Embed Size (px)
Citation preview

BASIC NEUROSCIENCES, GENETICS AND IMMUNOLOGY - ORIGINAL ARTICLE
From caffeine to fish waste: amine compounds present in foodand drugs and their interactions with primary amine oxidase
Aldo Olivieri • Daniel Rico • Zhied Khiari •
Gary Henehan • Jeff O’Sullivan • Keith Tipton
Received: 29 September 2010 / Accepted: 16 February 2011
� Springer-Verlag 2011
Abstract Tissue bound primary amine oxidase (PrAO)
and its circulating plasma-soluble form are involved,
through their catalytic activity, in important cellular roles,
including the adhesion of lymphocytes to endothelial cells
during various inflammatory conditions, the regulation of
cell growth and maturation, extracellular matrix deposition
and maturation and glucose transport. PrAO catalyses the
oxidative deamination of several xenobiotics and has been
linked to vascular toxicity, due to the generation of cyto-
toxic aldehydes. In this study, a series of amines and
aldehydes contained in food and drugs were tested via a
high-throughput assay as potential substrates or inhibitors
of bovine plasma PrAO. Although none of the compounds
analyzed were found to be substrates for the enzyme, a
series of molecules, including caffeine, the antidiabetics
phenformin and tolbutamide and the antimicrobial pent-
amidine, were identified as PrAO inhibitors. Although the
inhibition observed was in the millimolar and micromolar
range, these data show that further work will be necessary
to elucidate whether the interaction of ingested biogenic or
xenobiotic amines with PrAO might adversely affect its
biological roles.
Keywords Primary amine oxidase � Semicarbazide-
sensitive amine oxidase � Xenobiotic(s) � Enzyme
inhibition
Abbreviations
PrAO Primary amine oxidase
SSAO Semicarbazide-sensitive amine oxidase
TPQ 3,4,5-Trihydroxyphenylalanine quinone
DAO Diamine oxidase
Introduction
Primary amine oxidase (PrAO) is the accepted name for the
copper-containing enzyme (EC 1.4.3.21) that catalyses the
oxidative deamination of endogenous and exogenous pri-
mary amines, according to the overall reaction:
RCH2NH2 þ O2 þ H2O �!PrAORCHOþ NH3 þ H2O2
It was previously classified with a group of other enzymes
as semicarbazide-sensitive amine oxidase (SSAO), EC
1.4.3.6 (Boyce et al. 2009). The sensitivity to inhibition by
carbonyl-group reagents, such as semicarbazide, is a result of
the presence of a 6-hydroxydopa: 2,4,5-trihydroxyphen-
ylalanine quinone (TPQ) residue as the redox cofactor. The
enzyme is active towards aliphatic and aromatic primary
amines but secondary amines, such as adrenaline are not
oxidized. The important physiological substrates are
believed to include methylamine and aminoacetone (see
O’Sullivan et al. 2004).
A. Olivieri (&) � K. Tipton
School of Biochemistry and Immunology, Trinity College,
Dublin 2, Ireland
e-mail: [email protected]
D. Rico � Z. Khiari � G. Henehan
Dublin Institute of Technology, School of Food Science and
Environmental Health, Cathal Brugha Street, Dublin 1, Ireland
J. O’Sullivan
Dublin Dental School and Hospital, Trinity College,
Dublin 2, Ireland
Present Address:A. Olivieri
Department of Pharmacology, University of Alberta,
9-70 Medical Sciences Building, Edmonton,
AB T6G 2H7, Canada
123
J Neural Transm
DOI 10.1007/s00702-011-0611-z

PrAO is a membrane-bound protein in mammalian
species (Lewinsohn 1984), although a plasma soluble form
of the enzyme also exists, which results from the proteo-
lytic cleavage of membrane-bound PrAO (Stolen et al.
2004; Ekblom et al. 2000). The amount of the plasma form
varies greatly between species. Although it is normally low
in the human, it has been shown to be increased in a
number of disease states (see O’Sullivan et al. 2004). The
products of the PrAO catalysed reaction, especially
hydrogen peroxide, have important cellular roles, including
the regulation of cell growth and maturation, extracellular
matrix deposition and maturation and glucose transport
capacity. It has been demonstrated that the generation of
H2O2 during the oxidation of amine substrates by PrAO
stimulates glucose uptake by the recruitment of insulin
receptors to the cell-surface (for review, see O’Sullivan
et al. 2004). In some tissues, during pathological insult,
PrAO functions as a vascular adhesion protein, VAP-1, that
mediates the slow rolling and adhesion of lymphocytes to
endothelial cells, contributing to their transmigration into
sites of inflammation (Jalkanen and Salmi 2001). H2O2
generated by substrate oxidation also appears to be nec-
essary for this adhesion process to occur (O’Sullivan et al.
2007; Jalkanen et al. 2007; Olivieri and Tipton 2011).
Since all these processes involve the catalytic activity of
the enzyme, they are inhibited by PrAO inhibitors. They
have also been shown to be attenuated by knock-out of the
PrAO (AOC3) gene (Bour et al. 2007; Stolen et al. 2005).
