Embed Size (px)
Citation preview

Genetic dissection of quantitative powdery mildewresistance loci in tetraploid wheat
Roi Ben-David • Zvi Peleg • Amos Dinoor •
Yehoshua Saranga • Abraham B. Korol •
Tzion Fahima
Received: 16 July 2014 / Accepted: 7 October 2014 / Published online: 15 November 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Durum wheat, Triticum turgidum ssp.
durum Desf., is an important crop particularly in the
Mediterranean basin. Powdery mildew, caused by the
pathogen Blumeria graminis f. sp. tritici (Bgt), is a
major disease of wheat that results in significant yield
losses worldwide. A recombinant inbred line (RIL)
population, derived from a cross between durum
wheat and wild emmer wheat, T. turgidum ssp.
dicoccoides, was used for genomic dissection of
quantitative and qualitative resistance loci against
wheat powdery mildew based on a genomic map of
[600 markers, evenly distributed across the A and B
genomes of tetraploid wheat. The genetic analysis of
the phenotypic reactions of the RIL population to two
Bgt isolates revealed two different resistance mecha-
nisms. The first is monogenic: a wild emmer wheat
allele in a single locus conferring complete resistance
to Bgt#15, previously designated as PmG16. The
second one is polygenic: a set of durum wheat alleles,
in five independent QTLs that control partial resis-
tance to Bgt#66 in the RIL population, with a LOD
score range of 3.4–19.8. One of them is a major
quantitative resistance locus (QRL) that was mapped
on chromosome 1A and explains 26.4 % of the
variance. In most of the detected QRLs, the durum
wheat alleles conferred resistance to powdery mildew.Electronic supplementary material The online version ofthis article (doi:10.1007/s11032-014-0178-0) contains supple-mentary material, which is available to authorized users.
R. Ben-David
Institute of Plant Sciences, Agricultural Research
Organization (ARO)-Volcani Center, Bet Dagan 5025000,
Israel
e-mail: [email protected]
Z. Peleg � Y. Saranga
The Robert H. Smith Faculty of Agriculture, Food and
Environment, The Robert H. Smith Institute of Plant
Sciences and Genetics in Agriculture, The Hebrew
University of Jerusalem, Rehovot 7610001, Israel
A. Dinoor
Department of Plant Pathology and Microbiology,
The Robert H. Smith Faculty of Agriculture, Food and
Environment, The Hebrew University of Jerusalem,
Rehovot 7610001, Israel
A. B. Korol � T. Fahima (&)
Institute of Evolution and the Department of Evolutionary
and Environmental Biology, Faculty of Natural Sciences,
University of Haifa, 199 Abba-Hushi Avenue,
Mount Carmel, Haifa 3498838, Israel
e-mail: [email protected]
123
Mol Breeding (2014) 34:1647–1658
DOI 10.1007/s11032-014-0178-0

These findings are exceptional in the sense that, so far,
only a few Pm alleles originated from a durum wheat
background. Therefore, our results emphasize the high
potential of exploiting the wide genetic diversity of
tetraploid wheat germplasm for wheat breeding using
modern wheat genomics tools.
Keywords Blumeria graminis f. sp. tritici � Durum
wheat � Powdery mildew � QRL � Quantitative
resistance �Wild emmer wheat
Introduction
Wheat (Triticum spp.) is one of the world’s most
important food crop, which provides about one-fifth of
the calories and proteins consumed by humans
(FAOstat 2012, http://www.faostat.fao.org). Durum
wheat [T. turgidum ssp. durum (Desf.) MacKey.]
(2n = 4x = 28, genome BBAA) is the second most
widespread Triticum species constituting 10–11 % of
the world’s wheat cultivation land and accounts for
about 5 % of total wheat production. The Mediterra-
nean basin, close to the initial wheat domestication
and cultivation region, is producing most (*75 %) of
the world’s durum grains, but durum wheat is an
important crop also in other wheat-growing regions
(Elias and Manthey 2005). Durum wheat is the main
source for production of pasta, couscous, burghul and
other Mediterranean local end-products (Nachit et al.
2001). The ever-increasing human population con-
comitantly with loss of agricultural land (due to
urbanization processes, industrialization, desertifica-
tion and climatic changes) and diminishing resource
availability pose serious challenges to world agricul-
ture. To feed the 9 billion people expected by 2050
(http://www.fao.org/wsfs/world-summit/en/), a sig-
nificant grain yield increase of approximately 44
million metric tons per year would be needed (Tester
and Langridge 2010). It is estimated that pathogens
can cause potential yield loss of 16–23 % with greater
losses in intensive production agri-systems (Oerke
2006). Developing new cultivars with improved dis-
ease resistance is an economically and environmen-
tally safe approach to reduce yield losses. This
solution, however, requires a comprehensive explo-
ration of potential genetic resources and an in-depth
understanding of their resistance mechanisms.
Powdery mildew, caused by the biotrophic patho-
gen Blumeria graminis (DC.) E.O. Speer f. sp. tritici
Em. Marchal (Bgt hereafter), is a foliar wheat disease
resulting in severe yield losses worldwide. B. graminis
is an obligate biotrophic fungus of the order Erysip-
hales (Ascomycota), family Erysiphaceae (Braun et al.
2000). The continuous threat for breakdown of race-
specific resistance to powdery mildew (e.g., Bennett
1984; Hsam and Zeller 2002) is forcing a consecutive
effort to uncover new genes and alleles from the
resistance gene reservoir. Studies conducted in the last
75 years to identify powdery mildew resistance in
durum wheat germplasm yielded only a few resources
(reviewed by Bennett 1984). A single dominant gene,
designated Mld, was identified in durum wheat cv
Yuma (Briggle 1960). This powdery mildew resis-
tance gene was combined with Pm2 to produce
resistance in a British spring wheat cultivar. This
resistance has broken down due to a matching
virulence gene in mildew populations (Bennett
1984). A phenotypic test for resistance to powdery
mildew in 92 wheat lines resulted in only one durum
line possessing resistance (Upadhyay et al. 1972).
Screening of 2,500 exotic durum wheat and 500 exotic
bread wheat lines facilitated the detection of 35
additional resistant durum lines (Bahadur et al. 1977,
1979(. The allele Pm3h and the gene PmDR147 were
both derived from durum wheat (Hsam and Zeller
2002; Zhu et al. 2004).
