Embed Size (px)
Citation preview

Zhou et al. BMC Genomics (2022) 23:11 https://doi.org/10.1186/s12864-021-08245-2
RESEARCH
Identification and expression analysis of histone modification gene (HM) family during somatic embryogenesis of oil palmLixia Zhou*†, Rajesh Yarra†, Longfei Jin, Yaodong Yang, Hongxing Cao and Zhihao Zhao
Abstract
Background: Oil palm (Elaeis guineensis, Jacq.) is an important vegetable oil-yielding plant. Somatic embryogenesis is a promising method to produce large-scale elite clones to meet the demand for palm oil. The epigenetic mechanisms such as histone modifications have emerged as critical factors during somatic embryogenesis. These histone modifi-cations are associated with the regulation of various genes controlling somatic embryogenesis. To date, none of the information is available on the histone modification gene (HM) family in oil palm.
Results: We reported the identification of 109 HM gene family members including 48 HMTs, 27 HDMs, 13 HATs, and 21 HDACs in the oil palm genome. Gene structural and motif analysis of EgHMs showed varied exon–intron organiza-tion and with conserved motifs among them. The identified 109 EgHMs were distributed unevenly across 16 chromo-somes and displayed tandem duplication in oil palm genome. Furthermore, relative expression analysis showed the differential expressional pattern of 99 candidate EgHM genes at different stages (non-embryogenic, embryogenic, somatic embryo) of somatic embryogenesis process in oil palm, suggesting the EgHMs play vital roles in somatic embryogenesis. Our study laid a foundation to understand the regulatory roles of several EgHM genes during somatic embryogenesis.
Conclusions: A total of 109 histone modification gene family members were identified in the oil palm genome via genome-wide analysis. The present study provides insightful information regarding HM gene’s structure, their distribu-tion, duplication in oil palm genome, and also their evolutionary relationship with other HM gene family members in Arabidopsis and rice. Finally, our study provided an essential role of oil palm HM genes during somatic embryogenesis process.
Keywords: HMs, Somatic embryogenesis, Oil palm, Real-time PCR
© The Author(s) 2021. Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http:// creat iveco mmons. org/ licen ses/ by/4. 0/. The Creative Commons Public Domain Dedication waiver (http:// creat iveco mmons. org/ publi cdoma in/ zero/1. 0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
BackgroundHistone modifications gene family (HMs) play a vital role in plant growth and developmental processes by histone modification processes (methylation, dem-ethylation, acetylation, and deacetylation) either by
activating or silencing the gene expression. These four different histone modification processes are regulated by four different HM gene family members such as HMTs (histone methyltransferases), HDMs (histone demethy-lases), HATs (histone acetylases), and HDACs (histone deacetylases) [1, 2]. Each of the HM gene family mem-bers also consisted of a varied number of subfamilies with different domain structures. The HMTs contained SDG (set domain group) and PRMT (protein argi-nine methyltransferases) sub-families. The HDMs con-tained HDMA (SWIRM and C-terminal domain) and
Open Access
*Correspondence: [email protected]†Lixia Zhou and Rajesh Yarra are contributed equally to this work.Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences/ Hainan Key Laboratory of Tropical Oil Crops Biology, Wenchang, Hainan 571339, P. R. China

Page 2 of 15Zhou et al. BMC Genomics (2022) 23:11
JMJ (JmjC domain-containing proteins) sub-families. The HATs contained HAG (GCN5-, ELP3-, and HAT1-like histone acetylases domain structure), HAM (MOZ-YBF2 domain), HAC (p300/CREB-binding protein structure), and HAF (TATA-binding protein-associated factors TAFII250) sub-families. The HDACs contained HDA (RPD3/HDA1 superfamily), SRT (silent informa-tion regulator 2), and HDT (HD2 families) sub-families [3–7]. Numerous previous studies described the role of HM gene family members in regulating the vegeta-tive and reproductive growth, biotic and abiotic stress responses, stress-related hormone signaling [4, 8–16]. Various genome-wide studies revealed the occurrence of a varied number of HM gene family members in various plants including a total of 198 HMs in Malus domestica [8]; 125 HMs in Lycopersicum esculentum [17]; 136 HMs in Citrus sinensis [18]; 87 HMs in Litchi chinensis [9]. However, to date, none of the studies were published on genome-wide studies of HM gene family members in oil palm genome.
African oil palm (Elaeis guineensis, Jacq.) is the most promising and productive oil crop to accomplish the increasing demand for vegetable oils around the world [19, 20]. The vegetative propagation for large-scale pro-duction of oil palm plants is not possible due to the absence of auxiliary shoots and traditional seed prop-agation is hampered by the low seed germination rate [21, 22]. Moreover, genetic improvement of oil palm plants via seed propagation is the most complicated [22]. A promising substitute for the large-scale produc-tion of oil palm seedlings is the extensive micropropa-gation of plants via tissue culture method of approach i.e. somatic embryogenesis (SE) [22, 23]. In plants, somatic embryogenesis is the method of producing somatic embryos under in vitro conditions without the fusion of the gametes [24]. Somatic embryogenesis is the reliable and powerful biotechnological approach for the micropropagation of plants with low seed germina-tion rates as well as long reproductive cycles [25]. How-ever, the somatic embryogenesis response is differed from species to species depending on their totipotency capacity. Though somatic embryogenesis protocols were well established for the propagation of oil palm, it’s essential to understand metabolic, genetic, epige-netic, morphogenetic factors that boost the somatic embryogenesis process in oil palm plants. Moreover, epigenetic mechanisms such as methylation, demeth-ylation, acetylation, and deacetylation are regulating the gene expression that modulates the capacity of somatic embryogenesis during tissue culture [26]. The epigenetic mechanisms that regulate the gene expres-sion at somatic embryogenesis have not been addressed much in oil palm. Identification of genes that are
responsible for regulating the epigenetic mechanisms during somatic embryogenesis of oil palm is vital for a better understanding of SE process. Up to now, none of the studies were reported on histone modification (HM) gene family members in oil palm during different somatic embryogenesis phases.
In this study, we identified a total of 109 HM gene family members (48 HMTs; 27 HDMs; 13 HATs, and 21 HDACs) in the oil palm genome via the bioinformatics approach. We also analyzed the EgHMs gene structure, motif analysis, phylogenetic analysis, synteny, promoter analysis, subcellular localization, and their location on 16 chromosomes in oil palm genome. Further, we ana-lyzed the expression patterns of all identified EgHMs at different stages of the somatic embryogenesis process of oil palm.
