Embed Size (px)
Citation preview

Journal of PathologyJ Pathol 2014; 233: 183–195Published online in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/path.4347
ORIGINAL PAPER
Human adipocyte function is impacted by mechanical cuesV Pellegrinelli,1,2* J Heuvingh,5 O du Roure,5 C Rouault,1,2,3,4 A Devulder,5 C Klein,1,2,3 M Lacasa,2,6 E Clément,5D Lacasa1,2,3,4 and K Clément1,2,3,4
1 INSERM, UMR S 1166, Nutriomics Team, Paris, France2 Sorbonne Universités, UPMC University of Paris 06, UMR S 1166, ICAN, Paris, France3 Université Paris Descartes, UMR S 872, Paris, France4 Institute of Cardiometabolism and Nutrition, ICAN, AP-HP, Pitié-Salpêtrière Hospital, Paris, France5 PMMH UMR 7636 CNRS/ESPCI/UPMC, Paris, France6 INSERM, UMR S 1138 Team 4, Paris, France
*Correspondence to: V Pellegrinelli, Wellcome Trust-MRC Institute of Metabolic Science, Metabolic Research Laboratories, University of Cambridge,United Kingdom. E-mail: [email protected]
AbstractFibrosis is a hallmark of human white adipose tissue (WAT) during obesity-induced chronic inflammation.The functional impact of increased interstitial fibrosis (peri-adipocyte fibrosis) on adjacent adipocytes remainsunknown. Here we developed a novel in vitro 3D culture system in which human adipocytes and decellularizedmaterial of adipose tissue (dMAT) from obese subjects are embedded in a peptide hydrogel. When cultured withdMAT, adipocytes showed decreased lipolysis and adipokine secretion and increased expression/production ofcytokines (IL-6, G-CSF) and fibrotic mediators (LOXL2 and the matricellular proteins THSB2 and CTGF). Moreover,some alterations including lipolytic activity and fibro-inflammation also occurred when the adipocyte/hydrogelculture was mechanically compressed. Notably, CTGF expression levels correlated with the amount of peri-adipocytefibrosis in WAT from obese individuals. Moreover, dMAT-dependent CTGF promoter activity, which depends on�1-integrin/cytoskeleton pathways, was enhanced in the presence of YAP, a mechanosensitive co-activator of TEADtranscription factors. Mutation of TEAD binding sites abolished the dMAT-induced promoter activity. In conclusion,fibrosis may negatively affect human adipocyte function via mechanosensitive molecules, in part stimulated bycell deformation.Copyright © 2014 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Keywords: 3D culture; fibrosis; adipocytes; human obesity
Received 25 September 2013; Revised 21 February 2014; Accepted 27 February 2014
No conflicts of interest were declared.
Introduction
The extracellular matrix (ECM), which plays a pivotalrole in cell differentiation, migration and adhesion, iscomposed of structural proteins such as collagens andadhesion proteins such as fibronectin and proteoglycans.Excess deposition of insoluble ECM proteins, includ-ing interstitial collagens, characterizes fibrosis, a finalpathological feature of many chronic inflammatory dis-eases. Sustained production of fibrogenic cytokines andactivation of fibrogenic cells, such as myofibroblasts,leads to tissue remodelling that destroys the architec-ture and functions of different organs (liver, lung andkidney) [1]. Recent clinical and animal model stud-ies have shown the importance of the ECM in whiteadipose tissue (WAT) [2]. WAT is characterized byunique adipocyte plasticity, illustrated by its ability torapidly expand during obesity progression and to dimin-ish in size to ensure proper systemic energy homeostasis.WhileWAT growth is associated with ECM remodelling
in normal physiological conditions, an imbalance in col-lagen synthesis and degradation may result in excesscollagen deposition and subsequent fibrosis accumula-tion in obesity [2,3]. We reported previously that humanobese WAT shows altered expression of genes thatencode ECM components, proteases and inflammatorymediators [4,5]. We and others further showed that theseproteins are associated with increased peri-adipocytefibrosis in human obese WAT [6,7]. Increased type VIcollagen deposits in subcutaneous WAT (SAT) of obesesubjects are associated with insulin resistance [7,8].Notably, in mouse models, genetic ablation of collagensor metalloproteases profoundly affects adipocyte sizeand has metabolic consequences. For example, ob/obmice lacking collagen VI develop inflammation andadipocyte hypertrophy in the absence of fibrotic deposits[9]. Despite severe obesity, such mice are protected frommetabolic complications.Moreover, adipocyte hypertro-phy can be limited by genetic ablation of MT1-MMP,which plays a major role in collagen I degradation.MT1-MMP mutant mice demonstrate increased fibrosis
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

184 V Pellegrinelli et al
around smaller adipocytes and develop severe metabolicdisorders [10].These studies highlight the effects of fibrillar net-
works on adipocyte biology and suggest that mechanicalforces may contribute to the development/maintenanceof fibrosis. Indeed, ECM initiates both biochemicaland mechanical signals that are converted into biologi-cal responses, a process termed ’mechanotransduction’.Experimental evidence suggests that mechanotransduc-tion in WAT could be a crucial factor in the regulationof lipid storage and influence preadipocyte differenti-ation. However, it is unknown whether mature humanadipocytes (ie unilocular adipocytes) are mechanosensi-tive or mechanoresponsive. In particular, the effects offibrosis on biological function of adipocytes in obesesubjects have not been determined.Using a novel experimental model consisting of a
three-dimensional (3D) culture of mature adipocytesand decellularized material of adipose tissue (dMAT)embedded in a peptide hydrogel, we investigatedwhether dMAT affected the biological function ofadipocytes. We applied mechanical forces to thehydrogel to determine whether induced alterations inadipocyte function were similar to those induced by cul-ture with dMAT. Finally, we investigated the role of themechanosensitive transcriptional complex YAP/TEADin the regulation of connective tissue growth factor(CTGF) expression. This multifunctional matricellularprotein, over-expressed in fibrotic tissues and organssuch as lung, liver and kidney [11–14], is implicatedin the induction of several collagen proteins, such ascollagens I and III [15].
Materials and methods
Isolation of human adipocytes from SATand preparation of 3D adipocyte gel culturesSAT biopsies from the abdominal region were obtainedfrom 37 non-obese young women undergoing plasticsurgery and from 12 obese subjects undergoing bariatricsurgery. The clinical characteristics of these subjectsare described in the supplementary material. Each sam-ple was digested with collagenase and filtered throughgauze tissue. Floating adipocytes were washed threetimes in 10% sucrose, and non-digested material fromadipose tissue (MAT) was collected from the gauze tis-sue. Tissue pieces were decellularized to obtain dMAT(see supplementary material, Supplementary methods).Collagen fibre integrity was verified using tomography,immunofluorescence and second-harmonic generation(SHG) microscopy (see supplementary material, FigureS1A, B). The dMAT preparation was repleted withfibronectin (20 μg/ml) [16]. Importantly, fibronectin,which is a main ECM adhesion protein, is abundant inobese WAT and in fibrotic deposits (see supplementarymaterial, Figure S1C) [17]. Prior to 3D culture, dMATwas incorporated into the hydrogel, sonicated for 30 min
to achieve a homogeneous distribution, and the hydro-gel was cut thinly to ensure homogeneous cultures. Thepeptidic hydrogel was then diluted with 20% sucrosesolution. Adipocytes (1 × 104/100 μl gel) were embed-ded into the hydrogel alone (control condition) or in thepresence of dMAT (0.05 g/ml adipocyte/hydrogel mix-ture). Samples were placed into wells of a 96-well platecontaining 200 μl Dulbecco’s modified Eagle’s medium(DMEM)/F12 plus 1% albumin and 1% antibiotics, sup-plemented with insulin (50 nM). The culture mediumwas changed daily, and the spent medium was storedat −80∘C until experimental measurements were made.Additional protocol details can be found in the interna-tional patent (PCT/IB2011/052241 [18]) and in FigureS2A (see supplementary material).