Several genetic maps of crosses between durum
wheat and wild emmer have been constructed (e.g.,
Peng et al. 2000; Peleg et al. 2008; Ji et al. 2008) and
used for identification and allocation of various
agronomically important traits in tetraploid wheat,
including powdery mildew resistance genes (e.g.,
Ben-David et al. 2010; Ji et al. 2008). Qualitative
disease resistance is under simple genetic control and
is well characterized. By contrast, other forms of
disease resistance controlled by genetically complex
systems are less understood (Tanksley 1993). Most
complex resistance traits are controlled by multiple
loci, and their phenotypes are measured quantitatively.
Young (1996) defined the genetic loci associated with
this kind of resistance as quantitative resistance loci
(QRLs). This term is frequently used synonymously
with the term ‘‘partial resistance loci.’’ The number of
QRLs tends to range from one to ten or more in any
one system, suggesting that the number of loci
involved in quantitative disease resistance (QDR)
1648 Mol Breeding (2014) 34:1647–1658
123

might generally be lower than for other agriculturally
important traits (e.g., growth rate, biomass production,
fertility) (Young 1996). The identification of QRLs
plays a major role in crop improvement implemented
through marker-assisted selection (Miedaner and
Korzun 2012) and provides the genetic basis for
positional cloning of resistance genes.
A recombinant inbred line (RIL) population
derived from a cross between durum wheat and its
direct progenitor, wild emmer wheat [T. turgidum ssp.
dicoccoides (Korn.) Thell.], was used to genetically
dissect grain yield and yield components (Peleg et al.
2009a, b, 2011) as well as mapping of a single
dominant powdery mildew resistance gene, desig-
nated as PmG16 (Ben-David et al. 2010). In the
current study, we used the same population to: (1)
characterize the quantitative response to a Bgt isolate,
which is virulent on PmG16; (2) determine the
chromosomal location and phenotypic effects of QRLs
associated with wheat response to powdery mildew;
and (3) assess the novelty of the new QRLs compared
to known powdery mildew QRLs.
Materials and methods
Plant and fungal materials
A population of 152 F7 RILs was developed using the
single-seed descent (SSD) approach, from a cross
between durum wheat (cv. Langdon, LDN hereafter)
and wild emmer wheat (accession G18-16) (Peleg
et al. 2008). This RIL population was subjected to a
quantitative phenotypic analysis after inoculation with
Bgt isolate #66, collected from a natural population of
wild emmer wheat in Ammiad, northern Israel
(32�560N, 35�320E; 290 m above sea level). This
isolate is part of the ‘‘Eshed-Dinoor mildew collec-
tion,’’ maintained at the Hebrew University, and is
virulent on PmG16 that segregates in the same RIL
population (Ben-David et al. 2010).
Phenotypic characterization
A qualitative phenotypic test was performed at the
seedling stage on detached leaf segments, as previ-
ously described in Ben-David et al. (2010). The
quantitative response to Bgt#66 was characterized by
counting the number of powdery mildew pustules per
cm2 at the seedling stage on detached leaf segments at
three different days post-inoculation (DPI) points.
Leaf segments of the tested lines were maintained in
polystyrene boxes with water agar (6 g agar L-1)
supplemented with 50 mg L-1 of benzimidazole. The
detached leaves were infected with powdery mildew
spores in a settling tower at inoculum density of *500
spores cm-2. The leaf segments were incubated at
15 �C with a 12-h photoperiod of white fluorescent
light (photosynthetic photon flux density of
55–65 lmol m-2 s-1). The density of the deposited
spores per cm2 and the proportion and density of
germinated spores were determined in each of the
polystyrene boxes (six random measurements per box)
24 h after inoculation using a light microscope.
Optimization of the quantitative phenotypic assay
was performed, and it is detailed in the supplementary
section (see Fig. 1S and additional text). Inoculation
of the complete RILs population was performed on
two independent dates (90 days apart). These two
dates serve as two separate environments in the
ANOVA and QRL analysis.
The two parental lines (LDN and G18-16) were also
characterized for reaction to Bgt#66 at the adult plant
stage (Zadoks index 57; Zadoks et al. 1974). The
parental lines were grown in 5L pots at a mildew-free
greenhouse and were inoculated with fresh spores of
Bgt#66. In order to avoid the risk of cross-contami-
nation, transparent plastic cylinders were used to cover
plants prior to inoculation with Bgt spores. The pots
were then transferred to a controlled growth chamber
(15 �C) and were kept closed in those plastic
cylinders.
Statistical analysis
The JMP� ver. 10.0 statistical packages (SAS Insti-
tute, Cary, NC) were used for all statistical analyses.
The mean density of pustules (average of three leaf
segments) for each time point was transformed to
H(X ? 1) before analysis of variance. A factorial
model was employed for the analysis of variance
(ANOVA), with RILs and environment as random
effects. To assess the effect of RIL and environment
on the disease severity, ANOVA was implemented at
both 10DPI and 14DPI. Broad sense heritability
estimates (h2) were calculated for each trait across
the two environments using ANOVA-based variance
components:
Mol Breeding (2014) 34:1647–1658 1649
123

h2 ¼ r2G=ðr2
G þ r2G�E=EÞ
where r2G ¼ MSRIL � MSRIL�Eð Þ=E½ �; r2
G�E ¼MSRIL�E and E is the number of environments and
MS is the mean square.
QRL analysis
A genetic linkage map of 2,317 cM was previously
developed by genotyping the 152 RIL mapping
population with 197 simple sequence repeat (SSR)
and 493 Diversity Array Technology (DArT) markers
(Peleg et al. 2008). DArT markers presented in the
above map by clone ID numbers were renamed with
the prefix ‘‘wPt,’’ ‘‘rPt’’ or ‘‘tPt’’ (corresponding to
wheat, rye or triticale, respectively) followed by a
number. A skeleton map comprised of 307 markers,
scattered along the 14 chromosomes (chr.) of the
tetraploid wheat genome (an average of one marker
per 7.5 cM), was used for QRL analysis. The QRL
analysis was performed with the MultiQTL package
(http://www.multiqtl.com) using the general interval
mapping analysis for the RIL-selfing population as
described by Peleg et al. (2009a, b). QRL detection
was carried out with a structured multi-step scheme
embedded within the software (Korol et al. 2001).
