Embed Size (px)
Citation preview

Human cystic fibrosis airway epithelia have reducedCl− conductance but not increased Na+ conductanceOmar A. Itania,1, Jeng-Haur Chena,b, Philip H. Karpa,b, Sarah Ernsta, Shaf Keshavjeec, Kalpaj Parekhd, Julia Klesney-Taita,Joseph Zabnera,2, and Michael J. Welsha,b,e,2
Departments of aInternal Medicine, dCardiothoracic Surgery, and eMolecular Physiology and Biophysics and bThe Howard Hughes Medical Institute, The Roy J.and Lucille A. Carver College of Medicine, University of Iowa, Iowa City, IA 52242; and cToronto Lung Transplant Program, University of Toronto, Toronto, ON,Canada M5G 2C4
Contributed by Michael J. Welsh, May 2, 2011 (sent for review February 1, 2011)
Loss of cystic fibrosis transmembrane conductance regulator (CFTR)anion channel function causes cystic fibrosis (CF) lung disease. CFTRis expressed in airway epithelia, but how CF alters electrolytetransport across airway epithelia has remained uncertain. Recentstudies of a porcine model showed that in vivo, excised, andcultured CFTR−/− and CFTRΔF508/ΔF508 airway epithelia lacked anionconductance, and they did not hyperabsorb Na+. Therefore, weasked whether Cl− and Na+ conductances were altered in humanCF airway epithelia. We studied differentiated primary cultures oftracheal/bronchial epithelia and found that transepithelial conduc-tance (Gt) under basal conditions and the cAMP-stimulated in-crease in Gt were markedly attenuated in CF epithelia comparedwith non-CF epithelia. These data reflect loss of the CFTR anionconductance. In CF and non-CF epithelia, the Na+ channel inhibitoramiloride produced similar reductions in Gt and Na+ absorption,indicating that Na+ conductance in CF epithelia did not exceed thatin non-CF epithelia. Consistent with previous reports, adding ami-loride caused greater reductions in transepithelial voltage andshort-circuit current in CF epithelia than in non-CF epithelia; thesechanges are attributed to loss of a Cl− conductance. These resultsindicate that Na+ conductance was not increased in these culturedCF tracheal/bronchial epithelia and point to loss of anion transportas key to airway epithelial dysfunction in CF.
chloride secretion | epithelial Na+ channels
Electrolyte transport by airway epithelia and submucosalglands controls the quantity and composition of airway sur-
face liquid. The cystic fibrosis transmembrane conductance reg-ulator (CFTR) anion channel is localized in the apical membraneof airway epithelia, and its loss in cystic fibrosis (CF) is thought toimpair pulmonary host defense (1–7).However, how loss of CFTRfunction changes electrolyte transport across airway epithelia hasbeen uncertain.The first evidence of defective airway epithelial ion transport
was the observation that the voltage (Vt) across nasal epitheliawas more electrically negative in persons who have CF than innon-CF controls (8). Applying amiloride, an inhibitor of epi-thelial Na+ channels (ENaC), caused a greater reduction innasal Vt (ΔVtamil) in CF epithelia. Subsequent studies revealedthat CF epithelia were Cl− impermeable compared with non-CFepithelia (9, 10). Studies of excised nasal epithelia in Ussingchambers yielded varying results that, as the authors noted,might be attributed to epithelial location (CF polyps vs. non-CFturbinates), tissue injury, medications, small sample sizes, chronicinfection, inflammation, and exclusion bias (11).To circumvent some of these limitations and to facilitate
studies of airway epithelial ion transport, many investigatorsestablished techniques for culturing human airway epithelial cellsat the air–liquid interface so that they differentiate and can bestudied in vitro (12). Loss of anion conductance has been ap-parent in nearly all studies of differentiated CF airway epithelialcultures (9, 10, 13, 14). In addition, some studies showed anincreased short-circuit current (Isc) and voltage under basalconditions (basal Isc and basal Vt, respectively) and increasedreductions with amiloride application (ΔIscamil and ΔVtamil, re-
spectively) (7). The increased basal Vt, ΔVtamil, basal Isc, andΔIscamil were interpreted to indicate that Na+ channel activity(i.e., Na+ conductance) was increased in CF epithelia (7, 15).Additional studies of recombinant CFTR and ENaC in MDCKcells and fibroblasts reported that CFTR inhibited ENaC chan-nels (16), although other studies in oocytes suggested that CFTRdid not inhibit ENaC (17). These reports plus additional studiesled to the hypothesis that CFTR inhibits ENaC and that loss ofCFTR increases ENaC Na+ conductance, resulting in Na+
hyperabsorption and reducing periciliary liquid depth to causeCF lung disease (for a review, see ref. 15).To understand CF pathogenesis better, we recently developed
pigs with either a targeted disruption of the CFTR gene or a ΔF508mutation (18–20).CFTR−/−andCFTRΔF508/ΔF508pigs spontaneouslydevelop lung disease that closely resembles that in personswho haveCF (20, 21). At birth, porcine CF airway epithelia are not inflamed,but they already exhibit defective host defense against bacteria.Moreover, newborn CFTR−/− and CFTRΔF508/ΔF508 pigs exhibit in-creased nasal Vt and ΔVtamil like that seen in persons who have CF.However, they do not havemore Na+ absorption than controls, andtheCF-induced elevations inbasalVt, basal Isc,ΔVtamil, andΔIscamilare caused by the lack of apical Cl− conductance, not by increasedNa+ conductance (20, 22).These observations led us to ask how lack of CFTR changes
electrolyte transport in primary cultures of differentiated humanairway epithelia. Our main goal was to determine if human CFepithelia have decreased Cl− conductance, increased Na+ con-ductance, or a combination of the two. Any of these alternativesmight explain the changes in Vt and Isc observed in CF.We began by examining the electrophysiological properties of
primary cultures of differentiated airway epithelia and measuredtransepithelial conductance (Gt), Isc, and Vt. We first focusedon Gt (the inverse of transepithelial resistance) for two mainreasons. First, Gt measurements are related directly to the per-meability of ion channels, and our two main questions are aboutCl− and Na+ channels. Second, compared with Isc and Vtmeasurements in airway epithelia, Gt is much less affected bymembrane voltages, ion concentrations, and ion flow in thephysiologic range. Gt is the sum of the cellular (Gc) and para-cellular (Gp) conductances, and Gc is determined by apical andbasolateral conductances. If CF epithelia have only a reducedCl− conductance, then Gt under basal conditions (basal Gt)should be decreased, and the converse should be true if increased
Author contributions: O.A.I., J.-H.C., P.H.K., J.Z., and M.J.W. designed research; O.A.I., J.-H.C.,P.H.K., S.E., J.Z., and M.J.W. performed research; S.K., K.P., and J.K.-T. contributed newreagents/analytic tools; O.A.I., J.-H.C., P.H.K., S.E., J.Z., and M.J.W. analyzed data; andO.A.I., J.-H.C., J.Z., and M.J.W. wrote the paper.
