Embed Size (px)
Citation preview

Imaging Circuit Formation in Zebrafish
Nikolas Nikolaou, Martin P. Meyer
MRC Centre for Developmental Neurobiology, King’s College London, Guy’s Hospital Campus,London SE1 1UL, United Kingdom
Received 25 August 2010; accepted 9 January 2011
ABSTRACT: The study of nervous system develop-
ment has been greatly facilitated by recent advances in
molecular biology and imaging techniques. These
approaches are perfectly suited to young transparent
zebrafish where they have allowed direct observation of
neural circuit assembly in vivo. In this review we will
highlight a number of key studies that have applied opti-
cal and genetic techniques in zebrafish to address ques-
tions relating to axonal and dendritic arbor develop-
ment, synapse assembly and neural plasticity. These
studies have revealed novel cellular phenomena and
modes of growth that may reflect general principles
governing the assembly of neural circuits. ' 2011 Wiley
Periodicals, Inc. Develop Neurobiol 72: 346–357, 2012
Keywords: zebrafish; imaging; synapse; dendrite; axon;
circuit; plasticity; molecular biology
INTRODUCTION
Perception and behavior are dependent on the estab-
lishment of precise synaptic connections within the
developing nervous system. Two fundamental goals
in developmental neuroscience are to understand how
neurons target one another during periods of synapto-
genesis, and how synapses themselves are assembled.
The structure of developing neurons is also highly
plastic. How does the structural plasticity of individ-
ual neurons relate to circuit assembly, and how is
such plasticity regulated? Recent advances in
imaging technology, molecular biology, and the GFP
revolution have made live imaging the tool of choice
for studying circuit assembly and plasticity (Niell and
Smith, 2004; Lichtman and Smith, 2008). These tech-
nological advances have enabled researches to label
and follow over time virtually any feature of the nerv-
ous system from populations of neurons (Godinho
et al., 2005; Mumm et al., 2006), to dendrites and
axons of individual neurons (O’Rourke et al., 1994;
Jontes et al., 2000; Trachtenberg et al., 2002; Bishop
et al., 2004; Haas et al., 2006), to subcellular struc-
tures within neurons such as dendritic spines (Trach-
tenberg et al., 2002; Bishop et al., 2004; Keck et al.,
2008; Yang et al., 2009) and synaptic vesicles (Meyer
and Smith, 2006; Ruthazer et al., 2006). Live imaging
of these structures during development has revealed
new cellular phenomena and modes of growth, and
has generated hypotheses on the mechanisms of cir-
cuit assembly and plasticity. In this review we
describe the features of zebrafish as a model organism
that make it ideally suited to take advantage of these
technological developments. Rather than provide a
comprehensive review of the literature we have
focused on a few key studies that illustrate how live
imaging in zebrafish has enabled researchers to
address key questions in the field. We will finish by
discussing how the latest generation of optogenetic
tools and their use in zebrafish are likely to advance
our understanding of circuit formation, function, and
plasticity.
Correspondence to:M.P. Meyer ([email protected]).Contract grant sponsor: Medical Research Council; contract
grant number: G0801242.Contract grant sponsor: Medical Research Council Career Devel-
opment; contract grant number: G0600107.
' 2011 Wiley Periodicals, Inc.Published online 19 January 2011 in Wiley Online Library(wileyonlinelibrary.com).DOI 10.1002/dneu.20874
346

Arbor Growth and Synapse Formation inthe Retinotectal Projection: A Process ofTrial and Error
A crucial step in establishing synaptic connectivity is
the elaboration of axonal and dendritic arbors. Arbor
size, shape, and position determine patterns of synap-
tic connectivity, and hence the functional properties
of neural circuits. The visual system is a prominent
model system for studying how arbor morphogenesis
underlies the formation of precise neuronal circuitry.
Connections in the visual system are topographically
organized. That is, neighboring retinal ganglion cells
(RGCs) project to neighboring neurons in the optic
tectum, the main RGC target in the zebrafish brain.
Targeting of RGC axons to correct termination zones
within the tectum, and restriction of arbor size within
the termination zone are crucial for generating pre-
cise topography. Of equal importance is the develop-
ment of dendritic arbors in postsynaptic tectal cells,
since these determine how RGC inputs to the brain
are sampled, and ultimately how visual information is
processed.
A number of key features make the zebrafish reti-
notectal projection particularly amenable to live
imaging studies. First, zebrafish embryos and larvae
are translucent, develop externally, and RGC axons
and tectal cell dendrites arborize just beneath the skin
overlying the optic tectum. These arbors are therefore
easily accessible to imaging without the need for sur-
gery, even with a standard confocal microscope. Sec-
ond, development of the visual system is rapid. The
first RGC axons reach the tectum between 2 and 3
days post-fertilization (dpf), and visually evoked
behaviors can be elicited soon thereafter (Easter and
Nicola, 1996). Indeed the bulk of RGC axon and tec-
tal dendritic arbor growth is complete by 6dpf (Niell
et al., 2004; Meyer and Smith, 2006). Thus, in one
over-night time-lapse imaging session a considerable
degree of arbor development can be observed. Third,
injection of plasmids into single cell stage embryos
results in transient mosaic expression of transgenes at
larval stages of development. This versatile and
straightforward technique has been exploited exten-
sively to achieve Golgi-like labeling of single neu-
rons with fluorescent proteins in vivo, and has
enabled fine details such as dendritic filopodia to be
resolved and followed over time using confocal or
two photon microscopy. Live in vivo imaging of
RGC axonal and tectal cell dendritic arbors in zebra-
fish has revealed dynamic aspects of arbor growth
that would be missed from analysis of static images.