First, the entire genome was screened for genetic
linkage using single-trait analysis (STA). Next, mul-
tiple interval mapping (MIM) was applied, which
incorporates interfering effects of other QRLs located
on a separate chromosome(s) into the model to reduce
the residual variation (Kao et al. 1999). MIM was
applied when more than one QRL was detected. The
hypotheses that a single locus or two-linked loci on the
considered chromosome could have an effect on one
or two quantitative traits were first tested by running
5,000 permutation tests (Churchill and Doerge 1994).
The hypothesis that one locus on the chromosome has
an effect on a given trait (H1) was compared with the
null hypothesis (H0) of no effect of the chromosome
on that trait. Once the genetic model was chosen,
5,000 bootstrap samples were run to estimate the
standard deviation of the main parameters: locus
effect, its chromosomal position, its LOD score, and
the proportion of explained variation (PEV). Finally,
to evaluate the genome-wide significance of estimates
obtained on a chromosome 9 trait basis, an approach
based on controlling the false discovery rate was used
to correct for multiple comparisons (Benjamini and
Hochberg 1995). The effect of an epistatic interaction
was examined by comparison of H0(e = 0), i.e.,
additive effects of the QRL, and H1(e = 0), i.e.,
assuming epistasis (Ronin et al. 1999).
Environmental specificity of QRL was determined
using two approaches. First, the two-environment
QRL model was compared against a sub-model
assuming equal effect of both environments, using
5,000 permutation tests to examine (Geno-
type 9 environment) interaction. To determine envi-
ronmental specificity, we used additional criteria
where the QRL must not only show significant
G 9 E interaction but must also show [0.05 differ-
ence between PEVs of the two environments. The
environment with the greater PEV value was declared
as the most influential. Second, when QRL was found
significant only for one environment but not for the
two-environment model, it was declared as an envi-
ronment-specific QRL.
Results
Screening of the parental lines with 42 Bgt isolates
This study is based on a cross between durum wheat
and wild emmer wheat. These two parental lines were
first tested for their response to a set of 42 Bgt isolates,
from the ‘‘Eshed-Dinoor mildew culture collection’’
collected from domesticated and wild wheat species
from different locations in Israel, and five isolates
from Switzerland collected from cultivated wheat
fields (Table S1, Ben-David et al. 2010). The wild
emmer wheat parent showed a resistance response to
29 of the Bgt isolates, while the durum wheat LDN
showed a resistance response to only a few isolates
(Ben-David et al. 2010). The wild parent (G18-16)
was highly resistant to the powdery mildew isolate
Bgt#15, whereas LDN was highly susceptible to this
isolate (IT scores 0 and 4, respectively). Therefore,
isolate Bgt#15 was previously used to map the
resistance loci harbored by the wild emmer wheat
parent, as a single dominant gene, designated PmG16
(Ben-David et al. 2010). Contrary to Bgt#15, Bgt#66 is
virulent on PmG16 and generated a unique quantita-
tive resistance reaction when tested on the parental
lines of the RIL population (Fig. 2S). The durum
wheat parent (LDN) showed slight powdery mildew
symptoms (IT = 1, designated R-), while the wild
1650 Mol Breeding (2014) 34:1647–1658
123

emmer parent (G18-16) showed a moderately suscep-
tible reaction (IT = 2–3, designated M). This reaction
of both parents was evident both at the seedling stage
(Zadoks index 12), as well as at the adult plant stage
(Zadoks index 57; Zadoks et al. 1974) (Fig. 1).
Quantitative phenotypic characterization
of inoculation with Bgt#66
Each Bgt isolate possesses specific phenotypic fea-
tures, such as mycelium growth rate, pustule produc-
tion rate and sporulation pattern, and therefore, one
needs to optimize the phenotypic measurements for
each isolate, prior to conducting large-scale experi-
ments. Based on the counting of pustule densities at
five time points, on five susceptible tetraploid lines
(Fig. 1S), we concluded that the most critical param-
eter in avoiding experimental error is the time of
measuring pustule densities after inoculation (DPI).
Early measurement of pustule densities could result in
false positives due to failing to distinguish between
established colonies and unsuccessful penetration sites
(manifested by hyper- sensitive-like responses), while
late counting may result in false negatives due to the
merging of mycelia of neighboring colonies. There-
fore, based on our preliminary assay, two time points,
10 and 14 DPI, were selected for further measurements
in order to optimize the accuracy of measurements of
pustule densities.
Table 1 presents the quantitative response, as
means of numbers of pustules per cm2 of leaf area,
for the two parental lines and the RIL population in
two environments at 10 and 14 DPI. G18-16 was
susceptible to Bgt isolate #66 with a mean range of
45.2–74.6 pustules per cm2 (Table 1), whereas LDN
was partially resistant with a mean range of 15.0–17.3
pustules per cm2 (Table 1). The RILs exhibited
transgressive segregation for quantitative resistance
to powdery mildew (range of 0–127.4 colonies per
cm2). The pustule densities measured in environment
II were prominently higher when compared to envi-
ronment I, while the two measurement points (10 and
14 DPI) within each environment were relatively
homogeneous. The susceptible wild emmer wheat
genotype 1–67 showed an average of 74.2 pustules per
cm2 across all replicates and was therefore used as a
G18-16LDN
a
b
c
d
Fig. 1 Phenotypic responses of the two parental lines, Langdon
(LDN) and G18-16, to inoculation with isolate Bgt#66. Leaf
segment and whole plant assays are presented: a, b phenotypic
response of LDN at adult plant stage and seedling stage,
respectively; c, d phenotypic response of G18-16 at adult plant
stage and seedling stage, respectively
Table 1 Mean values and data range of pustule densities at 10 and 14 days post-inoculation (DPI) of 152 F7 recombinant inbred
lines (LDN 9 G18-16) and the two parental lines under two environmental conditions (I and II)
DPI Environment I (pustules/cm2) Environment II (pustules/cm2)
RILs LDN G18-16 RILs LDN G18-16
Mean Range Mean Range
10 20 0–103.0 15 45.2 46.3 0.9–127.4 16.6 68.1
14 21.7 0–100.7 17.3 57.3 38.3 2.0–115.0 15.6 74.6
Mol Breeding (2014) 34:1647–1658 1651
123

positive control line in all experiments to monitor
infection conditions between inoculated boxes. The
average deposited spore density and the germination
rate values (number of spores with germ tubes/total
number of spores) for each box were measured in
order to ensure uniform inoculation density across the
assay. Analysis of variance of pustule densities carried
out on the segregating RIL population revealed highly
significant effects (P \ 0.001) of genotype and envi-
ronment at 10 and 14 DPI, significant interaction and
very high heritability (Table 2).