ResultsIdentification of HM gene family in oil palm genomeIn this investigation, a total of 109 EgHM gene family members such as 48 of HMTs (histone methyltrans-ferases); 27 of HDMs (histone demethylases); 13 of HATs (histone acetylases), and 21 of HDACs (histone deacetylases) were successfully identified in the oil palm genome via genome-wide analysis. All the EgHM family members are categorized into 11 subfamilies (SDG, PRMT, HDMA, JMJ, HAG, HAM, HAC, HAF, HAD, SRT & HDT) based on their protein domain architecture. The HMT family is included with 39SDGs and 9PRMTs; HDM family is contained with 3HDMAs and 24JMJs; HAT family is included with 4HAGs, 2HAMs, 6HACs, and 1HAF; HDAC family included with 15HADs, 3SRTs, and 3 HDT sub-families. All of the gene IDs of EgHM family members were provided in Supplementary Table 1. The pI and Molecular weight of the oil palm HM family members were ranged from 4.57 to 9.95 and 12.3 to 273.3 respectively (Supplemen-tary Table 1). The EgHMs are predominantly localized to the cytoplasmic regions (Supplementary Table 2).
Gene structure and conserved motif analysis of EgHM family membersWe analyzed the gene structure of all EgHM gene fam-ily members using the Gene Structure Display Server tool 2.0 (http:// gsds. cbi. pku. edu. cn/). We found the occur-rence of varied numbers (2–34) of exons among the HM gene family members (Fig. 1). The highest number (34) of exons was observed in EgHMT members i.e. EgSDG18 and the least number (2) of exons were observed in EgSDG39 (Fig. 1). We found10 conserved motifs among the 109 HM gene family of oil palm (Fig. 2).

Page 3 of 15Zhou et al. BMC Genomics (2022) 23:11
Fig. 1 Gene structural organization of 109 EgHMs. Coding sequences (CDS) are represented by orange color blocks; 3’ & 5’ UTRs are regions are represented by blue color blocks; intron regions are represented by black color blocks in the diagram

Page 4 of 15Zhou et al. BMC Genomics (2022) 23:11
Chromosomal distribution of EgHM members in oil palm genomeThe chromosomal distribution of 109 EgHM gene
family members was also examined across the 16 chromosomes of the African oil palm genome. The EgHM gene family members were unevenly distributed
Fig. 2 Occurrence of conserved motifs in EgHM proteins. The MEME tool was used to identify the ten numbers of motifs in EgHM proteins. Each motif in EgHM proteins has shown with different colors. The sequence logo of each motif represents the abundance of each amino acid in their motifs

Page 5 of 15Zhou et al. BMC Genomics (2022) 23:11
on the chromosomes (Fig. 3). Among the identified 109EgHMs, only 86 were mapped across the 16 chro-mosomes (Fig. 3). We have not observed the mapping of 26 EgHM members on any of the chromosomes. Chromosomes 1 and 9 had the highest number (9) of EgHM family genes, whereas chromosome 14 had only one EgHM family gene (Fig. 3). Apart from chromo-some 14, all of the remaining chromosomes at least contained 3 or more EgHM family genes.
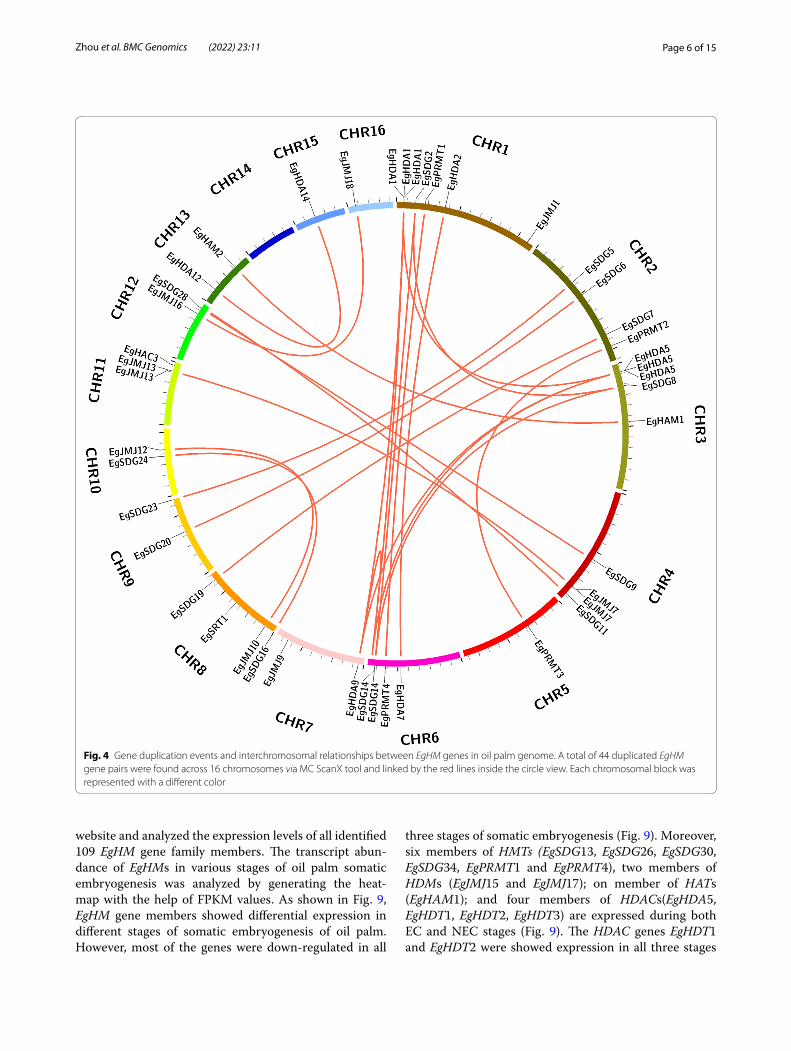
EgHM gene duplication in oil palm genomeTo know the expansion of the HM gene family in oil palm genome, we generated a gene duplication event diagram for duplicated blocks using a Circos algo-rithm. In a total, 37 pairs of EgHMs were identified from 16 chromosomes of oil palm, including 14 pairs of EgSDGs; 4 pairs of EgPRMTs; 6 pairs of EgJMJs; 2 pairs of EgHAMs; 1 pair of EgHACs; 9 pairs of EgH-DAs, and 1 pair of EgSRTs (Fig. 4). The paired EgHM duplicated genes were all located in different chromo-some blocks of oil palm genome (Fig. 4). Moreover, Chromosome 1 and Chromosome 6 had 6 and 5 num-ber of duplicated genes respectively. However, chro-mosome 14 block had no duplicated HM genes (Fig. 4). These results demonstrated the expansion of EgHM gene family that occurred through these duplicated regions.