The active site of the tissue-bound PrAO is located in
the extracellular domain (Jakobsson et al. 2005) making the
enzyme, together with its circulating plasma-soluble form,
a scavenger of potentially toxic amines in the blood. Food
products may contain a wide variety of biogenic amines
(see Table 1), which are often a consequence of enzymatic
decarboxylation (by microorganisms) of the corresponding
amino acids. Such amines may be sources of nitrogen and
Table 1 A selection of amines and amine derivatives that may be ingested and their pharmacological/toxic effects
Amine Sources Adverse effects
Histamine Fish, fermented sausages, cheese,
alcoholic drinks
Asthmatic attack, wheezing, decreased blood pressure,
urticaria, gastrointestinal symptoms, headache
Tyramine Cheese, pickled herring, chicken liver,
alcoholic drinks
Peripheral vasoconstriction, increases the cardiac output,
causes lacrimation and salivation, increases respiration,
increases blood sugar level, releases noradrenaline from
the sympathetic nervous system
2-phenylethylamine Chocolate, cheese, meat products Releases noradrenaline from the sympathetic nervous
system, increases the blood pressure, headache
Serotonin (5-hydroxytryptamine) Fruit (e.g. bananas) Inhibition of gastric secretion, stimulation of smooth
muscle, heart lesions (endomyocardial fibrosis, possibly)
Dopamine Bananas and other fruit, some vegetables Cardiotonic, antihypotensive (hypertensive crisis),
antioxidant and pro-oxidant
Tryptamine Cheese, meat and meat products,
fermented vegetables
Increases blood pressure
Methylamine Tobacco smoke (vapour phase), major
end-product of nicotine metabolism
Relatively non-toxic in the absence of SSAO
Allylamine Environment (used in the manufacture of
pharmaceuticals and vulcanized rubber)
Cardiovascular lesions
Volatile aliphatic amines Tobacco smoke (vapour phase)
Aromatic amines Tobacco smoke (particulate phase)
Nitrosamines Tobacco smoke (vapour and particulate
phase), salted, pickled and fermented
foods
Carcinogenic
Heterocyclic amines Cooked foods (cooking method) Carcinogenic
Spermine and spermidine Ubiquitous in food (e.g. fish, meat and
vegetable products)
Cell proliferation and differentiation
Cadaverine Fish, cheese, meat and meat products,
fermented vegetables
Hypotension, bradycardia, lockjaw, paresis of the
extremities, potentiates the toxicity of other amines
Putrescine Ubiquitous in food (e.g. fish) Cell proliferation and differentiation, hypotension,
bradycardia, lockjaw, paresis of the extremities,
potentiates the toxicity of other amines
Agmatine Fish, meat and vegetable products Modulation of pain, anticonvulsive effects
Adapted from Strolin Benedetti et al. (2007)
A. Olivieri et al.
123

are precursors for the synthesis of hormones, alkaloids,
nucleic acids and proteins, however, they can also pose
toxicological risks (KaroviCova and Kohajdova 2003;
Boobis et al. 2009). Because of microbiological decarbox-
ylation, biogenic amines are present in fermented products
(e.g. cheese 5–4500 mg kg-1, wine 5–130 mg dm-3, beer
2.8–13 mg dm-3, sauerkraut 110–300 mg kg-1) and in
improperly kept food (such as fish 2400–5000 mg kg-1,
beef liver about 340 mg kg-1, prepared meats 10–700
mg kg-1), (KaroviCova and Kohajdova 2003).
PrAO also catalyses the oxidative deamination of a
number of xenobiotics, including mescaline and the anti-
malarial drug primaquine (see O’Sullivan et al. 2004;
Tipton and Strolin Benedetti 2001). Metabolism of the
xenobiotic allylamine by PrAO produces acrolein, which
causes vascular toxicity (Hysmith and Boor 1988).
A common source of xenobiotic amines would be
through intake of drugs and additives present in food.
A number of antidiabetic drugs contain amino groups.
Metformin (dimethylbiguanide, see Fig. 2) is among the
most widely prescribed agents in the treatment of diabetes.
A related compound, aminoguanidine, still used as an
antidiabetic, has been reported to be a potent PrAO
inhibitor (Yu and Zuo 1997; Prevot et al. 2007). The anti-
inflammatory methylguanidine (Fig. 2) is an inhibitor of
diamine oxidase, DAO, another member of the SSAO
family of enzymes (Gang et al. 1976). Other guanidine
compounds include 1-phenethylbiguanide hydrochloride
(phenformin), which was used as antidiabetic, although its
use was discontinued for toxicity problems, 1-butylbigua-
nide (butformin), still used as antidiabetic, and the anti-
microbial pentamidine (see Fig. 2 for chemical structures
of phenformin, butformin and pentamidine). Phenformin
and pentamidine were also reported to inhibit DAO (Cubrıa
et al. 1991).
These and other dietary nitrogenous compounds, such
as caffeine, in tea, coffee, soft drinks and the now
popular ‘energy’ drinks, were assayed in this work for
their possible interaction with PrAO (see also Olivieri
and Tipton 2011). Some common dietary aldehydes
including vanillin, syringaldehyde, salicylaldehyde, cyn-
namaldehyde and anisaldehyde were also tested for any
potential interaction with the enzyme (see Figs. 1a, b and
2 for structures).
Given the multi-functionality of PrAO, especially in
relation to its roles in cell adhesion, maturation and glucose
uptake, it is important to know whether possible interactions
with biogenic and xenobiotic amines might occur after die-
tary intake or administration as drugs, since they might affect
the physiological roles of the enzyme. This work was
designed to test the inhibition of PrAO by a variety of such
compounds, as listed in Tables 2, 3 and 4, in order to gain
insights into possible interactions and to help identify lead
compounds for future drug developments.
Materials and methods
Sources of reagents
Bovine plasma PrAO was obtained by the supplier BioVar
Ltd, Yerevan, Armenia. Other enzymes and chemicals used
in this study were obtained from Sigma-Aldrich, unless
otherwise indicated.