Characteristics of the detected QRLs
A whole-genome genetic linkage map of 2,317 cM
with an average of one marker per 7.5 cM was used for
dissection of the mechanism controlling the wheat
plant response to Bgt#66. A total of five significant
QRLs, scattered across 5 out of 14 chromosomes of the
tetraploid wheat, was detected under two environ-
ments at 10 and 14 DPI (Table 3; Fig. 2). In all QRLs
but one, the durum wheat allele (LDN) contributed to
improved powdery mildew resistance (lower density
of fungal colonies on the leaves). Only in one QRL
(5B), the favorable allele was contributed by the wild
parent (G18-16). Only the QRL on 7A exhibited a
significant G 9 E interaction. No significant two-
locus epistasis was detected between any pair of the
QRLs. Since the measurements of colony densities at
10 DPI provided, in most cases, a higher resolution, we
used the second measurement (14 DPI) as another
confirmation to the QRLs identified at 10 DPI.
A total of five significant QRLs conferring powdery
mildew resistance were identified at 10 DPI, with
LOD scores ranging between 3.4 and 16.3, and
explaining 1.3-26.6 % of the variance (Table 3).
Higher resistance was conferred by the LDN alleles
at four loci (1A, 2B, 6B and 7A) and by the G18-16
allele at one locus (5B). One QRL showed significant
G 9 E interaction (7A). Two QRLs (5B and 6B) were
detected only under environment II (Table 3). Three
significant QRLs conferring powdery mildew resis-
tance were detected at 14 DPI, with LOD scores
ranging between 3.9 and 19.8, explaining 0.3–26.4 %
of the variance (Table 3). These QRLs, detected at 14
DPI, provided further validation for the three major
QRLs detected at 10 DPI. The five significant QRLs,
detected at least in one of the two environments, at 10
or 14 DPI were designated as QPm#66-1A, QPm#-
662B, QPm#66-5B, QPm#-666B and QPm#-667A
(Fig. 2).
Discussion
Crosses between durum wheat cultivars are commonly
used in durum wheat genetic studies. However, the
narrow genetic variation possessed by these crosses
limits the ability to detect and enrich the durum
resistance gene reservoir with novel alleles. To
overcome this limitation, we have used a whole
genome analysis of a tetraploid wheat cross between
durum wheat and wild emmer wheat. Using this
approach, we were able to detect novel QRLs due to
the high variability of wild and domesticated wheat
germplasm. Moreover, the recent advances in the
development and use of high-density genetic and
genomic maps (Maccaferri et al. 2014a, b e.g., SNP
based) will greatly enhance the genetic detection
power and the chances to genetically dissect novel
plant resistance alleles and QRLs (e.g., Marone et al.
2013).
Based on multiple observations of responses to
different types of pathogens in various crop-plants,
quantitative disease resistance was suggested as more
durable than typical R-gene-mediated resistance
(reviewed by Parlevliet 2002; Marone et al. 2013).
The polygenic nature of QDR also affects the selection
pressure on the pathogen. Even if the pathogen
develops a mutation that enables it to overcome a
single QRL, the pathogen can gain only a marginal
Table 2 Analyses of variance for the effects of genotype and
environment on pustules’ density at 10 and 14 days post-
inoculation (DPI) in a segregating F7 RIL population
(LDN 9 G18-16)
Source of variation d.f. Mean square
10 DPI 14 DPI
Genotype (G) 136 2,652*** 1,959***
Environment (E) 1 135,825*** 55,784***
G 9 E 136 920*** 683***
Experimental error 548 393 301
Heritability (h2) 0.97 0.97
Heritability estimates (h2) of the RIL population are presented
for all traits
*** Significance at P \ 0.001
1652 Mol Breeding (2014) 34:1647–1658
123

Ta
ble
3B
iom
etri
cal
par
amet
ers
of
QR
Ls
affe
ctin
gp
ow
der
ym
ild
ewre
sist
ance
inte
trap
loid
wh
eat
RIL
po
pu
lati
on
(LD
N9
G1
8-1
6)
Tra
it/c
hro
mo
som
eP
osi
tio
n(c
M)
Nea
rest
mar
ker
LO
Da
En
vir
on
men
tI
En
vir
on
men
tII
Fav
ora
ble
alle
led
G9
Ee
P.E
.V.b
dc
P.E
.V.
d
10
da
ysp
ost
-in
ocu
lati
on
1A
30
.47
±5
.29
Xg
wm
30
83
16
.3*
**
0.2
66
3.0
66
±0
.46
30
.17
63
.74
8±
0.6
97
Ln
.s.
2B
16
8.4
2±
7.9
9X
wP
t-2
72
47
.3*
**
0.0
78
1.6
08
±0
.51
40
.07
72
.45
5±
0.1
86
Ln
.s.
5B
89
.67
±1
7.5
0X
wP
t-6
02
23
.9*
*–
–0
.05
7-
1.8
72
±0
.42
7G
6B
45
.74
±1
9.2
1X
wP
t-0
55
43
.4*
––
0.0
45
1.6
44
±0
.39
9L
7A
42
.80
±1
4.3
5X
tPt-
17
55
4.9
**
0.0
13
0.2
41
±0
.55
50
.11
52
.95
6±
0.9
53
L*
**
14
da
ysp
ost
-in
ocu
lati
on
1A
31
.52
±4
.49
Xg
wm
30
83
19
.8*
**
0.2
64
2.5
74
±0
.17
40
.23
02
.81
5±
0.4
52
Ln
.s.
2B
17
0.6
0±
8.9
2w
Pt-
21
35
7.3
**
*0
.05
71
.12
7±
0.3
68
0.1
03
1.8
48
±0
.39
3L
n.s
.
