Embed Size (px)
Citation preview

Human ERK1 Induces Filamentous Growth and Cell WallRemodeling Pathways in Saccharomyces cerevisiae*
Received for publication, December 16, 1999, and in revised form, April 27, 2000Published, JBC Papers in Press, April 27, 2000, DOI 10.1074/jbc.M910024199
Josephine M. Atienza‡, Michael Suh‡, Ioannis Xenarios§¶, Ralf Landgraf§, and John Colicelli‡i
From the ‡Department of Biological Chemistry and the Molecular Biology Institute, UCLA School of Medicine and theMolecular Biology Institute and the §UCLA-DOE Laboratory of Structural Biology & Molecular Medicine,Los Angeles, California 90095
Expression of an activated extracellular signal-regu-lated kinase 1 (ERK1) construct in yeast cells was usedto examine the conservation of function among mitogen-activated protein (MAP) kinases. Sequence alignment ofthe human MAP kinase ERK1 with all Saccharomycescerevisiae kinases reveals a particularly strong kinshipwith Kss1p (invasive growth promoting MAP kinase),Fus3p (pheromone response MAP/ERK kinase), andMpk1p (cell wall remodeling MAP kinase). A fusion pro-tein of constitutively active human MAP/ERK kinase 1(MEK) and human ERK1 was introduced under regu-lated expression into yeast cells. The fusion protein(MEK/ERK) induced a filamentation response elementpromoter and led to a growth retardation effect concom-itant with a morphological change resulting in elon-gated cells, bipolar budding, and multicell chains. In-duction of filamentous growth was also observed fordiploid cells following MEK/ERK expression in liquidculture. Neither haploids nor diploids, however, showedmarked penetration of agar medium. These effects couldbe triggered by either moderate MEK/ERK expression at37 °C or by high level MEK/ERK expression at 30 °C. Thecombination of high level MEK/ERK expression and37 °C resulted in cell death. The deleterious effects ofMEK/ERK expression and high temperature were signif-icantly mitigated by 1 M sorbitol, which also enhancedthe filamentous phenotype. MEK/ERK was able to con-stitutively activate a cell wall maintenance reportergene, suggesting misregulation of this pathway. In con-trast, MEK/ERK effectively blocked expression from apheromone-responsive element promoter and inhibitedmating. These results are consistent with MEK/ERK pro-moting filamentous growth and altering the cell wallthrough its ability to partially mimic Kss1p and stimu-late a pathway normally controlled by Mpk1p, whileappearing to inhibit the normal functioning of the struc-turally related yeast MAP kinase Fus3p.
Mitogen-activated protein (MAP)1 kinases constitute a fam-ily of enzymes that control a variety of functions in eukaryotic
cells. In response to an extracellular stimulus, particular MAPkinases become activated following the sequential triggering ofupstream kinases, producing a kinase cascade. This results inthe phosphorylation of substrates involved in both short termand long term (e.g. transcription-mediated) cell changes. Astheir name implies, activation of some MAP kinases can lead toa proliferative response. However, it is now clear that membersof this family of kinases can initiate a variety of other cell fatepathways, such as differentiation and/or cell cycle arrest. Insome cases, the activation of a given MAP kinase can lead todivergent cell fates depending upon the initiating stimulus orthe cell type (1, 2). The regulation of MAP kinases and theirsignaling specificity are controlled at multiple levels (reviewedin Refs. 3 and 4). The association of some MAP kinases andupstream cascade components with scaffold proteins appears tobe one way of orchestrating both incoming activation signalsand outgoing substrate phosphorylation signals (reviewed inRef. 5). Subcellular localization is another point that is utilizedfor regulating the accessibility of MAP kinases to their sub-strates. In particular, most MAP kinases have both cytoplasmicand nuclear substrates, necessitating the use of nuclear importmechanisms to direct signaling (6). Signal attenuation throughthe intervention of appropriate phosphatases (reviewed in Ref.7) and feedback phosphorylation (8) are also involved. Incorpo-rated within these regulatory mechanisms is the ability tomodulate both the intensity and duration of phosphorylationsignals, directly influencing the cell response (1). Once a MAPkinase is activated and properly localized, the final outcome ispresumably dictated by substrate availability in a given celltype and by the intrinsic substrate specificity of each MAPkinase.
As with other isozyme families, it is likely that the strikinglevels of sequence identity among the MAP kinases reflectsboth an evolutionary kinship of the genes encoding these en-zymes and a structural constraint dictated by a shared cata-lytic mechanism. Conversely, the sequence variations amongMAP kinases presumably highlight residues not strictly requiredfor core enzyme function and should include determinants ofisozyme-specific properties, such as substrate preference.
The yeast Saccharomyces cerevisiae encodes six MAP ki-nases (9) that share extensive sequence identity among them-selves and with mammalian MAP kinases. For most of theyeast MAP kinase enzymes, activating extracellular stimuli,upstream kinase cascades, and phenotypic outcomes have beencharacterized (reviewed in Ref. 10). In particular, Fus3p andKss1p have been studied in great detail and have served asparadigms for the study of MAP kinase regulation and func-tion. In wild type cells, Fus3p responds to pheromone binding,and its activation produces a mating response that includesadoption of an elongated morphology (shmoo). Kss1p acts tocontrol invasive (filamentous) growth, which results from dif-
* This work was supported by National Institutes of Health GrantNS31911. The costs of publication of this article were defrayed in partby the payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
¶ A fellow of the Swiss National Fund for Research.i To whom correspondence should be addressed. Tel.: 310-206-7800;
Fax: 310-206-5272; E-mail: [email protected] The abbreviations used are: MAP, mitogen-activated protein; ERK,
extracellular signal-regulated kinase; FRE, filamentation response el-ement; GST, glutathione S-transferase; HA, hemagglutinin; MEK, mi-togen-activated protein kinase/extracellular signal-regulated kinase ki-nase; PRE, pheromone-responsive element.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 275, No. 27, Issue of July 7, pp. 20638–20646, 2000© 2000 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org20638
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

ferent cues in haploid and diploid cells. And Kss1p has beenimplicated in the maintenance of cell wall integrity duringvegetative growth (11). The Fus3 and Kss1 proteins sharestriking sequence and function relatedness (12, 13). BothFus3p and Kss1p respond to the same upstream activatingkinases (Ste11p and Ste7p), and gene deletion experimentshave indicated that Kss1p can functionally substitute forFus3p in the pheromone response, although at lower efficiency(12, 14). In addition, both Fus3p and Kss1p activate promotersthat utilize the Ste12p transcription activator. Pathway dis-crimination is achieved in part through the intervention of thescaffold protein Ste5p, which preferentially recruits Fus3p intoa pheromone-responsive kinase cascade (5) and through theintervention of Ste12p-collaborating transcription factors, suchas Tec1p, which is believed to be filamentous growth-specific.The distinct properties of Fus3p and Kss1p are also likely toreflect differences in substrate specificity dictated by sequencevariation.
