Embed Size (px)
Citation preview

ARTHRITIS & RHEUMATISMVol. 43, No. 5, May 2000, pp 1156–1164© 2000, American College of Rheumatology
GENETIC ENHANCEMENT OF MATRIX SYNTHESISBY ARTICULAR CHONDROCYTES
Comparison of Different Growth Factor Genes in the Presence and Absence of Interleukin-1
P. SMITH, F. D. SHULER, H. I. GEORGESCU, S. C. GHIVIZZANI, B. JOHNSTONE, C. NIYIBIZI,P. D. ROBBINS, and C. H. EVANS
Objective. To determine whether articular chon-drocytes express growth factor genes delivered by ad-enoviral vectors and whether expression of these genesinfluences matrix synthesis in the presence and absenceof interleukin-1 (IL-1).
Methods. Monolayer cultures of rabbit articularchondrocytes were infected with recombinant adenovi-rus carrying genes encoding the following growth fac-tors: insulin-like growth factor 1 (IGF-1), transforminggrowth factor b1 (TGFb1), and bone morphogeneticprotein 2 (BMP-2). As a control, cells were transducedwith the lac Z gene. Cultures were also treated with eachgrowth factor supplied as a protein. Levels of geneexpression were noted, and the synthesis of proteogly-can, collagen, and noncollagenous proteins was mea-sured by radiolabeling. Collagen was typed by sodiumdodecyl sulfate–polyacrylamide gel electrophoresis andautoradiography. The effects of growth factor genetransfer on proteoglycan synthesis in the presence ofIL-1 were also measured.
Results. The expression of all transgenes was highfollowing adenoviral transduction. Proteoglycan synthe-sis was stimulated ;8-fold by the BMP-2 gene and2–3-fold by the IGF-1 gene. The effects of BMP-2 andIGF-1 genes were additive upon cotransduction. Synthe-
sis of collagen and noncollagenous proteins, in contrast,was most strongly stimulated by the IGF-1 gene. In eachcase, collagen typing confirmed the synthesis of type IIcollagen. IL-1 suppressed proteoglycan synthesis by50–60%. IGF-1 and TGFb genes restored proteoglycansynthesis to control levels in the presence of IL-1. TheBMP-2 gene, in contrast, elevated proteoglycan synthe-sis beyond control levels in the presence of IL-1.
Conclusion. Transfer of growth factor genes toarticular chondrocytes can greatly increase matrix syn-thesis in vitro, even in the presence of the inflammatorycytokine IL-1. This result encourages the further devel-opment of gene therapy for the repair of damagedcartilage.
Articular cartilage is frequently damaged as aresult of disease or injury, yet it has poor regenerativeproperties (1,2). Indeed, partial-thickness lesions mounta negligible restorative response and persist indefinitely(3). Full-thickness lesions that penetrate the subchon-dral bone heal through the ingress of cells from theunderlying blood vessels and bone marrow (4). Al-though the repair tissue that forms in this way hascertain similarities to articular cartilage, it is essen-tially a scar, which ultimately fails. There is clearly apressing clinical need for improved methods of carti-lage repair (1,2).
One promising approach to achieving this aimrelies upon the abilities of certain growth factors toincrease the synthesis and deposition of extracellularmatrix components by chondrocytes. Prominent in thisregard are transforming growth factor b (TGFb),insulin-like growth factor 1 (IGF-1), bone morphoge-netic proteins (BMPs) 2 and 7, fibroblast growth factors,and hepatocyte growth factor (5–10). Despite the favor-
Supported in part by NIAMS grant AR-43820.P. Smith, MD, F. D. Shuler, MD, H. I. Georgescu, BS, S. C.
Ghivizzani, PhD, C. Niyibizi, PhD, P. D. Robbins, PhD, C. H. Evans,PhD, DSc: University of Pittsburgh School of Medicine, Pittsburgh,Pennsylvania; B. Johnstone, PhD: Case Western Reserve University,Cleveland, Ohio.
Drs. Evans and Robbins are both consultants for Valentis, Inc.Address reprint requests to C. H. Evans, PhD, DSc, Center
for Molecular Orthopedics, Harvard Medical School, 221 LongwoodAvenue, BL-152, Boston, MA 02115.
Submitted for publication August 31, 1999; accepted inrevised form December 17, 1999.
1156

able biologic properties of these molecules, their clinicalapplication is hindered by delivery problems. In partic-ular, it is difficult to maintain therapeutic levels ofgrowth factors locally within the lesion for the lengthof time that is likely to be needed to achieve repair.For example, the half-life of BMP-2 after introductioninto a full-thickness lesion in rabbit articular cartilage is4 days (11). Various types of slow-release devices arebeing developed and evaluated in response to thisproblem. Gene transfer offers an alternative potentialsolution (12).