7A
50
.13
±2
1.1
1X
gw
m8
71
a3
.9*
0.0
45
-0
.54
5±
0.6
48
0.0
30
1.3
29
±0
.74
6L
*
aL
OD
sco
res
that
wer
efo
un
dsi
gn
ifica
nt
wh
enco
mp
arin
gh
yp
oth
eses
H1
(th
ere
isa
QT
Lin
the
chro
mo
som
e)v
ersu
sH
0(n
oef
fect
of
the
chro
mo
som
eo
nth
etr
ait)
,u
sin
g5
,00
0
per
mu
tati
on
ste
st(C
hu
rch
ill
and
Do
erg
e1
99
4)
bP
rop
ort
ion
of
exp
lain
edv
aria
tio
no
fth
etr
ait
cT
he
adap
tiv
eef
fect
of
anal
lele
mea
sure
das
on
e-h
alf
of
the
mea
nd
iffe
ren
cein
ho
mo
zyg
ote
sw
ith
and
wit
ho
ut
the
alle
led
Fav
ora
ble
par
enta
lal
lele
con
trib
uti
ng
tob
ette
rp
ow
der
ym
ild
ewre
sist
ance
(lo
wer
pu
stu
led
ensi
ties
),L
ang
do
n(L
)an
dG
18
-16
(G)
eG
eno
typ
e9
env
iro
nm
ent
inte
ract
ion
,te
sted
by
com
par
ing
the
mo
del
wit
hn
ewsu
b-m
od
elin
wh
ich
bo
then
vir
on
men
tsh
ave
equ
alef
fect
,u
sin
g5
,00
0p
erm
uta
tio
ns
test
.T
his
test
isn
ot
app
lica
ble
wh
enQ
TL
issp
ecifi
cfo
ro
nly
on
een
vir
on
men
to
rin
case
of
the
two
-QT
Lm
od
el
*,
**
,*
**
and
n.s
.in
dic
ate
sig
nifi
can
ceat
PB
0.0
5,
0.0
1,
0.0
01
or
no
n-s
ign
ifica
nt
effe
ct,
resp
ecti
vel
y
Mol Breeding (2014) 34:1647–1658 1653
123

advantage and therefore will have only a marginal
fitness advantage over other pathogen strains (Poland
et al. 2008). In the current study, we have used QRL
analysis to study plant–pathogen interactions and
discuss the results in light of previous QDR studies.
Bgt#66 was virulent on PmG16, but was avirulent
(IT = 0) on a set of 17 differential wheat lines that
carry Pm alleles (i.e., Pm1a, Pm1b, Pm2, Pm3a,
Pm3b, Pm3c, Pm3d, Pm4a, Pm4b, Pm5a, Pm5b, Pm6,
Pm7, Pm8, Pm9 and Pm17) (Ben-David 2011).
Therefore, isolate Bgt#66 allowed us to expose partial
resistance QRL alleles derived from the durum wheat
parental line. Five significant QRLs associated with
the phenotypic reactions to inoculation of tetraploid
wheat with Bgt#66 were identified and mapped.
Interestingly, most QRLs affecting improved resis-
tance to the Bgt#66 were conferred by the domesti-
cated durum wheat (LDN) alleles. These findings are
unique in the sense that, so far, only two Pm genes are
known to have originated from a durum wheat
background (Hsam and Zeller 2002; Huang et al.
2004; Zhu et al. 2004). A major QRL was detected on
chr. 1A (QPm#66-1A, LOD 16.3 and 19.8 for 10 and
14 DPI, respectively) explaining 26.6 and 26.4 % of
the variance in the two environments, at 10 and 14 DPI
(Table 3; Fig. 3). The implementation of the two
Fig. 2 Likelihood intervals for quantitative resistance loci
(QRLs) associated with resistance to powdery mildew isolate
Bgt#66 in recombinant inbred line (RIL) population of the cross
between Langdon and G18-16. Molecular markers are shown on
the left with map distances and QRLs on the right. The
approximate centromere position is indicated by black rectangle
1654 Mol Breeding (2014) 34:1647–1658
123
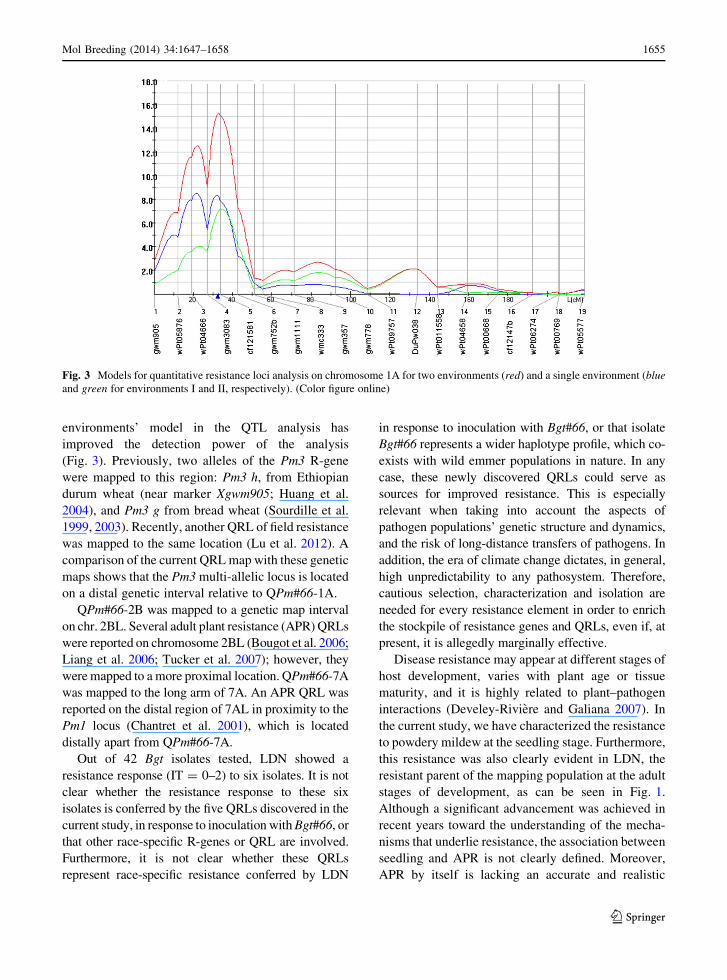
environments’ model in the QTL analysis has
improved the detection power of the analysis
(Fig. 3). Previously, two alleles of the Pm3 R-gene
were mapped to this region: Pm3 h, from Ethiopian
durum wheat (near marker Xgwm905; Huang et al.
2004), and Pm3 g from bread wheat (Sourdille et al.
1999, 2003). Recently, another QRL of field resistance
was mapped to the same location (Lu et al. 2012). A
comparison of the current QRL map with these genetic
maps shows that the Pm3 multi-allelic locus is located
on a distal genetic interval relative to QPm#66-1A.
QPm#66-2B was mapped to a genetic map interval
on chr. 2BL. Several adult plant resistance (APR) QRLs
were reported on chromosome 2BL (Bougot et al. 2006;
Liang et al. 2006; Tucker et al. 2007); however, they
were mapped to a more proximal location. QPm#66-7A
was mapped to the long arm of 7A. An APR QRL was
reported on the distal region of 7AL in proximity to the
Pm1 locus (Chantret et al. 2001), which is located
distally apart from QPm#66-7A.