Conflict of interest statement: M.J.W. is a co-founder of Exemplar Genetics, which islicensing materials and technology related to this work.
Freely available online through the PNAS open access option.1Present address: Department of Biology, University of Balamand, El-Koura, Lebanon.2To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1106695108/-/DCSupplemental.
10260–10265 | PNAS | June 21, 2011 | vol. 108 | no. 25 www.pnas.org/cgi/doi/10.1073/pnas.1106695108
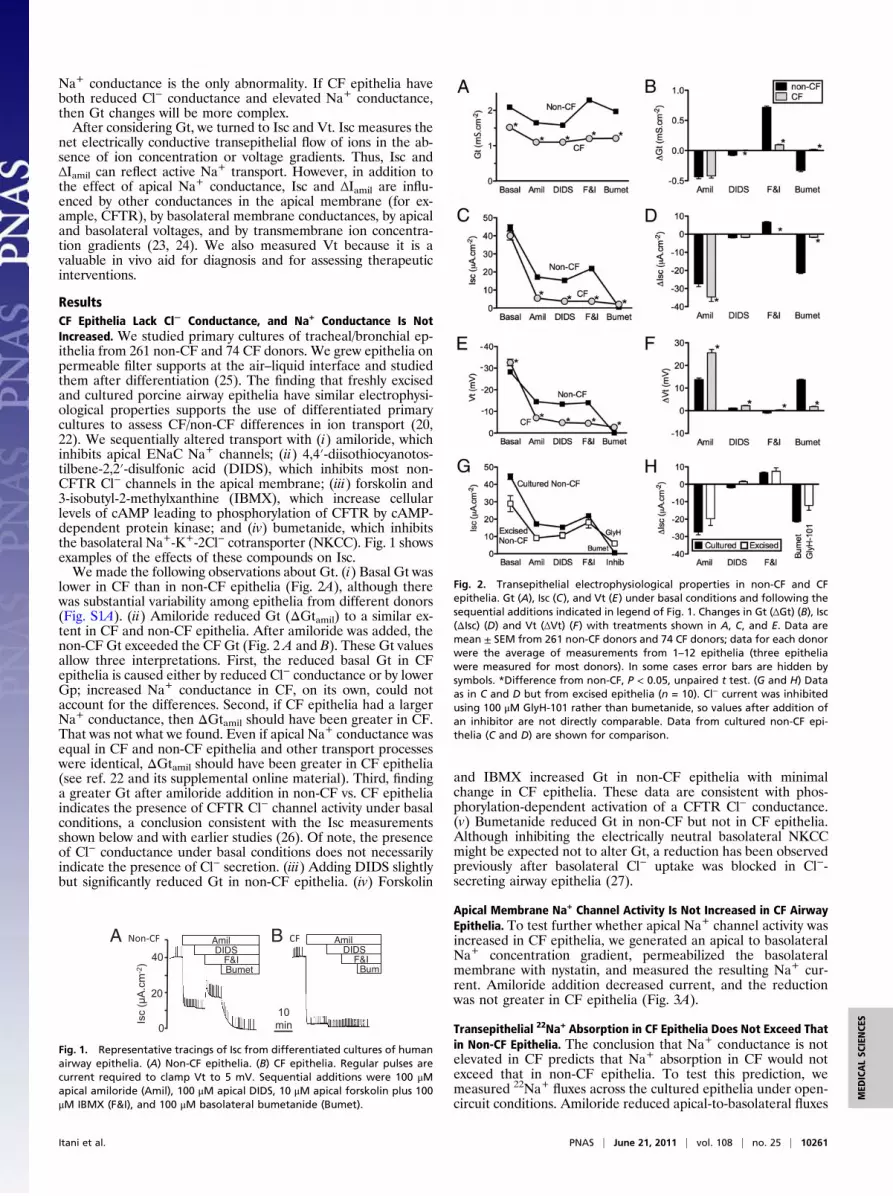
Na+ conductance is the only abnormality. If CF epithelia haveboth reduced Cl− conductance and elevated Na+ conductance,then Gt changes will be more complex.After considering Gt, we turned to Isc and Vt. Isc measures the
net electrically conductive transepithelial flow of ions in the ab-sence of ion concentration or voltage gradients. Thus, Isc andΔIamil can reflect active Na+ transport. However, in addition tothe effect of apical Na+ conductance, Isc and ΔIamil are influ-enced by other conductances in the apical membrane (for ex-ample, CFTR), by basolateral membrane conductances, by apicaland basolateral voltages, and by transmembrane ion concentra-tion gradients (23, 24). We also measured Vt because it is avaluable in vivo aid for diagnosis and for assessing therapeuticinterventions.