Gene-based approaches to cartilage repair in-volve the transfer of the appropriate genes to cellswithin the lesion. The simplest approach would be tointroduce the vectors intraarticularly by direct in vivoinjection. Although certain vectors may be able totransduce articular chondrocytes in situ by this means(13), they also transduce synovium with high efficiencyand do not appear to localize at sites of cartilagedamage. Ex vivo approaches, although more tedious, canbe designed to target areas of damage and allow theemployment of a greater range of vectors. Moreover, exvivo methods involve the introduction of cells into thelesion (14). This may be important because there isextensive death of chondrocytes in rheumatoid arthritisand osteoarthritis (15), and even in healthy cartilage,chondrocytes adjacent to sites of injury undergo apopto-sis (16).
Ex vivo cellular treatment of cartilaginous lesionsis already in clinical use (17). In these procedures,autologous cartilage is harvested, and the chondrocytesare expanded in culture and then transplanted into thedamaged site. Ex vivo gene treatment of cartilaginouslesions is a logical extension of this procedure (14). Thefeasibility of ex vivo gene transfer to experimentallyproduced defects in articular cartilage has already beendemonstrated in laboratory animals (18–20). Moreover,in preliminary studies, in vitro transfer to chondrocytesof complementary DNA (cDNA) encoding TGFb1 wasshown to enhance matrix synthesis (21,22). As a preludeto studies in experimental animals, it is necessary todetermine which gene, or combinations of genes, givesthe best result.
In the present experiments, we compared IGF-1,BMP-2, and TGFb1. Because interleukin-1 (IL-1), apowerful inhibitor of matrix synthesis (23), is likely to bepresent in diseased and injured joints, we also evaluatedthe ability of these growth factors to enhance matrixsynthesis in the presence of IL-1.
MATERIALS AND METHODS
Adenoviral vectors. First-generation, DE1DE3, sero-type 5 adenoviral vectors were used for these studies. Trans-genes encoding human IGF-1, BMP-2, TGFb1, or, as acontrol, the lac Z gene of Escherichia coli, were inserted intothe E1 region of the virus by cre-lox recombination (24). Ineach case, gene expression was driven by the human cytomeg-alovirus early promoter. The resulting vectors were designatedAdIGF-1, AdBMP-2, AdTGFb1, and AdLacZ, respectively.
Cell isolation and transduction. Articular chondro-cytes were isolated from articular cartilage recovered from theknee and shoulder joints of skeletally mature NZW rabbits,using the method described by Green (25). Cells were seededat a density of 2 3 104 cells/well into monolayer cultures on24-well plates using Ham’s F-12 nutrient medium supple-mented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin in a humidified atmosphere of 5% CO2 at 37°C.The cultures became confluent after 7–10 days, and typicallycontained 2 3 105 cells/well.
Confluent cells were rinsed twice with sterile Gey’sbalanced salt solution (GBSS) to remove traces of serum inpreparation for adenoviral transduction. Cells were transducedwith adenoviruses that contained genes encoding IGF-1,TGFb1, BMP-2, or b-galactosidase (lac Z gene). Transductionwas performed in 300 ml of GBSS for 1 hour at 37°C at variousmultiplicities of infection (MOIs), as indicated in the text.Following transduction, the supernatant was aspirated andreplaced with 500 ml of Dulbecco’s modified Eagle’s medium(DMEM) supplemented with 1% FBS and antibiotics.
Exogenous recombinant human IGF-1 (R&D Systems,Minneapolis, MN) or recombinant human BMP-2 (a kind giftfrom Genetics Institute, Cambridge, MA) protein was addedto the media of the nontransduced control groups. Recombi-nant human IL-1b (2 ng/ml) was also added to the appropriategroups at this time. Cells were then incubated for 48 hoursbefore the addition of radionucleotides for the determinationof matrix synthesis.
IGF-1 and BMP-2 measurements. The concentrationof IGF-1 in cell supernatants was determined at 48 hoursfollowing transduction using a commercially available enzyme-linked immunosorbent assay (ELISA) kit and protocol (Diag-nostics Laboratory Systems, Webster, TX).
The BMP-2 concentration was also determined byELISA at 48 hours following transduction. Because only 1anti–BMP-2 antibody was available, purified BMP-2 standardsor samples containing BMP-2 were preadsorbed onto 96-wellplates by overnight incubation at 4°C. The plates were thenwashed thoroughly with phosphate buffered saline (PBS),blocked by incubating with 3% (weight/volume) bovine serumalbumin (BSA) for 4 hours at 4°C, and incubated overnightwith 100 ml of primary antibody (murine anti-human BMP-2;kindly supplied by Genetics Institute) diluted 1:1,000 in 1%BSA. Plates were then washed with PBS containing 0.05%Tween 20 and incubated with secondary antibody (goat anti-mouse IgG) conjugated to alkaline phosphatase. After 2 hours’incubation at room temperature, plates were washed, incu-bated with substrate, and the optical density at 405 nm wasread on a plate reader.
