-ir projection,there would seem little question but that the EW is thedominant source of this. Using a viral transneuronaltracing approach, Loewy and colleagues (1978) have iden-tified the EW as contributing to the premotor innervationof the stellate ganglion (which supplies the heart), as wellas other autonomic targets (Jansen et al., 1992; 1995,Smith et al., 1998).
Of the early forays into UCN biology, clearly the greatestinterest has been sparked by the observations that cen-trally administered UCN is more potent than CRF ininhibiting food intake, but less so in inducing anxiety-like
effects and locomotor activity (Spina et al., 1996), suggest-ing a clear segregation of stress-related function to the twopeptide systems. UCN-ir projections have been identifiedin several cell groups that may be considered as potentialsites of action for the anorexic effect. These include theparaventricular nucleus, where local microinjections ofCRF have been found to inhibit feeding, whereas compa-rable placements in the ventromedial nucleus or lateralhypothalamic area do not (Krahn et al., 1988).Also notewor-thy in this regard is the dorsal raphe nucleus, whichreceives a UCN-ir input and is a prominent site of CRF-R2expression (Chalmers et al., 1995). The serotonergic sys-tem has itself been prominently implicated in the inhibi-tory control of food intake (e.g., Simansky, 1996) and is apopular target for appetite-suppressing pharmaceuticals.The paraventricular nucleus projects to the EW (Swanson,1977), receives a serotonergic input from the dorsal raphe(Sawchenko et al., 1983), and has also been implicated as asite at which the appetite-suppressing effects of serotoninmay be exerted (Morley et al., 1985; Simansky, 1996),raising the possibility that the effects of UCN and seroto-nin on appetitive behavior may tap into a common system.
With respect to feeding-related effects of UCN, it is alsoworthy of mention that the cranial nerve motor nucleisubserving orofacial and upper alimentary function broadlyexpress UCN, and several appear to receive substantialUCN-ir inputs. In view of evidence to indicate that theorofacial reflexes that mediate the acceptance or rejectionof taste substances are organized at brainstem levels (Grilland Norgren, 1978a,b), the possibility that appetitiveeffects of central UCN may have some basis in influencesexerted at the levels of the final common path would seemworthy of consideration (Sawchenko, 1998).
Knowledge of the range of functions that may be associ-ated with UCN-ir elements in and around the EW willrequire a fuller appreciation of the organization of theafferent connections of this cell group. The single study tohave explicitly addressed this topic, a survey of brainsteminputs to EW in cat (Breen et al., 1983), identified potentialsources of input arising from many sites highlighted hereas targets of UCN-ir projections. This included neurons inthe NTS and parabrachial nuclei, defining potential sub-strates for visceral sensory control of EW mechanisms, aswell as ones arising from the brainstem raphe nuclei. Thelatter may be relevant to the findings that nociceptiveunits have been identified in the EW region (Innis andAghajanian, 1986) and that the EW has been reported todisplay very prominent activational responses (immediate-early gene induction) to noxious visceral stimuli (Lanteri-Minet et al., 1993). Descending inputs to EW have not beensystematically examined but are known to include aninput from the paraventricular nucleus of the hypothala-mus (Swanson, 1977), a pivotal structure in central auto-nomic and neuroendocrine control. Thus, although severaldifferent authors have suggested that the EW is impli-cated in the regulation of central autonomic function, acohesive view of its place in this context is lacking.
ACKNOWLEDGMENTS
W.W.V. and P.E.S. were recipients of NIH grant DK-26741. They are Investigators of the Foundation for Re-search and the Foundation for Medical Research, respec-tively, for which this work was partly conducted. J.C.B., avisiting scientist on leave from the Department of Anatomy-
UROCORTIN EXPRESSION IN BRAIN 309

ICB, University of Sao Paulo, Brazil, was supported bygrants from CNPq (203301/87-0 and 300583/91-5—J.C.B.is a CNPq investigator). The current address for R.A.R. is:MCP Hahnemann University School of Medicine, Philadel-phia, PA. We are grateful to Belle Wamsley and KrisTrulock for excellent editorial and photography/graphicsassistance, respectively.
LITERATURE CITED
Abercrombie M. 1946. Estimation of nuclear populations from microtomepopulations from microtome sections. Anat Rec 94:239–247.