ResultsCF Epithelia Lack Cl− Conductance, and Na+ Conductance Is NotIncreased. We studied primary cultures of tracheal/bronchial ep-ithelia from 261 non-CF and 74 CF donors. We grew epithelia onpermeable filter supports at the air–liquid interface and studiedthem after differentiation (25). The finding that freshly excisedand cultured porcine airway epithelia have similar electrophysi-ological properties supports the use of differentiated primarycultures to assess CF/non-CF differences in ion transport (20,22). We sequentially altered transport with (i) amiloride, whichinhibits apical ENaC Na+ channels; (ii) 4,4′-diisothiocyanotos-tilbene-2,2′-disulfonic acid (DIDS), which inhibits most non-CFTR Cl− channels in the apical membrane; (iii) forskolin and3-isobutyl-2-methylxanthine (IBMX), which increase cellularlevels of cAMP leading to phosphorylation of CFTR by cAMP-dependent protein kinase; and (iv) bumetanide, which inhibitsthe basolateral Na+-K+-2Cl− cotransporter (NKCC). Fig. 1 showsexamples of the effects of these compounds on Isc.We made the following observations about Gt. (i) Basal Gt was
lower in CF than in non-CF epithelia (Fig. 2A), although therewas substantial variability among epithelia from different donors(Fig. S1A). (ii) Amiloride reduced Gt (ΔGtamil) to a similar ex-tent in CF and non-CF epithelia. After amiloride was added, thenon-CF Gt exceeded the CF Gt (Fig. 2 A and B). These Gt valuesallow three interpretations. First, the reduced basal Gt in CFepithelia is caused either by reduced Cl− conductance or by lowerGp; increased Na+ conductance in CF, on its own, could notaccount for the differences. Second, if CF epithelia had a largerNa+ conductance, then ΔGtamil should have been greater in CF.That was not what we found. Even if apical Na+ conductance wasequal in CF and non-CF epithelia and other transport processeswere identical, ΔGtamil should have been greater in CF epithelia(see ref. 22 and its supplemental online material). Third, findinga greater Gt after amiloride addition in non-CF vs. CF epitheliaindicates the presence of CFTR Cl− channel activity under basalconditions, a conclusion consistent with the Isc measurementsshown below and with earlier studies (26). Of note, the presenceof Cl− conductance under basal conditions does not necessarilyindicate the presence of Cl− secretion. (iii) Adding DIDS slightlybut significantly reduced Gt in non-CF epithelia. (iv) Forskolin
and IBMX increased Gt in non-CF epithelia with minimalchange in CF epithelia. These data are consistent with phos-phorylation-dependent activation of a CFTR Cl− conductance.(v) Bumetanide reduced Gt in non-CF but not in CF epithelia.Although inhibiting the electrically neutral basolateral NKCCmight be expected not to alter Gt, a reduction has been observedpreviously after basolateral Cl− uptake was blocked in Cl−-secreting airway epithelia (27).
Apical Membrane Na+ Channel Activity Is Not Increased in CF AirwayEpithelia. To test further whether apical Na+ channel activity wasincreased in CF epithelia, we generated an apical to basolateralNa+ concentration gradient, permeabilized the basolateralmembrane with nystatin, and measured the resulting Na+ cur-rent. Amiloride addition decreased current, and the reductionwas not greater in CF epithelia (Fig. 3A).
Transepithelial 22Na+ Absorption in CF Epithelia Does Not Exceed Thatin Non-CF Epithelia. The conclusion that Na+ conductance is notelevated in CF predicts that Na+ absorption in CF would notexceed that in non-CF epithelia. To test this prediction, wemeasured 22Na+ fluxes across the cultured epithelia under open-circuit conditions. Amiloride reduced apical-to-basolateral fluxes
Fig. 1. Representative tracings of Isc from differentiated cultures of humanairway epithelia. (A) Non-CF epithelia. (B) CF epithelia. Regular pulses arecurrent required to clamp Vt to 5 mV. Sequential additions were 100 μMapical amiloride (Amil), 100 μM apical DIDS, 10 μM apical forskolin plus 100μM IBMX (F&I), and 100 μM basolateral bumetanide (Bumet).
Fig. 2. Transepithelial electrophysiological properties in non-CF and CFepithelia. Gt (A), Isc (C), and Vt (E) under basal conditions and following thesequential additions indicated in legend of Fig. 1. Changes in Gt (ΔGt) (B), Isc(ΔIsc) (D) and Vt (ΔVt) (F) with treatments shown in A, C, and E. Data aremean ± SEM from 261 non-CF donors and 74 CF donors; data for each donorwere the average of measurements from 1–12 epithelia (three epitheliawere measured for most donors). In some cases error bars are hidden bysymbols. *Difference from non-CF, P < 0.05, unpaired t test. (G and H) Dataas in C and D but from excised epithelia (n = 10). Cl− current was inhibitedusing 100 μM GlyH-101 rather than bumetanide, so values after addition ofan inhibitor are not directly comparable. Data from cultured non-CF epi-thelia (C and D) are shown for comparison.
Itani et al. PNAS | June 21, 2011 | vol. 108 | no. 25 | 10261
MED
ICALSC
IENCE
S

in both non-CF and CF epithelia (Fig. 3B). The net rate of Na+absorption did not differ between non-CF and CF epithelia.The Gt data, measurements of Na+ current when the baso-
lateral membrane was permeabilized, and 22Na+ flux results in-dicate that Na+ conductance was not increased in CF epithelia.
Amiloride Causes Greater Isc and Vt Changes in CF Epithelia. Wecompared Isc in CF and non-CF epithelia. (i) Basal Isc did notdiffer statistically between the two groups (Fig. 2C and Fig. S1B).(ii) Amiloride reduced Isc more in CF than in non-CF epithelia(Fig. 2 C and D). After amiloride was added, Isc was greater innon-CF than in CF epithelia because of the presence of Cl−secretion. This result is consistent with constitutive CFTR ac-tivity under basal conditions. These data also support the con-clusion that the greater ΔIscamil in CF was caused by the lack ofa Cl− conductance. (iii) DIDS had minimal effects in both CFand non-CF epithelia. (iv) Increasing cellular levels of cAMPincreased Isc, and inhibiting basolateral Cl− entry with bumeta-nide reduced Isc in non-CF but not in CF epithelia. Theseobservations have been made often and indicate CFTR-dependent Cl− secretion.We also measured Vt. (i) Basal Vt was more electrically
negative in CF epithelia than in non-CF epithelia (Fig. 2E andFig. S1B). (ii) Amiloride reduced Vt almost twice as much in CFepithelia as in non-CF epithelia (Fig. 2 E and F). The lesserΔVtamil in non-CF epithelia was caused largely by an amiloride-insensitive Vt from non-ENaC–mediated active ion transport.(iii) Adding forskolin and IBMX had minimal effects on Vt innon-CF epithelia. (iv) After bumetanide was added to inhibit Cl−
entry into cells, and thereby transepithelial Cl− secretion, non-CF Vt fell to values close to those in CF epithelia, i.e., near zero.