Enzyme assay procedures
The activity of PrAO was determined following the pro-
duction of H2O2 at 498 nm, by the method of Holt and
Palcic (2006), in the presence of 5 mM benzylamine and
10 lg/ml PrAO. The chromogenic solution for the detec-
tion of H2O2 contained 1 mM vanillic acid, 0.5 mM
4-aminoantipyrine and horseradish peroxidase (4 U/ml) in
a ‘physiological’ HEPES buffer system (100 mM HEPES,
280 mM NaCl, 10 mM KCl, 4 mM CaCl2, 2.8 mM
MgCl2). The pH of the buffer was adjusted to 7.4 with
0.1 M NaOH. Assays were completed in a reaction volume
of 300 ll in 96-well microtitre plates, at 37�C, using a
SpectraMax 340PC plate reader (Molecular Devices, Inc.
Sunnyvale, CA 94089-1136, USA).
Control assays of the coupling system, in the pres-
ence of 10 lM H2O2, 1 mU/ml HRP but in the absence
of PrAO, showed that none of the compounds affected
the peroxidase chromogenic detection system. Each
compound was assayed in triplicate, at a final con-
centration of 1 mM and after a preincubation time of
0 min or 60 min, at 37�C, before adding substrate. If
any compound was found to show substantial PrAO
inhibition at 0 min and/or after 60 min of preincuba-
tion, it was further analyzed at different concentrations
and/or different times of preincubation to elucidate the
inhibition type.
Inhibition of bovine plasma PrAO in the presence
of protein hydrolysates from fish-waste sources
The samples shown in Table 5 were obtained from mack-
erel viscera, bones and gelatin after hydrolysis with dif-
ferent peptidases. All samples were diluted to a working
concentration of 1 mg/ml in assay buffer and added at 0, 60
and 120 min to the reaction mixture, for pre-incubation at
37�C, before adding substrate. Although the proteolysis
was completed at various pH values (see Table 5), control
experiments showed that when the compounds were diluted
From caffeine to fish waste
123
at 1 mg/ml in buffer the pH was 7.4 at their assay
concentrations.
Kinetics of Inhibition
IC50 values were determined by plotting the initial rates,
expressed in A498 nm 9 10-3 min-1 vs log [inhibitor] and
fitting the resulting plot to a sigmoidal curve, whose
point of inflection represents the log IC50 value for the
inhibitor studied. The initial rates (v = abs498 nm 9
10-3 min-1) of H2O2 formation were determined at 37�C
and pH 7.4. Data were then fitted to the Michaelis–
Menten equation by non-linear regression and the value
of Ki was determined from the dependence of the kinetic
parameters on the inhibitor concentration. The curves
were fitted with the aid of the computer software
GraphPad Prism, version 5.00.
Statistical analysis
Data collected were the mean values ± S.E.M. of at least
three independent experiments. Two-way ANOVA and
Bonferroni post tests were completed with the aid of the
computer software GraphPad Prism, version 5.00. Differ-
ences between two groups of data were considered
statistically significant when comparison tests resulted in
two-tailed P value \ 0.05.
Results
Effects of caffeine and polysaccharides
With the exception of caffeine, none of the compounds
shown in Table 3 showed significant inhibition of PrAO.