ARTICULAR CARTILAGE AND GROWTH FACTOR GENES 1157

Proteoglycan and collagen synthesis. The synthesis ofmatrix molecules was measured by previously published meth-ods (26,27). To measure proteoglycan synthesis, the mediawere aspirated following the initial 48-hour incubation periodand replaced with 500 ml of fresh DMEM with 1% FBS.Exogenous growth factors and IL-1 were also readministeredto the appropriate wells. Each culture well received 20 mCi/mlof 35S-labeled Na2SO4, and the cells were incubated for anadditional 24 hours. The synthesis of proteoglycans was deter-mined in the cell layer and the media by the incorporation of35SO4
22 into glycosaminoglycans. Size-exclusion chromatogra-phy with PD10 columns (Pharmacia, Piscataway, NJ) was usedto separate the glycosaminoglycans from unincorporated label.
Protein synthesis was determined by the incorporationof 3H-proline into collagen and noncollagenous proteins in thefollowing manner. When the cells reached confluence, themedium was removed and replaced with serumless mediumcontaining 50 mg/ml of ascorbic acid, 50 mg/ml ofb-aminopropionitrile, and 20 mCi/ml of 3H-proline for 24hours. After incubation, the conditioned medium and cell layerwere collected and combined, and the relative collagen synthe-sis was determined by 3H-proline incorporation using a mod-ified collagenase digestion method.
Briefly, after 24 hours of culture in the presence of theradioisotope, a carrier protein (2 mg/ml of pepsin-solubilizedbovine type II collagen) was added, and the proteins wereprecipitated by addition of 10% trichloroacetic acid (TCA).The TCA-precipitated proteins were recovered by centrifuga-tion at 14,000 revolutions per minute for 30 minutes at 4°C.The recovered protein pellets were washed repeatedly with 5%TCA to remove the unbound isotope. The washed pellets wereresuspended in the collagenase buffer (5 mM CaCl2 and 3 mMN-ethylmaleimide), the pH was adjusted to 7, and the suspen-sions were incubated at 37°C for 2 hours. TCA was then added,and the mixture was centrifuged for 30 minutes at 14,000 rpm.The supernatants were removed and were used as enzymeblanks. The pellets were resuspended in the collagenase buffer,the pH was adjusted to 7 by the addition of NaOH, andpurified bacterial collagenase ABC form III (Advanced Bio-factures, Lynbrook, NY) was added (20 units/ml) and incu-bated for 2 hours at 37°C.
The collagenase-digestible proteins were separatedfrom nondigested proteins by the addition of 5% TCA fol-lowed by centrifugation at 14,000 rpm for 30 minutes. Thepellets were redissolved in 0.2M NaOH, and aliquots of thepellets and the radioactivities of the supernatants were mea-sured to determine the level of collagen and noncollagenousprotein synthesis. The data were corrected for cell number bycounting cells in parallel wells that received the same factorsand were plated at the same density as in the experimentalwells.
Determination of collagen phenotype. Collagen phe-notyping was also performed on each sample, as describedelsewhere (27). Briefly, following the 24-hour labeling periodwith 3H-proline, all samples underwent pepsin (100 mg/ml)digestion for 4 hours. The pepsin-resistant chains were precip-itated with 3M NaCl followed by dialysis against NH4HCO3 for5 days. Samples were then dried and separated by 7.5% sodiumdodecyl sulfate–polyacrylamide gel electrophoresis. Gels weretreated with EN3HANCE (DuPont NEN, Boston, MA) priorto autoradiography.
RESULTS
IGF-1 gene expression and matrix synthesis.Resting cultures of rabbit articular chondrocytes se-creted 3.9 6 1.7 ng/ml (mean 6 SD) IGF-1 into theirculture media. Infection with AdIGF-1 increased IGF-1production dramatically, in a dose-dependent manner(Figure 1A). Experiments with the AdLacZ vector con-firmed that at an MOI of 300, all chondrocytes had beentransduced (results not shown); the number of viralgenomes per cell under these conditions is not known.IGF-1 protein was added to certain cultures to confirmthe accuracy of the ELISA. Of interest is the observationthat the recovery of IGF-1 was always much greater than100% (Figure 1A). This discrepancy could not be ac-counted for by the presence of IGF-1 in the serum(results not shown) and is most likely explained by theability of IGF-1 to induce its own synthesis.
Monolayer cultures of chondrocytes readily in-corporated radiolabeled precursors into matrix macro-molecules. In multiple experiments, the radioactivities ofthe various matrix molecules (mean 6 SD) were asfollows: for proteoglycan in the cell layer 4,620 6 1,104disintegrations per minute, proteoglycan in the medium7,465 6 692 dpm, collagen 2,415 6 354 dpm, andnoncollagenous proteins 42,487 6 4,719. Infection ofchondrocytes with AdLacZ did not alter matrix synthesisby chondrocytes (results not shown).
Both IGF-1 protein and transduction with IGF-1cDNA enhanced proteoglycan synthesis ;2–3-fold (Fig-ure 1B). Due to biologic variability, statistical signifi-cance was only achieved at the highest concentration ofIGF-1 protein and 300 MOI of virus. Proteoglycansynthesis declined at the highest MOI, despite the highlevels of IGF-1 produced at this dose of virus. In allcases, increased proteoglycan deposition was noted inthe cell layer, as well as in the conditioned media,suggesting that the newly synthesized proteoglycan pro-duced in response to IGF-1 was incorporated into astable, cell-associated matrix.