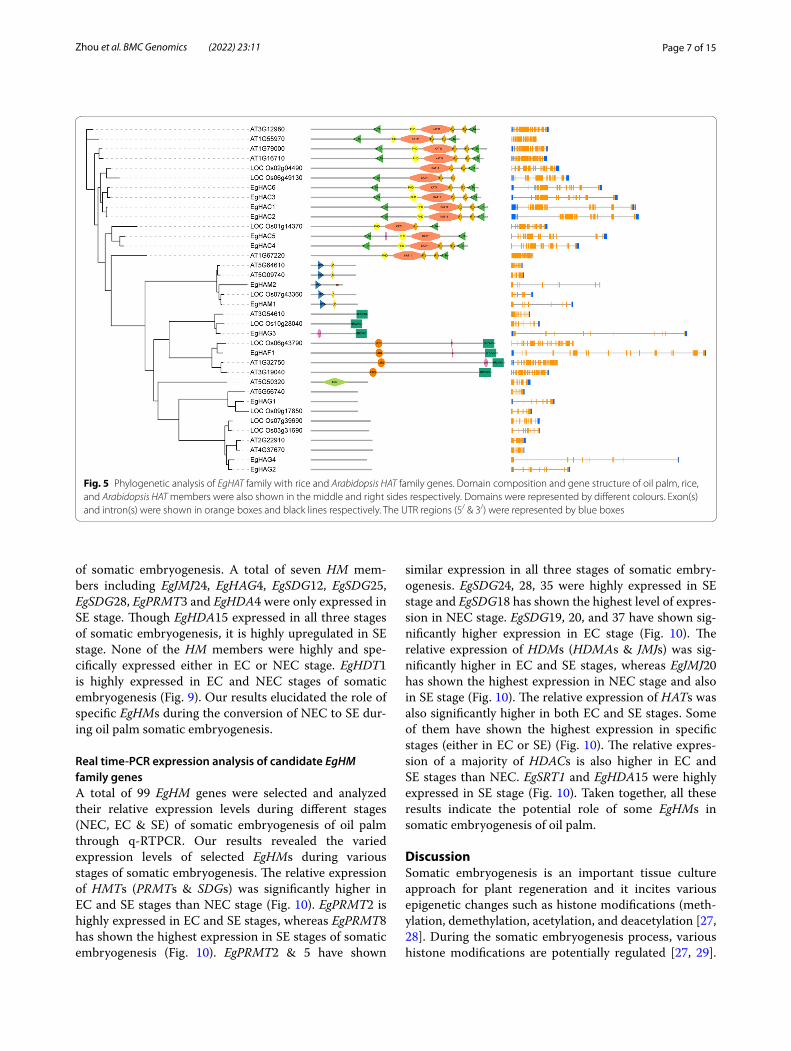
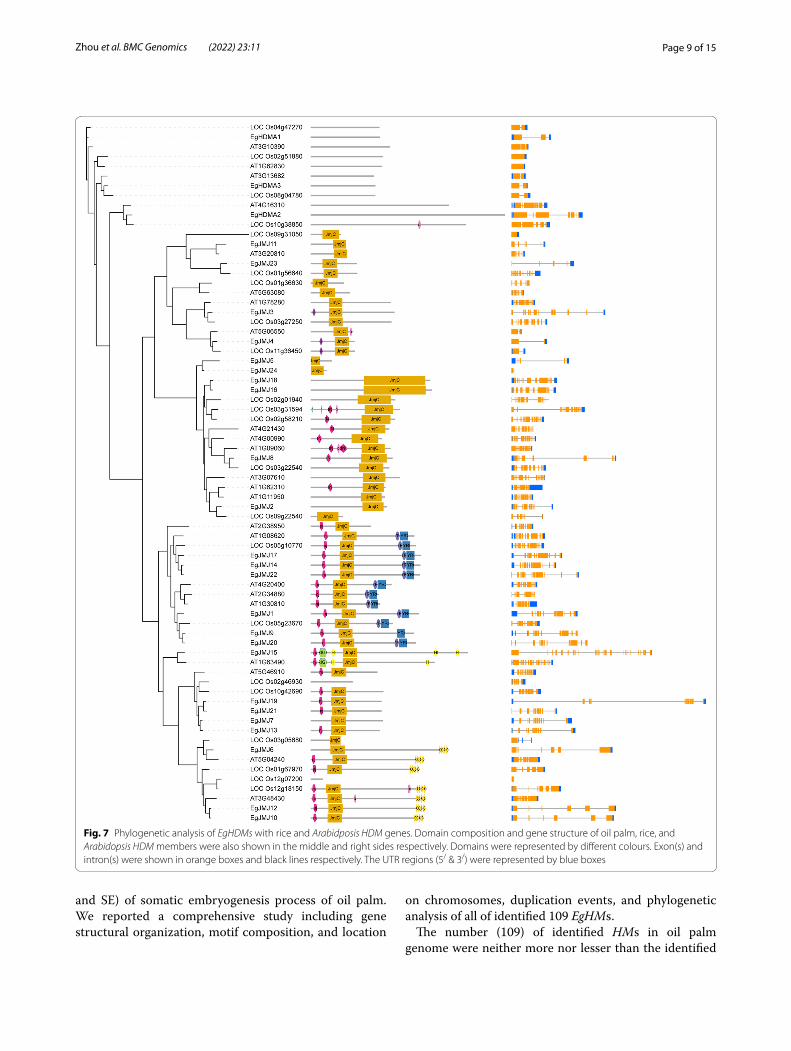
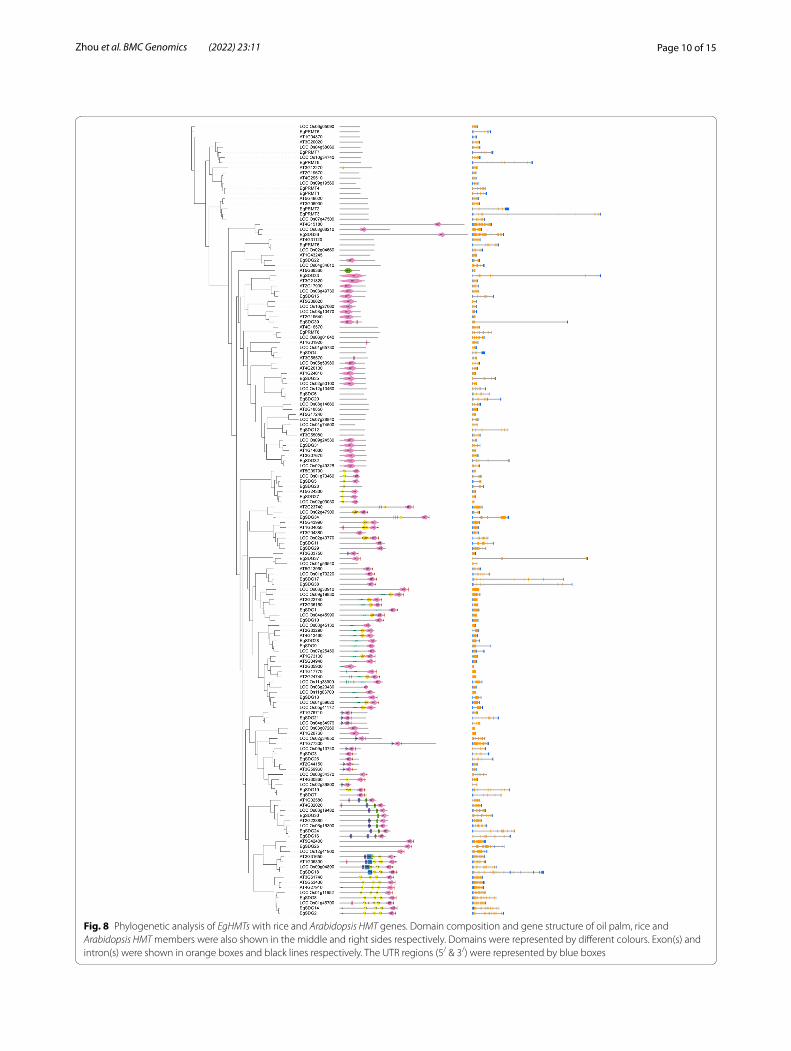
Phylogenetic analysis between oil palm, rice, and Arabidopsis HM gene familyTo elucidate the evolutionary relationship between oil palm, rice, Arabidopsis, we generated the rooted phy-logenetic trees for each HM gene family (HAT, HDAC, HDM, and HMT) (Figs. 5, 6, 7 and 8). The phylogenetic tree for subfamilies of each HM gene family was classi-fied and clustered into diverse trends. The phylogenetic tree of HAT family showed that all the HAG, HAM, HAC, and HAFs of oil palm, rice, and Arabidopsis were not clustered together and they clustered in a species-spe-cific manner and also in mixed type (Fig. 5). For HDAC family, HDTs and SRTs clustered in a species-specific manner, whereas HDAs clustered together (Fig. 6). The phylogenetic tree of HDMs (HDMAs and JMJs) also showed a different trend with a species-specific type of clustering (Fig. 7). The phylogenetic tree of HMT family revealed that all the SDGs and PRMTs genes were clus-tered together (Fig. 8). Altogether, our results indicated that there is a clear evolutionary relationship and diversi-fication between oil palm, rice, and Arabidopsis HM gene family members.
Expression levels of EgHMs in different somatic embryogenic stages (Embryogenic calli, Non‑embryogenic calli, and somatic embryos)Based on the available transcriptome data of oil palm EC, NEC, and SE (https:// www. ncbi. nlm. nih. gov/ biopr oject/ PRJNA 699335) stages were downloaded from the NCBI
Fig. 3 Chromosomal distribution of EgHM genes across 16 chromosomes of oil palm genome. Chromosome numbers are indicated on the top of each chromosome. The vertical greyscale on the left side represents the length of the oil palm chromosomes