Agnello D, Bertini R, Sacco S, Meazza C, Villa P, Ghezzi P. 1998.Corticosteroid-independent inhibition of tumor necrosis factor produc-tion by the neuropeptide urocortin. Am J Physiol 275:E757–E762.
Ando T, Rivier J, Yanaihara H, Arimura A. 1998. Peripheral corticotropin-releasing factor mediates the elevation of plasma IL-6 by immobiliza-tion stress in rats. Am J Physiol 275:R1461–R1467.
Asaba K, Makino S, Hashimoto K. 1998. Effect of urocortin on ACTHsecretion from rat anterior pituitary in vitro and in vivo: comparisonwith corticotropin-releasing hormone. Brain Res 806:95–103.
Aschoff A, Ostwald J. 1988. Distribution of cochlear efferents and olivo-collicular neurons in the brainstem of rat and guinea pig. Exp Brain Res71:241–251.
Bamberger CM, Wald M, Bamberger AM, Ergun S, Beil FU, Schulte HM.1998. Human lymphocytes produce urocortin, but not corticotropin-releasing hormone. J Clin Endocrinol Metab 83:708–711.
Bentivoglio M, Kuypers HGJM, Catsman-Berrevoets CE, Loewe H, DannO. 1980. Two new fluorescent retrograde neuronal tracers which aretransported over long distances. Neurosci Lett 18:25–30.
Bittencourt JC, Presse F, Arias C, Peto C, Vaughan J, Vale W, SawchenkoPE. 1992. The melanin-concentrating hormone system of the rat brain:an immuno- and hybridization histochemical characterization. J CompNeurol 319:218–245.
Bittencourt JC, Vaughan J, Vale W, Sawchenko PE. 1995. Evidence for theexistence in rat brain of a sauvigine/urotensin-like peptide, distinctfrom CRF and in a position to interact with CRF receptors. Soc NeurosciAbstr 21:359.
Breen LA, Burde RM, Loewy AD. 1983. Brainstem connections to theEdinger-Westphal nucleus of the cat: a retrograde tracer study. BrainRes 261:303–306.
Brown M. 1986. Corticotropin releasing factor: central nervous system sitesof action. Brain Res 399:10–14.
Brown MC, Liberman MC, Benson TE, Ryugo DK. 1988. Brainstembranches from olivocochlear axons in cats and rodents. J Comp Neurol278:591–603.
Brown MR, Fisher LA. 1985. Corticotropin-releasing factor: effects on theautonomic nervous system and visceral system. Fed Proc 44:243–248.
Brown MR, Fisher LA, Webb V, Vale WW, Rivier JE. 1985. Corticotropin-releasing factor: a physiologic regulator of adrenal epinephrine secre-tion. Brain Res 328:355–357.
Brownstein MJ, Mezey E. 1986. Multiple chemical messengers in hypotha-lamic magnocellular neurons. In: T Hokfelt, K Fuxe, B Pernow, editors.Progress in brain research—coexistence of neuronal messengers: a newprinciple in chemical transmission. New York: Elsevier, p 161–168.
Ceccatelli S, Villar MJ, Goldstein M, Hokfelt T. 1989. Expression of c-Fosimmunoreactivity in transmitter-characterized neurons after stress.Proc Natl Acad Sci USA 86:9569–9573.
Ceccatelli S, Cortes R, Hokfelt T. 1991. Effect of reserpine and colchicine onneuropeptide mRNA levels in the rat hypothalamic paraventricularnucleus. Mol Brain Res 9:57–69.
Cepoi D, Sutton S, Arias C, Sawchenko PE, Vale WW. 1999. Ovine genomicurocortin: cloning, pharmacologic characterization, and distribution ofcentral mRNA. Mol Brain Res 68:109–118.
Chadwick DJ, Marsh J, Ackrill K. 1993. Corticotropin-releasing factor:Ciba Foundation Symposium 172. London: John Wiley.
Chalmers DT, Lovenberg TW, De Souza EB. 1995. Localization of novelcorticotropin-releasing factor receptor (CRF2) mRNA expression tospecific sub-cortical nuclei in rat brain: comparison with CRF1 receptormRNA expression. J Neurosci 15:6340–6350.
Choi RC, Leung PW, Dong TT, Wan DC, Tsim KW. 1996. Calcitoningene-related peptide increases the expression of acetylcholinesterase incultured chick myotubes. Neurosci Lett 217:165–168.