IGF-1 protein had a more modest effect on thesynthesis of collagen and noncollagenous proteins by thechondrocytes, but the stimulatory effects of transducingthe IGF-1 cDNA were very marked (Figure 1C). Even atan MOI of 100, where IGF-1 accumulated in the me-dium to ;80 ng/ml (Figure 1A), an approximately 3-foldincrement was seen (Figure 1C). Addition of 100 ng/mlof protein, in contrast, achieved only about a 1.5-foldincrement, despite the presence of ;160 ng/ml IGF-1 inthe culture medium (Figure 1A).
1158 SMITH ET AL

Collagen typing confirmed that IGF-1, whetherdelivered as a protein or as a gene, maintained thephenotype of the chondrocytes (Figure 1D). The pre-dominant collagen produced by the cells under allconditions was type II collagen, with barely detectableamounts of a2(I), which would indicate the synthesis oftype I collagen. A prominent band was seen at theposition where the a1(V) and a1(XI) chains comigrate,
indicating the presence of types V and XI collagens.Articular chondrocytes are known to produce both ofthese types of collagens.
BMP-2 gene expression and matrix synthesis.Resting cultures of rabbit articular chondrocytes ac-cumulated 15.5 6 2.3 ng/ml (mean 6 SD) of BMP-2in their culture media. Infection with the AdBMP-2virus increased BMP-2 production dramatically
Figure 1. Production of insulin-like growth factor 1 (IGF-1) by transduced chondrocytes and its effects on matrix synthesis. A, Content of IGF-1in conditioned media following transduction with the IGF-1 gene and addition of IGF-1 protein. Cultures of chondrocytes were infected withAdIGF-1 at the indicated multiplicity of infection (MOI) or were supplemented with IGF-1 at the indicated dosages. After 48 hours, all conditionedmedia were assayed for IGF-1. B, Effects of IGF-1 protein and gene on the synthesis of proteoglycans by chondrocytes. Incorporation of 35SO4
22
into proteoglycans present in the conditioned media and the cell layer was measured under the indicated conditions (columns are stacked). C, Effectsof IGF-1 protein and gene on the synthesis of collagen and noncollagenous proteins by chondrocytes. Incorporation of 3H-proline intocollagenase-sensitive and collagenase-resistant proteins was measured under the indicated conditions. Values are the mean and SD (n 5 6). p 5 P ,0.05 versus control. D, Effects of IGF-1 protein and gene on the types of collagen synthesized by chondrocytes. Chondrocytes were incubated with3H-proline under the conditions indicated. Pepsin-resistant proteins were then separated by sodium dodecyl sulfate–polyacrylamide gelelectrophoresis and subjected to autoradiography.
ARTICULAR CARTILAGE AND GROWTH FACTOR GENES 1159

(Figure 2A). Unlike IGF-1, BMP-2 did not appear toinduce its own synthesis.
BMP-2 had a very large effect on proteoglycansynthesis (Figure 2B). Both the recombinant proteinsand the gene produced increases of 6–9-fold in both thesoluble and cell-associated fractions. The effects ofBMP-2 on collagen synthesis were more modest, with a2–3-fold increase occurring under all conditions (Figure
2C). For unknown reasons, there was no evidence of adose response.
Recombinant BMP-2 did, however, provoke alarge increase in the synthesis of noncollagenous pro-teins, but, oddly, this was not seen with the BMP-2 gene(Figure 2C). The reason for this is obscure.
Collagen typing (Figure 2D) confirmed the main-tenance of the type II collagen phenotype. It also
Figure 2. Production of bone morphogenetic protein 2 (BMP-2) by transduced chondrocytes and its effects on matrix synthesis. A, Content ofBMP-2 in conditioned media following transduction with the BMP-2 gene and addition of BMP-2 protein. Cultures of chondrocytes were infectedwith AdBMP-2 at the indicated multiplicity of infection (MOI) or were supplemented with BMP-2 at the indicated dosages. After 48 hours, allconditioned media were assayed for BMP-2. B, Effects of BMP-2 protein and gene on the synthesis of proteoglycans by chondrocytes. Incorporationof 35SO4
22 into proteoglycans present in the conditioned media and the cell layer was measured under the indicated conditions (columns arestacked). C, Effects of IGF-1 protein and gene on the synthesis of collagen and noncollagenous protein by chondrocytes. Incorporation of 3H-prolineinto collagenase-sensitive and collagenase-resistant proteins was measured under the indicated conditions. Values are the mean and SD (n 5 6).p 5 P , 0.05 versus control. D, Effects of BMP-2 protein and gene on the types of collagen synthesized by chondrocytes. Chondrocytes wereincubated with 3H-proline under the indicated conditions. Pepsin-resistant proteins were then separated by sodium dodecyl sulfate–polyacrylamidegel electrophoresis and subjected to autoradiography.
1160 SMITH ET AL

confirmed, in an independent manner, the modest stim-ulatory effects of BMP-2 and the absence of a clear doseresponse as was noted in Figure 2C.