Excised Non-CF Epithelia Show Properties Similar to Those in CulturedEpithelia. We also studied bronchial epithelia excised from 10nonsmoking persons without chronic obstructive lung disease;some of the lungs had been removed for transplantation butwere not used for reasons that are unknown to us. Edge damageand potential injury during mounting can increase Gt markedly;the increase would be similar to an increase in Gp (28). Thus,
this preparation reduces Vt. Edge damage also reduces the areaof functioning epithelia in the Ussing chamber and therebyreduces Isc, but the effect on Isc is much less than on Gt. Wefound that values of basal Isc, ΔIscamil, and cAMP-induced ΔIscwere reasonably close to those of cultured epithelia (Fig. 2G andH). The directions of Gt and Vt responses to the various inter-ventions were consistent with (although much smaller than)those in cultured epithelia (Fig. S2). We did not evaluate excisedCF bronchial epithelia, because the tissue we obtained usuallywas not suitable for study because of advanced disease.
Paradoxical Finding That ΔVtamil and ΔIscamil but Not Na+ Conductance
Are Increased in CF. It may seem paradoxical that these CF epi-thelia do not have increased Na+ conductance but have a greaterbasal Vt, ΔVtamil, and ΔIscamil than non-CF epithelia. Considerfive points. (i) The non-CF epithelia have substantial CFTR Cl−conductance under basal conditions, consistent with earlierstudies (26). That Cl− conductance is missing in CF. (ii) Addinga Cl− channel to a Na+-conductive apical membrane introducesanother electromotive force generated by the apical Cl− con-centration gradient, shunts voltage generated by Na+ channels,and alters the effect on the voltage of current generated at thebasolateral membrane (23, 24). The resulting changes in apical(and basolateral) voltage then alter Vt and Isc. Subtracting a Cl−conductance has the opposite effects. (iii) Equivalent electricalcircuit models show that loss of apical Cl− conductance can in-crease ΔVtamil and ΔIscamil even when apical Na+ conductance isheld constant (23, 24). Although the responsible processes arecomplex, one of several factors is that amiloride hyperpolarizesthe apical membrane, increasing the driving force for apical Cl−exit and thereby minimizing ΔIscamil compared with CF epithelia,which lack CFTR anion channels. (iv) In porcine nasal epithelia,CF/non-CF differences in basal Vt, Isc, ΔVtamil, and ΔIscamil arecaused by the lack of CFTR Cl− conductance (22). These dif-ferences are absent or less striking in porcine tracheal/bronchialepithelia where CFTR Cl− conductance is minimal underbasal conditions. (v) The paradoxical influence on electrophysi-ological properties of the absence of Cl− conductance is readilyapparent in the sweat gland duct. In CF ducts, Na+ absorption isdecreased by ∼80%, but absolute Vt can be increased 10-fold(29). Decreased Cl− conductance, not increased Na+ conduc-tance, causes the increased Vt.To test further the conclusion that CF/non-CF differences in
basal Vt, ΔVtamil, and ΔIscamil were caused by lack of CFTR Cl−conductance rather than by increased Na+ conductance, we didseveral additional studies.
Reducing Cl− Conductance Increases ΔIscamil and ΔVtamil in Non-CFEpithelia. We predicted that reducing the CFTR Cl− conduc-tance of non-CF epithelia would minimize CF/non-CF differ-ences in Vt, ΔVtamil, and ΔIscamil. To test this prediction, weadded forskolin and IBMX to culture medium for 16–24 h andthen withdrew it when epithelia were mounted in Ussingchambers. Withdrawal of chronic cAMP stimulation minimizesbasal CFTR activity, although it also increases Na+ conductancein both non-CF and CF epithelia (30), perhaps by increasingENaC expression (31). After amiloride addition, Gt did notdiffer between CF and non-CF epithelia, consistent with minimalCFTR conductance in non-CF epithelia (Fig. 4A). Importantly,with the reduced Cl− conductance, values of basal Vt, ΔVtamil,and ΔIscamil did not exceed or were only slightly greater thanthose in non-CF epithelia (compare Fig. 4 B and C with Fig. 2).We predicted a similar effect if we removed Cl− from the
bathing solutions. Fig. 5 shows that in Cl−-free solutions, basalVt, ΔVtamil, and ΔIscamil did not differ between CF and non-CFepithelia. Of note, Cl−-free solutions reduced basal Isc in bothnon-CF and CF epithelia; we do not know the mechanism.We predicted that inhibiting CFTR would increase Vt, ΔVtamil,
and ΔIscamil. We applied CFTRinh-172, which inhibits CFTR(32) but also may have a small inhibitory effect on Na+ transport(33), and found that it reduced Gt as expected (Fig. 6A). This
A
B
Fig. 3. Apical Na+ current and Na+ fluxes in non-CF and CF epithelia. (A)Amiloride-sensitive current in epithelia with a permeabilized basolateralmembrane and an apical-to-basolateral Na+ concentration gradient. n = 11non-CF and 8 CF epithelia, all from different donors. (B) Open-circuit Na+
unidirectional and net Na+ flux rates determined isotopically under basalconditions and after addition of 100 μM amiloride apically. JNaap-bl indicatesNa+ flux from the apical to the basolateral surface, JNabl-ap indicates flux inthe opposite direction, and JNanet indicates net flux. n = 9 non-CF and 9 CFdonors with one pair of epithelia from each donor. In matched epithelia, Vtwas −20 ± 5 mV for non-CF and −24 ± 4 mV for CF epithelia; Isc was 44 ± 7μA·cm−2 for non-CF and 47 ± 8 μA·cm−2 for CF; Gt was 2.2 ± 0.3 mS·cm−2 fornon-CF and 1.8 ± 0.2 mS·cm−2 for CF.