For example, when presented with static images of a
developing axonal or dendritic arbor one would
assume that these arbors grow much like a tree, with
continuous elongation of branches, and a gradual
increase in arbor complexity as new branches are
added. Time-lapse imaging in zebrafish has shown
that the process of RGC axonal and tectal dendritic
arbor growth is much more dynamic. During develop-
mental periods growing axonal and dendritic arbors
are continually remodeled by the concurrent exten-
sion and retraction of filopodia, only a small fraction
of which are maintained as branches in the mature
arbor (Kaethner and Stuermer, 1992, 1997; Niell
et al., 2004; Meyer and Smith, 2006). Thus, time-
lapse imaging has shown that arbor development is
more a process of trial-and-error rather than simple
forward monotonic growth.
How is the structural plasticity of developing
axons and dendrites related to circuit formation? By
imaging RGC axons and tectal cell dendrites express-
ing fluorescent pre- and postsynaptic marker proteins,
respectively, it was found that the dynamic turnover
of axonal and dendritic branches is accompanied by
the turnover of many nascent synapses (Niell et al.,
2004; Meyer and Smith, 2006) (see Fig. 1). Thus, the
dynamic rearrangement of arbors serves to sample
many potential synaptic partners, and generates a
large pool of nascent synaptic connections, of which
only a small number are kept. Importantly, the pro-
cess of arbor growth and synapse formation appear to
be mechanistically linked. Maintenance of a synapse
stabilizes the axonal or dendritic filopodium on which
it is located and stabilized filopodia mature into
branches. Synapses are also the sites of new branch
addition. Imaging with high temporal resolution over
many hours has shown that axonal and dendritic
arbors grow by iterative rounds of filopodium exten-
sion from, and selective stabilization by synapses
(Niell et al., 2004; Meyer and Smith, 2006). These
studies suggest that by modulating two distinct
aspects of arbor growth—branch formation and
branch stability, the process of synapse formation
itself could guide growth of axonal and dendritic
arbors. Modeling studies have suggested that such a
\synaptotropic" mechanism can generate specific
dendritic arborization patterns independently of den-
dritic guidance molecules or genetically predeter-
mined branching programs (Niell, 2006; see Fig. 2).
Instead, formation of a correctly shaped and posi-
tioned dendrite depends solely on the distribution of
presynaptic partners, and on a local stochastic process
of branch extension and selective stabilization. In
other words, the extent and complexity of a dendritic
arbor will arise as a consequence of local interactions
with neighboring axons. Doesn’t this just offload the
Imaging Circuit Formation in Zebrafish 347
Developmental Neurobiology

task of circuit patterning onto the precise positioning
of axons? To a certain extent the answer is yes, but
since in the retinotectal projection a synaptotropic
mechanism also operates in presynaptic RGCs, the
initial targeting of axons need only be approximate.
A synaptotropic mechanism operating in both pre-
and postsynaptic neurons could facilitate self-organi-
zation of precise circuitry from only approximately
positioned axons. Indeed, the prevailing view in the
field of visual map development is that genetically
encoded axon guidance cues establish crude retino-
topic positioning of axons that are then refined by
neural activity (Ruthazer and Cline, 2004; Huberman
et al., 2008). It is therefore possible that activity at
nascent synapses modulates both axon branch stabil-
ity and branch formation resulting in a tight control
of axonal arbor structure, and hence map refinement.
In the following section we will describe how imag-
ing in zebrafish has been used to ask precisely how
neural activity modulates the structural development
of RGC axons.
Regulation of Axonal Arbor Growth byNeural Activity
Perturbations in neural activity followed by studies of
fixed tissue can show the effect of the perturbation on
final arbor form. Live imaging can go one step further
by revealing precisely how activity in a growing neu-
ron affects the process of arbor growth. For example,
time-lapse imaging can reveal whether an increase in
the number of branches arises through an increased
rate of branch formation or branch stabilization. Two
recent studies have used molecular genetic
approaches to suppress synaptic transmission in
RGCs in zebrafish and then used time-lapse imaging
to examine the effect on axon arbor development. In
the first study by Hua et al., synaptic transmission in
single RGCs was suppressed by expression of a mu-
tant form of the synaptic vesicle protein, VAMP2/
Synaptobrevin 2 (Hua et al., 2005). Time-lapse imag-
ing revealed that singly silenced axons formed fewer
branches than active ones, and as a consequence
Figure 1 Series of images from time-lapse imaging of axonal and dendritic arbor growth and
synapse formation in the zebrafish tectum. (A) The growth of an RGC axon arbor occurs by an iter-
ative sequence of presynaptic punctum (green) formation and filopodial (red) stabilization. A filo-
podium (arrow) is seen to extend and nascent puncta (arrowheads) are formed soon after. The
selective maintenance of these synapses lead to stabilization of the filopodium which matures into
a stable branch. Two other filopodia are formed and eliminated in close proximity. Scale bar ¼2 lm. (B) Similarly, the dendritic arbor growth of tectal cells is characterized by the presence of
many transient filopodia (red) and postsynaptic puncta (green), indicated by arrows and arrow-
heads, respectively. Scale bar ¼ 5 lm. Image in (A) reproduced from Meyer and Smith (2006)
with permission (Copyright 2006 by the Society for Neuroscience), and image in (B) reproduced
from Niell et al. (2004) with permission (Copyright 2004 by Nature Publishing Group).