Growth factor combinations. From the datashown so far, a combination of AdIGF-1 and AdBMP-2seemed promising for further evaluation. This combina-tion was tested at 2 different MOIs. As shown in Figure3A, these 2 vectors had an additive effect on proteogly-can synthesis. A similar effect was noted with a combi-nation of AdIGF-1 and AdTGFb1 (Figure 3B), althoughthe maximum increase in proteoglycan synthesis was lessthan that achieved with equivalent amounts of AdIGF-1and AdBMP-2.
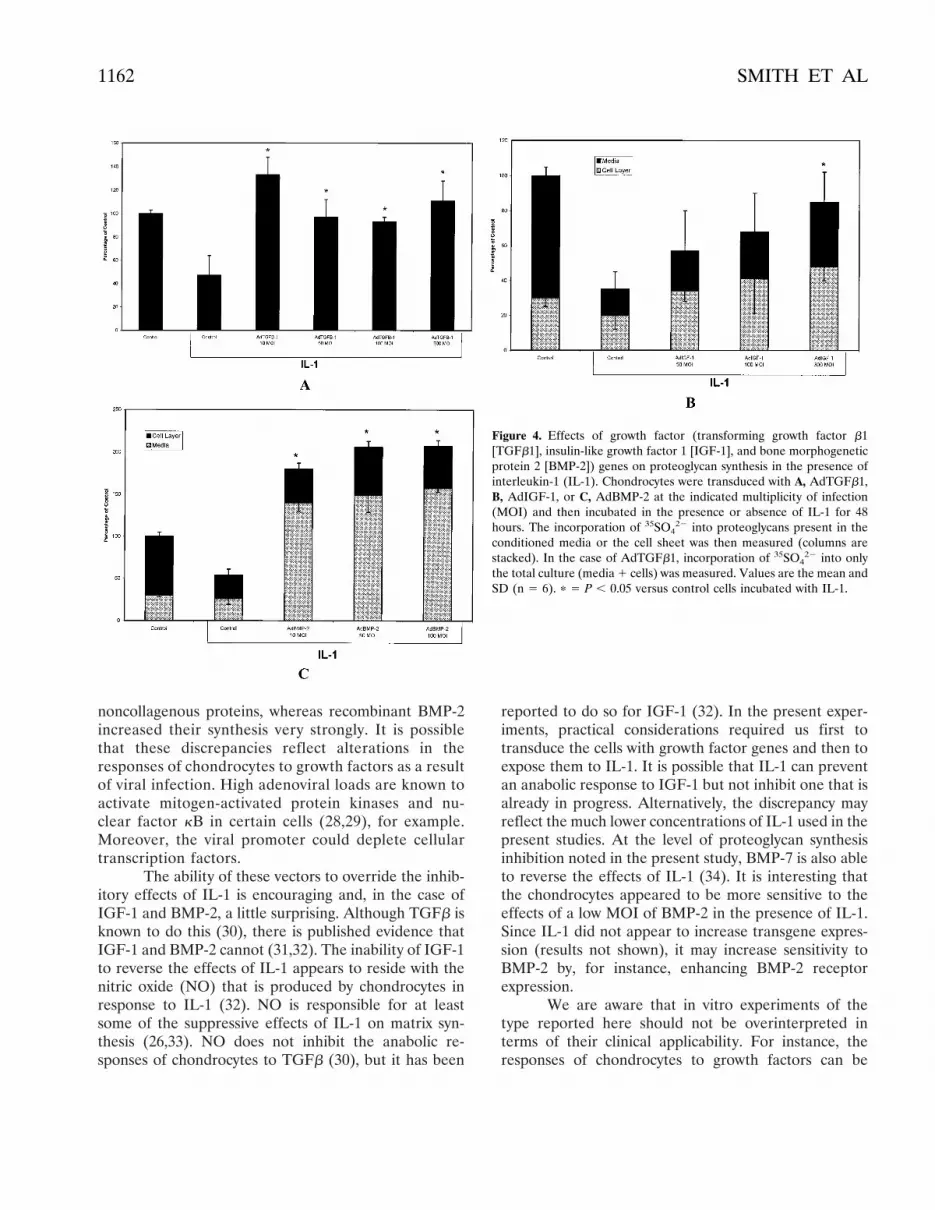
Effects of IL-1. It is well known that IL-1 inhibitsproteoglycan synthesis by articular chondrocytes (23).Both AdTGFb1 and AdIGF-1 were able to reverse thiseffect but were unable, at the MOIs used here, toincrease proteoglycan synthesis beyond the control level(Figures 4A and B). AdBMP-2, in contrast, not onlyreversed the suppressive effects of IL-1, but also in-creased proteoglycan synthesis to approximately doublethe control level (Figure 4C). Oddly, at an MOI of only10, AdBMP-2 produced a full stimulatory effect,whereas in the absence of IL-1, this concentration ofvirus was only weakly active (Figure 2B).