Chung RY, Mason P, Strassman A, Maciewicz R. 1987. Edinger-Westphal
nucleus: cells that project to spinal cord contain corticotropin-releasingfactor. Neurosci Lett 83:13–19.
Cox KH, DeLeon DV, Angerer LM, Angerer RC. 1984. Detection of mRNAsin sea urchin embryos by in situ hybridization using asymmetric RNAprobes. Dev Biol 101:485–502.
Cunningham ET Jr, Sawchenko PE. 1991. Reflex control of magnocellularvasopressin and oxytocin secretion. TINS 14:406–411.
Elverland HH. 1978. Ascending and intrinsic projections of the superiorolivary complex in the cat. Exp Brain Res 32:117–134.
Fisher LA. 1993. Central actions of corticotropin-releasing factor onautonomic nervous activity and cardio-vascular functioning. In: DJChadwick, J Marsh, K Ackrill, editors. Corticotropin-releasing factor:Ciba Foundation Symposium 172. London: John Wiley, p 243–257.
Glendenning KK, Masterton RB. 1983. Acoustic chiasm: efferent projec-tions of the lateral superior olive. J Neurosci 3:1521–1537.
Grill HJ, Norgren R. 1978a. The taste reactivity test. I. Mimetic responsesto gustatory stimuli in neurologically normal rats. Brain Res 143:263–279.
Grill HJ, Norgren R. 1978b. The tast reactivity test. II. Mimetic responsesto gustatory stimuli in chronic thalamic and chronic decerebrate rats.Brain Res 143:281–297.
Hara Y, Ueta Y, Isse T, Kabashima N, Shibuya I, Hattori Y, Yamashita H.1997a. Increase of urocortin-like immunoreactivity in the rat hypo-thalamo-neurohypophysial system after salt loading and hypophysec-tomy. Neurosci Lett 227:127–130.
Hara Y, Ueta Y, Isse T, Kabashima N, Shibuya I, Hattori Y, Yamashita H.1997b. Increase of urocortin-like immunoreactivity in the rat supraop-tic nucleus after dehydration but not food deprivation. Neurosci Lett229:65–68.
Herkenham M. 1978. The connections of the nucleus reuniens thalami:evidence for a direct thalamo-hippocampal pathway in the rat. J CompNeurol 177:589–610.
Herkenham M. 1987. Mismatches between neurotransmitter and receptorlocalizations in brain: observations and implications. Neuroscience23:1–38.
Hinrichsen CFL, Watson CD. 1984. The facial nucleus of the rat. Represen-tation of facial muscles revealed by retrograde transport of horseradishperoxidase. Anat Rec 209:407–415.
Hosoba M, Bando T, Tsukahara N. 1978. The cerebellar control of accommo-dation of the eye in the cat. Brain Res 153:495–505.
Iino K, Sasano H, Oki Y, Andoh N, Shin R-W, Kitamoto T, Totsune K,Takahashi K, Suzuki H, Nagura H, Yoshimi T. 1997. Urocortin expres-sion in human pituitary gland and pituitary adenoma. J Clin Endocri-nol Metab 82:3842–3850.
Ikeda KT, Togo K, Sato S, Ebisawa T, Tokudome G, Hosoya T, Harada M,Nakagawa O, Nakao K. 1998. Urocortin, a newly identified corticotropin-releasing factor-related mammalian peptide, stimulates atrial natri-uretic peptide and brain natriuretic peptide secretions from neonatalrat cardiomyocytes. Biochem Biophys Res Commun 250:298–304.
Imaki J, Imaki T, Vale W, Sawchenko PE. 1991. Distribution of corticotropin-releasing factor mRNA and immunoreactivity in the central auditorysystem of the rat. Brain Res 547:28–36.
Innis RB, Aghajanian GK. 1986. Cholecystokinin-containing and nocicep-tive neurons in rat Edinger-Westphal nucleus. Brain Res 363:230–238.
Jansen AS, Ter Horst GJ, Mettenleiter TC, Loewy AD. 1992. CNS cellgroups projecting to the submandibular parasympathetic preganglionicneurons in the rat: a retrograde transneuronal viral cell body labelingstudy. Brain Res 572:253–260.