Mechanical deformation of the 3D culture systemMechanical force was used to deform hydrogels. Briefly,we crafted a plastic culture lid in which small columnshad been inserted (column diameter = 4 mm; columnlength = 15 mm). The modified lid was placed on topof a multiwell culture plate, such that each column wasinserted into a single well and compressed/deformedthe hydrogel in a controlled manner. Deformation wasdefined as the ΔH:H ratio, where H is the height ofthe gel before compression and (H – ΔH) is the heightof the deformed gel. Downward pressure from the topthus produced compression of the gel vertically in therange 0–50% (maximal deformation with the gel heightbeing halved). The range was achieved by spacers in thefour empty wells at the corners of the plate, two at 0%compression height and two at 50% compression height(see supplementary material, Figure S5A).
Monitoring adipocyte metabolismLipolysis assay
The adipocyte/hydrogel was incubated in Krebs–Ringerbicarbonate buffer supplemented with 3% bovine serumalbumin (BSA) for 4 h at 37∘C. Lipolytic activity wasevaluated by measuring the release of glycerol andnon-esterified fatty acids (NEFAs) after stimulation withisoproterenol 1 μM, forskolin 10 μM and DbcAMP0.5mM.
Adipocyte secretory functions
Inflammatory factors produced by adipocytes after 3days of culture with dMAT were measured using aMultiplex immunoassay. Colorimetric ELISA kits wereused to determine the adipokine concentrations in themedium in different experimental conditions, accordingto the manufacturer’s instructions.
Phosphorylated kinase array
We determined the phosphorylation profile of 43kinases and their protein substrates in adipocytes that
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 185
were exposed to dMAT for 30 min or that were sub-jected to mechanical deformation according to themanufacturer’s instructions (Human Phospho-KinaseAntibody Array, ARY003B, R&D Systems). Briefly,1 ml cell lysate was incubated with the membranesovernight at 4∘C. The membranes were then incu-bated with the detection antibody cocktail for 2 hat room temperature. After washing, the membraneswere incubated with streptavidin–HRP for 30 min atroom temperature. Specific signals were detected byincubation with ECL detection solution (GE Health-care), followed by immediate exposure to X-rayfilms. Signals obtained after a 5 min exposure wereanalysed.
Plasmids, transfection experiments and luciferaseassayThe construction of a 1000 bp sequence correspond-ing to the controlling sequence of the human CTGFgene is presented in Supplementary methods (seesupplementary material). The pGL3–CTGF–lucplasmid was generated by subcloning the 1000 bpCTGF promoter fragment into the pGL3 basic vec-tor upstream of the luciferase gene. CTGF–TB3 orTB2/TB3 mutants were generated by site-directedmutagenesis. HEK cells were transiently transfectedwith either the pGL3–CTGF–luc construct alone(WT, mutated with TB3, or both TB2/TB3 sites) orco-transfected with the pCMV–Flag–YAP–4SAconstruct (Addgene, Cambridge, MA, USA) [19].Transfected cells were cultured in the 3D culture systemwith or without dMAT, as described in Supplementarymethods.
Statistical analysisExperiments were repeated at least four times, usingadipocytes from different lean subjects. Statisticalanalyses were performed using GraphPad Software(San Diego, CA, USA). The values were expressed asmean ± SEM of (n) independent experiments. Com-parisons between two conditions (adipocyte cultureversus adipocyte culture/mechanical compression) wereanalysed using theWilcoxon non-parametric paired test.Comparisons among more than two groups were per-formed using one-way analysis of variance (ANOVA),followed by a post hoc test. Differences were consideredsignificant when p < 0.05.
Supplementary experimental proceduresSupplementary methods (see supplementary mate-rial) contains the clinical characteristics of lean andobese subjects, sources of all antibodies and chem-ical products, adipocyte flattening analysis, proteinand gene expression analysis with the list of primersequences used for real-time PCR (Table S1), quan-tification of total/peri-cellular fibrosis in obese SAT,OCT/SHG/confocal imaging analysis, Multiplex assayresults and experimental models.
Results
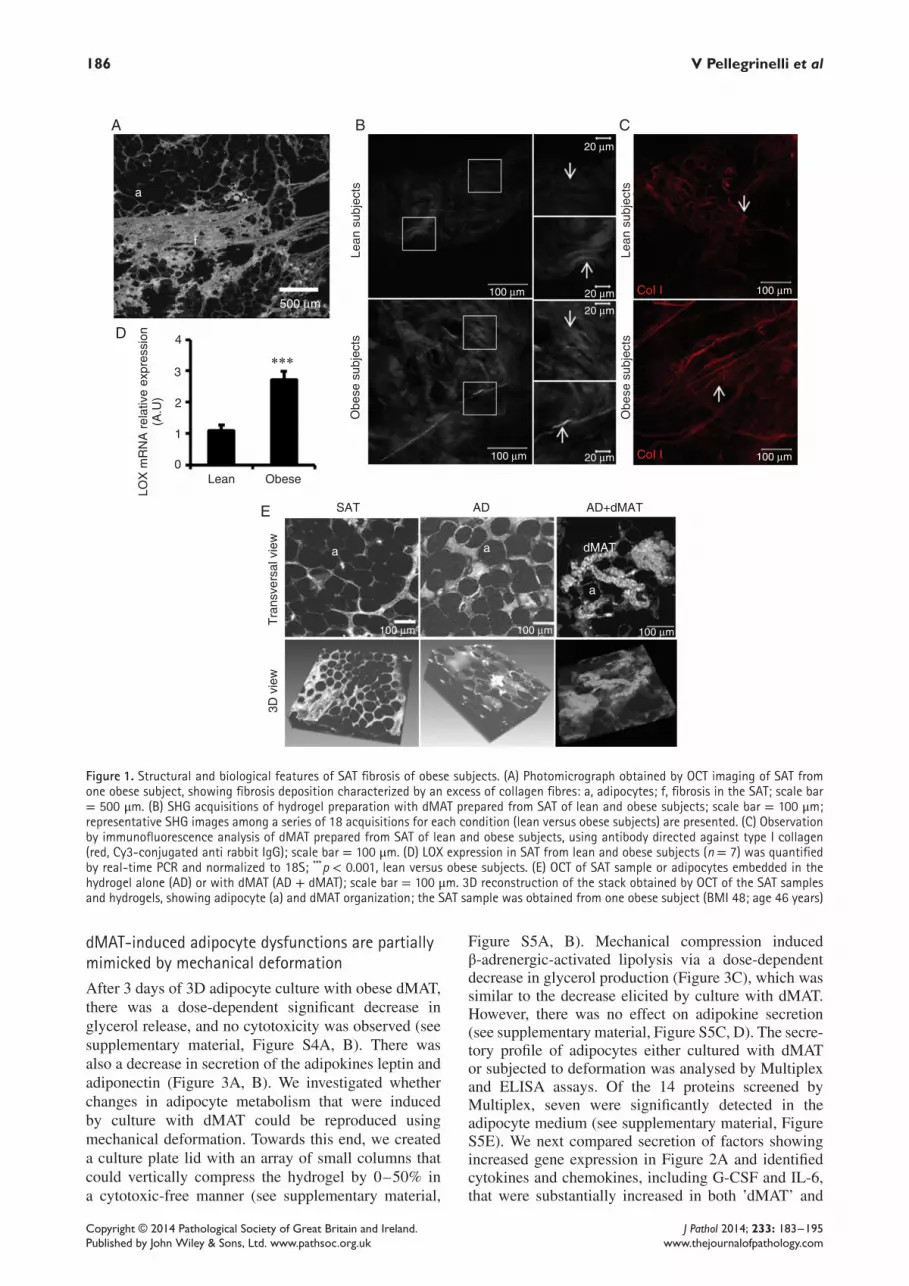
Fibrotic depots of SAT from obese subjects showcollagen accumulation and structural remodellingTomographic imaging (optical coherence tomography,OCT) was used to examine in situ SAT samples fromobese subjects, demonstrating thick bands of collagenfibres throughout the tissue (Figure 1A). Adipocyteswere surrounded by collagen fibres. Insoluble fibresextracted from obese SAT (obSAT dMAT)were predom-inantly type I, III and VI collagen proteins; in contrast,non-digested obSAT showed greater collagen diversity(see supplementary material, Figure S1C, D). Moreover,second harmonic generation (SHG) imaging, whichreveals the detailed structural organization of collagenfibrils, showed an enhanced signal in obSAT dMATcompared to lean SAT dMAT (Figure 1B). Specifically,collagen fibre organization in obSAT dMAT appearedmore compact and linearized compared to lean SATdMAT (Figure 1B, C). The expression of lysyl oxydase(LOX), a major collagen-crosslinking enzyme, was sub-stantially increased in obese SAT (Figure 1D). Theseresults suggest greater crosslinking of collagen fibres inobSAT dMAT versus lean SAT dMAT and that this mayimpact adipocyte function.