Page 6 of 15Zhou et al. BMC Genomics (2022) 23:11
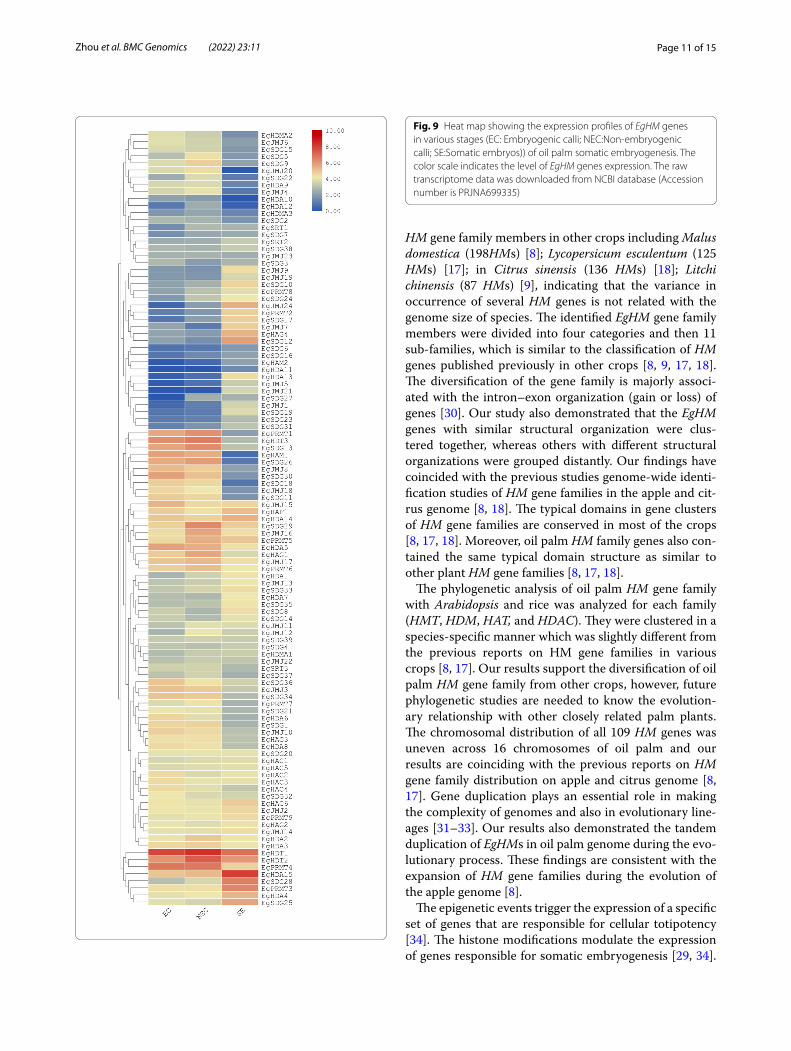
website and analyzed the expression levels of all identified 109 EgHM gene family members. The transcript abun-dance of EgHMs in various stages of oil palm somatic embryogenesis was analyzed by generating the heat-map with the help of FPKM values. As shown in Fig. 9, EgHM gene members showed differential expression in different stages of somatic embryogenesis of oil palm. However, most of the genes were down-regulated in all
three stages of somatic embryogenesis (Fig. 9). Moreover, six members of HMTs (EgSDG13, EgSDG26, EgSDG30, EgSDG34, EgPRMT1 and EgPRMT4), two members of HDMs (EgJMJ15 and EgJMJ17); on member of HATs (EgHAM1); and four members of HDACs(EgHDA5, EgHDT1, EgHDT2, EgHDT3) are expressed during both EC and NEC stages (Fig. 9). The HDAC genes EgHDT1 and EgHDT2 were showed expression in all three stages
Fig. 4 Gene duplication events and interchromosomal relationships between EgHM genes in oil palm genome. A total of 44 duplicated EgHM gene pairs were found across 16 chromosomes via MC ScanX tool and linked by the red lines inside the circle view. Each chromosomal block was represented with a different color
Page 7 of 15Zhou et al. BMC Genomics (2022) 23:11
of somatic embryogenesis. A total of seven HM mem-bers including EgJMJ24, EgHAG4, EgSDG12, EgSDG25, EgSDG28, EgPRMT3 and EgHDA4 were only expressed in SE stage. Though EgHDA15 expressed in all three stages of somatic embryogenesis, it is highly upregulated in SE stage. None of the HM members were highly and spe-cifically expressed either in EC or NEC stage. EgHDT1 is highly expressed in EC and NEC stages of somatic embryogenesis (Fig. 9). Our results elucidated the role of specific EgHMs during the conversion of NEC to SE dur-ing oil palm somatic embryogenesis.