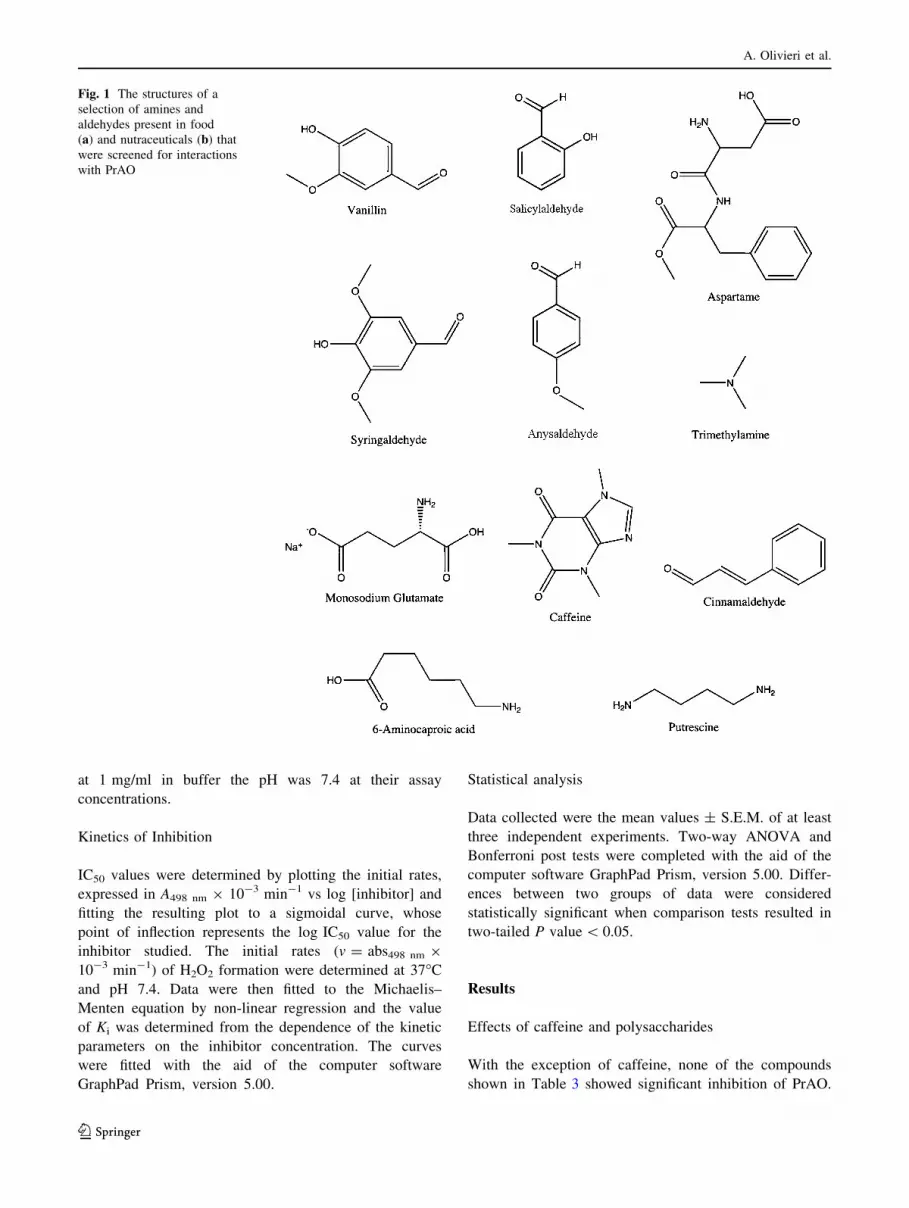
Fig. 1 The structures of a
selection of amines and
aldehydes present in food
(a) and nutraceuticals (b) that
were screened for interactions
with PrAO
A. Olivieri et al.
123

All were screened at concentrations of 1 and 10 mM,
except chondroitin sulphates A and B, which were tested in
the concentration range of 1–5 mg/ml and chitosan, where
a range of 0.1–10 mg/ml was used. It was not possible to
assess the effects of chitin, because of its insolubility.
Caffeine was an effective inhibitor with an IC50 of
0.8 ± 0.3 mM, as shown in Fig. 3. The inhibition, which
was not time dependent, was previously shown of a mixed-
type inhibitor with respect to methylamine, with a Ki and
Ki0 values of approximately 1.0 and 8.0 mM (Olivieri and
Tipton 2011).
Screening of common antidiabetic drugs as potential
inhibitors of PrAO
The compounds screened as potential PrAO inhibitors and
the concentration ranges used in the assays are listed in
Table 6. Phenformin (Fig. 4) was shown to inhibit PrAO
weakly (IC50 = 26.7 ± 1.1 mM), with no time depen-
dence observed when the enzyme was preincubated with
inhibitor for 1 h prior to the addition of substrate. Tolbu-
tamide was also a weak PrAO inhibitor, causing 20%
inhibition at a concentration of 1.0 mM, with no time
dependence. Butformin, metformin and methylguanidine
were not found to inhibit PrAO in the range 0–10 mM.
Phenytoin, aminotriazole and pentamidine and their
potential interactions with PrAO
The anti-epilectic drug phenytoin did not inhibit PrAO at
concentrations up 1 mM and preincubation for up to 1 h.
The weed killer aminotriazole was a weak inhibitor (ca.
20%) of the enzyme when used in the concentration range
0–1.0 mM and incubation times up to 1 h. No time
dependence was observed (see Table 7). However, the
antimicrobial drug pentamidine was found to inhibit PrAO
in the range 0.1–100 lM, with IC50 *3.5 lM with no
time-dependence observed (Fig. 5).
Inhibition of bovine plasma PrAO by protein
hydrolysates from fish-waste sources
14 samples containing protein hydrolysates from mackerel
bones, viscera and gelatin (see Table 5 for more informa-
tion on the samples) were assayed as potential PrAO
inhibitors, with significant findings that may be summa-
rized as follows (see Fig. 6a, b):
Samples 1 and 2 (from mackerel head and skin gelatin,
respectively, hydrolysed with pepsin) showed approxi-
mately 15% inhibition of PrAO at 0.25 mg/ml when
assayed without preincubation with the enzyme. This
inhibition increased to about 50% after incubation for 1 or
2 h prior to the addition of benzylamine.
Samples 7 and 8 (from mackerel viscera, hydrolysed by
endogenous proteases and with Aspergillus oryzae prote-
ase, respectively) behaved similarly to samples 1 and 2 at
time zero when present in the assayed without preincuba-
tion with the enzyme. Inhibition of PrAO was time
dependent, with approximately 30 and 50% inhibition after
incubation with the enzyme for 1 and 2 h, respectively,
before adding benzylamine.
Samples 11 and 12 (from mackerel viscera, hydrolysed
by trypsin and chymotrypsin, respectively) showed inhi-
bition of PrAO after 1 and 2 h of preincubation with the
enzyme prior to the addition of substrate. However, they
did not show any inhibition of PrAO at time zero.
None of the other hydrolysates tested had any significant
effects on PrAO activity.
Discussion
Determination of hydrogen peroxide formation indicated
that none of the compounds studied were substrates for
PrAO in the concentration ranges used. The ‘‘food addi-
tives’’ analysed were found to have no significant effects
on PrAO activity under the conditions used in this work.
Although substrate oxidation by PrAO yields an aldehyde
product and, therefore, product inhibition might be
Fig. 1 continued
From caffeine to fish waste
123

expected, the inhibition of the enzyme from pig plasma by
benzylaldehyde was reported to be competitive with
respect to benzylamine with a Ki of about 0.7 mM for the
enzyme from pig plasma (Taylor et al. 1972), none of the
aldehydes investigated had a significant effect on PrAO
activity. Neither the sweetener aspartame nor the flavour
enhancer monosodium glutamate inhibited PrAO. The
failure of 6-aminocaproic acid to inhibit was interesting,
since this can be regarded as an a-deaminated analogue of
L-lysine, which was reported to be an inhibitor of the
enzyme (Olivieri et al. 2007, 2010). The failure of chitosan,
which is widely used as an aid to slimming (see Jull et al.
2008) and the chondroitin sulphates to inhibit PrAO con-
trasts with the behaviour of free aminosugars, which have
been shown to inhibit the enzyme O’Sullivan et al. 2007).