S. cerevisiae haploids normally propagate as spherical, axialbudding, individual cells in culture. Filamentous growth rep-resents a differentiated state that requires the coordinatedinduction of cell elongation, bipolar budding, incomplete cellseparation, and invasion (e.g. of agar medium). Similarchanges can occur in diploid cells. Kss1p is a principal regula-tor controlling the switch to filamentous growth in both hap-loids and diploids (12, 15). These phenotypes also require dra-matic alterations in the cellular cytoskeleton and the cell walland are likely mediated by multiple pathways (16–18).
A key regulator of cell wall integrity is the MAP kinaseMpk1p. The activity of this kinase is itself controlled by theupstream kinases Mkk1/2p and Bck1p. This MAP kinase cas-cade is regulated by Pkc1p, which appears to respond to up-stream signaling from Rho1p in response to a membrane local-ized sensor of cell integrity encoded by the WSC1/HCS77/SLG1gene (19–22). This pathway must also be recruited for theorganized remodeling of the cell wall in the transition to mat-ing or filamentous morphologies (23–25). Hallmarks of disrup-tion in this signaling pathway include cell lysis that is exacer-bated by high temperature and mitigated by isoosmotic growthmedia (e.g. media with 1 M sorbitol) and by heightened sensi-tivity to caffeine (20, 26, 27).
We used inducible expression of an activated human ERK1in yeast cells to examine issues of MAP kinase substrate spec-ificity controlling cellular responses. Primary sequence align-ments have previously suggested a close relationship betweenmammalian ERK1 and particular yeast MAP kinases. We pres-ent experimental data and a modified alignment approach toaddress structure/function conservation.
Changes in cell morphology and specific reporter gene acti-vation reported here suggest that human ERK1 functions as anactivator of filamentous morphology and cell wall remodelingpathways normally regulated by the yeast kinases Kss1p andMpk1p and inhibits signals normally channeled through thehighly related kinase Fus3p. The inducible system describedhere may also prove useful for the isolation of human MAPkinase interaction partners and regulators.
EXPERIMENTAL PROCEDURES
Strains and Growth Assays—Escherichia coli strain DH5a was usedfor the construction and propagation of plasmids. Yeast strains used inthis work are listed in Table I. Transformations were performed usinga modified lithium acetate method (28). Cells expressing MEK/ERK orthe control plasmid (pYNH) were grown in synthetic complete mediumwithout uracil or leucine, unless otherwise indicated. Growth assayswere carried out by inoculating 105 cells from a log phase culture into 10ml of selective medium with or without Cu21 (0.5 mM) or sorbitol (1 M)and at 30 or 37 °C. Quantitative matings were carried out essentially asdescribed previously (29) using 105 tester cells mixed with 107 partner
cells in YPD diluted 10-fold with water prior to plating on syntheticcomplete-histidine medium. Cells were examined using a Nikon Dia-phot 200 with a 3100 objective and a phase condenser.
Plasmids—The MEK/ERK plasmid was constructed from humanMEK1 and human ERK1 clones generously provided by Natalie Ahnand Melanie Cobb, respectively. The constitutively active mutantMEK1 (DN3/S218W/S222D) was polymerase chain reaction-amplifiedwith the primers TCAGGTCGACCGGAGTTGGAAGCGCGTTA andTACGAAGCTTGCTAGCGACGCCAGCAGCATGGGTT and initiallycloned into the SalI and HindIII sites of pBluescript KS (Stratagene).This construct (pKS-MEK/ERK) was then digested with NheI and NotIfollowed by ligation with the NheI to NotI ERK1 fragment ofpCEP4Erk1. The MEK1/ERK1 fusion fragment was then released withSalI and NotI and cloned directly into pYNH, a form of pYEX (AMRADBiotech) that had been previously modified to change the unique EcoRIsite to NotI and to include an HA epitope sequence between the BamHIand SalI sites. To create the MEK1-only construct, the pKS-MEK/ERKplasmid was cut with NheI and NotI to release the ERK1 sequencesthen ligated in the presence of adaptor oligonucleotides. The resultingSalI to NotI fragment of MEK1 was then moved to the modified pYEXdescribed above. Similarly, to make the ERK1-only construct, MEKsequences were released from the fusion construct by digestion withSalI plus NheI followed by ligation in the presence of appropriateadaptor oligonucleotides. The PRE(FUS1) and FRE(ty1) reporter con-structs were provided by the laboratory of Dr. Gerald Fink (Massachu-setts Institute of Technology, Cambridge, MA). The FKS2 reporter wasprovided by Dr. David Levin (Johns Hopkins University, Baltimore,MD). All three reporter plasmids originally contained URA3 markergenes, and each was changed to HIS3 by homologous recombination.
Immunoblots, Protein Purification, and Kinases Assays—HA-MEK/ERK fusion protein was expressed in SP1. Cultures were grown tologarithmic phase and induced with CuSO4 to a final concentration of0.5 mM for 3 h. Cultures were resuspended in phosphate-buffered salineplus leupeptin, pepstatin, and phenylmethylsulfonyl fluoride. Theywere lysed as described previously (28). 50 mg of total extract wasloaded on a gel and transferred to nitrocellulose. Membrane was probedwith HA monoclonal antibody (12CA5), and detected by ECL (Amer-sham Pharmacia Biotech). Immunoprecipitations and kinase assayswere done as described previously (30) except that lysates were precip-itated with HA antibody (1 ml of ascites from 12CA5 hybridoma) for 1 h.Kinase reactions were carried out in a total volume of 40 ml using 2 mgof purified Elk1 (New England BioLabs) as substrate.
GST-FAR1 and GST-STE12 were prepared from BL21 cells trans-fected with expression plasmids from Matthias Peter (Swiss Institutefor Experimental Cancer Research) and Stan Fields (University ofWashington), respectively. Purification on glutathione beads (Amer-sham Pharmacia Biotech) was according to the manufacturer’s instruc-tions using 50 ml of bead volume for extract from a 200 ml culture. MEK,ERK, and MEK/ERK proteins were immunoprecipitated from yeast cellextracts (approximately 8 mg of total protein) using HA antibody. Ki-nase assays were performed using immunoprecipitated material resus-pended in buffer (10 mM Tris (pH 7.5), 10 mM MgCl, 100 mM sodiumvanadate). Assays were carried out at 30 °C for 30 min, and the prod-ucts were separated by SDS-polyacrylamide gel electrophoresis.