10262 | www.pnas.org/cgi/doi/10.1073/pnas.1106695108 Itani et al.

result, as well as the finding that removing Cl− reduced Gt morein non-CF than in CF epithelia (Fig. 5 A and D) indicates that,under basal conditions, CFTR channels show some constitutiveactivity. CFTRinh-172 also acutely increased basal Vt (Fig. 6C).Isc under basal conditions was not altered; that result was notsurprising, because Isc was similar in non-CF and CF epithelia(Fig. 6B). CFTRinh-172 also increased ΔIscamil and ΔVtamil (Fig.6 E and F). These results indicate that acutely inhibiting CFTRincreases Vt, ΔVtamil, and ΔIscamil so that they are more similarto the values in CF epithelia.Each of these three maneuvers has limitations. Withdrawal of
chronic cAMP stimulation also increases Na+ channel activityand probably affects other processes, removing Cl−-reducedbasal Isc and possibly causing changes in cell volume, andCFTRinh-172 can have other effects in addition to inhibitingCFTR. It is possible that each of these interventions could af-fect non-CF and CF epithelia differently. However, consistentwith our measurements of ΔGtamil, all the data indicate that non-CF/CF differences in values of basal Vt, ΔVtamil, and ΔIscamilare not the consequence of increased apical Na+ conductancebut instead result from the presence/absence of an apicalCl− conductance.
Estimates of Gp in Non-CF and CF Epithelia. Because Gp influencesGt and thereby Vt, we asked if it was reduced in CF epithelia.We estimated Gp by measuring Gt after Gc was minimized asfollows: CFTR anion conductance was decreased by withdrawalof chronic cAMP stimulation; ENaC was inhibited with ami-loride; and other apical Cl− channels were inhibited with DIDS.Under these conditions, Gt was the same in non-CF and CFepithelia (Fig. 7). Those Gt values also were similar to those inepithelia genetically lacking CFTR function (i.e., CF) and trea-ted with amiloride and DIDS but not by withdrawal of chroniccAMP stimulation. These data suggest that Gp did not differbetween CF and non-CF epithelia. This conclusion also is con-sistent with passive (basolateral-to-apical) 22Na+ fluxes, whichoccur primarily through the paracellular pathway; these fluxes
did not differ between non-CF and CF epithelia (Fig. 3). PassiveNa+ fluxes also were similar in wild-type and CFTR−/− porcineepithelia (22).
Basal Amil DIDS F&I Bumet0
1
2
3
Non-CF
CF
** *
Gt(
mS
.cm
-2)
Basal Amil DIDS F&I Bumet0
15
30
45-
-
-Non-CF
CF* * * *
Vt(
mV
)
Basal Amil DIDS F&I Bumet0
20
40
60
80
Non-CF
CF
* * * *
Isc
(µA
.cm
-2)
B
C
A
-1.5
-1.0
-0.5
0.0
Non-CF CF
*
Gt a
mil(m
S.c
m-2
)
0
10
20
30
40
Non-CF CF
*
Vt a
mil(m
V)
-80
-60
-40
-20
0
Non-CF CFIs
c am
il(µ
A.c
m-2
)
Fig. 4. Transepithelial electrophysiological properties in non-CF and CFepithelia following withdrawal of chronic cAMP stimulation. Left panelsindicate Gt (A), Isc (B), and Vt (C) under basal conditions and after additionsindicated in legend of Fig. 1. Right panels show changes with amilorideaddition. In some cases error bars are hidden by symbols. n = epithelia from261 non-CF donors and 105 CF donors; data for each donor were the averageof measurements from 1–12 epithelia (three epithelia were measured formost donors). *P < 0.05, unpaired t test.
0
1
2
3
Basal Amil
CF
Non-CF
*
*
Gt(
mS
.cm
-2)
0
1
2
3
Basal Amil
CF
Non-CF
Gt(
mS
.cm
-2)
0
10
20
30
Basal Amil
CF
Non-CF
-
-
-
*
*
Vt(
mV
)
0
10
20
30
Basal Amil
CF
Non-CF
-
-
-
Vt(
mV
)
0
20
40
60
Basal Amil
CF
Non-CF
*
0
20
40
60
Basal Amil
CF
Non-CF
A B C
D E FCl- free solution
Cl- containing solution
Fig. 5. Effect of Cl−-free solution on the electrophysiological response toamiloride. Gt, Isc, and Vt under basal conditions and after amiloride additionin Cl−-containing (A–C) and Cl−-free (D–F) solutions. n = 12 non-CF and 13 CFepithelia for Cl−-containing solutions and 15 non-CF and 14 CF epithelia forCl−-free solutions. *Difference in CF vs. non-CF epithelia, P < 0.05, unpairedt test.
0
2
4
Basal INH
Gt(
mS
.cm
-2)
0
1
2
Vehicle INH
-
-
Gt a
mil(m
S.c
m-2
)
0
20
40
Basal INH
-
-Vt(
mV
)
0
50
100
Basal INH
Isc
(µA
.cm
-2)
0
40
80
INHVehicle
-
-
Isc a
mil(µ
A.c
m-2
)
0
20
40
INHVehicle
Vt a
mil(m
V)
A B C
D E F
Fig. 6. Effect of CFTRinh-172 on electrophysiological properties of non-CFairway epithelia. (A–C) Gt, Isc, and Vt before and after addition of 100 μMCFTRinh-172 (INH). Each symbol indicates a different donor; n = 9. CFTRinh-172decreased Gt and increased Vt but did not alter Isc (P = 0.0016, 0.0024, and0.17, respectively, paired t test). Vehicle treatment (not shown) did not alterGt, Vt, or Isc (all P > 0.27, paired t test). (D–F) ΔGtamil, ΔIscamil, and ΔVtamil inepithelia treated with vehicle or CFTRinh-172. Each set of data points indi-cates a pair of epithelia from an individual donor with one receiving CFTRinh-172 and the other vehicle. ΔIscamil and ΔVtamil but not ΔGtamil were greaterin epithelia treated with CFTRinh-172 (P = 0.003, 0.0002, and 0.27, respec-tively, paired t test).