3D culture of adipocytes with dMAT from obesesubjectsAdipocyte culture is technically challenging, due tobuoyancy and fragility stemming from their relativelylarge diameter and unilocular structure. Cell viabilityis reduced after 24–48 h in two-dimensional (2D) cul-ture [20]. Self-assembling peptides provide a nanofibrescaffold which is suitable for long-term 3D hepatocyteculture [21]. In these 3D cultures, which more closelymimic a physiological context, we observed that leanSAT adipocytes were protected from cell lysis and thatthe responses to insulin, lipolytic activity and adipokinesecretion were maintained for 1 week (see supplemen-tary material, Figure S2). Subsequently, adipocytes werecultured either alone in the hydrogel or in the pres-ence of obese dMAT (Figure 1E). Interestingly, dMATseemed to affect adipocyte morphology (see supplemen-tary material, Figure S3A), with a net tendency towardslarger values (ie flatter cells) in the presence of dMAT;the mean flattening value was 22.7 ± 0.9% (n = 254)versus 18.1 ± 0.7% (n = 348) without dMAT (see sup-plementary material, Figure S3B), corresponding to adeformation of 10%. In parallel, 3D OCT imaging ofadipocytes embedded in a hydrogel subjected to uniax-ial (vertical) compression (Figure 2A, B) revealed thatdeformation from 0% to 50% induced a mean adipocytevertical deformation of 2.3 ± 0.5% to 9.2 ± 1.0% (n =16) (Figure 2C). Thus, flattening deformation inducedby dMAT was comparable to uniaxial deformation ofthe adipocytes by 10%, which corresponded to a 50%compression of the hydrogel sample.
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com
186 V Pellegrinelli et al
Lean
2
LOX
mR
NA
rel
ativ
e ex
pres
sion
(A.U
)
Obe
se s
ubje
cts
Obe
se s
ubje
cts
Lean
sub
ject
s
Lean
sub
ject
s
3
4
A B C
D
E
500 µm100 µm 20 µm
20 µm
20 µm
20 µm100 µm
100 µm 100 µm 100 µm
100 µm
100 µm
a
f
a a
a
dMAT
1
0Obese
SAT
CoI I
CoI I
Tra
nsve
rsal
vie
w3D
vie
w
AD AD+dMAT
∗∗∗
Figure 1. Structural and biological features of SAT fibrosis of obese subjects. (A) Photomicrograph obtained by OCT imaging of SAT fromone obese subject, showing fibrosis deposition characterized by an excess of collagen fibres: a, adipocytes; f, fibrosis in the SAT; scale bar= 500 μm. (B) SHG acquisitions of hydrogel preparation with dMAT prepared from SAT of lean and obese subjects; scale bar = 100 μm;representative SHG images among a series of 18 acquisitions for each condition (lean versus obese subjects) are presented. (C) Observationby immunofluorescence analysis of dMAT prepared from SAT of lean and obese subjects, using antibody directed against type I collagen(red, Cy3-conjugated anti rabbit IgG); scale bar = 100 μm. (D) LOX expression in SAT from lean and obese subjects (n = 7) was quantifiedby real-time PCR and normalized to 18S; ***p < 0.001, lean versus obese subjects. (E) OCT of SAT sample or adipocytes embedded in thehydrogel alone (AD) or with dMAT (AD + dMAT); scale bar = 100 μm. 3D reconstruction of the stack obtained by OCT of the SAT samplesand hydrogels, showing adipocyte (a) and dMAT organization; the SAT sample was obtained from one obese subject (BMI 48; age 46 years)
dMAT-induced adipocyte dysfunctions are partiallymimicked by mechanical deformationAfter 3 days of 3D adipocyte culture with obese dMAT,there was a dose-dependent significant decrease inglycerol release, and no cytotoxicity was observed (seesupplementary material, Figure S4A, B). There wasalso a decrease in secretion of the adipokines leptin andadiponectin (Figure 3A, B). We investigated whetherchanges in adipocyte metabolism that were inducedby culture with dMAT could be reproduced usingmechanical deformation. Towards this end, we createda culture plate lid with an array of small columns thatcould vertically compress the hydrogel by 0–50% ina cytotoxic-free manner (see supplementary material,
Figure S5A, B). Mechanical compression inducedβ-adrenergic-activated lipolysis via a dose-dependentdecrease in glycerol production (Figure 3C), which wassimilar to the decrease elicited by culture with dMAT.However, there was no effect on adipokine secretion(see supplementary material, Figure S5C, D). The secre-tory profile of adipocytes either cultured with dMATor subjected to deformation was analysed by Multiplexand ELISA assays. Of the 14 proteins screened byMultiplex, seven were significantly detected in theadipocyte medium (see supplementary material, FigureS5E). We next compared secretion of factors showingincreased gene expression in Figure 2A and identifiedcytokines and chemokines, including G-CSF and IL-6,that were substantially increased in both ’dMAT’ and
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 187
100 µm
49%
Hydrogel deformation (%)
20 30 40 501000
2
4
6
8
Adi
pocy
te d
efor
mat
ion
(%) 10
C
BA
42%
28%
21%
0%
Figure 2. Mechanical deformation and adipocyte morphology. (A) OCT transverse views of a group of adipocytes embedded in hydrogeland submitted to vertical compression (the applied deformation is indicated in white). These transverse views are the top 200 μm of a 7mm-thick sample; scale bar = 100 μm. (B) Superposition of the reconstructed 3D shape of one adipocyte at zero (green) and maximal (red)value of applied deformation. Compression in the vertical direction leads to expansion in the horizontal directions (flattening); black barsindicate 50 μm. (C) Deformations of individual adipocytes as a function of applied deformation to the whole sample. Deformations aremeasured in the direction of the compressive force. The analysis was conducted on 16 different adipocytes
’deformation’ conditions (Figure 3D). Interestingly,this inflammatory response did not increase whenadipocytes were both cultured with dMAT and sub-jected to mechanical compression (see supplementarymaterial, Figure S5F). Finally, although WAT fromlean individuals exhibits some interstitial fibrosis, thedMAT from lean WAT did not alter adipocyte functionor increase inflammation (see supplementary material,Figure S6).