Gene Expression Assays—Liquid b-galactosidase assays for PRE andFRE reporters were performed and quantified by established tech-niques (31). Liquid b-galactosidase assays for the FKS2 reporter wereperformed by another method (32) due to the difficulty of lysing cellsgrown at 39 °C. The enzyme activity units for each method are distinctand not comparable. Yeast strains W303-1A and SP1 were used (see
TABLE IList of strains
Strains Genotype
SP1 MATa ade8 his3 leu2 trp1 ura3 can1a
W303–1A MATa ade2 his3 leu2 trp1 ura3 can1b
L5684 MATa leu2 ura3 RED11c
L6149 MATa his3 leu2 ura3 tec1::HIS3c
JTY256 MATa ade2 his3 leu2 trp1 ura3 can1 ste12::URA3d
L6018 MATa his3 leu2 trp1 ura3 fus3::TRP1 RED11c
DC14 MATa his1a
DC17 MATa his1a
DC124 MATa ade8 his4 leu2 trp1 ura3a
a Cold Spring Harbor Laboratory strain collection.b Dr. Rodney Rothstein laboratory strain collection.c Dr. Gerald Fink laboratory strain collection.d Dr. Jeremy Thorner laboratory strain collection.
MAP Kinase Functional Conservation 20639
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Table II). Pheromone induction was assayed in W303-1A cells usingpSB234, a 2-m based PRE(FUS1)::lacZ reporter construct. This con-struct was transformed into yeast cells containing pYNH or pYME.Transformants were grown overnight in selective medium, switched toYPD, and treated with a-Factor (Sigma) to a final concentration of 2.5mM for 2.5 h. Cultures were then lysed and assayed. Similar resultswere obtained in the SP1 strain background. Filamentous inductionwas assayed in SP1 cells using a modified pLG669Z, a 2-m basedFRE(Ty1)::lacZ reporter construct. Similar results were obtained forthis reporter in the W303-1A strain background. Induction of the cellwall remodeling pathway was assayed in SP1 cells using anFKS2(-706)::lacZ reporter construct.
Sequence Alignments—All data base searches were performed withthe complete set of yeast open reading frames. Initial multiple align-ments were obtained by applying ClustalW (33). Generalized profileconstruction and searches were run locally using the PFTOOLS pack-age, version 2.0. Evolutionary surface patch analysis was used to iden-tify functional surface patches in ERK1 and the yeast MAP kinases. Inshort, conservation difference scores are calculated for windows of po-sitions, with each window representing a profile of residues adjacent inthree-dimensional space rather than primary sequence. Relatednessbetween sequences A and B can be viewed as a minimum conservationdifference score between two data sets (including or excluding B). Ashift in local sequence similarities was used to identify regions of theprotein in which the “local phylogeny” differs from that of the fullsequence. The contribution of specific proteins to this “shift score” canbe evaluated as described above for the conservation difference score.This analysis is an extension of the previously described evolutionarytracing analysis (34, 35) and will be discussed in detail elsewhere.2
RESULTS
Constitutively Active Human ERK1 Causes Growth Arrest inYeast—We created an active MAP kinase by connecting humanERK1 sequences to a constitutively active mutant form of hu-man MEK1 (37). The resulting fusion protein, MEK/ERK, wasalso expected to show constitutive ERK1 kinase activity result-ing from the steady state phosphorylation of this enzyme.MEK/ERK, which carries an HA epitope tag, was detectable inyeast extracts and appeared to have approximately the pre-dicted molecular weight (Fig. 1). The expression vector usedincluded both URA3 and leu2d selectable markers and em-ployed a CUP1 promoter for Cu21-inducible expression.Growth in medium without leucine results in selection for highplasmid copy number and gave rise to a significant increase ofprotein levels, presumably due to basal expression from theCUP1 promoter. The addition of Cu21 produced a further in-crease in the amount of protein (Fig. 1), that was maximal after1–3 h (data not shown).
Enzymatic activity was demonstrated by kinase assays ofimmunoprecipitated MEK/ERK protein from yeast cell lysates.MEK/ERK immunoprecipitated from yeast showed autophos-phorylation and was capable of phosphorylating purified mam-malian ELK1 protein, a well characterized ERK1 substrate(Fig. 2). This was not true for activated MEK alone. Immuno-purified ERK appeared to be phosphorylated in the kinase
reaction, perhaps reflecting the presence of a co-purified endog-enous yeast MEK-type kinase, and this resulted in relativelyweak kinase activity for ELK1. However, a mixture of MEKand ERK extracts resulted in substantial ERK phosphorylationand high level ELK1 phosphorylation. These results confirmedthat, when expressed in yeast, the combination of constitu-tively activated human MEK with wild type human ERK,whether separate or covalently attached as a fusion protein,still shows normal substrate reactivity. Indeed, a similar con-struct fusing MEK1 with ERK2 has been reported to functionas a constitutively active kinase in mammalian cells (38).
MEK/ERK Causes Growth Retardation and MorphologicalChanges in Yeast—We examined the growth properties of hap-loid (3A) and diploid (3B) yeast cells expressing the MEK/ERKfusion and observed severe growth suppression. When grown at30 °C, this phenotype was strongest following induction of theCUP1 promoter. Cells expressing MEK or ERK constructsalone had no discernible growth defect (data not shown).
The effect of elevated temperature on cell growth was alsoexamined. High temperature is a well studied stress conditionthat triggers a variety of cell responses, including cell wallremodeling (21, 39). When cultured at 37 °C, cells expressingMEK/ERK showed a marked long term reduction in growthcompared with cells expressing vector only or MEK/ERK cellsgrown at 30 °C (Fig. 3). In the presence of CuSO4, which in-duces higher levels of MEK/ERK expression, the negative ef-
2 R. Landgraf, I. Xenarios, and D. Eisenberg, manuscript inpreparation.
FIG. 2. Catalytic activity of MEK/ERK. Extracts from cultures ofyeast cells transformed with the indicated construct were subjected toimmunoprecipitation using epitope tag (HA) antibody. Precipitated ma-terial was subjected to a kinase reaction in the presence of absence ofpurified ELK1 protein. Molecular weight markers (kDa) are indicatedat the left.
TABLE IIMating efficiency (diploid colonies per mating pair)
SP1 (MATa) cells were paired with DC14 (MATa), DC17 (MATa) orDC124 (MATa) cells and mating was assayed as described under “Ex-perimental Procedures.” V and ME indicate vector and MEK/ERK con-struct transformants, respectively. DC14 and DC17 matings were per-formed in triplicate. DC124 mating repeated in a separate experiment(data not shown) that gave consistent results.