Real time‑PCR expression analysis of candidate EgHM family genesA total of 99 EgHM genes were selected and analyzed their relative expression levels during different stages (NEC, EC & SE) of somatic embryogenesis of oil palm through q-RTPCR. Our results revealed the varied expression levels of selected EgHMs during various stages of somatic embryogenesis. The relative expression of HMTs (PRMTs & SDGs) was significantly higher in EC and SE stages than NEC stage (Fig. 10). EgPRMT2 is highly expressed in EC and SE stages, whereas EgPRMT8 has shown the highest expression in SE stages of somatic embryogenesis (Fig. 10). EgPRMT2 & 5 have shown
similar expression in all three stages of somatic embry-ogenesis. EgSDG24, 28, 35 were highly expressed in SE stage and EgSDG18 has shown the highest level of expres-sion in NEC stage. EgSDG19, 20, and 37 have shown sig-nificantly higher expression in EC stage (Fig. 10). The relative expression of HDMs (HDMAs & JMJs) was sig-nificantly higher in EC and SE stages, whereas EgJMJ20 has shown the highest expression in NEC stage and also in SE stage (Fig. 10). The relative expression of HATs was also significantly higher in both EC and SE stages. Some of them have shown the highest expression in specific stages (either in EC or SE) (Fig. 10). The relative expres-sion of a majority of HDACs is also higher in EC and SE stages than NEC. EgSRT1 and EgHDA15 were highly expressed in SE stage (Fig. 10). Taken together, all these results indicate the potential role of some EgHMs in somatic embryogenesis of oil palm.
DiscussionSomatic embryogenesis is an important tissue culture approach for plant regeneration and it incites various epigenetic changes such as histone modifications (meth-ylation, demethylation, acetylation, and deacetylation [27, 28]. During the somatic embryogenesis process, various histone modifications are potentially regulated [27, 29].
Fig. 5 Phylogenetic analysis of EgHAT family with rice and Arabidopsis HAT family genes. Domain composition and gene structure of oil palm, rice, and Arabidopsis HAT members were also shown in the middle and right sides respectively. Domains were represented by different colours. Exon(s) and intron(s) were shown in orange boxes and black lines respectively. The UTR regions (5/ & 3/) were represented by blue boxes

Page 8 of 15Zhou et al. BMC Genomics (2022) 23:11
The histone modifications activity leads to the altered gene expression of somatic embryogenesis process [27]. These histone modification changes regulated by vari-ous genes during somatic embryos potentially affect the development of somatic embryos. It’s necessary to know the information of various genes that regulate the his-tone modifications during somatic embryogenesis. To date, none of the studies provided brief information on histone modification genes of oil palm during somatic
embryogenesis process. To the best of our knowledge, this is the first report on the identification and character-ization of HM genes in oil palm during somatic embryo-genesis process. In our study, we identified a total of 109 HM genes including 48 of HMTs (histone methyltrans-ferases); 27 of HDMs (histone demethylases); 13 of HATs (histone acetylases), and 21 of HDACs (histone deacety-lases) through genome-wide analysis and analyzed their expression patterns during different stages (NEC, EC,
Fig. 6 Phylogenetic analysis of EgHDACs with rice and Arabidopsis HDAC genes. Domain composition and gene structure of oil palm, rice, and Arabidopsis HDAC members were also shown in the middle and right sides respectively. Domains were represented by different colours. Exon(s) and intron(s) were shown in orange boxes and black lines respectively. The UTR regions (5/ & 3/) were represented by blue boxes
Page 9 of 15Zhou et al. BMC Genomics (2022) 23:11
and SE) of somatic embryogenesis process of oil palm. We reported a comprehensive study including gene structural organization, motif composition, and location
on chromosomes, duplication events, and phylogenetic analysis of all of identified 109 EgHMs.
The number (109) of identified HMs in oil palm genome were neither more nor lesser than the identified
Fig. 7 Phylogenetic analysis of EgHDMs with rice and Arabidposis HDM genes. Domain composition and gene structure of oil palm, rice, and Arabidopsis HDM members were also shown in the middle and right sides respectively. Domains were represented by different colours. Exon(s) and intron(s) were shown in orange boxes and black lines respectively. The UTR regions (5/ & 3/) were represented by blue boxes
Page 10 of 15Zhou et al. BMC Genomics (2022) 23:11
Fig. 8 Phylogenetic analysis of EgHMTs with rice and Arabidopsis HMT genes. Domain composition and gene structure of oil palm, rice and Arabidopsis HMT members were also shown in the middle and right sides respectively. Domains were represented by different colours. Exon(s) and intron(s) were shown in orange boxes and black lines respectively. The UTR regions (5/ & 3/) were represented by blue boxes
Page 11 of 15Zhou et al. BMC Genomics (2022) 23:11
HM gene family members in other crops including Malus domestica (198HMs) [8]; Lycopersicum esculentum (125 HMs) [17]; in Citrus sinensis (136 HMs) [18]; Litchi chinensis (87 HMs) [9], indicating that the variance in occurrence of several HM genes is not related with the genome size of species. The identified EgHM gene family members were divided into four categories and then 11 sub-families, which is similar to the classification of HM genes published previously in other crops [8, 9, 17, 18]. The diversification of the gene family is majorly associ-ated with the intron–exon organization (gain or loss) of genes [30]. Our study also demonstrated that the EgHM genes with similar structural organization were clus-tered together, whereas others with different structural organizations were grouped distantly. Our findings have coincided with the previous studies genome-wide identi-fication studies of HM gene families in the apple and cit-rus genome [8, 18]. The typical domains in gene clusters of HM gene families are conserved in most of the crops [8, 17, 18]. Moreover, oil palm HM family genes also con-tained the same typical domain structure as similar to other plant HM gene families [8, 17, 18].