dMAT and mechanical compression exert theireffects via β1-integrin-dependent pathwayin adipocytesWe next determined whether β1-integrin signallingmediated dMAT-induced mechanotransduction inadipocytes. In the presence of β1-integrin blockingantibody, dMAT-induced IL-6 secretion was abol-ished, while adipokine secretion remained reduced(Figure 4A; see also supplementary material, FigureS7). External forces transmitted by integrins can acti-vate distinct signalling pathways that involve focaladhesion proteins, caveolin-1 and their downstreamkinase targets (eg FAK, ROCK and ERK, respectively),as well as actin-dependent pathways that induce Srckinase activation [22]. Short-term dMAT exposure byadipocytes induced phosphorylation of several pro-teins, such as caveolin-1 tyrosine 14 phosphorylation(Figure 4B). It also induced the phosphorylation ofseveral kinases (eg FAK, ERK 1/2, AMPKα2 andGSK-3α/β) and their downstream targets (eg STAT
proteins and HSP27; Figure 4C; see also supplementarymaterial, Figure S8A). Furthermore, culture of differ-entiated preadipocytes with dMAT caused an increasein actin stress fibres detected by phalloidin staining,suggesting increased isometric cytoskeletal tension (seesupplementary material, Figure S8B).
dMAT-exposed or mechanically-compressedadipocytes increase inflammatory and fibrotic geneexpressionMechanical compression modifies gene expression inseveral cellular models [23,24]. We thus examinedchanges in gene expression in adipocytes that werecultured with obese dMAT or that were compressed.Several genes involved in adipocyte metabolism,inflammation, ECM remodelling and the endoplasmicreticulum (ER) stress response have been described asmechanosensitive genes in previous studies (marked #in Figure S8C; see supplementary material) [25–27].Whereas expression of metabolic proteins such asGLUT-4, FABP-4 and CD36 was not modified in eitherof these conditions, expression of ER stress markergenes ATF4, HSPA5 and C/EBPζ/CHOP was increasedin dMAT-cultured adipocytes or those subjected tocompression. There was also a significantly increasedexpression of inflammatory (IL-6 and CCL5) andECM remodelling [LOXL2, CTGF, thrombospondin 2(THSB2) and COL1A1] genes in adipocytes culturedwith dMAT. Mechanical deformation induced similarresponses, except that the expression of LOXL2 and
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

188 V Pellegrinelli et al
1.2
Basal
Iso
AD
AD+dMAT
AD
AD+dMAT
AD+DEF
Basal
R2 = 0.7453
R2 = 0.6919
p = 0.029
p = 0.051
Iso
G-CSF IL-820
Hydrogel deformation (%)
30 40 501000
200
400
600
IL-6 CCL5 VEGF
1
0.8
0.6
Sec
retio
n(f
old
over
con
trol
, AD
)
Sec
retio
n(f
old
over
con
trol
, AD
)
∗∗
∗∗
∗
∗
∗∗
∗∗
∗∗∗
∗
#
∗∗
0.4
0.2
0
0
2
4
6
8
10
12
LeptinAD
2
Gly
cero
l rel
ease
(fol
d ov
er c
ontr
ol, A
D b
asal
)
Gly
cero
l rel
ease
(µM
)
3
4
5
6A B
C D
1
0AD+dMAT Adiponectin
Figure 3. Metabolic and secretory functions of adipocytes from lean SAT cultured with obese dMAT or submitted to mechanicaldeformations. (A) Lipolytic activity was evaluated by the glycerol release from either adipocytes cultured alone (AD) or with dMAT (AD +dMAT) after 3 days in the 3D culture model. Glycerol release was measured in the basal (white bars) or β-adrenergic-stimulated conditions;isoproterenol, Iso (1 μM; black bars) for 4 h. The results are presented as the fold difference between the basal condition in AD and otherconditions; all data are presented as mean ± SEM of six independent experiments; *p < 0.05, **p < 0.01, basal versus Iso conditions in ADand AD + dMAT; #p < 0.05, AD versus AD + dMAT in Iso conditions. (B) Adipokine secretion was measured using ELISA. Comparison ofleptin and adiponectin secretions from either adipocytes alone (AD, white bars) or adipocytes cultured with dMAT (AD + dMAT, black bars)for 3 days in the 3D culture model. The results are presented as the fold variation of adipokine secretions in AD + dMAT compared to AD;data are mean ± SEM of six independent experiments; **p < 0.01 AD versus AD + dMAT. (C) Effects of mechanical constraint on lipolyticactivity of adipocytes evaluated for each deformation from 0% to 50%. Glycerol and NEFA release were measured under basal (white line)or stimulated conditions, isoproterenol (Iso, 1 μM; black line) for 4 h in the 3D culture model. The glycerol and NEFA release negativelycorrelated with the mechanical deformation for stimulated conditions. Data are presented as mean ± SEM of five independent experiments.(D) Significant changes in the secretion of inflammatory molecules by adipocytes in the 3D culture with dMAT (AD + dMAT, black bars)compared to either adipocytes cultured alone (control AD, white bars) or adipocytes subjected to 50% mechanical deformation (AD +DEF, grey bars). The results are presented as the fold differences over the control; data are presented as mean ± SEM of six independentexperiments; *p< 0.05, **p< 0.01, AD versus AD + dMAT or AD + DEF. The SAT of lean subjects (23.05 ± 0.38 kg/m2) was used to prepareadipocytes and SAT of obese subjects (mean BMI = 45.2 ± 1.5 kg/m2) to prepare dMAT
COL1A1 did not increase (Figure 5A). Moreover, LOX(which was increased in WAT during obesity) wasmodified neither by dMAT nor compression expo-sure (see supplementary material, Figure S8D). Wenext explored the biochemical role of dMAT-inducedadipocyte response by studying the effect of solubleforms of ECM components on adipocytes cultured in3D. Incorporation of soluble collagen I and fibronectininto the hydrogel had no effect on lipolytic activityor ER stress, but decreased adipokine secretion andinduced kinase phosphorylation (HSP27, STAT5 andERK1/2) and the expression of inflammatory/profibroticgenes (IL-6, CCL5, CTGF, LOXL2; see supplementarymaterial, Figure S9A–D). However, these soluble pro-teins could not completely reproduce the inflammatoryresponse induced by dMAT at the secretion level (seesupplementary material, Figure S9E, F). Fibronectinalone had no effect but acted synergistically to increase
the inflammatory effects of dMAT (see supplementarymaterial, Figure S10).
CTGF is a potential mediator of WAT fibrosisand adipocyte mechanotransductionCTGF, LOXL2 and THSB2 mRNA was increased inadipocytes cultured with obese dMAT compared to con-trol (Figure 5). The clinical relevance of these results issupported by the observation that CTGF, LOXL2 andTHSB2 mRNA levels correlate with total fibrosis inthe SAT of obese subjects. However, only the expres-sion of CTGF mRNA correlated with peri-adipocytefibrosis (Table 1). It is unknown whether CTGF, ahallmark of several fibrotic diseases, plays a role inWAT fibrosis. Re-examination of data obtained froma pan-genomic cDNA microarray performed previ-ously [4] showed that CTGF is up-regulated in obese
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 189
50
STAT2 (Y689)HSP27 (S78/S82)STAT5a (Y694)AMPKα2 (T172)STAT5b (Y699)Src (Y419)Chk-2 (T68)AKT (T308)CREB (S133)MSK 1/2 (S376/360)P70 S6K (T389)Hck (Y411)Yes (Y426)
Lyn (Y397)STAT6 (Y641)P53 (S46)AKT (S473)ERK 1/2 (T202/Y204/T185/Y187)STAT5a/b (Y694/Y699)FAK (Y397)TOR (S2448)c-jun (S63)PRAS-40 (T246)EGFR (Y1086)P53 (S15)RSK 1/2/3 (S380/S386/S377)STAT3 (Y705)P70 S6K (T421/S424)HSP60
GSK-3α /β (S21/S9)
ADAD
A C
B
AD+dMATIL
-6 s
ecre
tion
(fol
d ov
er c
ontr
ol, A
D)
ns
IgG10
1
2
3
ab β1-int
∗∗#
AD+dMAT
Kinase phosphorylation level (A.U)
AD+DEF
AD
pY14 Cav-1
Cav-1
AD+dMAT AD+DEF
0
Figure 4. β1-integrin- and kinase-dependent pathways activated in fibrotic or constraint adipocytes. (A) ELISA analysis of IL-6 secretion byadipocytes in the 3D culture with dMAT (AD+ dMAT, black bars) compared to adipocytes cultured alone (control AD, white bars). Adipocyteswere treated with IgG1 or β1-integrin neutralizing antibody (ab β1-int). Data are presented as mean± SEM of five independent experiments;#p< 0.05, AD versus AD+ dMAT; **p< 0.01, IgG1 versus ab int-β1 in AD+ dMAT; ns, not significant. (B) Cell lysates prepared from adipocytescultured for 30 min in the 3D hydrogel alone (control, AD) or exposed to dMAT (AD + dMAT) or 50% mechanical deformation (AD + DEF)were immunoblotted to detect caveolin-1 phosporylated on tyrosine 14 (Y14 Cav-1). The figure shows one representative immunoblot amongfour independent experiments; *p< 0.05, AD versus AD + dMAT or AD + DEF. (C) Cell lysates prepared from four independent experimentsof adipocytes cultured for 30 min in the 3D hydrogel alone (control, AD) or exposed to dMAT (AD + dMAT) or 50% mechanical deformation(AD + DEF) were pooled and analysed for the phosphorylation of different kinases, using the Human Phospho-Kinase Antibody Array kit.Heat maps with graded shades of red represent the quantifications of membrane array performed by ImageJ software and presented inFigure S4B (see supplementary material)
WAT and down-regulated after weight loss, empha-sizing the potential role of CTGF in obesity. Theseresults were confirmed by PCR analysis, whereasLOXL-2 expression showed a non-significant increasedexpression in WAT from obese subjects (see supple-mentary material, Figure S11). Moreover, when humanpreadipocytes, the major pro-fibrotic cells in WATduring obesity [16], were treated with human recombi-nant CTGF, they increased expression of COL1, COL3and fibronectin (Figure 5B, C). In contrast, CTGFdoes not directly impact adipocyte function (data notshown).