Out of 42 Bgt isolates tested, LDN showed a
resistance response (IT = 0–2) to six isolates. It is not
clear whether the resistance response to these six
isolates is conferred by the five QRLs discovered in the
current study, in response to inoculation with Bgt#66, or
that other race-specific R-genes or QRL are involved.
Furthermore, it is not clear whether these QRLs
represent race-specific resistance conferred by LDN
in response to inoculation with Bgt#66, or that isolate
Bgt#66 represents a wider haplotype profile, which co-
exists with wild emmer populations in nature. In any
case, these newly discovered QRLs could serve as
sources for improved resistance. This is especially
relevant when taking into account the aspects of
pathogen populations’ genetic structure and dynamics,
and the risk of long-distance transfers of pathogens. In
addition, the era of climate change dictates, in general,
high unpredictability to any pathosystem. Therefore,
cautious selection, characterization and isolation are
needed for every resistance element in order to enrich
the stockpile of resistance genes and QRLs, even if, at
present, it is allegedly marginally effective.
Disease resistance may appear at different stages of
host development, varies with plant age or tissue
maturity, and it is highly related to plant–pathogen
interactions (Develey-Riviere and Galiana 2007). In
the current study, we have characterized the resistance
to powdery mildew at the seedling stage. Furthermore,
this resistance was also clearly evident in LDN, the
resistant parent of the mapping population at the adult
stages of development, as can be seen in Fig. 1.
Although a significant advancement was achieved in
recent years toward the understanding of the mecha-
nisms that underlie resistance, the association between
seedling and APR is not clearly defined. Moreover,
APR by itself is lacking an accurate and realistic
Fig. 3 Models for quantitative resistance loci analysis on chromosome 1A for two environments (red) and a single environment (blue
and green for environments I and II, respectively). (Color figure online)
Mol Breeding (2014) 34:1647–1658 1655
123

definition. Quantitative APR gene is considered to be
expressed when major APR resistance genes are
absent from the genetic background or when only a
defeated forms of the R-gene is evident (Wang et al.
2005). While it can be assumed that APR genes are
more durable, cases of race-specific APR loci were
also reported and therefore should be taken into
consideration when used by breeders (Li et al. 2014).
It has been suggested that some QRLs or APR
condition a weaker form of R-gene-mediated defense
although the possibility that other mechanisms could
provide isolate-specific resistance was not ruled out
(Parlevliet and Zadoks 1977). In light of this hypoth-
esis, phenotypic variance and durability can be
explained by a minor-gene-for-minor-gene interac-
tion, where virulence genes having a minor effect in
the pathogen correspond to resistance genes having a
minor effect in the host. Poland et al. (2008) assumed
that qualitative and quantitative disease resistance
might only be two ends of a continuum, with R-genes
and QRLs resembling the two extremes of the
spectrum. Although selection favors R-genes with
strong effects, pathogen evolution can erode the
effectiveness of R-genes, converting them into QRLs.
This phenomenon, known as ‘‘residual resistance,’’
has been observed in wheat powdery mildew (Nass
et al. 1981) and wheat stem rust (Brodny et al. 1986):
When a pathogen strain overcomes an R-gene, the
level of disease in the presence of the ‘‘defeated’’
R-gene is reduced relative to the level of disease in the
absence of the R allele. QTL analysis can help test the
hypothesis that QRLs are variants of R-genes that have
been (partially) overcome by their respective patho-
gens. Combining DNA markers and QTL mapping,
complex forms of disease resistance and their under-
lying genes are now far more accessible for investi-
gation (e.g., Marone et al. 2013). Positional cloning of
R-genes and QRLs is still a challenging task in wheat.
Only two partial resistance genes are cloned so far in
wheat, shedding some light on the mechanisms
involved (Yr36, Fu et al. 2009; Lr34, Krattinger
et al. 2009). Yr36 provides partial resistance to all
stripe rust races tested, while Lr34 confers resistance
to multiple pathogens. Hence, we assume that the
resistance mechanisms conferred by the QRLs
described in the current study are different from those
of Yr36 and Lr34. Further study is needed to reveal the
genetic and physiological differences between quali-
tative and quantitative disease resistance.
While the reaction to Bgt#66 expresses a partial
resistance response of the domesticated parent and
susceptible response of the wild parent, the response
to another isolate, Bgt#15, was susceptibility in
LDN and resistance in G18-16. This response
facilitated the mapping of an effective R-gene
(PmG16) on chr. 7A (Ben-David et al. 2010). Using
the same mapping population as in this study,
PmG16 was genetically and physically mapped to
the long arm of chromosome 7A, on wheat
chromosome deletion bin 7AL-16 0.86-1.00. Hence,
we have used the same genetic platform to identify
and map a new powdery mildew resistance gene
from wild emmer and to map new QRLs derived
from durum wheat. The high potential of wild
emmer wheat as a reservoir for new powdery
mildew resistance genes has been shown previously
(Ben-David et al. 2010; Xie et al. 2012). In contrast,
durum wheat is underrepresented in the current
published Pm catalog and could be an important
genetic source.
In conclusion, the genetic analysis of the pheno-
typic reactions of the RIL population at the seedling
stage to the two Bgt isolates describes two different
resistance mechanisms. The first is monogenic: a
wild emmer wheat allele in a single locus conferring
complete resistance to Bgt#15. The second is
polygenic: a set of durum wheat alleles, in five
independent QTLs, controls partial resistance to
Bgt#66 in the RIL population. Bgt belongs to a
group of pathogens, which holds the highest risk to
wheat crops due to their dual reproduction system
and a high degree of gene/genotype flow (McDonald
and Linde 2002). Thus, breeding efforts should be
focused on sources of quantitative resistance that
will need to be renewed regularly in order to be able
to stay ahead of the pathogen. The identified new
resistance alleles from durum wheat could contrib-
ute to wheat breeding for Bgt resistance by precise
exploitation of the available and well-studied LDN
genetic platform. The results of the current study
shed light on evolutionary mechanisms associated
with the development and distribution of powdery
mildew resistance genes in wheat.
Acknowledgments This study was supported by The Israel
Science Foundation Grant #205/08 and equipment Grants
1478/04 and #1719/08. The authors thank A. Fahoum and M.