348 Nikolaou and Meyer
Developmental Neurobiology

developed smaller arbors. Using similar techniques
Ben Fredj et al., suppressed synaptic transmission in
single RGCs using targeted expression of tetanus
toxin light-chain fused to EGFP (TeNT-Lc-EGFP)
(Ben Fredj et al., 2010). In contrast to the findings of
Hua et al., single RGC axons silenced by TeNT-Lc-
EGFP expression were larger than wild-type axons,
and time-lapse analysis revealed a failure of these
axons to arrest branch formation events. However,
both studies found that phenotypes were only appa-
rent when single RGCs were silenced in a field of
active ones. Widespread suppression of activity in
RGCs using either the mutant form of VAMP2 or
TeNT-Lc-EGFP resulted in arbors that were of nor-
mal size. Thus, relative activity levels or activity-de-
pendent competition between RGCs appear to regu-
late the size of territory occupied by axonal arbors. A
plausible explanation that could account for the dif-
ferent outcomes of single cell silencing is that the
degree of presynaptic suppression achieved with
the two approaches is different. Over-expression of
the VAMP2 mutant has been shown to suppress
stimulus-evoked vesicular release but does not
abolish it entirely (Sorensen et al., 2002), whereas
TeNT-Lc-EGFP is highly effective at blocking both
stimulus evoked and spontaneous release of synap-
tic vesicles. The weak residual activity in RGCs
expressing the VAMP2 mutant could therefore
induce plasticity mechanisms analogous to long-
term depression (LTD), which in the developing
retinotectal projection may lead to synapse elimina-
tion. Since synapses appear to be correlated with
both stabilization and formation of axonal branches
(Alsina et al., 2001; Meyer and Smith, 2006; Ruth-
azer et al., 2006), synapse elimination would lead
to an increased rate of branch elimination, a
reduced rate of branch formation and hence smaller
arbors. LTD, by definition, only occurs at synapses
with at least some activity. Therefore, in axons that
express TeNT-Lc-EGFP, where synaptic activity is
almost entirely abolished, LTD and synapse elimi-
nation are not induced, permitting branches to
extend beyond their normal territory. In this view,
competition punishes the weakly active axon more
than the completely silent one. While this possibil-
ity has not been explored directly, the contrasting
findings of these two studies suggest that different
manipulations of synaptic function could have very
different consequences for the growth of RGC
axons. To complicate matters further, other manipu-
lations that alter synaptic function have resulted in
non-competitive structural phenotypes. For exam-
ple, the blumenkohl (blu) mutant, initially identified
in a screen for zebrafish retinotectal projection
defects, is characterized by enlarged termination
zones and defasciculation of RGC axons (Baier et
al., 1996). The blu mutation maps to vglut2a, a
member of the vesicular glutamate transporter fam-
Figure 2 Synaptogenesis guides the growth and branching of dendritic arbors. Modeling the
sequence of events during synaptotropic dendritic growth in a situation where there is a non-homo-
geneous distribution of axons that are, receptive to synapse formation (dashed green boxes). Many
filopodia are extended (red), but only those which form synapses (green) are maintained as stable
branches (black). As a result, dendritic arbor growth is restricted to regions containing appropriate
presynaptic partners (dashed boxes). Image reproduced from Niell (2006) with permission (Copy-
right 2006 by Elsevier Ltd).
Imaging Circuit Formation in Zebrafish 349
Developmental Neurobiology

ily, which are responsible for filling synaptic
vesicles with the excitatory neurotransmitter gluta-
mate (Smear et al., 2007). Using patch-clamp
experiments in vivo it was found that synaptic
vesicles at blu retinotectal synapses contain less
glutamate, and that these synapses fatigue prema-
turely under high-frequency stimulation. Mutant
synapses appeared to function appropriately under
normal conditions however, suggesting that other
glutamate transporters can partially compensate for
the loss of vglut2a. Labeling of individual RGC
axons in the blu mutant reveals an increase in
RGC axon branch number and coverage area in the
tectum and an increase in the frequency of minia-
ture excitatory postsynaptic currents (mEPSCs) in
postsynaptic tectal cells. The increase in mEPSC
frequency may reflect an increase in the number of
presynaptic release sites in the enlarged axonal
arbors. The findings of the blu mutant study are
similar to those in the study that used TeNT-Lc-
EGFP to suppress synaptic transmission in that
expansion of RGC axonal arbor area was observed.