dMAT-dependent regulation of CTGF promoteractivity through β1-integrin/cytoskeletal signallingBecause CTGF is both pro-fibrotic and a fibrosis targetgene, we investigated its transcriptional regulationby dMAT in our 3D culture systems. We generateda CTGF–luciferase construct (pGL3–CTGF–luc)corresponding to the proximal sequence (−1000/0)and transiently transfected it into HEK cells cul-tured with dMAT in the 3D system in the presenceof a blocking β1-integrin antibody or pharma-cological inhibitors. dMAT-induced luciferase
Table 1. Spearman correlations between fibrosis (total andpericellular) and the expression of fibrosis-target genes (CTGF ,LOXL2 and THSB2)
R p
Total fibrosis, CTGF 0.554 0.009Pericellular fibrosis 0.655 0.038Total fibrosis, LOXL2 0.483 0.031Pericellular fibrosis 0.412 NSTotal fibrosis, THSB2 0.497 0.026Pericellular fibrosis 0.207 NS
NS, not significant.Both total and pericellular fibrosis were quantified by using red picrosiriusstaining of SAT biopsies from obese subjects (obese individual cohort from [6];BMI ± SEM, 48.2 ± 0.8).
activity was reduced in the presence of a block-ing β1-integrin antibody (Figure 6A), LatrunculinB, cytochalasin B, LY 294002 and Sarcatinib,but not with Y27632; these agents inhibit actinpolymerization, microtubule polymerization, PI3-kinase, Src kinase and ROCK inhibitor, respectively(Figure 6B). Taken together, these results suggest thatβ1-integrin-dependent/cytoskeleton-dependent path-ways play critical roles in the mechanical regulation ofthe CTGF promoter.
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

190 V Pellegrinelli et al
AD
0
3
2
1
0
5
4
1
2
3
0COL1A1 COL1A2 COL3A1 FN
Control
Control
∗
∗
∗
∗∗∗
∗ ∗ ∗∗
∗∗ ∗
∗∗∗
∗∗∗
∗
∗
∗ ∗ ∗
∗∗ ∗∗
Con
trol
mR
NA
rel
ativ
e ex
pres
sion
(fol
d ov
er c
ontr
ol, A
D)
mR
NA
rel
ativ
e ex
pres
sion
(fol
d ov
er c
ontr
ol)
Imm
unof
luor
esce
nt p
rote
in(f
old
over
con
trol
)
CT
GF
(250
ng/
mL)
CTGF(250 ng/ml)
CTGF(250 ng/ml)
ATF4 HSPA5 IL-6 CCL5
Collagen I
20 µm 20 µm
20 µm20 µm
Fibronectin
Collagen I Fibronectin
DDR2 CTGF THSB2 LOXL2 COL1A1C/EBPζ
1
2
3
4
5
6
7A
B C
AD+dMATAD+DEF
Figure 5. Gene expression profile in adipocytes exposed to obese dMAT and mechanical deformation. (A) Significant differences in the geneexpression in the 3D culture of adipocytes with dMAT (AD + dMAT, black bars) compared to either adipocytes cultured alone (control AD,white bars) or adipocytes subjected to 50% mechanical deformation (AD + DEF; grey bars). The results are presented as the fold differencesover control adipocytes. Data are presented as mean ± SEM of six independent experiments; *p< 0.05, **p< 0.01, AD versus AD + dMAT orAD + DEF. (B, C) CTGF effects on ECM components of human preadipocytes. Human preadipocytes were treated by recombinant CTGF (250ng/ml) for 3 days. Cells were then collected for RNA extraction or fixed for immunofluorescence analysis. (B) Gene expression of COL1A1,COL1A2, COL3A1 and FN in control (white bars) and preadipocytes treated by CTGF (250 ng/ml; black bars). (C) Immunofluorescenceanalysis of control and CTGF-treated preadipocytes using antibodies directed against collagen type I, (red, Cy3-conjugated anti rabbit IgG)and fibronectin (green, Cy2-conjugated anti mouse IgG; scale bar = 20 μm. Graphs represent quantification of the fluorescent signal. Dataare presented as mean ± SEM of six or seven independent experiments; *p< 0.05, **p< 0.01, control versus CTGF. The SAT of obese subjects(mean BMI = 45.2 ± 1.5 kg/m2) was used to prepare dMAT
dMAT induces CTGF promoter activity through aYAP–TEAD-dependent pathwayYes-associated protein (YAP) is induced in responseto mechanical cues such as cytoskeleton tension [28].Moreover, YAP, which co-activates the TEA-DNAbinding domain family member (TEAD), up-regulatesthe promoter activity and expression of CTGF [29].In mature adipocytes, YAP and TEAD1 expressionincreased in the presence of dMAT in the 3D cul-ture system (Figure 7A). Further, YAP and CTGFexpression were positively correlated in these cells(Figure 7B). The phosphorylated form of YAP wasdecreased in adipocytes cultured with dMAT, suggest-ing that it may have undergone nuclear translocation;notably, this is where YAP exerts its co-regulator activ-ity (Figure 7C). To assess the importance of YAP indMAT-induced activity of the CTGF promoter, weused a vector that expressed a constitutively activeform of YAP, YAP-4SA [19]. Our data showed thatYAP-4SA strongly induced CTGF promoter activitycompared to non-co-transfected cells and maintainedthe dMAT response. Given this effect of YAP-4SA, we
next validated the importance of TEADs. Two majorbinding sites for TEADs that regulate CTGF expression,TB2 and TB3, have been identified [29]. We generatedmutations in these sites and transiently co-transfectedHEK cells with the different pGL3–CTGF–luc con-structs (ie wild-type or mutated in the TB2/TB3sites) and YAP-4SA. Importantly, both TB2 and TB3mutations were needed to completely suppress bothYAP-4SA- and dMAT-induced CTGF promoter activityin co-transfected cells (Figure 7D).