DISCUSSION
These data confirm that it is possible to gain largeincreases in matrix synthesis by the genetic modification
of chondrocytes. Transduction with AdBMP-2 provokeda particularly large increase in proteoglycan synthesis,and AdIGF-1 stimulated collagen synthesis very dramat-ically. This suggests that a combination of the two couldbe particularly useful. When combined, these 2 viruseshad an additive effect on proteoglycan synthesis, butcollagen synthesis was not evaluated.
Although AdTGFb1 is also able to increasemarkedly the synthesis of proteoglycan and collagen bychondrocytes (21,22), it needs to be used cautiouslywithin joints. Direct injection of AdTGFb1 into rabbits’knee joints provokes considerable pathology, includingsynovial thickening, inflammation, and loss of articularcartilage with concurrent cartilage formation in thesynovium (Mi Z et al: unpublished observations). Intra-articular injection of AdIGF-1 or AdBMP-2, in contrast,is not accompanied by adverse reactions of this type (MiZ et al: unpublished observations). However, proteogly-can synthesis by articular cartilage recovered from thesejoints is modestly elevated compared with the valuesreported here. This suggests that gene transfer to chon-drocytes might be more efficient than gene transfer tosynovium as a means of increasing matrix deposition.
In most cases, the responses of the chondrocytesto transfer of a growth factor gene were similar to thoseobtained with the growth factor itself, but there wereanomalies. AdIGF-1, for example, increased collagensynthesis much more strongly than did IGF-1 protein.AdBMP-2, in contrast, barely affected the synthesis of
Figure 3. Effects of growth factor (insulin-like growth factor 1 [IGF-1] and transforming growth factor b1 [TGFb1]) gene combinations onproteoglycan synthesis by chondrocytes. Cultures of chondrocytes were infected with a combination of A, AdIGF-1 and AdBMP-2 or B, AdIGF-1and AdTGFb1 at the indicated multiplicity of infection (MOI). The incorporation of 35SO4
22 into proteoglycans present in the conditionedmedium and cell layer was then measured (columns are stacked). Values are the mean and SD (n 5 6). p 5 P , 0.05 versus control.
ARTICULAR CARTILAGE AND GROWTH FACTOR GENES 1161
noncollagenous proteins, whereas recombinant BMP-2increased their synthesis very strongly. It is possiblethat these discrepancies reflect alterations in theresponses of chondrocytes to growth factors as a resultof viral infection. High adenoviral loads are known toactivate mitogen-activated protein kinases and nu-clear factor kB in certain cells (28,29), for example.Moreover, the viral promoter could deplete cellulartranscription factors.
The ability of these vectors to override the inhib-itory effects of IL-1 is encouraging and, in the case ofIGF-1 and BMP-2, a little surprising. Although TGFb isknown to do this (30), there is published evidence thatIGF-1 and BMP-2 cannot (31,32). The inability of IGF-1to reverse the effects of IL-1 appears to reside with thenitric oxide (NO) that is produced by chondrocytes inresponse to IL-1 (32). NO is responsible for at leastsome of the suppressive effects of IL-1 on matrix syn-thesis (26,33). NO does not inhibit the anabolic re-sponses of chondrocytes to TGFb (30), but it has been
reported to do so for IGF-1 (32). In the present exper-iments, practical considerations required us first totransduce the cells with growth factor genes and then toexpose them to IL-1. It is possible that IL-1 can preventan anabolic response to IGF-1 but not inhibit one that isalready in progress. Alternatively, the discrepancy mayreflect the much lower concentrations of IL-1 used in thepresent studies. At the level of proteoglycan synthesisinhibition noted in the present study, BMP-7 is also ableto reverse the effects of IL-1 (34). It is interesting thatthe chondrocytes appeared to be more sensitive to theeffects of a low MOI of BMP-2 in the presence of IL-1.Since IL-1 did not appear to increase transgene expres-sion (results not shown), it may increase sensitivity toBMP-2 by, for instance, enhancing BMP-2 receptorexpression.
We are aware that in vitro experiments of thetype reported here should not be overinterpreted interms of their clinical applicability. For instance, theresponses of chondrocytes to growth factors can be
Figure 4. Effects of growth factor (transforming growth factor b1[TGFb1], insulin-like growth factor 1 [IGF-1], and bone morphogeneticprotein 2 [BMP-2]) genes on proteoglycan synthesis in the presence ofinterleukin-1 (IL-1). Chondrocytes were transduced with A, AdTGFb1,B, AdIGF-1, or C, AdBMP-2 at the indicated multiplicity of infection(MOI) and then incubated in the presence or absence of IL-1 for 48hours. The incorporation of 35SO4
22 into proteoglycans present in theconditioned media or the cell sheet was then measured (columns arestacked). In the case of AdTGFb1, incorporation of 35SO4
22 into onlythe total culture (media 1 cells) was measured. Values are the mean andSD (n 5 6). p 5 P , 0.05 versus control cells incubated with IL-1.