The phylogenetic analysis of oil palm HM gene family with Arabidopsis and rice was analyzed for each family (HMT, HDM, HAT, and HDAC). They were clustered in a species-specific manner which was slightly different from the previous reports on HM gene families in various crops [8, 17]. Our results support the diversification of oil palm HM gene family from other crops, however, future phylogenetic studies are needed to know the evolution-ary relationship with other closely related palm plants. The chromosomal distribution of all 109 HM genes was uneven across 16 chromosomes of oil palm and our results are coinciding with the previous reports on HM gene family distribution on apple and citrus genome [8, 17]. Gene duplication plays an essential role in making the complexity of genomes and also in evolutionary line-ages [31–33]. Our results also demonstrated the tandem duplication of EgHMs in oil palm genome during the evo-lutionary process. These findings are consistent with the expansion of HM gene families during the evolution of the apple genome [8].
The epigenetic events trigger the expression of a specific set of genes that are responsible for cellular totipotency [34]. The histone modifications modulate the expression of genes responsible for somatic embryogenesis [29, 34].
Fig. 9 Heat map showing the expression profiles of EgHM genes in various stages (EC: Embryogenic calli; NEC:Non-embryogenic calli; SE:Somatic embryos)) of oil palm somatic embryogenesis. The color scale indicates the level of EgHM genes expression. The raw transcriptome data was downloaded from NCBI database (Accession number is PRJNA699335)

Page 12 of 15Zhou et al. BMC Genomics (2022) 23:11
In our study, several genes encoding histone methyla-tion, demethylation, histone acetylation, and histone dea-cetylation were identified through genome-wide study and analyzed their expression analysis during different stages (NEC, EC & SE) of somatic embryogenesis. In this study, several EgHMs displayed their differential expres-sion between different stages of somatic embryogenesis of oil palm as shown in Fig. 10. Based on qPCR data, EgPRMT2, EgPRMT3, EgPRMT5, EgPRMT6, EgSDG1, EgSDG3, EgSDG15, EgSDG16, EgSDG21, EgSDG23, EgSDG26, EgSDG27, EgSDG33, EgHDMA2, EgJMJ2, EgJMJ5, EgJMJ6, EgJMJ8, EgJMJ12, EgJMJ13, EgJMJ19, EgJMJ24, EgHAM1, EgHAC3, EgHAC5, EgHAC6, EgHAG1, EgHAG2, EgHAG3, EgHDT1, EgHDT2, EgHDT3, EgHDA11 & EgHDA14 showed their expression levels during both embryogenic calli and somatic embryo stages, revealed their prominent role in somatic embryo-genesis. These identified EgHMs may provide insightful information to elucidate the molecular mechanisms asso-ciated with the somatic embryogenesis of oil palm.
Material and methodsEgHMs identification in oil palm genomeThe pfam database was used to identify the EgHM gene family members using the Hidden Markov Model pro-files of published IDs of each type (HMT, HDM, HAT & HDAC) of HM genes [17, 18]. The pfam IDs belongs each
type of HM genes were used as a query to search for HM gene members in oil palm genome database [35] using HMMER3.0 tool. Further, we also retrieved the unavail-able sequences of EgHMs using the known HM gene sequences of Arabidopsis (http:// www. arabi dopsis. org/) and Oryza sativa (http:// rice. plant biolo gy. msu. edu/) through blast search in oil palm genome database (http:// palmx plore. mpob. gov. my/ palmX plore/).
We also predicted the coding sequence length of each HM gene family member using Blastn search against oil palm genome database. The identified putative oil palm HMs genes, such as HMTs (SDGs and PRMTs), HDMs (HDMAs and JMJs), HATs (HAGs, HAMs, HACs, and HAFs), and HDACs (HDAs, SRTs, and HDTs) were identified finally based on the highly conserved domains. Additionally, the M.wt, pI values of oil palm HM gene members were also determined with the help of ExPASy (https:// web. expasy. org/ compu te_ pi/). The online tool “CELLO” (http:// cello. life. nctu. edu. tw/) was further used to predict the subcellular localization of all oil palm HM gene family members.
Gene structure, Conserved motifs analysis of EgHMsThe 109 EgHM genes structural analysis (intron–exon organization) was analyzed by the Gene Structure Dis-play Server (http:// gsds. cbi. pku. edu. cn/). The conserved motifs analysis of all identified EgHM proteins were
Fig. 10 Relative expression analysis of 99 EgHM candidate genes in various stages of somatic embryogenesis in oil palm. The X-axis represents different stages (SE, Somatic embryos; NEC, non-embryogenic calli; EC, embryogenic calli); Y-axis represents relative expression of specific EgHM gene. Data represent the mean ± SE of three replicates. Asterisks represent significant differences at P ≤ 0.05(*) and P ≤ 0.01(**)

Page 13 of 15Zhou et al. BMC Genomics (2022) 23:11
investigated by MEME tool (http:// meme- suite. org/ tools/ meme).
EgHM gene duplications, phylogenetic relationships, and their distribution on chromosomesWe explored the duplications of 109 EgHM gene family members in oil palm genome using the MCScanX tool with default parameters [36, 37]. Further, we mapped the location of all 109 EgHMs across 16 chromosomes of oil palm from the available genome database of oil palm. We also generated the phylogenetic tree for each type (HMT, HDM, HAT & HDAC) of oil palm HMs with HM genes Arabidopsis and Oryza sativa using MEGA 7.0 [38] by Maximum Likelihood method, with a bootstrap value of 1000 replications.
Plant materialsThe Oil palm (Elaeis guineensis; pisifera, thin-shelled African oil palm) plants were grown in the coconut field of Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences, Wenchang, China. All the plants were grown under institutional regulatory issues. All the plant materials are collected by the correspond-ing author of this research work. The immature zygotic embryos were selected as explants for oil palm tissue culture and followed the method as described by Silva et al. (2012) [39] with few modifications. The calli (non-embryogenic or embryogenic) was induced from imma-ture zygotic embryos on callus induction media (CIM).The explants were cultured on CIM: (1/2 MS medium supplemented with 30 mg/l picloram, 100 mg/l casein hydrolysate, 500 mg/l L-glutamine, 200 mg/l aspargine, 200 mg/l arginine, 2 mg/l glycine, 100 mg/l adenine sulfate, 100 mg/l citric acid, 100 mg/l ascorbic acid, 30,000 mg/l sucrose, and 3,000 mg/l Phytagel). The EC and NEC were induced after 3 months of repeated sub-cultures. The Somatic embryogenesis (SE) was induced by transferring EC to somatic embryo induction medium (SIM: CIM without picloram). The somatic embryo was induced in SIM after four months of culture. The EC, NEC, and SEs (torpedo) were collected at their stages and then quickly frozen in liquid nitrogen followed by the storage of -70 °C for subsequent RNA extraction.