Discussion
In obesity, human WAT undergoes major remodellingwith excess insoluble collagen fibres [4,6]. An increasein connective fibres may exert mechanical forces onsurrounding adipocytes (peri-adipocyte fibrosis) andthereby contribute to the metabolic disorders asso-ciated with obesity [9]. However, fibrosis-inducedfactors that alter human WAT biology and the poten-tial mechanosensitive responses of adipocytes remainunknown. Our human in vitro cellular study, which was
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 191
HEKA B
ns
∗
∗ ∗∗∗
# #HEK+dMAT
Control
CT
GF
pro
mot
er a
ctiv
ity(f
old
over
con
trol
, HE
K)
CT
GF
pro
mot
er a
ctiv
ity(f
old
over
con
trol
, HE
K)
0
1
2
3
0
1
2
34
IgG ab β1-int
Contro
l
Latru
nculi
n B
Cytoca
lasin
B
LY29
4002
Sarca
tinib
Y2763
2
HEKHEK+dMAT
Figure 6. Roles of β1-integrin and cytoskeleton pathways in CTGF promoter activity. HEK cells were transiently transfected with thepGL3–CTGF proximal promoter–Luc construct and cultured alone (HEK, white bars) or in presence of dMAT (0.05 g/ml; HEK + dMAT,black bars) for 48 h. The luciferase activity was normalized against β-galactosidase activity for each well (three wells/condition). The resultsare presented as the fold difference over the control (HEK). (A) CTGF promoter activity in HEK cells treated with IgG1 (control, IgG 5 μg/ml)or β1-integrin neutralizing antibody (ab β1-int, 5 μg/ml). Data are presented as mean ± SEM of three independent experiments; #p< 0.05,HEK versus HEK + dMAT; *p < 0.05, control IgG1 versus ab int-β1 in HEK + dMAT. (B) CTGF promoter activity in HEK cells treated withinhibitors of actin (Latrunculin B, 20 μM) or microtubule polymerization (cytochalasin B, 20 μM), PI 3-kinase (LY 294002, 25 μM), Src kinase(sarcatinib, 100 nM) or Rock kinase (Y27632, 10 μM). Data are presented as mean ± SEM of five independent experiments; #p < 0.05, HEKversus HEK + dMAT; *p < 0.05, control versus inhibitors in HEK + dMAT. The SAT of obese subjects (mean BMI = 45.2 ± 1.5 kg/m2) wasused to prepare dMAT
based on the convergence of biophysics and biology,provides new insights regarding the link between fibro-sis and mature adipocyte dysfunction. Here we showedthat dMAT prepared from obese, but not lean WATaffected lipolysis and adipokine secretion by adipocytesand promoted fibro-inflammation. The alteration ofstimulated β-adrenergic lipolysis is a feature charac-terizing adipocytes from obese subjects [30]. Althoughadipocyte hypertrophy and associated inflammationcharacterized WAT from obese subjects, heterogene-ity of adipocyte size is also observed with smalleradipocytes in fibrotic regions (Figure 1). Removingcollagen VI in obese mice (COL6 KO Ob/ob mice)leads to larger adipocytes in association with metabolicimprovements [9]. Moreover, MT1-MMP KO micedisplay WAT fibrosis and associated lipodystrophyassociated with a defective leptin production [10]. Thus,we hypothesized that adipocytes present in fibroticdepots become less metabolically active because ofincreased tension from the rigid surrounding matrix inobese WAT. In the present study, both SHG imaging andLOX expression analysis suggested increased collagenfibre crosslinking in WAT from obese subjects, whichmight explain the altered metabolism of adipocytes.Pharmacological inhibition of LOX improves metabolicparameters and reduces inflammation in mice [31]. Ourfindings are in agreement with recent clinical resultsshowing associations between fibrosis and physicalmeasures of stiffness in human adipose tissue [32].
Here we observed increased expression of IL-6 andCCL5 and the ECM-associated proteins CTGF andLOXL-2 in adipocytes cultured with dMAT or sub-jected to deformation. These proteins, which are alsoproduced by other WAT cells, such as macrophages,preadipocytes and endothelial cells, are up-regulatedin response to mechanical stress in other cell types[25,27,33,34]. However, mechanical force alone didnot reproduce all of the dMAT-induced alterations inadipocytes, such as decreases in adipokine secretion.Thus, other mechanisms, such as biochemical cuesthat depend on the composition of the dMAT, may beinvolved. This hypothesis is supported by our findingsthat the incorporation of type I collagen and fibronectininto the adipocyte hydrogel culture induced expres-sion of pro-inflammatory/fibrotic genes and decreasedadipokine secretion. Overall. our findings propose thatadipocyte metabolism and secretory functions are influ-enced by both biochemical and mechanical cues (seesupplementary material, Table S2).We were particularly interested in how dMAT acts
on adipocytes leading to CTGF induction; while CTGFinhibits adipocyte differentiation [35], its role hasnever been linked to WAT fibrosis during obesity. Wefound a correlation between CTGF expression andperi-adipocyte fibrosis in human WAT. Furthermore,CTGF induced production of collagen and fibronectinin human preadipocytes, the main fibrosis-producingcells in obese WAT [16]. This suggests that CTGF isa pro-fibrotic factor contributing to the maintenance
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

192 V Pellegrinelli et al
0.2
0.15
0.1
YAP
mR
NA
rel
ativ
e ex
pres
sion
mR
NA
rel
ativ
e ex
pres
sion
(fol
d ov
er c
ontr
ol, A
D)
0.05
00 0.02
YAP
0
1
2
3
4
TEAD1
AD
1 0.25± 0.1∗∗
0.37± 0.11∗
pS127 YAP
Total YAP
AD+dMAT AD+DEF
CT
GF
pro
mot
er a
ctiv
ity(f
old
over
con
trol
, HE
K)
TEAD2 TEAD3 TEAD4
35282114
76
4
2
0Control YAP-4SA YAP-4SA TB3
mutationYAP-4SATB2/TB3mutations
CTGF mRNA relative expression0.04 0.06
HEK
ns
**
**
∗∗
∗
#
##
##
******
R2 = 0.653p = 0.0014AD
A B
C D
AD+dMAT
HEK+dMAT
Figure 7. Role of YAP–TEAD transcriptional complex in dMAT-induced regulation of CTGF. (A) Differences in the gene expression of YAPand TEADs in the 3D culture of adipocytes with dMAT (AD + dMAT, black bars) compared to adipocytes cultured alone (control AD, whitebars). The results are presented as the fold differences over control AD. Data are presented as mean ± SEM of six independent experiments;*p < 0.05, **p < 0.01, AD versus AD + dMAT. (B) Correlation analysis between the gene expressions of CTGF and YAP was performed in3D cultured adipocytes using the Spearman regression test. (C) Cell lysates prepared from adipocytes cultured for 24 h in the 3D settingalone (control, AD) or exposed to dMAT (AD + dMAT) or 50% mechanical deformation (AD + DEF) were immunoblotted to detect total and127serine phosphorylated (S127 YAP) form of YAP. The results are presented as the fold differences over the control (AD). The figure showsa representative immunoblot among four independent experiments; *p< 0.05, AD versus AD + DEF; **p< 0.01, AD versus AD + dMAT. TheSAT of lean subjects (mean BMI = 23.05 ± 0.38 kg/m2) was used to prepare adipocytes and SAT of obese subjects (mean BMI = 45.2 ±1.5 kg/m2) to prepare dMAT. (D) HEK cells were transiently transfected with pGL3–CTGF proximal promoter–Luc construct, mutated or not(control) for TB3 or TB2/TB3 binding sites and cultured alone (HEK, white bars) or in the presence of dMAT, 0.05 g/ml (HEK + dMAT, blackbars) for 48 h. HEK cells were co-transfected with empty vector or constitutively active YAP form (YAP-4SA). The luciferase activity wasnormalized against β-galactosidase activity for each well (three wells/condition). The results are presented as the fold difference over thecontrol (HEK cells without dMAT). Data are presented as mean ± SEM of five independent experiments; ##p< 0.01 HEK versus HEK + dMAT;**p< 0.01, YAP-4SA versus YAP-4SA TB3 mutation or YAP-4SA TB2/TB3 mutations in HEK + dMAT; ***p< 0.001, YAP-4SA versus control orYAP-4SA TB2/TB3 mutations in HEK + dMAT. The SAT of obese subjects (mean BMI = 45.2 ± 1.5 kg/m2) was used to prepare dMAT