DC14 DC17 DC124-V DC124-ME
SP1-V ,1 3 1025 0.5 6 0.1 0.5 0.007SP1-ME ,1 3 1025 0.009 6 0.003 0.007 0.003SP1-V (Cu21) 0.5 6 0.1 0.2 0.003SP1-ME (Cu21) 0.005 6 0.003 0.006 0.0004
FIG. 1. Expression of MEK/ERK fusion protein in yeast. Ex-tracts of cultures transformed with MEK/ERK or vector were analyzedby immunoblot using epitope tag (HA) antibody. Molecular weightmarkers (kDa) are indicated at the left.
MAP Kinase Functional Conservation20640
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

fects of high temperature were exacerbated. Under these con-ditions, the MEK/ERK-expressing cells were unable to growand showed no recovery after transfer to fresh medium withoutCuSO4 at 30 °C (data not shown).
We next examined the effect of 1 M sorbitol on growth ofMEK/ERK-expressing cells. This condition has been shown torescue a variety of yeast cell wall integrity mutations, presum-ably by providing an isoosmotic environment in which cell walldefects can be tolerated. We observed that sorbitol rescued thegrowth defect of MEK/ERK-expressing cells with CuSO4-in-duced levels at 30 °C as well as uninduced (basal) levels at37 °C. The combined effects of high MEK/ERK expression andelevated temperature, however, could not be rescued by sorbi-tol (Fig. 3).
An examination of MEK/ERK-expressing cells under variousgrowth conditions showed several morphological alterations(Fig. 4A). These included an elongated cell morphology andshort linear chains indicative of polar cell division withoutseparation. The changes were most pronounced when cellswere grown in sorbitol and were enhanced by induction of high
level MEK/ERK expression. These features are reminiscent ofcells undergoing invasive growth, a filamentous phenotype nor-mally controlled by an endogenous yeast kinase cascade utiliz-ing the Kss1p MAP kinase.
Diploid cells can normally undergo a related type of filamen-tous growth producing pseudohyphae cells (40). In response tonitrogen starvation conditions, cells become elongated, switchto a unipolar budding pattern, do not fully separate leading tocell chains, and exhibit agar invasive properties. The switch topseudohyphae growth, like filamentous growth in haploids, iscontrolled in part by Kss1p. We expressed MEK/ERK in diploidcells and observed a striking shift to predominantly elongatedcells with a high proportion of cells found in short chains andclumps (Fig. 4A). It should be noted that this apparent induc-tion of filamentous growth took place in liquid medium withnormal levels of ammonium as nitrogen source, conditions notnormally conducive to this growth conversion. As with hap-loids, 1 M sorbitol enhanced these effects, whereas elevatedMEK/ERK expression appeared to produce a somewhat dis-torted cell shape.
FIG. 3. Growth suppression byMEK/ERK in haploid and diploidcells. A, the haploid strain SP1 wastransformed with the indicated plasmid,and equal inoculums were cultured in se-lective medium (see under “ExperimentalProcedures”) with or without CuSO4 andsorbitol. Parallel cultures at 30 °C and37 °C were analyzed after 48 h growth.A600 is indicated at the left. Data are froma representative experiment. B, the dip-loid strain SP1/DC124 was transformedand treated as described above. Data pre-sented are mean values from an experi-ment carried out in triplicate.
MAP Kinase Functional Conservation 20641
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from
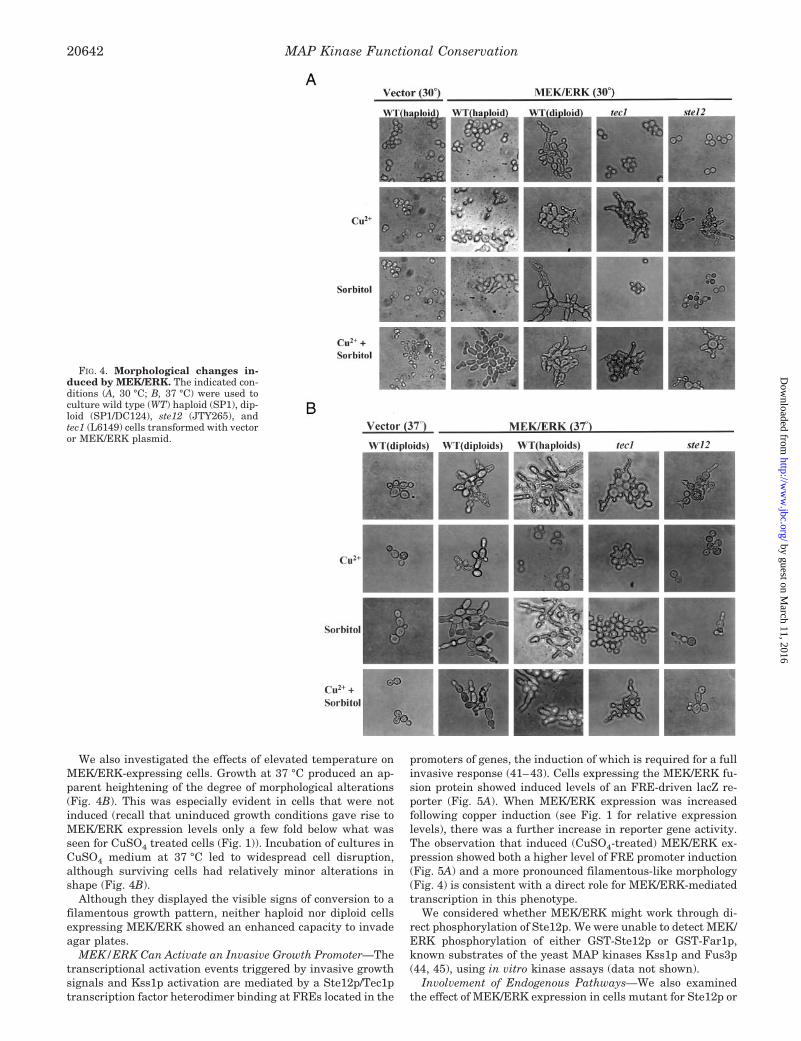
We also investigated the effects of elevated temperature onMEK/ERK-expressing cells. Growth at 37 °C produced an ap-parent heightening of the degree of morphological alterations(Fig. 4B). This was especially evident in cells that were notinduced (recall that uninduced growth conditions gave rise toMEK/ERK expression levels only a few fold below what wasseen for CuSO4 treated cells (Fig. 1)). Incubation of cultures inCuSO4 medium at 37 °C led to widespread cell disruption,although surviving cells had relatively minor alterations inshape (Fig. 4B).