Jansen ASP, Wessendorf MW, Loewy AD. 1995. Transneuronal labeling ofCNS neuropeptide and monoamine neurons after pseudorabies virusinjections into the stellate ganglion. Brain Res 683:1–24.
Jones DN, Kortekaas R, Salde PD, Middlemiss DN, Hagan JJ. 1998. Thebehavioural effects of corticotropin-releasing factor-related peptides inrats. Psychopharmacology (Berl) 138:124–132.
Kimura I, Okazaki M, Nojima H. 1997. Mutual dependence of calcitonin-gene related peptide and acetylcholine release in neuromuscular prepa-rations. Eur J Pharmacol 330:123–128.
Klooster J, Beckers HJM, Vrensen GFJM, van der Want JJL. 1993. Theperipheral and central projections of the Edinger-Westphal nucleus inthe rat. A light and electron microscopic tracing study. Brain Res632:260–273.
Kozicz T, Yanaihara H, Arimura A. 1998. Distribution of urocortin-likeimmunoreactivity in the central nervous system of the rat. J CompNeurol 391:1–10.
Krahn DD, Gosnell BA, Levine AS, Morley JE. 1988. Behavioral effects of
310 J.C. BITTENCOURT ET AL.

corticotropin-releasing factor: localization and characterization of cen-tral effects. Brain Res 443:63–69.
Lanteri-Minet M, Isnardon P, De Pommery J, Menetrey D. 1993. Spinal andhindbrain structures involved in visceroception and visceronociceptionas revealed by the expression of Fos, Jun and Krox-24 proteins.Neuroscience 55:737–753.
Lederis K, Letter A, McMaster D, Moore G, Schlesinger D. 1982. Completeamino acid sequence of urotensin I, a hypotensive and corticotropin-releasing neuropeptide from Catostomus. Science 218:162–164.
Leitch IM, Boura AL, Botti C, Read MA, Walters WA, Smith R. 1998.Vasodilator actions of urocortin and related peptides in the humanperfused placenta in vitro. J Clin Endocrinol Metab 83:4510–4513.
Loewy AD. 1990. Autonomic control of the eye. In: AD Loewy, KM Spyer,editors. Central regulation of autonomic functions. New York: OxfordUniversity Press, p 268–285.
Loewy AD, Saper CB. 1978. Edinger-Westphal nucleus: projections to thebrain stem and spinal cord in the cat. Brain Res 150:1–27.
Loewy AD, Saper CB, Yamodis ND. 1978. Re-evaluation of the efferentprojections of the Edinger-Westphal nucleus. Brain Res 141:153–159.
Maciewicz R, Phipps BS, Grenier J, Poletti CE. 1984. Edinger-Westphalnucleus: cholecystokinin immunocytochemistry and projections to spi-nal cord and trigeminal nucleus in the cat. Brain Res 299:139–145.
Martin X, Dolivo M. 1983. Neuronal and transneuronal tracing in thetrigeminal system of the rat using herpes virus suis. Brain Res273:253–276.
Masuzawa M, Oki Y, Ozawa M, Watanabe F, Yoshimi T. 1999. Corticotropin-releasing factor but not urocortin is involved in adrenalectomy-inducedadrenocorticotropin release. J Neuroendocrinol 11:71–74.
Montecucchi PC, Henschen A. 1981. Amino acid composition and sequenceanalysis of sauvagine, a new active peptide from the skin of Phyllome-dusa sauvagei. Int J Peptide Protein Res 18:113–120.
Morley JE, Levine AS, Gosnell BA, Krahn DD. 1985. Peptides as centralregulators of feeding. Brain Res Bull 14:511–519.
Mundy WR, Tilson HA. 1990. Neurotoxic effects of colchicine. Neurotoxicol-ogy 1:539–547.
Nahon JL, Presse F, Bittencourt JC, Sawchenko P, Vale W. 1989. The ratmelanin-concentrating hormone mRNA encodes multiple putative neu-ropeptides coexpressed in the dorsolateral hypothalamus. Endocrinol-ogy 125:2056–2065.
New HV, Mudge AW. 1986. Calcitonin gene-related peptide regulatesmuscle acetylcholine receptor synthesis. Nature 323:809–811.
Okamoto S, Ishikawa I, Kimura K, Saito M. 1998. Potent suppressiveeffects of urocortin on splenic lymphocyte activity in rats. NeuroReport9:4035–4039.