Caffeine, which is probably among the most commonly
ingested drugs, since it is present in coffee, tea, numerous
soft drinks and ‘energy’ drinks has been shown to be a
PrAO inhibitor. Caffeine is a purine, containing an imid-
azole ring, thus it is possible that it might bind to an
inhibitory imidazoline binding site (I2) on PrAO (see Mu
et al. 1994; Holt et al. 2008; Olivieri and Tipton 2011). A
Fig. 2 The structures of a
selection of antidiabetic, anti-
epileptic and antimicrobial
drugs screened for interactions
with PrAO in this study
A. Olivieri et al.
123

‘‘typical’’ cup of coffee (5 oz) contains 40–170 mg of
caffeine (depending on how the coffee is brewed). Around
40 mg are contained in the average cup of tea (5 oz), a can
of Coke (12 oz) and in an espresso shot (1 oz). Some
‘energy’ drinks contain a higher quantity of caffeine,
varying from 80 to 300 mg. Caffeine is easily absorbed
into the bloodstream from the digestive tract and easily
moves out of the blood and into the tissues. The half-life
for caffeine in the bloodstream is about 3–4 h, while its
maximum concentration is achieved in 0.5–1 h after
ingestion (Klosterman 2006). Momoi et al. (2008) reported
that the ingestion of a regular cup of coffee corresponds to
a caffeine blood concentration of 5–10 lM, 30 min after
consumption. Battram et al. (2006) showed that acute
administration of 4.5 mg/kg (a regular cup of coffee would
provide about 1–2 mg/kg of caffeine) to 10 non-caffeine
users and 1 caffeine user of average body weight of 76 kg
resulted in a blood caffeine concentration that peaked at
36.8 ± 1.5 lM. These values remain well below the IC50
of 0.8–1.0 mM calculated in this work for the inhibition of
PrAO by caffeine. However, it cannot be excluded that
consumption of high concentrations of caffeine in a short
time might lead to significant physiological levels of PrAO
inhibition. Since PrAO is involved in regulating the uptake
of glucose (Enrique-Tarancon et al. 2000; El Hadri et al.
2002; Morin et al. 2002; Zorzano et al. 2003) it would be
interesting to investigate whether small, chronic levels of
PrAO inhibition among subjects who use caffeine in excess
might affect glucose uptake. Further work will be neces-
sary to show whether this inhibition of PrAO has any
physiological implications.
Several other amine compounds, commonly used as
antidiabetic drugs, were investigated in this work. 1,1-
Dimethylbiguanide HCl (metformin), one of the most
commonly used drugs in the treatment of diabetes and
2-butyl-1-(diaminomethylidene)-guanidine (butformin) did
not result in any significant PrAO inhibition. However,
phenformin (1-phenethylbiguanide�HCl) and tolbutamide
both acted as weak PrAO inhibitors. Such inhibition is
unlikely to be significant at the plasma levels reported after
administration of these antidiabetic drugs (see Marchetti
et al. 1987). Other compounds screened included the
antiepileptic drug diphenyldantoin (Phenytoin), which did
not inhibit PrAO and the weed-killer aminotriazole (con-
sisting of a pyrrolic ring and a primary amino group),
which acted as PrAO inhibitor, albeit a poor one.
The most potent inhibitor found among all the com-
pounds screened for this work was the antimicrobial
drug pentamidine. Pentamidine (1,5-bis(4-amidinophenoxy)
pentane) is a diamidine that has been reported to be a DAO
inhibitor and is used to treat pneumocystis carinii pneu-
monia (an infection common among immunosuppressed
patients; see Hughes et al. 1978). The fact that it is also an
inhibitor of PrAO from this source was surprising since not
many diamines display this property. Other polyamines,
such as spermidine, were reported to be able to bind to
PrAO at both the active site and a second binding site close
to the TPQ cofactor (Holt et al. 2008). It cannot be
excluded that pentamidine might bind to PrAO in a similar
fashion. Furthermore, the aromatic rings in proximity to the
amino groups of the molecule (see Fig. 2) might play a role
in the binding. The IC50 value for pentamidine was similar
to the maximum concentration of this drug reported in
human plasma after a standard 4 mg/kg dose administered
by intramuscular injection (Sands et al. 1985). Therefore,
Table 3 Some amines and aldehydes, contained in food and addi-
tives, that were screened as potential inhibitors of PrAO in this study
Aldehydes: vanillin,
syringaldehyde, cynnamaldehyde
Food flavourings
Aspartame Sweetener
Caffeine Coffee, tea, soft drinks
Monosodium glutamate Meat, food additive
Chitin, chitosan Vegetable, etc., products
6-Aminocaproic acid and
7-aminoheptanoic acid
From lysine metabolism
Trimethylamine oxide Fish
Chondroitin sulphate A and B Nutraceuticals, used for the
treatment of ostheoarthritis
Polyamines: putrescine,
spermidine, spermine and
cadaverine
Seafoods, etc.
Table 4 Amine compounds that were considered potential PrAO
inhibitors
Phenytoin (diphenyldantoin) Anti-epileptic
Aminotriazole Weed killer, carcinogen
Pentamidine Antimicrobial
Table 2 A selection of common antidiabetic drugs that were
screened as potential inhibitors of PrAO in this study
1,1-dimethylbiguanide HCl (metformin)—in common use
1-Phenethylbiguanide HCl (phenformin, phenethylbiguanide HCl)—
discontinued for toxicity problemsa
1-Butylbiguanide (butformin, 2-butyl-1-(diaminomethylidene)-
guanidine)—still commonly used
Aminoguanidine—reported to be antidiabetic and anti-ageingb
Methylguanidine—anti-inflammatory also formed in vivoc
Tolbutamide—one of the sulfonylurea group of drugs
a Phenformin is reported to inhibit diamine oxidase (Cubrıa et al.
1991)b Known to inhibit SSAO (Yu and Zuo 1997)c Inhibits DAO (Gang et al. 1976)
From caffeine to fish waste
123

a transient inhibition of PrAO might be expected after
administration of this drug.