Although they displayed the visible signs of conversion to afilamentous growth pattern, neither haploid nor diploid cellsexpressing MEK/ERK showed an enhanced capacity to invadeagar plates.
MEK/ERK Can Activate an Invasive Growth Promoter—Thetranscriptional activation events triggered by invasive growthsignals and Kss1p activation are mediated by a Ste12p/Tec1ptranscription factor heterodimer binding at FREs located in the
promoters of genes, the induction of which is required for a fullinvasive response (41–43). Cells expressing the MEK/ERK fu-sion protein showed induced levels of an FRE-driven lacZ re-porter (Fig. 5A). When MEK/ERK expression was increasedfollowing copper induction (see Fig. 1 for relative expressionlevels), there was a further increase in reporter gene activity.The observation that induced (CuSO4-treated) MEK/ERK ex-pression showed both a higher level of FRE promoter induction(Fig. 5A) and a more pronounced filamentous-like morphology(Fig. 4) is consistent with a direct role for MEK/ERK-mediatedtranscription in this phenotype.
We considered whether MEK/ERK might work through di-rect phosphorylation of Ste12p. We were unable to detect MEK/ERK phosphorylation of either GST-Ste12p or GST-Far1p,known substrates of the yeast MAP kinases Kss1p and Fus3p(44, 45), using in vitro kinase assays (data not shown).
Involvement of Endogenous Pathways—We also examinedthe effect of MEK/ERK expression in cells mutant for Ste12p or
FIG. 4. Morphological changes in-duced by MEK/ERK. The indicated con-ditions (A, 30 °C; B, 37 °C) were used toculture wild type (WT) haploid (SP1), dip-loid (SP1/DC124), ste12 (JTY265), andtec1 (L6149) cells transformed with vectoror MEK/ERK plasmid.
MAP Kinase Functional Conservation20642
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Tec1p, proteins that regulate transcription from FRE promot-ers. Although induction of the FRE reporter construct waseliminated in the ste12 strain (Fig. 5), significant morphologi-cal changes still resulted following MEK/ERK expression (Fig.4). Unlike wild type cells, however, the cell shape deviatedconsiderably from classic filamentous morphology even in thepresence of sorbitol. In addition, either CuSO4 or 37 °C wererequired for any effect. These results suggest that some mor-phological alterations resulting from high level MEK/ERK ex-pression are not strictly dependent on the multipurpose tran-scription factor Ste12p. Similar results were obtained in a tec1strain expressing MEK/ERK (Fig. 4). These findings are con-
sistent with a model that some aspects of the MEK/ERK ex-pression phenotype involve the Ste12p-Tec1p heterodimertranscription factor working at FRE type promoters. However,because significant morphological changes are induced byMEK/ERK in the absence of either Ste12p or Tec1p, otherfactors must be mediating this response.
MEK/ERK Disrupts Cell Wall Integrity—ERK1 has beencategorized within the same subgroup of MAP kinases asKss1p, Fus1p, and Mpk1p (9, 46). Alignment of the full cata-lytic domain of the ERK1 protein (344 residues) with all six S.cerevisiae MAP kinases showed a particularly strong relation-ship with Kss1p and Fus3p. When alignment analysis was
FIG. 5. MEK/ERK expression differ-entially regulates MAP kinase path-way reporters. Wild type haploid cells(see under “Experimental Procedures”)transformed with vector or MEK/ERKconstructs, together with a FRE::lacZ re-porter (A), FKS2::lacZ reporter (B), orPRE::lacZ reporter (C), were grown underthe indicated conditions and assayed forlacZ expression. Different lacZ units (Aand C versus B) reflect different assaytechniques used (see under “Experimen-tal Procedures” and “Results”).
MAP Kinase Functional Conservation 20643
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

restricted to the 63-amino acid active site domain directlyimplicated in substrate specificity (including the P11, lip, andL13 regions) (47, 48), a close similarity between ERK1 andMpk1p was also revealed (Fig. 6). The “docking site” of a MAPkinase is an important determinant for interaction with regu-lators and substrates (49). In this sequence, ERK1 shows clearrelatedness to Kss1p, Fus3p, and Mpk1p. We also performedevolutionary surface patch analysis to identify amino acid po-sitions associated with MAP kinase isozyme specificity. Thismethod uses a reference protein structure (ERK1) to studyconservation/divergence of neighbor residues that may not becontiguous in the primary sequence. The 63-amino acid activesite domain showed the highest conservation scores amongdata sets including or excluding Kss1p and Mpk1p (data notshown). Shift scores were then used to highlight regions ofdivergent phylogeny that are dependent on Kss1p and Mpk1pinclusion in the data set. This revealed contiguous surfacepatches representing putative Kss1p/Mpk1p functional speci-ficity regions that are also closely related to ERK1. Theseresults support a structural basis for the observed Kss1p-likephenotypes resulting from ERK1 expression in yeast. They alsosuggested a possible functional relatedness between ERK1 andMpk1p.
We therefore considered the possible involvement of cell wallintegrity pathways in the phenotypes resulting from MEK/ERK expression. The observations that elevated temperatureand 1 M sorbitol enhanced the filamentous morphology of MEK/ERK-expressing yeast cells suggested that the requisite remod-eling of the cell wall was not taking place efficiently. Anotherhallmark of cell wall maintenance defects is heightened caf-feine sensitivity (20, 26, 27). We observed a marked increase incaffeine sensitivity in MEK/ERK-expressing cells (Fig. 7), aresult consistent with cell wall disruption but perhaps attrib-
utable to nonspecific effects of MEK/ERK expression. To exam-ine this further, we utilized a reporter construct derived fromthe FKS2 (50) promoter known to be responsive to this path-way following activation by high temperature conditions (39).At 25 °C the expression of MEK/ERK produced a 4-fold in-crease in reporter gene activity (Fig. 5B). At elevated levels ofMEK/ERK, there was a 20-fold difference, correlating MEK/ERK expression directly with activation of this promoter. In-cubation of vector transformed cells at 39 °C resulted in astrong induction of the reporter, as expected. This level was notincreased by MEK/ERK, suggesting that the conditions usedmay have yielded maximum reporter expression.