1162 SMITH ET AL

strongly influenced by the surrounding matrix (35) andstate of differentiation (36). Moreover, during aging andwith the onset of osteoarthritis, chondrocytes lose theirresponsiveness to IGF-1 (37,38), whereas responses toTGFb are increased in osteoarthritis (39). Transfer of anIGF-1 gene may therefore be most applicable to cases ofcartilaginous injury in younger individuals. With BMP-2,there is a concern that ossification may occur (40).Nevertheless, the approaches described in this reportseem worthy of further development as a possible meansof addressing a serious and pressing clinical need.
ACKNOWLEDGMENTS
We thank Genetics Institute, Inc. for kindly supplyingthe BMP-2 protein, BMP-2 cDNA, and anti–BMP-2 antibodiesused in these studies, Dr. Janey Whalen for conducting theBMP-2 ELISA, and Mrs. Lou Duerring for kindly typing themanuscript.
REFERENCES
1. Buckwalter JA, Mankin HJ. Articular cartilage repair and trans-plantation. Arthritis Rheum 1998;41:1331–42.
2. O’Driscoll SW. The healing and regeneration of articular cartilage.J Bone Joint Surg Am 1998;80A:1795–812.
3. Meachim G. The effect of scarification on articular cartilage in therabbit. J Bone Joint Surg Br 1963;45B:150–61.
4. Meachim G, Roberts C. Repair of the joint surface from subar-ticular tissue in the rabbit knee. J Anat 1971;109:317–27.
5. Trippel SB. Growth factor actions on articular cartilage. J Rheu-matol 1995;43 Suppl:129–32.
6. Sellers RS, Peluso D, Morris EA. The effects of recombinanthuman bone morphogenetic protein-2 (rhBMP-2) on the healingof full-thickness defects of articular cartilage. J Bone Joint SurgAm 1997;79A:1452–63.
7. Morales TI. Transforming growth factor-b stimulates synthesis ofproteoglycan aggregates in calf articular cartilage organ cultures.Arch Biochem Biophys 1988;157:1352–9.
8. Tyler JA. Insulin-like growth factor 1 can decrease degradationand promote synthesis of proteoglycan in cartilage exposed tocytokines. Biochem J 1989;260:543–8.
9. Flechtenmacher J, Huch K, Thonar EJ-MA, Mollenhauer JA,Davies SR, Schmid TM, et al. Recombinant human osteogenicprotein 1 is a potent stimulator of the synthesis of cartilageproteoglycans and collagens by human articular chondrocytes.Arthritis Rheum 1996;39:1896–904.
10. Wakitani S, Imoto K, Kimura T, Ochi T, Matsumoto K, NakamuraT. Hepatocyte growth factor stimulated cartilage repair: fullthickness articular cartilage defect studied in rabbit knees. ActaOrthop Scand 1997;68:474–80.
11. Glasson SS, Kim HD, D’Augusta DA, Seeherman HJ, Morris EA.rhBMP-2 residence in full-thickness cartilage defects. 2nd Inter-national Symposium of the International Cartilage Repair Society;1998 Nov 15–18; Boston.
12. Evans CH, Robbins PD. Possible orthopaedic applications of genetherapy. J Bone Joint Surg Am 1995;77A:1103–14.
13. Tomita T, Hashimoto H, Tomita N, Morishita R, Lee SB, Ha-yashida K, et al. In vivo direct gene transfer into articular cartilageby intraarticular injection mediated by HVJ (Sendai virus) andliposomes. Arthritis Rheum 1997;40:901–6.
14. Evans CH, Robbins PD. Genetically augmented tissue engineeringof the musculoskeletal system. Clin Orthop Rel Res 1999;367S:S410–8.
15. Blanco FJ, Guitian R, Vazquez-Martul E, de Toro FJ, Galdo F.Osteoarthritis chondrocytes die by apoptosis: a possible pathwayfor osteoarthritis pathology. Arthritis Rheum 1998;41:284–9.
16. Tew SR, Kwan APL, Hann A, Thomsom BM, Archer CW. Thereactions of articular cartilage to experimental wounding: role ofapoptosis. Arthritis Rheum 2000;43:215–25.
17. Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, PetersonL. Treatment of deep cartilage defects in the knee with autologouschondrocyte transplantation. N Engl J Med 1994;331:889–95.
18. Kang R, Marui T, Ghivizzani SC, Nita IM, Georgescu HI, Suh JK,et al. Ex vivo gene transfer to chondrocytes in full-thicknessarticular cartilage defects: a feasibility study. Osteoarthritis Carti-lage 1997;5:139–43.
19. Baragi VM, Renkiewicz RR, Qiu L, Brammer D, Riley JM, SiglerRE, et al. Transplantation of adenovirally transduced allogeneicchondrocytes into articular cartilage defects in vivo. OsteoarthritisCartilage 1997;5:275–82.
20. Mason JM, Grande DA, Barcia M, Grant R, Pergolizza RG,Breitbart AS. Expression of human bone morphogenetic protein 7in primary rabbit periosteal cells: potential utility in gene therapyfor osteochondral repair. Gene Ther 1998;5:1098–104.