Itani et al. PNAS | June 21, 2011 | vol. 108 | no. 25 | 10263
MED
ICALSC
IENCE
S

DiscussionCF Airway Epithelia Have Reduced Anion Conductance but NotIncreased Na+ Conductance. Our data allow four main conclusionsabout electrolyte transport in CF vs. non-CF airway epithelia.First, CF airway epithelia have a markedly reduced trans-
epithelial Cl− conductance under basal conditions. Moreover,there was no increase in Cl− conductance after cellular levels ofcAMP were increased. These data are consistent with previousstudies done in vivo, in excised tissue, and in cultured epitheliafrom humans, pigs, ferrets, and mice (1, 7, 20, 22, 34, 35). Theseresults were expected, because CFTR is a phosphorylation-regulated anion channel (36).Second, non-CF epithelia had some constitutive CFTR Cl−
conductance under basal conditions. This result is consistent withprevious observations (26).Third, our measurements of Gt, apical Na+ current, and
22Na+ fluxes indicate that Na+ conductance in CF epithelia didnot exceed that in non-CF epithelia.Fourth, we found that basal Vt, ΔVtamil, and ΔIscamil were
greater in CF than in non-CF epithelia. Our data indicate thatloss of a Cl− conductance was responsible for these electro-physiological changes in CF epithelia. This conclusion was sup-ported by studies that reduced the Cl− conductance of non-CF epithelia.
Comparison of Our Results with Previous Studies. Our conclusionthat CF epithelia do not have increased Na+ conductance con-trasts with some previous interpretations that have been based,in part, on in vivo measurements of nasal Vt and measurementsof Vt and Isc (or equivalent current) in cultured epithelia (7, 15).As our results here and in porcine epithelia indicate, using thosevalues to make conclusions about Na+ conductance and Na+
absorption might lead to the wrong conclusion. However, otherstudies also have been done. With 22Na+ flux measurementsmade under short-circuit conditions, one study reported greaternet Na+ absorption in CF epithelia, and another reported nodifference between non-CF and CF epithelia (9, 11). However,22Na+ flux rates performed under open-circuit conditions inhuman (37, 38) and porcine (22) epithelia showed no differencebetween non-CF and CF epithelia; those results agree with ourdata. Moreover, open-circuit conditions better reflect in vivoconditions than short-circuit conditions.Measurements of the rate of liquid absorption, which depends in
part on Na+ conductance, have shown increased (39, 40), similar(41), and reduced (30) rates in CF vs. non-CF epithelia. Althoughwe did not measure liquid absorption, our data predict that CFabsorption would not exceed non-CF absorption rates. The ex-planation for differences between studies might relate to cultureconditions, the state of epithelial differentiation, the time of studyafter seeding, and the activity of CFTR (30). It also is noteworthythat in some reports (40) Gt did not differ between the two groupsof epithelia, whereas we found that basal Gt was reduced in CF.
After liquid was added to the surface of airway epithelia,periciliary liquid depth has been reported to be decreased incultured CF epithelia under equilibrium conditions (39, 41);however, such reductions do not necessarily indicate Na+hyperabsorption and also might result from lack of Cl− secretionor alterations in other processes. A comparison of CF and non-CF cultured airway epithelia under equilibrium conditions foundno difference in periciliary liquid depth (42). Moreover, peri-ciliary liquid depth was not reduced in the trachea of newbornCFTR−/− pigs (22). In humans with CF, there was a tendency fora reduced airway surface liquid height, although disease wasadvanced at the time of the study, and the comparison with non-CF was not statistically significant (43). Finally, the inherentvariability among epithelia from different donors can limit con-clusions based on epithelia from small numbers of donors.
This Study Has Advantages and Limitations. This study has someadvantages. (i) We studied differentiated primary cultures ofairway epithelia, avoiding potential limitations of studying celllines in which genetic drift is possible. This approach also elim-inates the possibility that recombinant CFTR might be expressedin cell types and at levels different from the endogenous protein.(ii) Study of cultured epithelia minimizes acute effects of in-flammatory mediators. (iii) We report values for all the non-CFand CF epithelial cultures from our laboratories; we applied nospecific criteria in selecting epithelia for study, a process thatcould introduce unintended biases.Our study also has limitations. (i) We used cultured and ex-
cised epithelia rather than studying epithelia in vivo. However,methodology to measure Isc and Gt in vivo are not available, andin vivo CF epithelia are subject to inflammation, infection, andremodeling. (ii) Epithelia were produced with cells removedfrom lungs that often were diseased; it is unknown whetherdisease status might have consequences for electrolyte transportafter the cells are cultured. (iii) ENaC channel activity is regu-lated by proteolysis of its extracellular domain and insertion intothe membrane (44, 45). Our study might not detect effects of thein vivo CF environment that modify ENaC or its regulators (46).(iv) We did not study HCO3
− transport, which may play an im-portant role in CF airway pathogenesis. (v) We did not studyseveral other channels, pumps, and transporters that likely playimportant roles in airway epithelial function and CF patho-physiology. (vi) Culture methods can influence electrophysio-logical properties (12, 47). However, methods that generatedifferentiated airway epithelia, as ours do (25), do not markedlyalter responses to amiloride and cAMP-elevating agents (12).Moreover, our cultured and excised non-CF epithelia had rela-tively similar basal Isc and responses to pharmacological inter-ventions. In addition, we recently found similar properties forporcine airway epithelia studied in vivo, as freshly excised tissue,and as differentiated cultures (20, 22). These comparisons pro-vide some assurance that our culture procedures can provide aninformative model for investigating ion transport. Nevertheless,we recognize that there may be differences between CF and non-CF human epithelia that are not reflected in primary cultures.(vii) Airway region can influence electrophysiological properties,and our study was limited to epithelia cultured from largebronchi and trachea. As an example, basal Isc and ΔIscamil weregreater in CF than in non-CF porcine nasal epithelia but did notdiffer between CF and non-CF tracheal/bronchial epithelia (22).Those differences were caused by greater CFTR-mediated Cl−conductance under basal conditions in nasal vs. tracheal/bron-chial epithelia and by quantitative differences in other transportprocesses, including basolateral K+ channels.