MEK/ERK Expression Inhibits Mating—In contrast to theFRE and FKS2 promoter activation results, we observed thatthe MEK/ERK fusion protein was unable to induce expressionfrom the FUS1 promoter, which includes a pheromone re-sponse element (Fig. 5C). This reporter is strongly induced bytreatment of cells with pheromone (a-Factor), which initiates akinase cascade that terminates with activation of the MAPkinase Fus3p. Expression of MEK/ERK had a potent inhibitoryeffect on the induction of this reporter by pheromone. Themagnitude of the pheromone signal suppression, like those ofthe FRE and FKS2 promoter signal activation, was directlyrelated to MEK/ERK expression levels. Expression of MEKalone or ERK1 alone, which is mostly inactive (Fig. 2), did notcause reduction in the pheromone-induced reporter signal(data not shown).
The mating competence of MEK/ERK-expressing cells wasalso severely compromised (Table II). MATa cells (SP1) ex-pressing MEK/ERK showed a greater than 50-fold reduction inmating efficiency with a MATa tester strain (DC17). This effectwas also seen when MEK/ERK was expressed in MATa cells(DC124) and the block was greater in matings between strains
FIG. 6. Sequence alignment of human ERK1 and the six S. cerevisiae MAP kinases (Kss1p, Fus3p, Mpk1p, Hog1p, Mlp1p, andSmk1p). The regions presented (catalytic core (top panel) and substrate interaction/docking domain (bottom panel)) were chosen based on minimalconservation difference scores. Columns indicate the “shift” or divergent phylogeny score uncorrected (white) or corrected (black) for surfaceexposure. Structural domains (L, loop; a, a-helix) are designated according to established nomenclature (47). Amino acid positions for eachsequence are given at the left. Gaps are indicated by dashes. Black bars denote regions identified by this analysis as signature sequences for Kss1pand Mpk1p.
MAP Kinase Functional Conservation20644
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

that each expressed MEK/ERK. An increase in MEK/ERK lev-els (CuSO4 medium) did not lead to significant further de-creases in mating efficiencies, however, suggesting that theexpression levels without induction (Fig. 1) are sufficient forthe full effect on mating, as assayed here. There was no detect-able mating between MEK/ERK-expressing MATa cells and aMATa tester strain (DC14), indicating no loss in mating part-ner discrimination.
DISCUSSION
The human ERK1 protein shares extensive sequence iden-tity with all six MAP kinases from S. cerevisiae. Alignmentsfocusing specifically on key structural elements involved insubstrate interactions (catalytic core and docking region), to-gether with surface patch analysis, placed ERK1 in a subgroupwith Kss1p, Fus3p, and Mpk1p. Although previous studieshave noted the close sequence relationship between ERK1 andKss1p (46, 51, 52), this work represents a comprehensive anal-ysis with all yeast MAP kinase sequences and reveals a func-tional correlation. Integration of experimental data and struc-ture-based analysis, as described here, should aid in developinguseful approaches to the study of isozymes in general and MAPkinases in particular.
Expression of MEK/ERK, a constitutively active kinase, trig-gered phenotypes reminiscent of filamentous growth, suggest-ing that MEK/ERK can at least partially mimic activatedKss1p. Similar changes were induced by MEK/ERK expressionin diploid cells, in which filamentous growth normally requiresnitrogen starvation. Because neither constitutively active MEKnor wild type ERK alone produced these results, we concludethat this filamentous behavior is dependent on the activity ofERK stimulated by constitutively active MEK.
Characterization of the observed phenotypes as filamentousis supported by the ability of MEK/ERK to induce an FREreporter construct. This activity was absent in ste12 cells, con-sistent with the requirement for Ste12p to activate this pro-moter. However, the ste12 mutant cells expressing MEK/ERKstill displayed some changes in morphology. This suggests theinvolvement of other pathways and is consistent with previousgenetic analysis of the filamentous response (16). Indeed, ste12mutants have a greatly reduced, but not absent, response toinvasive growth signals (16, 53). In addition, filamentousgrowth induced by overexpression of Whi2p and Phd1p or byactivation of PKA have also been shown to be mostly Ste12p
independent (16, 18, 54). It should be noted that Ste12p, whichis also a component of the mating signal pathway, is not essen-tial for some pheromone-induced morphological changes (23).We observed that tec1 mutant cells showed similar abnormalmorphologies in response to MEK/ERK expression. Tec1p part-ners with Ste12p in the control of at least some filamentousresponse element promoters (16, 41, 55, 56). The observationthat ste12 and tec1 mutants respond similarly to MEK/ERKexpression, with either mutation blocking the cell elongationseen in wild type cells, is consistent with the known propertiesof the Ste12p/Tec1p heterodimer. Indeed, these mutations ap-pear to have similar, though not identical, effects on geneexpression (57).
Kss1p and Fus3p have a special relationship; each is moreakin to the other than to any of the remaining yeast MAPkinases, and each can influence responses normally controlledby the other. Kss1p, for example, can block the Fus3p inductionof pheromone-responsive genes (58). This effect has been pos-tulated to require the inactive form of Kss1p in the stabiliza-tion of PRE complexes (59). MEK/ERK can also block phero-mone-responsive gene transcription as well as mating. Becausea constitutively active construct was required, however, thisinhibitory effect may indicate a different type of repressionfrom that mediated by Kss1p.
The fact that MEK/ERK can activate an FRE-driven reporterbut strongly inhibits a PRE-driven reporter may reflect boththe influence of specific components (e.g. Tec1p) and differencesin the context of Ste12p (59, 60) that alter its interaction withMEK/ERK. Also, the persistent expression of MEK/ERK (evenwithout copper) might lead to down-regulation of pathwaycomponents.
Our data indicate that MEK/ERK expression results in dis-ruption of cell wall integrity, a pathway regulated by Pkc1pand the MAP kinase Mpk1p in response to stress and differen-tiation signals. Incubation at 37 °C, a condition known to in-duce cell wall remodeling, heightened both the growth retar-dation and morphological changes resulting from MEK/ERKexpression. In addition,1 M sorbitol not only ameliorated thegrowth retardation resulting from MEK/ERK expression butalso enhanced the filamentous-like phenotype. Taken togetherwith the induction of the FKS2 reporter, these alterationsstrongly indicate cell wall disruptions resulting from activatedERK. The slow growth, heat sensitivity, and caffeine sensitiv-ity phenotypes associated with MEK/ERK may result fromoverexpression of genes that are only transiently induced dur-ing a normal response to heat or the adoption of new morphol-ogies in mating or a switch to invasive growth. Indeed, al-though most studies of this pathway have employedinactivating mutations, overactivation of the homologous path-way in Schizosaccharomyces pombe leads to aberrant cell mor-phology and cell growth defects (61, 62) similar to what we havedescribed. Whether MEK/ERK works by recognizing and phos-phorylating Mpk1p substrates or by acting, like Kss1p, througha parallel cell wall pathway that also regulates FKS2 expres-sion in vegetative cells (11) is not yet clear.