Chatzav for their excellent technical assistance and to Dr.
T. Kis-Papo for scientific editing of the manuscript.
1656 Mol Breeding (2014) 34:1647–1658
123

Conflict of interest All authors of the manuscript have
declared no conflicts of interest.
References
Bahadur P, Sinha VC, Ruiker SK, Upadhyaya YM (1977)
Horizontal and vertical resistance in wheat-exotic durums.
Indian J Genet Plant Breed 37:328–334
Bahadur P, Sinha VC, Ruiker SK, Upadhyaya YM (1979)
Sources of resistance to rusts and powdery mildew in
wheat. Indian J Genet Plant Breed 39:402–411
Ben-David R (2011) Molecular mapping of powdery mildew
resistance genes derived from the Triticum turgidum gene
pool. Ph.D. thesis submitted to The University of Haifa,
Haifa, Israel
Ben-David R, Xie W, Peleg Z, Saranga Y, Dinoor A, Fahima T
(2010) Identification and mapping of powdery mildew
resistance gene PmG16, derived from wild emmer wheat,
Triticum dicoccoides. Theor Appl Genet 121:499–510
Benjamini Y, Hochberg Y (1995) Controlling the false discov-
ery rate: a practical and powerful approach to multiple
testing. J R Stat Soc Series B Stat Methodol 57:289–300
Bennett FGA (1984) Resistance to powdery mildew in wheat: a
review of its use in agriculture and breeding programmes.
Plant Pathol 33:279–300
Bougot Y, Lemoine J, Pavoine MT, Guyomar’ch H, Gautier V,
Muranty H, Barloy D (2006) A major QTL effect con-
trolling resistance to powdery mildew in winter wheat at
the adult plant stage. Plant Breed 125:550–556
Braun U, Cook RTA, Inman AJ, Shin HD (2000) The taxonomy
of the powdery mildew fungi. In: Belanger RR, Bushnell
WR, Aleid JD, Carver TLW (eds) The powdery mildews: a
comprehensive treatise. APS Press, St. Paul, MN, USA,
pp 13–55
Briggle LW (1960) Source of resistance to Erysiphe graminis f.
sp. tritici from tetraploid Triticum species. Agron Abstr
1960–65, 45
Brodny U, Nelson RR, Gregory LV (1986) The residual and
interactive expression of ‘‘defeated’’ wheat stem rust
resistance genes. Phytopathology 76:546–549
Chantret N, Mingeot D, Sourdille P, Bernard M, Jacquemin JM,
Doussinault G (2001) A major QTL for powdery mildew
resistance is stable over time and at two development
stages in winter wheat. Theor Appl Genet 103:962–971
Churchill GA, Doerge RW (1994) Empirical threshold values
for quantitative trait mapping. Genetics 138:963–971
Develey-Riviere M-P, Galiana E (2007) Resistance to patho-
gens and host developmental stage: a multifaceted rela-
tionship within the plant kingdom. New Phytol
175:405–416
Elias EM, Manthey FA (2005) End products. In: Royo C, Nachit
MN, Di Fonzo N, Araus JL, Pfeiffer WH, Slafer GA (eds)
Durum wheat breeding. Current approaches and future
strategies. Food Academic Press, The Haworth Press, New
York, pp 63–86
FAOstat (2012) Available by food and agriculture organization.
http://faostat.fao.org/. Accessed 17 Sept 2012
Fu D, Uauy C, Distelfeld A, Blechl A, Epstein L, Chen X, Sela
H, Fahima T, Dubcovsky J (2009) A kinase-START gene
confers temperature-dependent resistance to wheat stripe
rust. Science 323:1357–1360
Hsam SLK, Zeller FJ (2002) Breeding for powdery mildew
resistance in common wheat (Triticum aestivum L.). In:
Belanger RR, Bushnell WR, Dik AJ, Carver TLW (eds)
The powdery mildews, a comprehensive treatise. APS
Press, St. Paul, pp 219–238
Huang XQ, Hsam SLK, Mohler V, Roder MS, Zeller F (2004)
Genetic mapping of three alleles at the Pm3 locus confer-
ring powdery mildew resistance in common wheat (Triti-
cum aestivum L.). Genome 47:1130–1136
Ji X, Xie C, Ni Z, Yang T, Nevo E, Fahima T, Liu Z, Sun Q
(2008) Identification and genetic mapping of a powdery
mildew resistance gene in wild emmer (Triticum dicocco-
ides) accession IW72 from Israel. Euphytica 159:385–390
Kao CH, Zeng ZB, Teasdale RD (1999) Multiple interval map-
ping for quantitative trait loci. Genetics 152:1203–1216
Korol AB, Ronin YI, Itskovich AM, Peng J, Nevo E (2001)
Enhanced efficiency of quantitative trait loci mapping
analysis based on multivariate complexes of quantitative
traits. Genetics 157:1789–1803
Krattinger SG, Lagudah ES, Spielmeyer W, Singh RP, Huerta-
Espino J, McFadden H, Bossolini E, Selter LL, Keller B
(2009) A putative ABC transporter confers durable resis-
tance to multiple fungal pathogens in wheat. Science
323:1360–1363
Li Z, Lan C, He Z, Singh RP, Rosewarne GM, Chen X, Xia X
(2014) Overview and application of QTL for adult plant
resistance to leaf rust and powdery mildew in wheat. Crop
Sci 54:1907–1925
Liang SS, Suenaga K, He ZH, Wang ZL, Liu HY, Wang DS,
Singh RP, Sourdille P, Xia XC (2006) Quantitative trait
loci mapping for adult-plant resistance to powdery mildew
in bread wheat. Phytopathol 96:784–789
Lu Q, Bjørnstad A, Ren Y, Asad M, Xia X, Chen X, Ji F, Shi J,
Lillemo M (2012) Partial resistance to powdery mildew in
German spring wheat ‘Naxos’ is based on multiple genes
with stable effects in diverse environments. Theor Appl
Genet 110:1401–1409
Maccaferri M, Cane’ MA, Sanguineti MC, Salvi S, Colalongo
MC, Massi A, Clarke F, Knox R, Pozniak CJ, Clarke JM,
Fahima T, Dubcovsky J, Xu S, Ammar K, Karsai I, Vida G,
Tuberosa R (2014a) A consensus framework map of durum
wheat (Triticum durum Desf.) suitable for linkage dis-
equilibrium analysis and genome-wide association map-
ping. BMC Genomics 15:873