21. Arai Y, Kubo T, Kobayashi K, Takeshita K, Takahashi K, Ikeda T,et al. Adenovirus vector-mediated gene transduction to chondro-cytes: in vitro evaluation of therapeutic efficacy of transforminggrowth factor-b1 and heat shock protein 70 gene transduction.J Rheumatol 1997;24:1787–95.
22. Shuler FD, Georgescu HI, Niyibizi C, Studer RK, Mi Z, JohnstoneB, et al. Increased matrix synthesis following transfer of a trans-forming growth factor b1 gene into articular chondrocytes. J Or-thop Res. In press.
23. Tyler JA. Articular cartilage cultured with catabolin (piginterleukin-1) synthesizes a decreased number of normal proteo-glycan molecules. Biochem J 1985;227:869–78.
24. Hardy S, Kitamura M, Harris-Stansil T, Yumin D, Phillips ML.Construction of adenovirus vectors through Cre-Lox recombina-tion. J Virol 1997;71:1842–9.
25. Green WT. Behavior of articular chondrocytes in cell culture. ClinOrthop 1971;75:248–60.
26. Taskiran D, Stefanovic-Racic M, Georgescu HI, Evans CH. Nitricoxide mediates suppression of cartilage proteoglycan synthesis byinterleukin-1. Biochem Biophys Res Commun 1994;200:142–8.
27. Cao M, Westerhausen-Larson A, Niyibizi C, Kavalkovich K,Georgescu HI, Rizzo CF, et al. Nitric oxide inhibits the synthesisof type II collagen without altering COL2A1 mRNA abundance:prolyl hydroxylase as a possible target. Biochem J 1997;325:305–10.
28. Bruder JT, Kovesdi I. Adenovirus infection stimulates the Raf/MAPK signaling pathway and induces interleukin-8 expression.J Virol 1997;71:398–404.
29. Clesham GJ, Adam PJ, Proudfoot D, Flynn PD, Efstathiou S,Weissberg PL. High adenoviral loads stimulate NFkB-dependentgene expression in human vascular smooth muscle cells. GeneTher 1998;5:174–80.
30. Studer RK, Georgescu HI, Miller LA, Evans CH. Inhibition oftransforming growth factor b production by nitric oxide–treatedchondrocytes: implications for matrix synthesis. Arthritis Rheum1999;42:248–57.
31. Glansbeek HL, van Beuningen HM, Vitters EL, Morris EA, vander Kraan PM, van den Berg WB. Bone morphogenetic protein 2stimulates articular cartilage proteoglycan synthesis in vivo butdoes not counteract interleukin-1a effects on proteoglycan synthe-sis and content. Arthritis Rheum 1997;40:1020–8.
32. Van de Loo FAJ, Arntz OJ, van Enckevort FHJ, van Lent PLEM,van den Berg WB. Reduced cartilage proteoglycan loss duringzymosan-induced gonarthritis in NOS2-deficient mice and in
ARTICULAR CARTILAGE AND GROWTH FACTOR GENES 1163

anti–interleukin-1–treated wild-type mice with unabated joint in-flammation. Arthritis Rheum 1998;41:634–46.
33. Stefanovic-Racic M, Mollers MO, Miller LA, Evans CH. Nitricoxide and proteoglycan turnover in rabbit articular cartilage.J Orthop Res 1997;15:422–9.
34. Huch K, Wilbrink B, Flechtenmacher J, Koepp HE, AydelotteMB, Sampath TK, et al. Effects of recombinant human osteogenicprotein 1 on the production of proteoglycan, prostaglandin E2, andinterleukin-1 receptor antagonist by human articular chondrocytescultured in the presence of interleukin-1b. Arthritis Rheum 1997;40:2157–61.
35. Qi WN, Scully SP. Extracellular collagen modulates the regulationof chondrocytes by transforming growth factor-b1. J Orthop Res1997;15:483–90.
36. Lafeber FPJG, van Roy HLAM, van der Kraan PM, van den BergWB, Bijlsma JWJ. Transforming growth factor-b predominantly
stimulates phenotypically changed chondrocytes in osteoarthritichuman cartilage. J Rheumatol 1997;24:536–42.
37. Dore S, Pelletier J-P, DiBattista JA, Tardif G, Brazeau P, Martel-Pelletier J. Human osteoarthritic chondrocytes possess an in-creased number of insulin-like growth factor 1 binding sites but areunresponsive to its stimulation: possible role of IGF-1–bindingproteins. Arthritis Rheum 1994;37:253–63.
38. Martin JA, Ellerbroek SM, Buckwalter JA. Age-related decline inchondrocyte response to insulin-like growth factor-I: the role ofgrowth factor binding proteins. J Orthop Res 1997;15:491–8.
39. Lafeber FFJG, van der Kraan PM, Huber-Brning O, van den BergWB, Bijlsma JWJ. Osteoarthritic human cartilage is more sensitiveto transforming growth factor b than is normal cartilage. Br JRheumatol 1993;32:281–6.
40. Reddie AH. Role of morphogenetic proteins in skeletal tissueengineering and regulation. Nat Biotechnol 1998;16:247–52.
1164 SMITH ET AL





