Okawara Y, Morley SD, Burzio LO, Xwiers H, Lederis K, Richter D. 1988.Cloning and sequence analysis of cDNA for corticotropin-releasingfactor precursor from the teleost fish Catostomus commersoni. ProcNatl Acad Sci USA 85:8439.
Okosi A, Brar BK, Chan M, D’Souza L, Smith E, Stephanou A, LatchmanDS, Chowdrey HS, Knight RA. 1998. Expression and protective effectsof urocortin in cardiac myocytes. Neuropeptides 32:167–171.
Panneton WM, Burton H. 1981. Corneal and periocular representationwithin the trigeminal sensory complex in the cat studied with transgan-glionic transport of horseradish peroxidase. J Comp Neurol 199:327–344.
Parkes DG, Vaughan J, Rivier J, Vale W, May CN. 1997. Cardiac ionotropicactions of urocortin in conscious sheep. Am J Physiol 272:H2115–H2122.
Paxinos G, Watson C. 1986. The Rat Brain in Stereotaxic Coordinates, 2nded. New York: Academic Press.
Petraglia F, Florio P, Gallo R, Simoncini T, Saviozzi M, Di Blasio AM,Vaughan J, Vale W. 1996. Human placenta and fetal membranesexpress human urocortin mRNA and peptide. J Clin Endocrinol Metab81:3807–3810.
Phipps BS, Maciewicz R, Sandrew BB, Poletti CE, Foote WE. 1983.Edinger-Westphal neurons that project of spinal cord contain substanceP. Neurosci Lett 36:125–131.
Pickard GE. 1985. Bifurcating axons of retinal ganglion cells terminate inthe hypothalamic suprachiasmatic nucleus and the intergeniculateleaflet of the thalamus. Neurosci Lett 55:211–217.
Poliak S, Mor F, Conlon P, Wong T, Ling N, Rivier J, Vale W, Steinman L.1997. Stress and autoimmunity: the neuropeptides corticotropin-releasing factor and urocortin suppress encephalomyelitis via effects onboth the hypothalamic-pituitary-adrenal axis and the immune system.J Immunol 158:5751–5756.
Potter E, Sutton S, Donaldson C, Chen R, Perrin M, Lewis K, SawchenkoPE, Vale W. 1994. Distribution of corticotropin-releasing factor receptormRNA expression in the rat brain and pituitary. Proc Natl Acad SciUSA 91:8777–8781.
Rasmussen GL. 1946. The olivary peduncle and other fiber projections ofthe superior olivary complex. J Comp Neurol 84:141–219.
Rosenfeld MG, Mermod J-J, Amara S, Swanson LW, Sawchenko PE, RivierJ, Vale WW, Evans RM. 1983. Production of a novel neuropeptideencoded by the calcitonin gene via tissue-specific RNA processing.Nature 304:129–135.
Roste RK, Dietrichs E. 1988. Cerebellar cortical and nuclear afferents fromthe Edinger-Westphal nucleus in the cat. Anat Embryol 178:59–65.
Safieddine S, Prior AMS, Eybalin M. 1997. Choline acetyltransferase,glutamate decarboxylase, tyrosine hydroxylase, calcitonin gene-relatedpeptide and opioid peptides coexist in lateral efferent neurons of rat andguinea-pig. Eur J Neurosci 9:356–367.
Sala C, Andreose JS, Fumagalli G, Lomo T. 1995. Calcitonin gene-relatedpeptide: possible role in formation and maintenance of neuromuscularjunctions. J Neurosci 15:520–528.
Saper CB. 1995. Central autonomic system. In: G. Paxinos, editor. The RatNervous System, 2nd ed. San Diego: Academic Press, p 107–128.
Saper CB, Loewy AD, Swanson LW, Cowan WM. 1976. Direct hypothalamo-autonomic connections. Brain Res 117:305–312.
Sawchenko PE. 1983. Central connections of the sensory and motor nucleiof the vagus nerve. J Auton Nerv Syst 9:13–26.
Sawchenko PE. 1987. Evidence for differential regulation of CRF- andvasopressin-immunoreactivities in parvocellular neurosecretory andautonomic-related projections of the paraventricular nucleus. BrainRes 437:253–263.