However, the two studies differ in one important as-
pect. In the study that used TeNT-Lc-EGFP (and also
the study that used the VAMP mutant), structural
changes in axons were only observed under competi-
tive conditions, i.e., when only a single cell was
silenced in a field of active ones. In the blu mutant,
where all RGCs are affected, alterations in single arbor
morphology were apparent. How can these differences
be reconciled? The authors of the blu study interpret
the expansion of RGC arbors, and the increased
mEPSC frequency as a homeostatic response to the
reduced glutamate content of vesicles. This contrasts
with the competitive forms of plasticity observed in
the Hua and Ben Fredj studies. Together the studies
outlined here may therefore represent graded suppres-
sion of synaptic function from mild (the blu mutant),
to severe (TeNT-Lc-EGFP). The type of plasticity
induced in RGC axons could change accordingly; mild
suppression could induce homeostatic changes
whereas more severe manipulations could induce com-
petitive forms of plasticity.
The increase in single axon arbor size in the blumutant is also likely to lead to greater convergence in
the blu tectum; an individual tectal cell is likely to be
postsynaptic partner to a greater number of RGCs,
and therefore respond to a larger part of the visual
scene. Indeed, using electrophysiological approaches,
tectal cell receptive fields were shown to be expanded
in the blu mutant relative to those in wild-type zebra-
fish. As would be predicted from the degradation of
the retinotopic map, blu mutants show impairment in
visually driven behaviors such as prey capture (Smear
et al., 2007). The study of the blu mutant therefore
nicely illustrates the feasibility of using multiple
parallel approaches in zebrafish to ask questions
relating to circuit formation. This study started with a
phenotype identified in a forward genetic screen and
used behavioral, electrophysiological, and imaging
techniques, to map out a path from genetic lesion, to
molecular and physiological deficits at an identified
synapse, to structural and functional alterations in
circuitry. However, this study also represents one of
the few examples that use electrophysiological tech-
niques in the visual system of zebrafish [see also
(Ramdya et al., 2006)]. The small size of tectal cells
make application of these techniques a considerable
challenge in zebrafish [although electrophysiological
approaches have been used extensively in studies of
circuitry elsewhere in the zebrafish nervous system
(McDearmid et al., 2006; McLean et al., 2007; Knogler
et al., 2010)]. In Xenopus tadpoles for example, there is
a long history of using both live imaging and electro-
physiology to study development of retinotectal cir-
cuitry (Alsina et al., 2001; Engert et al., 2002; Aizen-
man et al., 2003; Ruthazer et al., 2006).
Molecular Control of Axon Arbor Growthand Laminar Targeting
In addition to neural activity, transcription factors, re-
ceptor-ligand interactions, various signaling path-
ways, and local translational machinery, have all
been identified as contributors to the organization of
axonal and dendritic arbors and the placement of
these arbors in developing neuronal circuitry
(Huberman et al., 2010; Jan and Jan, 2010). In vivoimaging of RGC axons in two zebrafish mutants,
astray (ast) and dragnet (drg) have given us insight
into the molecular mechanisms regulating axon arbor
growth and laminar targeting of axons respectively.
In both vertebrates and invertebrates Roundabout
(Robo) and Slit proteins serve as repulsive axon guid-
ance cues. In zebrafish ast mutants that lack func-
tional robo2, RGC axons show pathfinding defects
consistent with the established role for these receptors
(Fricke et al., 2001). However, robo2 and one of its
ligands, slit1a are also expressed in the optic tectum,
suggesting additional roles for these signaling mole-
cules once RGC axons have reached their target
(Campbell et al., 2007). Analysis of single RGC axo-
nal arbors in ast mutants, and in zebrafish injected
with slit1a antisense morpholino oligonucleotides
revealed that both morphant and mutant arbors had
more branch tips, greater arbor area, and more
presynaptic release sites. Furthermore, time-lapse
350 Nikolaou and Meyer
Developmental Neurobiology

imaging revealed that ast and slit1a morphant arbors
stabilized earlier in development than wild-type ones
(Campbell et al., 2007). Together these findings
demonstrate a novel and important role for Robo-Slit
signaling in regulating morphogenesis of RGC axonal
arbors and development of the retinotopic map.
In addition to making retinotopic decisions, RGC
axons must make layer-specific targeting decisions.
RGC axons terminate in six retinorecipient layers
within the zebrafish tectum; the most superficial is
the stratum opticum (SO), three sublayers of the stra-
tum fibrosum et griseum superficiale (SFGS), the
stratum griseum centrale (SGC) and a deep layer bor-
dering the stratum album centrale (SAC), and the
stratum periventriculare (SPV). Imaging of single
RGC axons as they innervate the tectum shows that
in zebrafish, RGCs are confined to a single lamina,
and that laminar specificity is precise from the ear-
liest developmental stages (Xiao et al., 2005; Xiao
and Baier, 2007). Unlike retinotopic organization,
formation of synaptic laminae in the zebrafish tectum
appears to be an activity-independent process (Nevin
et al., 2008). A subset of RGC axons that project into
the SO and the two deep sublayers of the SFGS are
labeled in the Pou4f3:membraneGFP-expressingtransgenic line of zebrafish. This transgenic line has
been used in a forward genetic screen to identify mo-
lecular determinants of laminar choice (Xiao et al.,
2005). In one of these mutants, drg, the orderly lami-
nar distribution of RGC axons in the Pou4f3:mGFPtransgenic is disrupted (Xiao and Baier, 2007). Imag-
ing of single axons in the mutant showed that axons
often strayed from the SFGS to the SO, a phenotype
that was never observed in wild-type larvae. The
dragnet gene encodes collagen IVa5, a component of
the basement membrane that lines the surface of the
tectum. The distribution of heparan sulfate proteogly-
cans (HSPGs, a class of extracellular matrix mole-
cules involved in RGC axon guidance) which are nor-
mally associated with this basement membrane, are
disrupted in the mutant tectum. Data from the dragnetmutant suggests that collagen IVa5 is required to
anchor HSPGs in the basement membrane, which in
turn serves as a lamina targeting cue for RGC axons
(Xiao and Baier, 2007).