Maccaferri M, Ricci A, Salvi S, Milner SG, Noli E, Martelli PL,
Casadio R, Akhunov E, Scalabrin S, Vendramin V, Ammar
K, Blanco A, Desiderio F, Distelfeld A, Dubcovsky J,
Fahima T, Faris J, Korol A, Massi A, Mastrangelo AM,
Morgante M, Pozniak CJ, N’Diaye A, Xu S, Tuberosa R
(2014b) A high-density, SNP-based consensus map of
tetraploid wheat as a bridge to integrate durum and bread
wheat genomics and breeding. Plant Biotech J (in press)
Marone D, Russo MA, Laido G, De Vita P, Papa R, Blanco A,
Mastrangelo AM (2013) Genetic basis of qualitative and
quantitative resistance to powdery mildew in wheat: from
consensus regions to candidate genes. BMC Genom 14:562
McDonald BA, Linde C (2002) Pathogen population genetics,
evolutionary potential, and durable resistance. Annu Rev
Phytopathol 40:349–379
Mol Breeding (2014) 34:1647–1658 1657
123

Miedaner T, Korzun V (2012) Marker-Assisted selection for
disease resistance in wheat and barley breeding. Phytopa-
thology 102:560–566
Nachit MM, Elouafi I, Pagnotta MA, Elsaleh A, Lacono E,
Labhilili M, Asbati AP, Azrak M, Hazzam H, Benscher D,
Khairallah M, Ribaut JM, Tanzarella OA, Porceddu E,
Sorrels ME (2001) Molecular linkage map for an intra-
specific recombinant inbred population of durum wheat
(Triticum turgidum L. var. durum). Theor Appl Genet
102:177–186
Nass HA, Pedersen WL, MacKenzie DR, Nelson RR (1981) The
residual effects of some ‘‘defeated’’ powdery mildew
resistance genes in isolines of winter wheat. Phytopathol-
ogy 71:1315–1318
Oerke EC (2006) Crop losses to pests. J Agric Sci 144:31–43
Parlevliet JE (2002) Durability of resistance against fungal,
bacterial and viral pathogens: present situation. Euphytica
124:147–156
Parlevliet JE, Zadoks JC (1977) The integrated concept of dis-
ease resistance: a new view including horizontal and ver-
tical resistance in plants. Euphytica 26:5–21
Peleg Z, Saranga Y, Suprunova T, Ronin Y, Roder MS, Kilian
A, Korol AB, Fahima T (2008) High-density genetic map
of durum wheat 9 wild emmer wheat based on SSR and
DArT markers. Theor Appl Genet 117:103–115
Peleg Z, Cakmak I, Ozturk L, Yazici A, Jun Y, Budak H, Korol
A, Fahima T, Saranga Y (2009a) Quantitative trait loci
conferring grain mineral nutrient concentrations in durum
wheat 9 wild emmer wheat RIL population. Theor Appl
Genet 119:353–369
Peleg Z, Fahima T, Krugman T, Abbo S, Yakir D, Korol AB,
Saranga Y (2009b) Genomic dissection of drought resis-
tance in durum wheat 9 wild emmer wheat recombinant
inbreed line population. Plant, Cell Environ 32:758–779
Peleg Z, Fahima T, Korol AB, Abbo S, Saranga Y (2011)
Genetic analysis of wheat domestication and evolution
under domestication. J Exp Bot 62:5051–5061
Peng JH, Korol AB, Fahima T, Roder MS, Ronin YI, Li YC,
Nevo E (2000) Molecular genetic maps in wild emmer
wheat, Triticum dicoccoides: genome-wide coverage,
massive negative interference, and putative quasi-linkage.
Genome Res 10:1509–1531
Poland JA, Balint-Kurti PJ, Wisser Randall J, Pratt RC, Nelson
RJ (2008) Shades of gray: the world of quantitative disease
resistance. Trend Plant Sci 14:21–29
Ronin YI, Korol AB, Nevo E (1999) Single- and multiple-trait
mapping analysis of linked quantitative trait loci: some
asymptotic analytical approximations. Genetics 151:387–396
Sourdille P, Robe P, Tixier MH, Doussinault G, Pavoine MT,
Bernard M (1999) Location of Pm3 g, a powdery mildew
resistance allele in wheat, by using a monosomic analysis
and by identifying associated molecular markers. Euphy-
tica 110:193–198
Sourdille P, Cadalen T, Guyomarc’h H, Snape JW, Perretant
MR, Charmet G, Boeuf C, Bernard S, Bernard M (2003) An
update of the Courtot 9 Chinese Spring intervarietal
molecular marker linkage map for the QTL detection of
agronomic traits in wheat. Theor Appl Genet 106:530–538
Tanksley SD (1993) Mapping polygenes. Annu Rev Genet
27:205–233
Tester M, Langridge P (2010) Breeding technologies to increase
crop production in a changing world. Science 327:818–822
Tucker DM, Griffey CA, Liu S, Brown-Guedira G, Marshall DS,
Maroof MAS (2007) Confirmation of three quantitative
trait loci conferring adult plant resistance to powdery
mildew in two winter wheat populations. Euphytica
155:1–13
Upadhyay MK, Kumar R, Singhal NC (1972) Sources of
resistance to powdery mildew of wheat. Indian J Genet
Plant Breed 32:242–246
Wang ZL, Li LH, He ZH, Duan XY, Zhou YL, Chen XM,
Lillemo M, Singh RP, Wang H, Xia XC (2005) Seedling
and adult plant resistance to powdery mildew in Chinese
bread wheat cultivars and lines. Plant Dis 89:457–463
Xie W, Ben-David R, Zeng B, Distelfeld A, Roder MS, Dinoor
A, Fahima T (2012) Identification and characterization of a
novel powdery mildew resistance gene PmG3M derived
from wild emmer wheat, Triticum dicoccoides. Theor Appl
Genet 124:911–922
Young ND (1996) QTL mapping and quantitative disease
resistance in plants. Annu Rev Phytopathol 34:479–501
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for
the growth stages of cereals. Weed Res 14:415–421
Zhu ZD, Kong XY, Zhou RH (2004) Identification and micro-
satellite markers of a resistance gene to powdery mildew in
common wheat introgressed from Triticum durum. Acta
Botanica Sinica 46:867–872
1658 Mol Breeding (2014) 34:1647–1658
123





