Divergence between human ERK1 and yeast Kss1p shouldrepresent sequences not important for basic catalytic function.These would include amino acids required for interfacing withupstream activation or downstream signal attenuation pro-teins. Conversely, residues that are conserved between ERK1and Kss1p should include those involved directly or indirectlyin catalysis. Indeed, of the 12 residues found mutated in defec-tive Kss1p alleles and predicted to play a direct role in catalyticfunction (12), all are identical in ERK1. Amino acids involvedin the substrate specificity characteristics shared betweenERK1 and either Kss1p or Mpk1p may also be conserved.
FIG. 7. MEK/ERK expression causes caffeine sensitivity, a cellwall defect-associated phenotype. Yeast cells (SP1) transformedwith vector or MEK/ERK were streaked on selective media with orwithout caffeine (4 mM) and CuSO4 (0.5 mM) as indicated, and incubatedat 30 °C for 3 days.
MAP Kinase Functional Conservation 20645
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Despite the parallel between structural and functional relat-edness it is clear that ERK1 can not provide complete func-tional redundancy with either Kss1p or Mpk1p. This is in partdue to the unregulated activity of our constructs: ERK1 alone isinactive and unresponsive to endogenous yeast activators, andMEK/ERK is constitutively active and unresponsive to endog-enous yeast attenuators. More fundamentally, the kinship ofthe enzyme with Kss1p and Mpk1p likely extends to only asubset of target proteins.
Our findings clearly demonstrate that MEK/ERK can inducemultiple cytoplasmic and cytoskeletal alterations requisite forfilamentous growth. MEK/ERK expression also results in ab-errant activation of cell wall remodeling, a process that isnormally regulated to accompany cell shape changes. The fila-mentous morphology enhancement observed in 1 M sorbitolindeed suggests that this growth program is principally dic-tated by cytoskeletal changes that are then accommodated bycell wall modifications. Similarly, hog1 and pbs2 mutants ofyeast show pheromone pathway activation and apparent mat-ing projections, but only in the presence of 1 M sorbitol (63).MEK/ERK might act directly on Spa2p and/or SphIp, two regu-lators of polarized morphogenesis that interact with Mpk1p andits upstream activating kinases Mkk1p and Mkk2p (64, 65).
Numerous mammalian oncoproteins are known to activateMAP kinases, including ERK1 and ERK2. These signals havebeen directly associated with the induction of transformationand differentiation pathways (38), each of which involves ex-tensive cytoskeletal remodeling and morphological changes,and ERK kinase pathways have been implicated in the controlof metastasis (migration and tissue invasion) in tumor cells (36,66). The inducible model system described here may providethe basis for genetic isolation of mammalian ERK1 regulatorsthat could affect these functions.
Acknowledgments—We acknowledge the contributions to experimen-tal analysis by Mark Grieb, Daniel McDonnell, and Negin Sohrabi. Wealso thank Gerald Fink, David Levin, Jeremy Thorner, Ira Herskowitz,Mathias Peter, Stan Fields, Roymarie Ballester, Melanie Cobb, andNatalie Ahn for providing critical reagents and Fuyu Tamanoi andDavid Eisenberg for useful comments.
REFERENCES
1. Marshal, C. J. (1995) Cell 80, 179–1852. Brunner, D., Oellers, N. Szabad, J., Biggs, W. H., Zipursky, L. S., and Hafen,
E. (1994) Cell 76, 875–8883. Schaeffer, H. J., and Weber, M. J. (1999) Mol. Cell. Biol. 19, 2435–24444. Garrington, T. P., and Johnson, G. L. (1999) Curr. Opin. Cell Biol. 11, 211–2185. Whitmarsh, A. J., and Davis, R. J. (1998) Trends Biochem. Sci. 23, 481–4856. Khokhlatchev, A. V., Canagarajah, B., Wilsbacher, J., Robinson, M., Atkinson,
M., Goldsmith, E., and Cobb, M. H. (1998) Cell 93, 605–6157. Lewis, T. S., Shapiro, P. S., and Ahn, N. G. (1998) Adv. Cancer Res. 74, 49–1398. Errede, B., and Ge, Q. Y. (1996) Philos. Trans. R. Soc. Lond. B Biol. Sci. 351,
143–1489. Hunter, T., and Plowman, G. D. (1997) Trends Biochem. Sci. 22, 18–22
10. Madhani, H. D., and Fink, G. R. (1998) Trends Genet. 14, 151–15511. Lee, B. N., and Elion, E. A. (1999) Proc. Natl. Acad. Sci. U. S. A. 96,
12679–1268412. Madhani, H. D., Styles, C. A., and Fink, G. R. (1997) Cell 91, 673–68413. Cherkasova, V., Lyons, D. M., and Elion, E. A. (1999) Genetics 151, 989–100414. Elion, E. A., Brill, J. A., and Fink, G. R. (1991) Proc. Natl. Acad. Sci. U. S. A.
88, 9392–939615. Cook, J. G., Bardwell, L., and Thorner, J. (1997) Nature 390, 85–8816. Mosch, H.-U., and Fink, G. R. (1997) Genetics 145, 671–68417. Lorenz, M. C., Heitman, J. (1998) Genetics 150, 1443–145718. Pan, X., and Heitman, J. (1999) Mol. Cell. Biol. 19, 4874–488719. Kamada, Y., Qadota, H., Python, C. P., Anraku, Y., Ohya, Y., and Levin, D. E.
(1996) J. Biol. Chem. 271, 9193–919620. Verna, J., Lodder, A., Lee, K., Vagts, A., and Ballester, R. (1997) Proc. Natl.
Acad. Sci. U. S. A. 94, 13804–1380921. Rajavel, M., Philip, B., Buehrer, B. M., Errede, B., and Levin, D. E. (1999) Mol.
Cell. Biol. 19, 3969–397622. Jacoby, J. J., Nilius, S. M., and Heinisch, J. J. (1998) Mol. Gen. Genet. 258,
148–15523. Buehrer, B. M., and Errede, B. (1997) Mol. Cell. Biol. 17, 6517–652524. Zarzov, P., Mazzoni, C., and Mann, C. (1996) EMBO J. 15, 83–9125. Navarro-Garcıa, F., Alonso-Monge, R., Rico, H., Pla, J., Sentandreu, R., and
Nombela, C. (1998) Microbiology 144, 411–42426. Costigan, C., Gehrung, S., and Snyder, M. (1992) Mol. Cell. Biol. 12,
1162–117827. Yoshida, S., Ikeda, E., Uno, I., and Mitsuzawa, H. (1992) Mol. Gen. Genet. 231,
337–34428. Atienza, J. M., and Colicelli, J. (1998) Methods Companion Methods Enzymol.