EgHMs expression analysis from available transcriptome dataWe downloaded the transcriptome data of oil palm somatic embryogenesis stages (EC, NEC, and SE; Acces-sion number: PRJNA699335) from the Sequence Read Archive (SRA) database of NCBI. The heatmap (http:// www. omics hare. com/ tools) was generated to calculate EgHMs expression levels in three different stages of
somatic embryogenesis using RPKM values (RPKM= 10
6C
NL/
103 [32].
EgHMs gene expression analysis by quantitative real‑time PCRTotal RNA was extracted from the different stages of calli (NEC, EC, and SE) during somatic embryogenesis using RNAprep pure Plant Kit (Tiangen, Beijing, China) by fol-lowing the manufacturer’s instructions. The first-strand cDNA was synthesized using EasyScript® First-Strand cDNA Synthesis SuperMix (TransGen, Beijing, China) kit. The relative expression analysis of the 99 EgHM genes from the 109 identified HM genes was carried out by using the real-time qPCR method. The real-time qPCR was performed with SYBR® Select Master Mix (Thermo Fisher Scientific, Waltham, USA). The ABI QuantS-tudio™6 Flex quantitative real-time PCR instrument (Thermo Fisher, Waltham, USA) was used to analyze the data. The relative expression levels of 99 HM genes at different stages of somatic embryogenesis were calcu-lated via 2−ΔΔCt method. The qPCR primers for analyz-ing the expression of 99 HM genes were designed using QuantPrime qPCR primer designing tool (https:// quant prime. mpimp- golm. mpg. de/) and listed in Supplemen-tary Table 3. All the qPCR reactions were performed with three biological and three technical replications. The oil palm actin gene (EgActin1) was used as an internal con-trol to check the expression of EgHMs. The statistical sig-nificance at p < 0.05 was determined by using One-Way ANOVA.
ConclusionsIn conclusion, this is the first report on genome-wide analysis of histone modification genes (HMT, HDM, HAT, and HDAC) in oil palm. From this study, a total of 109 EgHMs were identified and analyzed candidate genes expression patterns during somatic embryogenesis of oil palm. Moreover, comprehensive information regarding their gene structure, motif, composition, chromosomal distribution, duplication events, evolutionary relation-ship with rice, and Arabidopsis was also reported. Fur-thermore, differential expression of various EgHMs at different stages of somatic embryogenesis was also elu-cidated through real-time PCR analysis indicating their potential involvement during the somatic embryogenesis of oil palm.
Supplementary InformationThe online version contains supplementary material available at https:// doi. org/ 10. 1186/ s12864- 021- 08245-2.
Additional file 1: Table 1. List of EgHMs in oil palm genome.

Page 14 of 15Zhou et al. BMC Genomics (2022) 23:11
Additional file 2: Table 2. Subcellular localaization prediction of 109 EgHM gene family.
Additional file 3: Table 3. List of qRT-PCR primer pairs used for realtive expression of EgHM gene family members in oil palm during somatic embryogenesis.
AcknowledgementsWe thank Mengdi Yang for her help during this research work.
Authors’ contributionsLZ assessed the conceptualization of research. LZ and RY conceived and designed the methodology. LZ, RY, LJ, YY, HC, and ZZ analyzed the data. RY prepared and wrote original manuscript draft. LZ supervised the research. All authors have read and approved.
FundingThis research work was financially supported by the National Natural Science Foundation of China (No: 31870670) and also basic support from the Chinese Academy of Tropical Agricultural Sciences, China.
Availability of data and materialsThe transcriptome data used in this study are available in the NCBI Sequence Read Archive (SRA) with BioProject accession number PRJNA699335. All data sets analyzed during this research are provided as Supplementary files and also included in the article. The data sets generated during this study are also available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participateAll methods in this research were carried out in accordance with relevant guidelines and regulations of Coconut Research Institute, Wenchang, China.
Consent for publicationNot applicable
Competing interestsThe authors declare that they have no competing interest.
Received: 18 August 2021 Accepted: 7 December 2021
References 1. Zhang X. The epigenetic landscape of plants Sci. 2008;320:489–92. 2. Klose RJ, Zhang Y. Regulation of histone methylation by demethylimina-
tion and demethylation. Nat Rev Mol Cell Biol. 2007;8:307–18. 3. Aquea F, Vega A, Timmermann T, Poupin MJ, Arce-Johnson P. Genome-
wide analysis of the SET DOMAIN GROUP family in grapevine. Plant Cell Rep. 2011;30:1087–97.
4. Hollender C, Liu Z. Histone deacetylase genes in Arabidopsis develop-ment. J Int Plant Biol. 2008;50:875–85.
5. Jiang D, Yang W, He Y, Amasino RM. Arabidopsis relatives of the human lysine-specific Demethylase1 repress the expression of FWA and FLOWERING LOCUS C and thus promote the floral transition. Plant Cell. 2007;19:2975–87.
6. Pandey R, Muller A, Napoli CA, Selinger DA, Pikaard CS, Richards EJ, Bender J, Mount DW, Jorgensen RA. Analysis of histone acetyltrans-ferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res. 2002;30:5036–55.
7. Feng Q, Wang HB, Ng HH, Erdjument-Bromage H, Tempst P, Struhl K, Zhang Y. Methylation of H3-lysine 79 is mediated by a new family of HMTases without a SET domain. Curr Biol. 2002;12:1052–8.
8. Fan S, Wang J, Lei C. Identification and characterization of histone modification gene family reveal their critical responses to flower
induction in apple. BMC Plant Biol. 2018;18:173. https:// doi. org/ 10. 1186/ s12870- 018- 1388-0.
9. Peng M, Ying P, Liu X, Li C, Xia R, Li J, Zhao M. Genome-Wide Identification of Histone Modifiers and Their Expression Patterns during Fruit Abscission in Litchi. Front Plant Sci. 2017;8:639.
10. Ahmad A, Cao X. Plant PRMTs broaden the scope of arginine methylation. J Genet Genomics. 2012;39:195–208.
11. Dong G, Ma DP, Li J. The histone methyltransferase SDG8 regulates shoot branching in Arabidopsis. Biochem Biophys Res Commun. 2008;373:659–64.