Zebrafish have been the focus of many mutant
screens and the studies outlined above illustrate how
mutagenesis allows the role of candidate molecules
to be tested in imaging experiments. However, for-
ward genetic screens are long-term, labor intensive
endeavors. Until recently techniques for reverse
genetic approaches in zebrafish have been limited to
mRNA knockdown strategies using modified anti-
sense morpholinos and TILLING for point mutations
among a library of chemically mutagenized gametes.
Both strategies have shortcomings: morpholinos for
example are active for only the first few days of
development and have many off target effects (Eisen
and Smith, 2008). However, the resistance of the
zebrafish genome to targeted sequence alteration has
recently been overcome through the use of zinc-finger
nucleases (ZFNs). Although still in its infancy, this
new technology promises to overcome what has been
a major gap in the repertoire of techniques available
to zebrafish researchers (Doyon et al., 2008).
Simultaneous Imaging ofPre and Postsynaptic Partners
The studies of axonal and dendritic arbor growth out-
lined above, while informative, have only observed
circuit assembly from either the pre- or postsynaptic
perspective. How interaction with other neurons mod-
ulates the growth process has only been inferred from
the expression of synaptic marker proteins within the
cells being imaged. Imaging pre- and postsynaptic
partners simultaneously as they contact one another
would provide a direct measure of how pre-post inter-
actions modulate axonal or dendritic arbor growth. A
time lapse imaging study in zebrafish has described
the behavior of pre- and postsynaptic neurons as they
contact one another in the spinal cord (Jontes et al.,
2000). The authors focused on Mauthner cells, which
are large, paired, hindbrain neurons that mediate the
escape response in zebrafish. Each Mauthner cell
axon crosses the ventral midline to extend down the
contralateral spinal cord to synapse on serially repeat-
ing sets of three primary motor neurons. The Mauth-
ner axon and motor neuron were visualized by label-
ing each cell type with lipophilic carbocyanide dyes
and time-lapse imaging was used to describe the
dynamic behavior of each cell during the course of
the interaction. The Mauthner growth cone was found
to move relatively smoothly down the spinal cord but
appeared to pause at, and interact with successive tar-
get cells (Fig. 3). Axonal varicosities, presumably
nascent synapses, later appeared at the points where
the axon stalled. Both the primary motor neurons and
the Mauthner axon had a large number of exploratory
filopodia, consistent with an active involvement of
both pre- and postsynaptic elements in establishing
synaptic contacts.
The above study took advantage of the relative
simplicity of the spinal cord wiring diagram, and of
the fact that the Mauthner axon forms synapses with
every motor neuron it passes. Together, these sim-
plify the task of labeling pre and postsynaptic part-
Imaging Circuit Formation in Zebrafish 351
Developmental Neurobiology

ners and of catching the initial interaction between
them. The complexity of other regions of the CNS
makes the job of labeling cells that are destined to
contact one another, and imaging the interaction,
much more challenging. However, in many regions
of the CNS, synaptic connections are arranged into
discrete layers. Such organization provides an assess-
able system for studying how axons and dendrites
target each other during circuit assembly. The inner
plexiform layer (IPL) of the vertebrate retina is a
particularly attractive model in this regard since the
cellular and synaptic organization of this structure is
highly stereotyped and almost crystalline in nature.
RGCs, amacrine cells, and biopolar cells form synap-
ses with one another in the IPL. These connections
are broadly divided into two layers: connections of
cells depolarized by light (ON cells) and cells hyper-
polarized by light (OFF cells) are localized to the
inner and outer halves of the IPL, respectively.
Within the ON and OFF layers are multiple, function-
ally distinct sublaminae. Time-lapse imaging of fluo-
rescently labeled RGCs, amacrine cells and bipolar
cells in zebrafish has revealed the sequence of events
that establish the synaptic laminae in the IPL (God-
inho et al., 2005; Mumm et al., 2006; Schroeter et al.,
2006). In the pax6-DF4::M-CFP transgenic line of
zebrafish cyan fluorescent protein is targeted to the
membranes of amacrine cells, allowing the individual
synaptic laminae within the IPL to be resolved (God-
inho et al., 2005). Time-lapse imaging of these fish
reveals that amacrine cells are the first to extend neu-
rites into the future IPL, and that laminar organization
emerges almost as soon amacrine cell neurites make
contact with one another (Godinho et al., 2005).