of fibro-inflammation in obese WAT and to adipocytedysfunction via mechanotransduction pathways.Adipocytes cultured in the presence of dMAT or sub-
jected to compressive forces were deformed (flattened)compared to control cells. Although compressive forceswere always in the same (vertical) direction, fibrosis caninduce compression in various directions. We showedthat both situations led to a flattening of the adipocytes.Mechanotransduction pathways linked to external ten-sion may thus be involved in the detection of suchdeformations. These pathways involve mechanorecep-tors [36] such as integrins and adaptor molecules [22].Mechanotransduction pathways may also involve thedisassembly of caveolae in adipocytes [37].Here we propose that β1-integrin pathways involv-
ing both the actin cytoskeleton and multiple kinases
could mediate dMAT-induced inflammation and CTGFexpression. Our results are in line with recent observa-tions that, during tumour progression, increased ECMstiffness enhances integrin, focal adhesion and PI3Ksignalling [38]. In cancer-associated fibroblasts (CAFs),increased ECM stiffness affects mechanotransductionby activation of Src-family kinases downstream ofcytoskeleton tension, which activate YAP [39]. To ourknowledge, our study is the first to report the involve-ment of the YAP/TEAD transcriptional complex indMAT-induced CTGF expression. This involvementsuggests that there may be a similar mechanism inmature adipocytes exposed to dMAT.In conclusion, this study provides new information
about the pathological effects of WAT fibrosis on humanadipocyte function (Figure 8). Specifically, our results
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 193
Collagen fibers
Adipokines
Biochemical andbiomechanical actions
Fibrosis
CTGFInflammation
Lipolysis
FAK FAKCav-1
βα βα
Src
ERKPI3K
CTGF
YAP
TEAD
Integrins
Actin cytoskeleton
Microtubules
Figure 8. Integrative scheme presenting hypothetic action of fibro-sis through biochemical and biomechanical cues on humanadipocyte functions and transcriptional regulation of CTGF expres-sion. During obesity, the hypertrophied adipose tissue undergoesexcessive deposition of collagen fibres around adipocytes, calledperi-adipocyte fibrosis. Through both biochemical and biomechani-cal actions, fibrosis could provoke metabolic and secretory dysfunc-tions of adipocytes with increased production of pro-inflammatoryand pro-fibrotic molecules, among them the matricellular proteinCTGF regulated by the mechanosensitive YAP–TEAD pathway. Thisphenomenon could then perpetuate a vicious cycle with the main-tenance of fibro-inflammation in adipose tissue of obese subjects
suggest that mechanosensitive molecules (YAP/TEADand CTGF) may constitute novel therapeutic targetsfor the prevention of WAT fibro-inflammation duringobesity.
Acknowledgements
We are grateful to Maude Legall and Nicolas Vente-clef for helpful discussions. We thank NutriOmic forcontributing to the biological sample collection andacknowledge the help of Christine Poitou-Bernert, Nico-las Veyrie and Jean-Luc Bouillot. We thank EugenieDalimier for performing OCT imaging (LL-Tech, Paris,France) and Perrine Frère for performing SHG anal-ysis (Imaging Facilities, Institut du Cerveau et de laMoelle épinière, Pitié Salpétrière, Paris, France). Themanuscript was edited by San Francisco Edit. Thisstudy was funded by the University Pierre et MarieCurie (Emergence Programme 2010), by the NationalAgency of Research (ANR Adipofib), by the Fon-dation pour la Recherche Medicale (FRM team) andby the European Community Seventh Framework Pro-gramme (adipokines as a drug to combat the adverseeffects of excess adipose tissue (ADAPT) project; GrantNo. 201100). The Nutriomic team was supported by aFrench government grant, which was managed by the
ANR (programme ’Investments for the Future’, GrantNo. ANR-10-IAHU]. We thank the Assistance Publiquedes Hôpitaux de Paris [APHP; Programmes of Clini-cal Investigation (PHRC) No. 0702) for its support ofhuman data collection.
Author contributions
VP andDL designed, performed and analyzed the resultsof the biological experiments and wrote the manuscript;JH, OdR, AD and EC conceptualized, performed andanalysed the biophysical experiments and were involvedin writing the manuscript; CR performed the Multiplexexperiments; CK performed the confocal analysis; MLdesigned and constructed the CTGF promoter/luciferasevectors; andKC analysed the biological experiments andwrote the manuscript with the input of all of the otherauthors.
Abbreviations
3D, three-dimensional; BMI, body mass index; BSA,bovine serum albumin; CCL2/MCP-1, chemokine(C–C) motif ligand 2/monocyte chemoattractant pro-tein 1; CCL5/RANTES, chemokine (C–C) motif ligand5/regulated on activation, normal T cell expressed andsecreted; C/EBP, CAAT/enhancer binding protein;COL, collagen; CTGF, connective tissue growth factor;dMAT, decellularized material of adipose tissue; ECM,extracellular matrix; ER, endoplasmic reticulum; FN,fibronectin; G-CSF, granulocyte colony-stimulatingfactor; IL, interleukin; LOX, lysyl oxidase; LOXL2,lysyl oxidase-like 2; MMP, metalloprotease; NEFA,non-esterified fatty acids; NF-κB, nuclear factor-κB;OCT, optical coherence tomography; Pref-1/Dlk1,preadipocyte factor 1/δ-like homologue 1; SAT, sub-cutaneous adipose tissue; SHG, second harmonicgeneration; SVF, stroma-vascular fraction; TEAD,TEAD domain family; THSB2, thrombospondin 2;VAT, visceral adipose tissue; WAT, white adiposetissue; YAP, yes-associated protein.
References1. Zeisberg M, Kalluri R. Cellular mechanisms of tissue fibrosis.
1. Common and organ-specific mechanisms associated with tissuefibrosis. Am J Physiol Cell Physiol 2013; 304: C216–225.
2. Divoux A, Clément K. Architecture and the extracellular matrix: thestill unappreciated components of the adipose tissue. Obes Rev Off JInt Assoc Study Obes 2011; 12: e494–503.
3. Sun K, Kusminski CM, Scherer PE. Adipose tissue remodeling andobesity. J Clin Invest 2011; 121: 2094–2101.
4. Henegar C, Tordjman J, Achard V, et al. Adipose tissue transcrip-tomic signature highlights the pathological relevance of extracellularmatrix in human obesity. Genome Biol 2008; 9: R14.
5. Cancello R, Henegar C, Viguerie N, et al. Reduction of macrophageinfiltration and chemoattractant gene expression changes in whiteadipose tissue of morbidly obese subjects after surgery-inducedweight loss. Diabetes 2005; 54: 2277–2286.
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

194 V Pellegrinelli et al
6. Divoux A, Tordjman J, Lacasa D, et al. Fibrosis in human adiposetissue: composition, distribution, and link with lipid metabolism andfat mass loss. Diabetes 2010; 59: 2817–2825.
7. Spencer M, Yao-Borengasser A, Unal R, et al. Adipose tissuemacrophages in insulin-resistant subjects are associated with col-lagen VI and fibrosis and demonstrate alternative activation. Am J
Physiol Endocrinol Metab 2010; 299: E1016–1027.8. Pasarica M, Gowronska-Kozak B, Burk D, et al. Adipose tissue col-
lagen VI in obesity. J Clin Endocrinol Metab 2009; 94: 5155–5162.9. Khan T, Muise ES, Iyengar P, et al. Metabolic dysregulation and
adipose tissue fibrosis: role of collagen VI. Mol Cell Biol 2009; 29:1575–1591.