Implications for CF. Several hypotheses about the pathogenesis ofCF airway disease postulate that abnormal electrolyte transportacross airway epithelia and submucosal glands makes an importantcontribution to disease. Our data indicate that loss of anion con-ductance is a key abnormality in CF airway epithelia. The increasedCF epithelial Vt and Isc responses to amiloride that sometimes are
0
1
2
cAMP - + - +Non-CF CF
withdrawal:
Gt a
mil
&D
IDS
(mS
.cm
-2)
*
Fig. 7. Estimated paracellular conductance. Data are Gt in the presence ofamiloride and DIDS for non-CF and CF epithelia. n = 261 non-CF and 74 CFepithelia under control conditions and 261 non-CF and 105 CF epitheliatreated with withdrawal of chronic cAMP stimulation. Value for untreatednon-CF epithelia differed from other conditions (P < 0.0001 one-way ANOVAand Bonferroni post test).
10264 | www.pnas.org/cgi/doi/10.1073/pnas.1106695108 Itani et al.

interpreted to mean that CF epithelia hyperabsorb Na+ could beattributed to loss of Cl− conductance rather than to increased ac-tivity of Na+ channels. Thus, our data correspond to studies ofnewborn CF pigs done at a time when they manifest a defect inhost defense against bacteria but not secondary manifestationsof the disease (21, 22). Our results also are consistent with studiesof CF sweat gland ducts and airway submucosal glands, whichshow reduced anion transport but not Na+ hyperabsorption (2, 48,49). Although our studies do not address the possibility that in-creased Na+ conductance might be important at some stage of thedisease or that inhibiting ENaC might benefit patients with CF,these results focus attention on defective anion transport in CFlung disease.
Materials and MethodsProcedures for developing primary cultures of human airway epithelia andobtaining excised epithelia are similar to those previously reported (25).
Electrophysiological and Na+ flux measurements are similar to those pre-viously described (22). Please see SI Materials and Methods for a detaileddescription of the methods.
ACKNOWLEDGMENTS. We thank Tami Nesselhauf, Janice Launspach, andTheresa Mayhew for excellent assistance. We appreciate the help andassistance of the Iowa Donor Network and are deeply grateful to thepersons who donated their lungs for these studies. We appreciate thevaluable assistance of the University of Iowa In Vitro Models and Cell CultureCore supported in part by Grants R458-CR07 from the Cystic FibrosisFoundation, HL51670 and HL61234 from the National Heart, Lung, andBlood Institute, and DK54759 from the National Institute of Diabetes andDigestive and Kidney Diseases. This work was supported by Grant HL51670from the National Heart, Lung, and Blood Institute. O.A.I. was a recipient ofan Iowa Cardiovascular Interdisciplinary Research Fellowship (HL007121).P.H.K. is a Research Specialist, and M.J.W. is an Investigator of The HowardHughes Medical Institute.
1. Welsh MJ, Ramsey BW, Accurso F, Cutting GR (2001) Cystic fibrosis. The Metabolicand Molecular Basis of Inherited Disease, eds Scriver CR, et al. (McGraw-Hill, NewYork), 8th Ed.
2. Quinton PM (1999) Physiological basis of cystic fibrosis: A historical perspective.Physiol Rev 79(1, Suppl):S3–S22.
3. Wine JJ (1999) The genesis of cystic fibrosis lung disease. J Clin Invest 103:309–312.4. Verkman AS, Song Y, Thiagarajah JR (2003) Role of airway surface liquid and
submucosal glands in cystic fibrosis lung disease. Am J Physiol Cell Physiol 284:C2–C15.5. Rowe SM, Miller S, Sorscher EJ (2005) Cystic fibrosis. N Engl J Med 352:1992–2001.6. Davis PB (2006) Cystic fibrosis since 1938. Am J Respir Crit Care Med 173:475–482.7. Boucher RC (2007) Airway surface dehydration in cystic fibrosis: Pathogenesis and
therapy. Annu Rev Med 58:157–170.8. Knowles M, Gatzy JT, Boucher RCJ (1981) Increased bioelectric potential difference
across respiratory epithelia in cystic fibrosis. N Engl J Med 305:1489–1495.9. Knowles MR, et al. (1983) Abnormal ion permeation through cystic fibrosis respiratory
epithelium. Science 221:1067–1070.10. Widdicombe JH, Welsh MJ, Finkbeiner WE (1985) Cystic fibrosis decreases the apical
membrane chloride permeability of monolayers cultured from cells of trachealepithelium. Proc Natl Acad Sci USA 82:6167–6171.
11. Boucher RCJ, Stutts MJ, Knowles MR, Cantley LC, Gatzy JT (1986) Na+ transport incystic fibrosis respiratory epithelia. Abnormal basal rate and response to adenylatecyclase activation. J Clin Invest 78:1245–1252.
12. Sachs LA, Finkbeiner WE, Widdicombe JH (2003) Effects of media on differentiation ofcultured human tracheal epithelium. In Vitro Cell Dev Biol Anim 39:56–62.
13. Smith JJ, Welsh MJ (1992) cAMP stimulates bicarbonate secretion across normal, butnot cystic fibrosis airway epithelia. J Clin Invest 89:1148–1153.
14. Standaert TA, et al. (2004) Standardized procedure for measurement of nasalpotential difference: An outcome measure in multicenter cystic fibrosis clinical trials.Pediatr Pulmonol 37:385–392.
15. Donaldson SH, Boucher RC (2007) Sodium channels and cystic fibrosis. Chest 132:1631–1636.
16. Stutts MJ, et al. (1995) CFTR as a cAMP-dependent regulator of sodium channels.Science 269:847–850.
17. Nagel G, et al. (2005) CFTR fails to inhibit the epithelial sodium channel ENaCexpressed in Xenopus laevis oocytes. J Physiol 564:671–682.
18. Rogers CS, et al. (2008) Disruption of the CFTR gene produces a model of cystic fibrosisin newborn pigs. Science 321:1837–1841.