Bipolar cells are the last to arrive in the IPL, after the
connections between RGCs and amacrine cells have
formed (Schroeter et al., 2006). Although RGCs are
born before amacrine cells they don’t extend den-
drites into the IPL until after the amacrine cell plexu-
sus develop. Indeed, studies of the lakritz/atoh7mutant zebrafish that lacks RGCs shows that RGCs
are not absolutely required for amacrine sublamina-
tion (Kay et al., 2004). These findings suggest that
the early plexusus formed by amacrine cell neurites
may prepattern the IPL and provide lamination cues
for RGC dendrites. By time-lapse imaging single
RGCs labeled with yellow fluorescent protein within
the pax6-DF4::M-CFP background, the timing of
RGC dendrite versus amacrine cell stratification
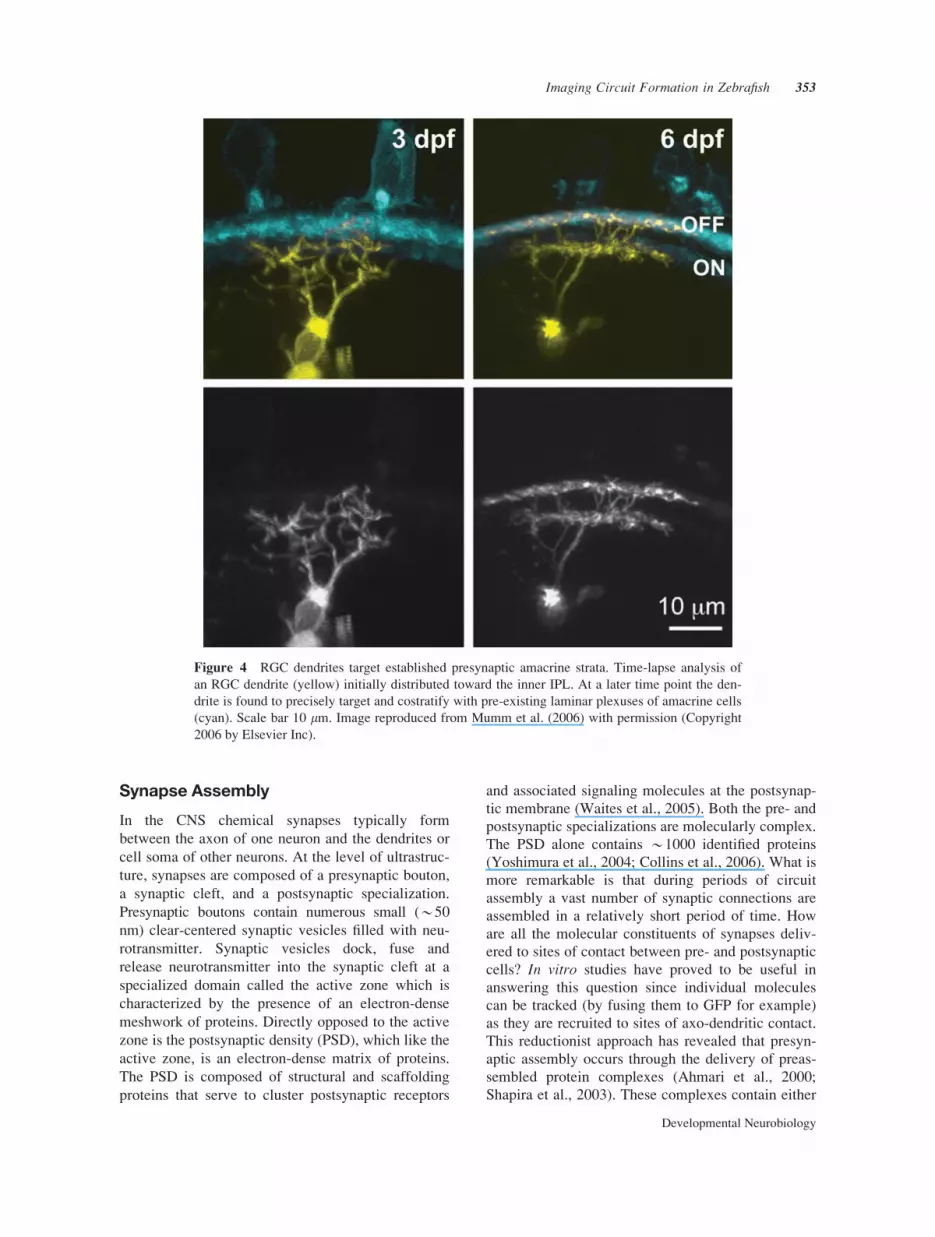
could be observed directly (Mumm et al., 2006).
While RGCs dendrites were found to exhibit diverse
growth and stratification patterns, most were found to
precisely target and costratify with pre-existing lami-
nar plexusus of amacrine cells (Fig. 4). Collectively
these observations suggest that amacrine cells first
target one another, perhaps by homotypic interac-
tions, to give rise to sublaminae, and that this process
is largely independent of RGCs. RGC dendrites then
use cues provided by these sublaminae to achieve
their final stratification pattern. These studies reveal
unexpected specificity in the targeting of dendrites
and also elegantly demonstrate the advantages of
using zebrafish and time-lapse imaging to study
mechanisms of circuit assembly.
Figure 3 Simultaneous imaging of the Mauthner growth
cone and primary motor neuron. The time-lapse sequence
of events show that the Mauthner growth cone is highly
active and transiently interacts with the motor neuron as it
migrates down the spinal cord. Although the growth cone
moved past its postsynaptic target without collapsing or sig-
nificantly altering its morphology, this interaction had a
subtle but detectable effect on growth cone dynamics. Scale
bar 10 lm. Image reproduced from Jontes et al. (2000) with
permission (Copyright 2000 by Nature Publishing Group).