12. Tian L, Fong MP, Wang JJ, Wei NE, Jiang H, Doerge RW, Chen ZJ. Reversible histone acetylation anddeacetylation mediate genome-wide, promoter-dependent and locus-specific changes in gene expression during plant development. Genetics. 2005;169:337–45.
13. Sheldon CC, Finnegan EJ, Dennis ES, Peacock WJ. Quantitative effects of vernalization on FLC and SOC1 expression. Plant J. 2006;45:871–83.
14. Chen ZJ, Tian L. Roles of dynamic and reversible histone acetyla-tion in plant development and polyploidy. Biochim Biophys Acta. 2007;1769:295–307.
15. Ma X, Lv S, Zhang C, Yang C. Histone deacetylases and their functions in plants. Plant Cell Rep. 2013;32:465–78.
16. Wang Z, Cao H, Chen F, Liu Y. The roles of histone acetylation in seed performance and plant development. Plant Physiol Biochem. 2014;84:125–33.
17. Cigliano RA, Sanseverino W, Cremona G, Ercolano MR, Conicella C, Con-siglio FM. Genome-wide analysis of histone modifiers in tomato: gaining an insight into their developmental roles. BMC Genomics. 2013;14:57.
18. Xu J, Xu H, Liu Y, Wang X, Xu Q, Deng X. Genome-wide identification of sweet orange (Citrus sinensis) histone modification gene families and their expression analysis during the fruit development and fruit-blue mold infection process. Front Plant Sci. 2015;6:607.
19. Barcelos E, Rios SA, Cunha RNV, Lopes R, Motoike SY, Babiychuk E, Skirycz A, Kushnir S. Oil palm natural diversity and the potential for yield improvement. Front Plant Sci. 2015;6:190. https:// doi. org/ 10. 3389/ fpls. 2015. 00190.
20. Corley RHV. How much palm oil do we need? Environ Sci Policy. 2019;12:134–9. https:// doi. org/ 10. 1016/j. envsci. 2008. 10. 011.
21. Cui J, Lamade E, Tcherkez G. Seed Germination in Oil Palm (Elaeis guineen-sis Jacq.): A review of metabolic pathways and control mechanisms. Int J Mol Sci. 2020;21(12):4227.
22. Weckx S, Inzé D, Maene L. Tissue culture of oil palm: finding the balance between mass propagation and somaclonal variation. Front Plant Sci. 2019;10:722.
23. Yarra R, Jin L, Zhao Z, Cao H. Progress in tissue culture and genetic trans-formation of Oil Palm: An Overview. Int J Mol Sci. 2019;20(21):5353.
24. Elhiti M, Stasolla C. Somatic embryogenesis: the molecular network regu-lating embryo formation. Somatic embryogenesis in ornamentals and its applications. Springer, Berlin. 2016;12:217–29.
25. Smertenko A, Bozhkov PV. Somatic embryogenesis: life and death pro-cesses during apical–basal patterning. J Exp Bot. 2014;65:1343–60.
26. Wang QM, Wang L. An evolutionary view of plant tissue culture: soma-clonal variation and selection. Plant Cell Rep. 2012;31:1535–47.
27. Li HL, Guo D, Zhu JH, Wang Y, Peng SQ. Identification of histone methyla-tion modifiers and their expression patterns during somatic embryogen-esis in Hevea brasiliensis. Genet Mol Biol. 2020;43(1): e20180141. https:// doi. org/ 10. 1590/ 1678- 4685- GMB- 2018- 0141.
28. Yakovlev IA, Carneros E, Lee Y. Transcriptional profiling of epigenetic regulators in somatic embryos during temperature induced formation of an epigenetic memory in Norway spruce. Planta. 2016;243:1237–49.
29. De-la-Peña C, Nic-Can GI, Galaz-Ávalos RM, Avilez-Montalvo R, Loyola-Vargas VM. The role of chromatin modifications in somatic embryogen-esis in plants. Front Plant Sci. 2015;6:635.
30. Xu GX, Guo CC, Shan HY, Kong HZ. Divergence of duplicate genes in exon-intron structure. P Natl Acad Sci USA. 2012;109(4):1187–92.
31. Si W, Hang T, Guo M, Chen Z, Liang Q, Gu L, Ding T. Whole-genome and transposed duplication contributes to the expansion and diversification of TLC genes in maize. Int J Mol Sci. 2019;20(21):5484.
32. Flagel LE, Wendel JF. Gene duplication and evolutionary novelty in plants. New Phytol. 2009;183(3):557–64.
33. Moore RC, Purugganan MD. The evolutionary dynamics of plant duplicate genes. Curr Opin Plant Bio. 2005;l8(2):122–8.

Page 15 of 15Zhou et al. BMC Genomics (2022) 23:11
• fast, convenient online submission
•
thorough peer review by experienced researchers in your field
• rapid publication on acceptance
• support for research data, including large and complex data types
•
gold Open Access which fosters wider collaboration and increased citations
maximum visibility for your research: over 100M website views per year •
At BMC, research is always in progress.
Learn more biomedcentral.com/submissions
Ready to submit your researchReady to submit your research ? Choose BMC and benefit from: ? Choose BMC and benefit from:
34. Pasternak T, Dudits D. Epigenetic Clues to Better Understanding of the Asexual Embryogenesis in planta and in vitro. Front Plant Sci. 2019;10:778. https:// doi. org/ 10. 3389/ fpls. 2019. 00778.
35. Sanusi NSNM, Rosli R, Halim MAA, Chan KL, Nagappan J, Azizi N, Amir-uddin N, Tatarinova TV, Low EL. PalmXplore: oil palm gene database. Database (Oxford). 2018;2018:bay095. https:// doi. org/ 10. 1093/ datab ase/ bay095.
36. Zhou LX, Yarra R. Genome-wide identification and expression analy-sis of bZIP transcription factors in oil palm (Elaeis guineensis Jacq.) under abiotic stress. Protoplasma 2021, Jul 1. https:// doi. org/ 10. 1007/ s00709- 021- 01666-6
37. Wang Y, Tang H, DeBarry JD, Tan X, Li J, Wang X, Lee T, Jin H, Marler B, Guo H, Kissinger JC, Paterson AH. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucl Acids Res. 2012;40(7):e49.
38. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
39. Silva RC, Luis ZG, Scherwinski-Pereira JE. Differential responses to somatic embryogenesis of different genotypes of Brazilian oil palm (Elaeis guineensis Jacq.). Plant Cell Tiss Org Cult. 2012;111:59–67.
Publisher’s NoteSpringer Nature remains neutral with regard to jurisdictional claims in pub-lished maps and institutional affiliations.





