The series of hydrolysates from mackerel gelatin (from
head and skin), viscera and frames were prepared using
different proteolytic enzymes under various conditions of
time and pH. Folador et al. (2006) showed that hydroly-
sates from the major fish wastes (heads, viscera, skin and
skeleton) contained approximately 5% lysine (of the total
amino acidic content), along with variable quantities of
biogenic amines, including histamine, tryptamine, putres-
cine, cadaverine, spermine, spermidine and tyramine.
Although these may make some contribution to the inhi-
bition observed, it was found that the nature of the pepti-
dase was critical for the formation of hydrolysates that
contained PrAO inhibitors. When gelatin from head and
skin was digested in pepsin, the hydrolysates produced
were found to inhibit PrAO in a time-dependent manner,
but this was not the case when the same hydrolysis was
carried out with trypsin or chymotrypsin. When the source
was mackerel viscera, the opposite occurred with trypsin
and chymotrypsin releasing material capable of inhibiting
PrAO after 1 and 2 h (s) of preincubation with the enzyme.
No PrAO inhibition was observed when the same hydrol-
ysates were digested in pepsin. The most effective prote-
olytic activities for mackerel viscera, in terms of generation
of PrAO inhibitors, were as a result of endogenous protease
activity (autolysis) and the protease preparation from
Table 5 Sources of protein
hydrolysates from fish-wasteSample
number
Source Enzyme used for
digestion
Hydrolysis
time (h)
pH Protein content
(mg/ml)
1 Head gelatine Pepsin 24 2.03 5.11
2 Skin gelatine Pepsin 24 2.02 4.96
3 Head gelatine Trypsin 24 7.59 5.04
4 Skin gelatine Trypsin 24 7.31 12.84
5 Head gelatine Chymotrypsin 24 7.7 3.68
6 Skin gelatine Chymotrypsin 24 7.59 11.48
7 Viscera Endogenous 24 6.97 2.36
8 Viscera Protease from
Aspergillus oryzae24 7.15 2.21
9 Viscera Protease from
Bacillus licheniformis24 7.2 6.82
10 Viscera Pepsin 24 2.95 7.17
11 Viscera Trypsin 24 7.2 3.01
12 Viscera Chymotrypsin 24 7.21 4.85
13 Frames Protease from
Aspergillus oryzae4 7.23 7.28
14 Frames Protease from
Bacillus licheniformis4 6.87 11.91
Fig. 3 The inhibition of PrAO by caffeine. The initial rates
(v = abs498 nm 9 10-3 min-1) of formation of H2O2 during the
oxidation of benzylamine by PrAO, were determined, at 37�C and pH
7.4, in the presence of the indicated concentrations of caffeine. The
curve fit and the value of IC50 = 0.8 ± 0.3 mM were obtained with
the aid of the computer software GraphPad Prism, version 5.00. The
x-axis is shown in logarithmic scale. All data were obtained from at
least three independent experiments, each completed in triplicate
Table 6 Screening of potential PrAO inhibitors among some com-
mon antidiabetic drugs
Compounds Concentration
range (mM)
Inhibition
1,1-dimethylbiguanide
HCl (Metformin)
0–10 None detected
1-Butylbiguanide
(butformin)
0–10 None detected
Methylguanidine 0–10 None detected
1-Phenethylbiguanide
HCl (Phenformin)
0–100 Yes
IC50 = 26.7 ± 1.1 mM
Tolbutamide 0–1 Yes
20 ± 1% at 1 mM
A. Olivieri et al.
123

Aspergillus oryzae. In contrast, the protease activity from a
Bacillus licheniformis preparation was ineffective. These
findings indicate that the effective inhibitors are peptides
generated by enzymes of appropriate specificities.
Although the specificities of chymotrypsin, pepsin and
trypsin are well known, the same cannot be said of the
peptidases from Aspergillus oryzae and Bacillus licheni-
formis, which appear to be complex mixtures of more than
one enzyme. Autolytic digestion is more complex in terms
Fig. 4 The inhibition of PrAO by 1-phenethylbiguanide�HCl (phen-
formin). The initial rates (v = abs498 nm 9 10-3 min-1) of formation
of H2O2, developed during the oxidation of benzylamine by PrAO,
were determined, at 37o C and pH 7.4, in the presence of the indicated
concentrations of phenformin. The curve fit and the value of
IC50 = 26.7 ± 1.1 mM were obtained with the aid of the computer
software GraphPad Prism, version 5.00. The x-axis is shown in
logarithmic scale. All data were obtained from at least three
independent experiments, each completed in triplicate. Error barsnot evident were smaller than the experimental points
Table 7 Screening of potential PrAO inhibitors among some drugs
and pesticides
Compounds Concentration
range
Inhibition
Phenytoin (diphenyldantoin) 0–1 mM None detected
Aminotriazole 0–1 mM Yes
20 ± 1% at 1 mM
Pentamidine 0–100 lM Yes
IC50 = 3.5 ± 0.50 lM
Fig. 5 The inhibition of PrAO by the antimicrobial pentamidine. The
initial rates (v = abs498 nm 9 10-3 min-1) of formation of H2O2,
formed during the oxidation of benzylamine by PrAO, were
determined, at 37�C and pH 7.4, in the presence of the indicated
concentrations of pentamidine. The curve fit and the value of
IC50 = 3.5 ± 0.50 lM were obtained with the aid of the computer
software GraphPad Prism, version 5.00. The x-axis is shown in
logarithmic scale. All data were obtained from at least three
independent experiments, each completed in triplicate. Error barswere smaller than the representation of the point
Fig. 6 Inhibition of bovine plasma PrAO in the presence of protein
hydrolysates from various sources. For information regarding the
nature of the samples refer to Table 7. All the samples were diluted to
a working concentration of 1 mg/ml in assay buffer. The final
concentration of the hydrolysates in the assay was 0.25 mg/ml. The
initial rates (v = abs498 nm 9 10-3 min-1) of formation of H2O2,
formed during the oxidation of benzylamine by PrAO, were
determined, at 37�C and pH 7.4. The data shown in the figure are
the mean values ± S.E.M. of three independent experiments, each
completed in triplicate. Statistical analysis (two-way ANOVA) was
completed with the aid of the computer software Prism, version 5.0.