352 Nikolaou and Meyer
Developmental Neurobiology
Synapse Assembly
In the CNS chemical synapses typically form
between the axon of one neuron and the dendrites or
cell soma of other neurons. At the level of ultrastruc-
ture, synapses are composed of a presynaptic bouton,
a synaptic cleft, and a postsynaptic specialization.
Presynaptic boutons contain numerous small (*50
nm) clear-centered synaptic vesicles filled with neu-
rotransmitter. Synaptic vesicles dock, fuse and
release neurotransmitter into the synaptic cleft at a
specialized domain called the active zone which is
characterized by the presence of an electron-dense
meshwork of proteins. Directly opposed to the active
zone is the postsynaptic density (PSD), which like the
active zone, is an electron-dense matrix of proteins.
The PSD is composed of structural and scaffolding
proteins that serve to cluster postsynaptic receptors
and associated signaling molecules at the postsynap-
tic membrane (Waites et al., 2005). Both the pre- and
postsynaptic specializations are molecularly complex.
The PSD alone contains *1000 identified proteins
(Yoshimura et al., 2004; Collins et al., 2006). What is
more remarkable is that during periods of circuit
assembly a vast number of synaptic connections are
assembled in a relatively short period of time. How
are all the molecular constituents of synapses deliv-
ered to sites of contact between pre- and postsynaptic
cells? In vitro studies have proved to be useful in
answering this question since individual molecules
can be tracked (by fusing them to GFP for example)
as they are recruited to sites of axo-dendritic contact.
This reductionist approach has revealed that presyn-
aptic assembly occurs through the delivery of preas-
sembled protein complexes (Ahmari et al., 2000;
Shapira et al., 2003). These complexes contain either
Figure 4 RGC dendrites target established presynaptic amacrine strata. Time-lapse analysis of
an RGC dendrite (yellow) initially distributed toward the inner IPL. At a later time point the den-
drite is found to precisely target and costratify with pre-existing laminar plexuses of amacrine cells
(cyan). Scale bar 10 lm. Image reproduced from Mumm et al. (2006) with permission (Copyright
2006 by Elsevier Inc).
Imaging Circuit Formation in Zebrafish 353
Developmental Neurobiology

synaptic vesicle precursors or active zone structural
proteins. The mechanisms of postsynaptic assembly
are more contentious. For example, there is evidence
that the postsynsptic scaffolding molecule, PSD-95 is
trafficked to the PSD by the gradual recruitment of
individual proteins, and through the delivery of pre-
fabricated protein complexes (Prange and Murphy,
2001; Bresler et al., 2004). While in vitro assays pro-
vide a powerful means to study the cell biology of
synapse assembly they cannot provide definitive
answers about how synapses are assembled in vivo.Many factors that are likely to impact synapse assem-
bly and stability such as molecular interactions with
neighboring cells and extracellular matrix, and
normal patterns of synaptic activity are perturbed
in vitro. Live imaging in zebrafish can complement
in vitro studies of synapse formation by providing the
natural three-dimensional environment of the nervous
system, normal physiological conditions and intact
sensory and synaptic input. Time-lapse imaging of
the synaptic vesicle protein, synaptophysin, fused to
GFP (syp-GFP) in RGC axons in zebrafish revealed
highly mobile clusters of syp-GFP within developing
axons, supporting the notion that presynaptic assem-
bly occurs via the recruitment of preassembled pack-
ets (Meyer and Smith, 2006). Similarly, the dynamic
behavior of a synaptic cell adhesion molecule, N-cad-
herin, was followed in Rohon-Beard sensory neurons
in the zebrafish spinal cord (Jontes et al., 2004; Latefi
et al., 2009). N-cadherin is thought to be involved in
the initial stages of synapse assembly and also syn-
apse stabilization. Like syp-GFP, an N-cadherin-GFP
fusion protein (N-cad-GFP) was transported within
axons as highly mobile protein clusters, while more
stable accumulations of N-cad-GFP corresponded to
synaptic sites. Many of the stable accumulations of
N-cad-GFP formed rapidly in the wake of migrating
growth cones suggesting that N-cadherin is recruited
early on during development of a synapse (Jontes et
al., 2004). Structure–function analysis revealed that
the extracellular domain of N-cadherin is required for
proper trafficking and targeting, and that post-transla-
tional processing of N-cadherin may be a mechanism
for regulating synaptogenesis in vivo (Jontes et al.,
2004; Latefi et al., 2009). Imaging the postsynaptic
marker protein PSD-95-GFP in zebrafish supports the
notion that postsynaptic assembly occurs very differ-
ently from presynaptic assembly. Imaging PSD-95-
GFP in zebrafish tectal cells in vivo showed that this
scaffolding protein concentrated at PSDs by the grad-
ual accumulation from an initially diffuse distribution
in dendrites (Niell et al., 2004). The modular recruit-
ment of PSD-95 to synaptic sites that has been
observed in vitro was not apparent in tectal cells invivo.