Sawchenko PE. 1998. Toward a new neurobiology of energy balance,appetite, and obesity: the anatomists weigh in. J Comp Neurol 402:435–441.
Sawchenko PE, Swanson LW. 1982. Immunohistochemical identification ofneurons in the paraventricular nucleus of the hypothalamus thatproject to the medulla or to the spinal cord in the rat. J Comp Neurol205:260–272.
Sawchenko PE, Swanson LW. 1983. The organization of forebrain afferentsto the paraventricular and supraoptic nuclei of the rat. J Comp Neurol218:121–144.
Sawchenko PE, Swanson LW. 1989. Organization of CRF immunoreactivecells and fibers in the rat brain: Immunohistochemical studies. In: EBDe Souza, CB Nemeroff, editors. Corticotropin-releasing Factor: Basicand Clinical Studies of a Neuropeptide. Boca Raton: CRF Press, p29–51.
Sawchenko PE, Swanson LW, Steinbusch HWM, Verhofstad AAJ. 1983. Thedistribution and cells of origin of serotonergic inputs to the paraventricu-lar and supraoptic nuclei of the rat. Brain Res 277:355–360.
Sawchenko PE, Cunningham ET Jr, Mortrud MT, Pfeiffer SW, Gerfen CR.1990. Phaseolus vulgaris-leucoagglutinin (PHA-L) anterograde axonaltransport technique. Methods Neurosci 3:247–260.
Sawchenko PE, Imaki T, Vale W. 1992. Colocalization of neuroactivesubstances in the endocrine hypothalamus. In: DJ Chadwick, J Marsh,editors. Functional Anatomy of the Neuroendocrine Hypothalamus:Ciba Foundation Symposium No. 168. London: John Wiley, p 16–42.
Schilling L, Kanzler C, Schmiedek P, Ehrenreich H. 1998. Characterizationof the relaxant action of urocortin, a new peptide related to corticotropin-releasing factor in the rat isolated basilar artery. Br J Pharmacol125:1164–1171.
Shu S, Ju G, Fan L. 1988. The glucose oxidase-DAB-nickel method inperoxidase histochemistry of the nervous system. Neurosci Lett 85:169–171.
Simansky KJ. 1996. Serotonergic control of the organization of feeding andsatiety. Behav Brain Res 73:37–42.
Simmons DM, Arriza JL, Swanson LW. 1989. A complete protocol for in situhybridization of messenger RNAs in brain and other tissues withradiolabeled single-stranded RNA probes. J Histotechnol 12:169–181.
Singh LK, Boucher W, Pang X, Letourneau R, Seretakis D, Green M,Theoharides TC. 1999. Potent mast cell degranulation and vascularpermeability triggered by urocortin through activation of corticotropin-releasing hormone receptors. J Pharmacol Exp Ther 288:1349–1356.
Skirboll L, Hokfelt T, Rehfeld J, Cuello AC, Dockray G. 1982. Coexistence ofsubstance P- and cholecystokinin-like immunoreactivity in neurons ofthe mesencephalic periaqueductal central gray. Neurosci Lett 28:35–39.
Smith JE, Jansen ASP, Gilbey MP, Loewy AD. 1998. CNS cell groupsprojecting to sympathetic outflow of tail artery: neural circuits involvedin heat loss in the rat. Brain Res 786:153–164.
UROCORTIN EXPRESSION IN BRAIN 311

Spina M, Merlo-Pich E, Chan RKW, Basso AM, Rivier J, Vale W, Koob GF.1996.Appetite-suppressing effects of urocortin, a CRF-related neuropep-tide. Science 273:1561–1564.
Stenzel-Poore MP, Heldwein KA, Stenzel P, Lee S, Vale WW. 1992.Characterization of the genomic corticotropin-releasing factor genefrom Xenopus laevis: two members of the CRF family exist in amphib-ians. Mol Endocrinol 6:1716.
Strausbaugh H, Irwin M. 1992. Central corticotropin-releasing hormonereduces cellular immunity. Brain Behav Immun 6:11–17.
Swanson LW. 1977. Immunohistochemical evidence for a neurophysin-containing autonomic pathway arising in the paraventricular nucleusof the hypothalamus. Brain Res 128:346–353.