Effect due to different samples P \ 0.0001, effect due to the time of
preincubation P \ 0.0001, interaction effect P \ 0.0001. The differ-ent columns were compared to their respective control (Ctrl) via
Bonferroni post test. *P \ 0.05, **P \ 0.001, ***P \ 0.0001
From caffeine to fish waste
123

of the enzymatic processes involved. The possibility that
amine formation, resulting from decarboxylation of any
free amino acids released during the process, might con-
tribute to the inhibition observed in these samples cannot
be excluded in this case. High levels of some amines can
also arise as a result of bacterial decarboxylation as fish
samples become stale. However, the observation that
digestion with some peptidases did not inhibit would sug-
gest that any amines generated in this way were not suf-
ficient to inhibit the enzyme.
Further analytical studies should be undertaken to
establish the nature of any inhibitory peptides present in
these hydrolysates as lead compounds for further devel-
opment as PrAO inhibitors.
Acknowledgments The authors wish to thank the Ashtown Food
Research Centre (AFRC), Dublin, Ireland for the provision of some of
the materials used in these studies and the Department of Agriculture,
Fisheries & Food (Ireland) for financial support towards part of this
work.
Conflict of interest The authors declare that they have no conflict
of interest.
References
Battram D, Arthur R, Weekes A, Graham T (2006) The glucose
intolerance induced by caffeinated coffee ingestion is less
pronounced than that due to alkaloid caffeine in men. J Nutr
136:1276–1280
Jull A, Ni Mhurchu C, Bennett, DA, Dunshea-Mooij, CA, Rodgers, A
(2008) Chitosan for overweight or obesity. Cochrane Database
Syst Rev CD003892
Boobis A, Watelet JB, Whomsley R, Strolin Benedetti M, Demoly P,
Tipton KF (2009) Drug interactions. Drug Metab Rev
41:486–527
Bour S, Prevot D, Guigne C, Stolen C, Jalkanen S, Valet P, Carpene C
(2007) Semicarbazide-sensitive amine oxidase substrates fail to
induce insulin-like effects in fat cells from AOC3 knockout
mice. J Neural Transm 114:829–833
Boyce S, Tipton KF, O’Sullivan MI, Davey GP, Motherway Gildea
M, McDonald AG, Olivieri A, O’Sullivan J (2009) Nomencla-
ture and potential functions of copper amine oxidases. In: Floris
G, Mondovi B (eds) Copper amine oxidases structures, catalytic
mechanisms and role in pathophysiology. CRC Press, Boca
Raton, FL, pp 5–17
Cubrıa J, Ordonez D, Alvarez-Bujidos M, Negro A, Ortız A (1991)
Inhibition of diamine oxidase from porcine kidney by pentam-
idine and other aminoguanidine compounds. Comp Biochem
Physiol B 100:543–546
Ekblom J, Gronvall JL, Garpenstrand H, Nillson S et al (2000) Is
semicarbazide-sensitive amine oxidase in blood plasma partly
derived from the skeleton? Neurobiology 8:129–135
El Hadri K, Moldes M, Mercier N, Andreani M, Pairault J, Feve B
(2002) Semicarbazide-sensitive amine oxidase in vascular
smooth muscle cells—differentiation-dependent expression and
role in glucose uptake. Arterioscler Thromb Vasc Biol 22:89–94
Enrique-Tarancon G, Castan I, Morin N, Marti L, Abella A, Camps
M, Casamitjana JR, Palacin M, Testar X, Degerman E, Carpene
C, Zorzano A (2000) Substrates of semicarbazide-sensitive
amine oxidase co-operate with vanadate to stimulate tyrosine
phosphorylation of insulin-receptor-substrate proteins, phospho-
inositide 3-kinase activity and GLUT4 translocation in adipose
cells. Biochem J 350:171–180
Folador JF, Karr-Lilienthal LK, Parsons CM, Bauer LL, Utterback
PL, Schasteen CS, Bechtel PJ, JR FaheyGC (2006) Fish meals,
fish components, and fish protein hydrolysates as potential
ingredients in pet foods. J Animal Sci 84:2752–2765
Gang V, Berneburg H, Henneman H, Hevendehl G (1976) Diamine
oxidase (histaminase) in chronic renal disease and its inhibition
in vitro by methylguanidine. Clin Nephrol 3:171–177
Holt A, Palcic MM (2006) A peroxidase-coupled continuous absor-
bance plate-reader assay for flavin monoamine oxidases, copper-
containing amine oxidases and related enzymes. Nat Protoc
1:2498–2505
Holt A, Smith D, Cendron L, Zanotti G, Rigo A, Di Paolo M (2008)
Multiple binding sites for substrates and modulators of semi-
carbazide-sensitive amine oxidases: kinetic consequences. Mol
Pharmacol 73:525–538
Hughes WT, Feldman S, Chaudhari SC, Ossi MJ, Cox F, Sanyal SK
(1978) Comparison of pentamidine isethionate and trimetho-
prim-sulfamethoxazole in the treatment of Pneumocystis cariniipneumonia. J Pediatr 92:285–291
Hysmith R, Boor P (1988) Role of benzylamine oxidase in the
cytotoxicity of allylamine toward aortic smooth muscle cells.