10. Chun TH, Hotary KB, Sabeh F, et al. A pericellular collagenasedirects the three-dimensional development of white adipose tissue.Cell 2006; 125: 577–591.
11. Phanish MK, Winn SK, Dockrell MEC. Connective tissue growthfactor (CTGF, CCN2) – a marker, mediator and therapeutic target forrenal fibrosis. Nephron Exp Nephrol 2010; 114: e83–92.
12. Gressner OA, Gressner AM. Connective tissue growth factor: afibrogenic master switch in fibrotic liver diseases. Liver Int Off J IntAssoc Study Liver 2008; 28: 1065–1079.
13. Lipson KE, Wong C, Teng Y, et al. CTGF is a central mediatorof tissue remodeling and fibrosis and its inhibition can reverse theprocess of fibrosis. Fibrogen Tissue Repair 2012; 5(suppl 1): S24.
14. Ponticos M, Holmes AM, Shi-wen X, et al. Pivotal role of connec-tive tissue growth factor in lung fibrosis: MAPK-dependent tran-scriptional activation of type I collagen. Arthritis Rheum 2009; 60:2142–2155.
15. De Winter P, Leoni P, Abraham D. Connective tissue growth factor:structure–function relationships of amosaic, multifunctional protein.Growth Factors Chur 2008; 26: 80–91.
16. Keophiphath M, Achard V, Henegar C, et al. Macrophage-secretedfactors promote a profibrotic phenotype in human preadipocytes.MolEndocrinol 2009; 23: 11–24.
17. Taleb S, Cancello R, et al. Cathepsin s promotes human preadipocytedifferentiation: possible involvement of fibronectin degradation.Endocrinology 2006; 147: 4950–4959.
18. Patent No.WO/2011/148310A1: http: //www.sumobrain.com/patents/wipo/Method-culturing-adipocytes/WO2011148310A1.pdf; cited29 December 2012.
19. Zhao B, Kim J, Ye X, et al. Both TEAD-binding and WW domainsare required for the growth stimulation and oncogenic transformationactivity of yes-associated protein. Cancer Res 2009; 69: 1089–1098.
20. Gesta S, Lolmède K, Daviaud D, et al. Culture of human adiposetissue explants leads to profound alteration of adipocyte gene expres-sion. Horm Metab Res 2003; 35: 158–163.
21. Wang S, Nagrath D, Chen PC, et al. Three-dimensional primaryhepatocyte culture in synthetic self-assembling peptide hydrogel.Tissue Eng A 2008; 14: 227–236.
22. Ingber DE. Cellular mechanotransduction: putting all the piecestogether again. FASEB J Off Publ Fed Am Soc Exp Biol 2006; 20:811–827.
23. Wang N, Tytell JD, Ingber DE. Mechanotransduction at a distance:mechanically coupling the extracellular matrix with the nucleus. NatRev Mol Cell Biol 2009; 10: 75–82.
24. Mammoto A, Mammoto T, Ingber DE. Mechanosensitive mecha-nisms in transcriptional regulation. J Cell Sci 2012; 125: 3061–3073.
25. Klein S, de Fougerolles AR, Blaikie P, et al. α5β1 integrin acti-vates an NF-κB-dependent program of gene expression important forangiogenesis and inflammation.Mol Cell Biol 2002; 22: 5912–5922.
26. Chaqour B, Yang R, Sha Q. Mechanical stretch modulates the pro-moter activity of the profibrotic factor CCN2 through increasedactin polymerization and NF-κB activation. J Biol Chem 2006; 281:20608–20622.
27. Yang R, Amir J, Liu H, Chaqour B. Mechanical strain activates aprogram of genes functionally involved in paracrine signaling ofangiogenesis. Physiol Genom 2008; 36: 1–14.
28. Halder G, Dupont S, Piccolo S. Transduction of mechanical andcytoskeletal cues by YAP and TAZ. Nat Rev Mol Cell Biol 2012; 13:591–600.
29. Zhao B, Ye X, Yu J, et al. TEAD mediates YAP-dependent geneinduction and growth control. Genes Dev 2008; 22: 1962–1971.
30. Horowitz JF, Klein S.Whole body and abdominal lipolytic sensitivityto epinephrine is suppressed in upper body obese women. Am JPhysiol Endocrinol Metab 2000; 278: E1144–1152.
31. Halberg N, Khan T, Trujillo ME, et al. Hypoxia-inducible factor 1αinduces fibrosis and insulin resistance in white adipose tissue. MolCell Biol 2009 ; 29: 4467–4483.
32. Abdennour M, Reggio S, Le Naour G, et al. Association of adiposetissue and liver fibrosis with tissue stiffness in morbid obesity: linkswith diabetes and BMI loss after gastric bypass. J Clin EndocrinolMetab 2014; jc20133253.
33. Okada M, Matsumori A, Ono K, et al. Cyclic stretch upregulatesproduction of interleukin-8 and monocyte chemotactic and activat-ing factor/monocyte chemoattractant protein-1 in human endothelialcells. Arterioscler Thromb Vasc Biol 1998; 18: 894–901.
34. Peterson JM, Pizza FX. Cytokines derived from cultured skeletalmuscle cells after mechanical strain promote neutrophil chemotaxisin vitro. J Appl Physiol (Bethesda, MD, 1985) 2009; 106: 130–137.
35. Tan JTM, McLennan SV, Song WW, et al. Connective tissue growthfactor inhibits adipocyte differentiation. Am J Physiol Cell Physiol
2008; 295: C740–751.36. Larsen M, Artym VV, et al. The matrix reorganized: extracellular
matrix remodeling and integrin signaling. Curr Opin Cell Biol 2006;18: 463–471.
37. Parton RG, Simons K. The multiple faces of caveolae. Nat Rev MolCell Biol 2007; 8: 185–194.
38. Levental KR, Yu H, Kass L, et al. Matrix crosslinking forcestumor progression by enhancing integrin signaling. Cell 2009; 139:891–906.
39. Calvo F, Ege N, Grande-Garcia A, et al. Mechanotransduction andYAP-dependent matrix remodelling is required for the generation andmaintenance of cancer-associated fibroblasts.Nat Cell Biol 2013; 15:637–646.
SUPPLEMENTARY MATERIAL ON THE INTERNETThe following supplementary material may be found in the online version of this article:
Supplementary methods
Figure S1. From WAT extracellular matrix to dMAT
Figure S2. 3D culture of human unilocular adipocytes, tissue microstructure and maintenance of viable mature adipocytes
Figure S3. Analysis of adipocyte flattening in culture with obese dMAT
Figure S4. Lipolytic activity of adipocytes cultured with obese dMAT in 3D hydrogel
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

Human adipocytes as mechanosensitive cells 195
Figure S5. Metabolic and secretory functions of adipocytes submitted to mechanical deformation and cultured with obese dMAT in 3D hydrogel
Figure S6. Metabolic and secretory functions of adipocytes cultured with dMAT from SAT of lean subjects in the 3D setting
Figure S7. Role of β1-integrin in mediating alterations of adipocytes induced by obese dMAT
Figure S8. Effect of obese dMAT on kinase phosphorylation profile, actin cytoskeleton remodelling and gene expression
Figure S9. Biochemical effects of soluble type I collagen/fibronectin on metabolic and secretory functions of human adipocytes
Figure S10. Effects of fibronectin alone or with dMAT exposure on human adipocyte functions
Figure S11. CTGF and LOXL2 expression in WAT during obesity
Table S1. List of primer sequences used for real-time PCR
Table S2. Recapitulative observations on the alterations of metabolic activities and secretory functions of human adipocytes
Copyright © 2014 Pathological Society of Great Britain and Ireland. J Pathol 2014; 233: 183–195Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com





