Swanson LW, Kuypers HGJM. 1980. The paraventricular nucleus of thehypothalamus: cytoarchitectonic subdivisions and organization of pro-jections to the pituitary, dorsal vagal complex, and spinal cord asdemonstrated by retrograde fluorescence double-labeling methods. JComp Neurol 194:555–570.
Takahashi K, Totsune K, Sone M, Murakami O, Satoh F, Arihara Z, SasanoH, Iino K, Mouri T. 1998. Regional distribution of urocortin-likeimmunoreactivity and expression of urocortin mRNA in the humanbrain. Peptides 19:643–647.
Tiedge H, Bloom FE, Richter D. 1999. RNA, whither goest thou? Science283:186–187.
Turnbull AV, Rivier C. 1997. Corticotropin-releasing factor (DRF) andendocrine responses to stress: CRF receptors, binding protein, andrelated peptides. Proc Soc Exp Biol Med 215:1–10.
Turnbull AV, Vale W, Rivier C. 1996. Urocortin, a corticotropin-releasingfactor-related mammalian peptide, inhibits edema due to thermalinjury in rats. Eur J Pharmacol 303:213–216.
Turnbull AV, Vaughan J, Rivier JE, Vale WW, Rivier C. 1999. Urocortin isnot a significant regulator of intermittent electrofootshock-inducedadrenocorticotropin secretion in the intact male rat. Endocrinology140:71–78.
Vale W, Speiss J, Rivier C, Rivier J. 1981. Characterization of a 41-residueovine hypothalamic peptide that stimulates secretion of corticotropinand b-endorphin. Science 213:1394–1397.
van Leeuwen FW, de Raay C, Swabb DF, Fisser B. 1979. The localization ofoxytocin, vasopressin, somatostatin and luteinizing hormone releasinghormone in the rat neurohypophysis. Cell Tissue Res 202:189–201.
Vaughan J, Donaldson C, Bittencourt J, Perrin MH, Lewis K, Sutton S,Chan R, Turnbull AV, Lovejoy D, Rivier C, Rivier J, Sawchenko PE, ValeW. 1995. Urocortin, a mammalian neuropeptide related to fish uroten-sin I and to corticotropin-releasing factor. Nature 378:287–292.
Vetter DE, Mugnaini E. 1992. Distribution and dendritic features of threegroups of rat olivocochlear neurons. Anat Embryol 185:1–16.
Vetter DE, Adams JC, Mugnaini E. 1991. Chemically distinct rat olivoco-chlear neurons. Synapse 7:21–43.
Warr WB, Guinan JJ Jr, White JS. 1986. Organization of the efferent fibers:the lateral and medial olivocochlear systems. In: RA Altschuler, RPBobbin, DW Hoffman, editors. Neurobiology of Hearing: the Cochlea.New York: Raven Press, p 333–348.
Webster WR. 1995. Auditory system. In: G Paxinos, editor. The Rat NervousSystem, 2nd ed. San Diego: Academic Press, pp 797–831.
Weiner JL, Buhler AV, Whatley VJ, Harris RA, Dunwiddie TV. 1998.Colchicine is a competitive antagonist at human recombinant gamma-aminobutyric acid A receptors. J Pharmacol Exp Ther 284:95–102.
Wong ML, al-Shekhlee A, Bongiorno PB, Esposito A, Khatri P, SternbergEM, Gold PW, Licinio J. 1996. Localization of urocortin messenger RNAin rat brain and pituitary. Mol Psychiatry 1:307–312.
Yamamoto H, Maeda T, Fujimura M, Fujimiya M. 1998. Urocortin-likeimmunoreactivity in the substantia nigra, ventral tegmental area inEdinger-Westphal nucleus of rat. Neurosci Lett 243:21–24.
Zamboni L, De Martino C. 1967. Buffered picric acid-formaldehyde: a new,rapid fixative for electron microscopy. J Cell Biol 35:148A.
312 J.C. BITTENCOURT ET AL.

![Cytochemical Studies of Corticotropin-Releasing Factor (CRF) Receptors in Anterior Lobe Corticotropes: Binding, Glucocorticoid Regulation, and Endocytosis of [Biotinyl-Ser 1 ]CRF*](https://img.pdfslide.net/doc/110x75/6354020725654d63c100dab3/cytochemical-studies-of-corticotropin-releasing-factor-crf-receptors-in-anterior.jpg)



