19. Meyerholz DK, Stoltz DA, Pezzulo AA, Welsh MJ (2010) Pathology of gastrointestinalorgans in a porcine model of cystic fibrosis. Am J Pathol 176:1377–1389.
20. Ostedgaard LS, et al. (2011) The ΔF508 mutation causes CFTR misprocessing and cysticfibrosis-like disease in pigs. Sci Transl Med 3: 74ra24.
21. Stoltz DA, et al. (2010) Cystic fibrosis pigs develop lung disease and exhibit defectivebacterial eradication at birth. Sci Transl Med 2(29): 29ra31.
22. Chen J-H, et al. (2010) Loss of anion transport without increased sodium absorptioncharacterizes newborn porcine cystic fibrosis airway epithelia. Cell 143:911–923.
23. Horisberger JD (2003) ENaC-CFTR interactions: The role of electrical coupling of ionfluxes explored in an epithelial cell model. Pflugers Arch 445:522–528.
24. Duszyk M, French AS (1991) An analytical model of ionic movements in airwayepithelial cells. J Theor Biol 151:231–247.
25. Karp PH, et al. (2002) An in vitro model of differentiated human airway epithelia:Methods and evaluation of primary cultures. Epithelial Cell Culture Protocols, edWise C (Humana, Totowa, NJ), Vol 188, pp 115–137.
26. Uyekubo SN, et al. (1998) cAMP-dependent absorption of chloride across airwayepithelium. Am J Physiol 275:L1219–L1227.
27. Welsh MJ (1983) Inhibition of chloride secretion by furosemide in canine trachealepithelium. J Membr Biol 71:219–226.
28. Helman SI, Miller DA (1973) Edge damage effect on electrical measurements of frogskin. Am J Physiol 225:972–977.
29. Quinton PM (2010) Role of epithelial HCO3⁻ transport in mucin secretion: Lessons from
cystic fibrosis. Am J Physiol Cell Physiol 299:C1222–C1233.30. Zabner J, Smith JJ, Karp PH, Widdicombe JH, Welsh MJ (1998) Loss of CFTR chloride
channels alters salt absorption by cystic fibrosis airway epithelia in vitro. Mol Cell 2:397–403.
31. Thomas CP, Campbell JR, Wright PJ, Husted RF (2004) cAMP-stimulated Na+ transportin H441 distal lung epithelial cells: Role of PKA, phosphatidylinositol 3-kinase, andsgk1. Am J Physiol Lung Cell Mol Physiol 287:L843–L851.
32. Ma T, et al. (2002) Thiazolidinone CFTR inhibitor identified by high-throughputscreening blocks cholera toxin-induced intestinal fluid secretion. J Clin Invest 110:1651–1658.
33. Wang XF, ReddyMM, Quinton PM (2004) Effects of a new cystic fibrosis transmembraneconductance regulator inhibitor on Cl− conductance in human sweat ducts. Exp Physiol89:417–425.
34. Grubb BR, Boucher RC (1999) Pathophysiology of gene-targeted mouse models forcystic fibrosis. Physiol Rev 79(1, Suppl):S193–S214.
35. Sun X, et al. (2010) Disease phenotype of a ferret CFTR-knockout model of cysticfibrosis. J Clin Invest 120:3149–3160.
36. Sheppard DN, Welsh MJ (1999) Structure and function of the CFTR chloride channel.Physiol Rev 79(1, Suppl):S23–S45.
37. Willumsen NJ, Boucher RCJ (1991) Sodium transport and intracellular sodium activityin cultured human nasal epithelium. Am J Physiol 261:C319–C331.
38. Willumsen NJ, Boucher RCJ (1991) Transcellular sodium transport in cultured cysticfibrosis human nasal epithelium. Am J Physiol 261:C332–C341.
39. Matsui H, et al. (1998) Evidence for periciliary liquid layer depletion, not abnormal ioncomposition, in the pathogenesis of cystic fibrosis airways disease. Cell 95:1005–1015.
40. Jiang C, Finkbeiner WE, Widdicombe JH, McCray PB, Jr., Miller SS (1993) Altered fluidtransport across airway epithelium in cystic fibrosis. Science 262:424–427.
41. Van Goor F, et al. (2009) Rescue of CF airway epithelial cell function in vitro by a CFTRpotentiator, VX-770. Proc Natl Acad Sci USA 106:18825–18830.
42. Derichs N, Jin BJ, Song Y, Finkbeiner WE, Verkman AS (2011) Hyperviscous airwaypericiliary and mucous liquid layers in cystic fibrosis measured by confocal fluorescencephotobleaching. FASEB J, 10.1096/fj.10-179549.
43. Griesenbach U, et al. (2011) Quantification of periciliary fluid (PCL) height in humanairway biopsies is feasible, but not suitable as a biomarker. Am J Respir Cell Mol Biol44:309–315.
44. Hughey RP, et al. (2004) Epithelial sodium channels are activated by furin-dependentproteolysis. J Biol Chem 279:18111–18114.
45. Snyder PM (2005) Minireview: Regulation of epithelial Na+ channel trafficking.Endocrinology 146:5079–5085.
46. Myerburg MM, Harvey PR, Heidrich EM, Pilewski JM, Butterworth MB (2010) Acuteregulation of the epithelial sodium channel in airway epithelia by proteases andtrafficking. Am J Respir Cell Mol Biol 43:712–719.
47. Widdicombe JH, Sachs LA, Finkbeiner WE (2003) Effects of growth surface ondifferentiation of cultures of human tracheal epithelium. In Vitro Cell Dev Biol Anim39:51–55.
48. Quinton PM (2007) Cystic fibrosis: Lessons from the sweat gland. Physiology(Bethesda) 22:212–225.
49. Joo NS, Irokawa T, Robbins RC, Wine JJ (2006) Hyposecretion, not hyperabsorption, isthe basic defect of cystic fibrosis airway glands. J Biol Chem 281:7392–7398.
Itani et al. PNAS | June 21, 2011 | vol. 108 | no. 25 | 10265
MED
ICALSC
IENCE
S





