14, 35–4229. Spain, B. H., Bowdish, K. S., Pacal, A. R., Staub, S. F., Koo, D., Chang, C. Y.,
Xie, W., and Colicelli, J. (1996) Mol. Cell. Biol. 16, 6698–670630. Afar, D., Han, L., McLaughlin, J., Wong, S., Dhaka, A., Parmar, K., Rosenberg,
N., Witte, O. N., and Colicelli, J. (1997) Immunity 6, 773–78231. Guarente, L. (1983) Methods Enzymol. 101, 181–19132. Rose, M. D., Winston, F., and Hieter, P. (1990) Methods in Yeast Genetics, Cold
Spring Harbor Laboratory, Cold Spring Harbor, NY33. Higgins, D. G., Thompson, J. D. & Gibson, T. J. (1996) Methods Enzymol. 266,
383–40234. Lichtarge, O., Bourne, H. R., and Cohen, F. E. (1996) J. Mol. Biol. 257,
342–35835. Landgraf, R., Fischer, D., and Eisenberg, D. S. (1999) Protein Eng. 12, 943–95136. Janulis, M., Silberman, S., Ambegaokar, A., Gutkind, J. S., and Schultz, R. M.
(1999) J. Biol. Chem. 274, 801–81337. Mansour, S. J., Matten, W. T., Hermann, A. S., Candia, J. M., Rong, S.,
Fukasawa, K., Vande Woude, G. F., and Ahn, N. G. (1994) Science 265,966–970
38. Robinson, M. J., Stippec, S. A., Goldsmith, E., White, M., and Cobb, M. H.(1998) Curr. Biol. 8, 1141–1150
39. Zhao, C., Jung, U. S., Garrett-Engele, P., Roe, T., Cyert, M. S., and Levin, D. E.(1998) Mol. Cell. Biol. 18, 1013–1022
40. Gimeno, C. J., Ljungdahl, P. O., Styles, C. A., and Fink, G. R. (1992) Cell 68,1077–1090
41. Madhani, H. D., Fink, G. R. (1997) Science 275, 1314–131742. Baur, M., Esch, R. K., and Errede, B. (1997) Mol. Cell. Biol. 17, 4330–433743. Lo, W. S., and Dranginis, A. M. (1998) Mol. Biol. Cell 9, 161–17144. Yuan, Y. O., Stroke, I. L., and Fields, S. (1993) Genes Dev. 7, 1584–159745. Peter, M., and Herskowitz, I. (1994) Science 265, 1228–123146. Ferrell, J. E. (1996) Curr. Top. Dev. Biol. 33, 1–6047. Zhang, F., Strand, A., Robbins, D., Cobb, M. H., and Goldsmith, E. J. (1994)
Nature 367, 704–71148. Canagarajah, B. J., Khokhlatchev, A., Cobb, M. H., and Goldsmith, E. J. (1997)
Cell 90, 859–86949. Tanoue, T., Adachi, M., Moriguchi, T., and Nishida, E. (2000) Nat. Cell Biol. 2,
110–11650. Mazur, P., Morin, N., Baginsky, W., el-Sherbeini, M., Clemas, J. A., Nielsen,
J. B., and Foor, F. (1995) Mol. Cell. Biol. 15, 5671–568151. Boulton, T. G., Yancopoulos, G. D., Gregory, J. S., Slaughter, C., Moomaw, C.,
Hsu, J., and Cobb, M. H. (1990) Science 249, 64–6752. Bardwell, L., Cook, J. G., Inouye, C. J., and Thorner, J. (1994) Dev. Biol. 166,
363–37953. Lo, H. J., Kohler, J. R., DiDomenico, B., Loebenberg, D., Cacciapuoti, A., and
Fink, G. R. (1997) Cell 90, 939–94954. Radcliffe, P. A., Binley, K. M., Trevethick, J., Hall, M., and Sudbery, P. E.
(1997) Microbiol. 143, 1867–187655. Gavrias, V., Andrianopoulos, A., Gimeno, C. J., and Timberlake, W. E. (1996)
Mol. Microbiol. 19, 1255–126356. Oehlen, L., and Cross, F. R. (1998) FEBS Lett. 429, 83–8857. Madhani, H. D., Galitski, T., Lander, E. S., and Fink, G. R. (1999) Proc. Natl.
Acad. Sci. U. S. A. 96, 12530–1253558. Bardwell, L., Cook, J. G., Voora, D., Baggott, D. M., Martinez, A. R., and
Thorner, J. (1998a) Genes Dev. 12, 2887–289859. Bardwell, L., Cook, J. G., Zhu-Shimoni, J. X., Voora, D., and Thorner, J.
(1998b) Proc. Natl. Acad. Sci. U. S. A. 95, 15400–1540560. Cook, J. G., Bardwell, L., Kron, S. J., and Thorner, J. (1996) Genes Dev. 10,
2831–284861. Toda, T., Shimanuki, M., and Yanagida, M. (1993) EMBO J. 12, 1987–199562. Hirata, D., Nakano, K., Fukui, Takenaka, H., Miyakawa, T., and Mabuchi, I.
(1998) J. Cell Sci. 111, 149–5963. O’Rourke, S. M., and Herskowitz, I. (1998) Genes Dev. 12, 2874–288664. Roemer, T., Vallier, L., Sheu, Y. J., and Snyder, M. (1998) J. Cell Sci. 111,
479–49465. Sheu, Y. J., Santos, B., Fortin, N., Costigan, C., and Snyder, M. (1998) Mol.
Cell. Biol. 18, 4053–406966. Webb, C. P., Van Aelst, L., Wigler, M. H., and Woude, G. F. (1998) Proc. Natl.
Acad. Sci. U. S. A. 95, 8773–8778
MAP Kinase Functional Conservation20646
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Josephine M. Atienza, Michael Suh, Ioannis Xenarios, Ralf Landgraf and John Colicelli Saccharomyces cerevisiae
Human ERK1 Induces Filamentous Growth and Cell Wall Remodeling Pathways in
doi: 10.1074/jbc.M910024199 originally published online April 27, 20002000, 275:20638-20646.J. Biol. Chem.
10.1074/jbc.M910024199Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/275/27/20638.full.html#ref-list-1
This article cites 65 references, 34 of which can be accessed free at
by guest on March 11, 2016
http://ww
w.jbc.org/
Dow
nloaded from





