Toxicology 51:133–145
Jakobsson E, Nilsson J, Ogg D, Kleywegt GJ (2005) Structure of
human semicarbazide-sensitive amine oxidase/vascular adhesion
protein-1. Acta Crystallogr Sect D 61:1550–1562
Jalkanen S, Salmi M (2001) Cell surface monoamine oxidases:
enzymes in search of a function. EMBO J 20:3893–3901
Jalkanen S, Karikoski M, Mercier N, Koskinen K, Henttinen T, Elima
K, Salmivirta K, Salmi M (2007) The oxidase activity of
vascular adhesion protein-1 (VAP-1) induces endothelial E- and
P-selectins and leukocyte binding. Blood 110:1864–1870
KaroviCova J, Kohajdova Z (2003) Biogenic amines. Food Chem Pap
59:70–79
Klosterman L (2006) The facts about caffeine. Marshall, Cavendish
Lewinsohn R (1984) Mammalian monoamine-oxidizing enzymes,
with special reference to benzylamine oxidase in human tissues.
Braz J Med Biol Res 17:223–256
Marchetti P, Benzi L, Cecchetti P, Giannarelli R, Boni C, Ciociaro D,
Ciccarone AM, Di Cianni G, Zappella A, Navalesi R (1987)
Plasma biguanide levels are correlated with metabolic effects in
diabetic patients. Clin Pharmacol Ther 41:450–454
Momoi N, Tinney JP, Liu LJ, Elshershari H, Hoffman PJ, Ralphe JC,
Keller BB, Tobita K (2008) Modest maternal caffeine exposure
affects developing embryonic cardiovascular function and
growth. Am J Physiol Heart Circ Physiol 294:H2248–H2256
Morin N, Visentin V, Calise D, Marti L, Zorzano A, Testar X, Valet
P, Fischer Y, Carpene C (2002) Tyramine stimulates glucose
uptake in insulin-sensitive tissues in vitro and in vivo via its
oxidation by amine oxidases. J Pharmacol Exp Ther
303:1238–1247
Mu D, Medzihradszky K, Adams G, Mayer P, Hines WM, Burlin-
game A, Smith A, Cai D, Klinman J (1994) Primary structures
for a mammalian cellular and serum copper amine oxidase.
J Biol Chem 269:9926–9932
O’Sullivan J, Unzeta M, Healy J, O’Sullivan MI, Davey G, Tipton KF
(2004) Semicarbazide-sensitive amine oxidases: enzymes with
quite a lot to do. Neurotoxicology 25:303–315
O’Sullivan J, Davey G, O’Sullivan M, Tipton KF (2007) Hydrogen
peroxide derived from amine oxidation mediates the interaction
between aminosugars and semicarbazide-sensitive amine oxi-
dase. J Neural Transm 114:751–756
A. Olivieri et al.
123

Olivieri A, Tipton KF (2011) Inhibition of bovine plasma semicarb-
azide-sensitive amine oxidase by caffeine. J Biochem Mol
Toxicol 25:26–27
Olivieri A, Tipton K, O’Sullivan J (2007) L-Lysine as a recognition
molecule for the VAP-1 function of SSAO. J Neural Transm
114:747–749
Olivieri A, O’Sullivan J, Alvarez Fortuny LR, Larrauri Vives I,
Tipton KF (2010) Interaction of L-lysine and soluble elastin with
the semicarbazide-sensitive amine oxidase in the context of its
vascular-adhesion and tissue maturation functions. Biochim
Biophys Acta (BBA) 1804:941–947
Prevot D, Soltesz Z, Abello V, Wanecq E, Valet P, Unzeta M,
Carpene C (2007) Prolonged treatment with aminoguanidine
strongly inhibits adipocyte semicarbazide-sensitive amine oxi-
dase and slightly reduces fat deposition in obese Zucker rats.
Pharmacol Res 56:70–79
Sands M, Kron MA, Brown RB (1985) Pentamidine: a review. Rev
Infect Dis 7:625–634
Stolen CM, Yegutkin GG, Kurkijarvi R, Bono P, Alitalo K, Jalkanen
S (2004) Origins of serum semicarbazide-sensitive amine
oxidase. Circ Res 95:50–57
Stolen CM, Marttila-Ichihara F, Koskinen K, Yegutkin GG, Turja R,
Bono P, Skurnik M, Hanninen A, Jalkanen S, Salmi M (2005)
Absence of the endothelial oxidase AOC3 leads to abnormal
leukocyte traffic in vivo. Immunity 22:105–115
Strolin Benedetti M, Tipton KF, Whomsley R (2007) Amine oxidases
and monooxygenases in the in vivo metabolism of xenobiotic
amines in humans: has the involvement of amine oxidases been
neglected? Fundam Clin Pharmacol 21:467–480
Taylor CE, Taylor RS, Rasmussen C, Knowles PF (1972) A catalytic
mechanism for the enzyme benzylamine oxidase from pig
plasma. Biochem J 130:713–728
Tipton KF, Strolin Benedetti M (2001) Amine oxidases and the
metabolism of xenobiotics. In: Ioannides C (ed) Enzyme systems
that metabolise drugs and other xenobiotics. Wiley, Chichester,
UK, pp 95–146
Yu PH, Zuo DM (1997) Aminoguanidine inhibits semicarbazide-
sensitive amine oxidase activity: implications for advanced
glycation and diabetic complications. Diabetologia 40:1243–1250
Zorzano A, Abella A, Marti L, Carpene C, Palacin M, Testar X (2003)
Semicarbazide-sensitive amine oxidase activity exerts insulin-
like effects on glucose metabolism and insulin-signaling path-
ways in adipose cells. Biochim Biophys Acta 1647:3–9
From caffeine to fish waste
123





