One advantage of in vitro systems is that it is
feasible to visualize contact events between pre and
postsynaptic neurons, and then see how and when
synaptic components are recruited to contact sites. As
we have already mentioned, imaging such contact
events in vivo represents a considerable challenge,
particularly in the CNS. The neuromuscular junction
(NMJ) however is more accessible to labeling and
imaging. Transgenic lines of zebrafish, that express
GFP in motor neurons, allow visualization of motor
axons as they innervate their muscle targets. This,
coupled with staining live embryos with fluorescently
tagged a-bungarotoxin [a toxin that binds to postsy-
naptic acetylcholine receptors (AChR)] enables si-
multaneous imaging of both the presynaptic and post-
synaptic specializations in vivo. Thus, the NMJ is an
attractive model for studying the sequence of events
during synapse assembly. Furthermore, because myo-
tubes and motor neurons develop in a rostral-to-cau-
dal progression, the whole sequence of events under-
lying NMJ assembly can be viewed in a single zebra-
fish embryo (Flanagan-Steet et al., 2005; Panzer et
al., 2005, 2006). Two time lapse imaging studies
have demonstrated that prepatterned AChRs exist on
muscle fibers before the arrival of motor axons (Fla-
nagan-Steet et al., 2005; Panzer et al., 2006). Follow-
ing contact with these clusters, presynaptic vesicles
rapidly clustered in motor axons (Fig. 5). These
observations suggested that at the NMJ, postsynaptic
differentiation precedes presynaptic differentiation
and that the formation of initial postsynaptic special-
izations does not require motor axons. Postsynaptic
differentiation was nonetheless found to be nerve de-
pendent, in that axons are crucial for the maturation
and maintenance of the postsynaptic apparatus.
CONCLUDING REMARKS
By imaging nervous system development in zebrafish
we have already gained valuable insight into the
mechanisms of nervous system assembly. The future
looks as, if not more promising. The recent develop-
ment of photo-switchable ion channels and pumps
such as channelrhodopsin (ChR2) and halorhodopsin
(NpHR) has already enabled non-invasive dissection
of circuit function in zebrafish (Douglass et al., 2008;
Arrenberg et al., 2009; Schoonheim et al., 2010).
These tools could also be used to address questions
relating to nervous system plasticity. For example, by
using them to control the firing pattern and rate
within a developing neuron we could ask what kinds
of activity patterns drive structural changes in neu-
rons. Genetically encoded reporters of synaptic func-
354 Nikolaou and Meyer
Developmental Neurobiology

tion could also provide a means to examine the emer-
gence of circuit function, functional plasticity at the
level of single synapses, and to relate synapse func-
tion to the structural plasticity of neurons (Dreosti et al.,
2009). For example, the functional properties of an
identified synapse could be related to the dynamic
behavior of the axonal or dendritic branch on which it
is located. Furthermore enhancer and gene trapping
approaches are generating hundreds of Gal4 driver lines
in which the expression of UAS-linked effectors can be
targeted to subpopulations of neurons (Scott et al.,
2007; Asakawa and Kawakami, 2009). This approach
promises to vastly expand the repertoire of neuronal
cell types that are accessible to manipulation and imag-
ing.
The authors thank Juan Burrone, Sarah Hammond, and
Paul Hunter for helpful comments on the manuscript.
Figure 5 Postsynaptic differentiation precedes and guides axon outgrowth. Time-lapse images of
a motor axon growth cone extending toward, contacting, and extending beyond pre-patterned AChR
clusters. The motor axon was labeled with VAMP-GFP (green) and the AChR clusters with a-bun-garotoxin (red). A number of pre-existing AChR clusters are seen ahead of the growth cone, which
eventually contacts the clusters one by one. In some cases VAMP-GFP positive clusters of presynap-
tic vesicles accumulate over pre-patterned AChR clusters (asterisks), possibly forming NMJs.
Toward the end of the time-lapse imaging the growth cone is seen to turn toward pre-patterned
AChR clusters at the dorsal end of the myotome (arrowheads). Scale bar 10 lm. Images reproduced
from Panzer et al. (2006) with permission (Copyright 2006 by the Society for Neuroscience).
Imaging Circuit Formation in Zebrafish 355
Developmental Neurobiology

REFERENCES
Ahmari SE, Buchanan J, Smith SJ. 2000. Assembly of pre-
synaptic active zones from cytoplasmic transport packets.
Nat Neurosci 3:445–451.
Aizenman CD, Akerman CJ, Jensen KR, Cline HT. 2003.
Visually driven regulation of intrinsic neuronal excitabil-
ity improves stimulus detection in vivo. Neuron 39:831–
842.
Alsina B, Vu T, Cohen-Cory S. 2001. Visualizing
synapse formation in arborizing optic axons in vivo:
Dynamics and modulation by BDNF. Nat Neurosci
4:1093–1101.
Arrenberg AB, Del BF, Baier H. 2009. Optical control of
zebrafish behavior with halorhodopsin. Proc Natl Acad
Sci USA 106:17968–17973.
Asakawa K, Kawakami K. 2009. The Tol2-mediated Gal4-
UAS method for gene and enhancer trapping in zebrafish.
Methods 49:275–281.
Baier H, Klostermann S, Trowe T, Karlstrom RO, Nus-
slein-Volhard C, Bonhoeffer F. 1996. Genetic dissection
of the retinotectal projection. Development 123:415–
425.
Ben FN, Hammond S, Otsuna H, Chien CB, Burrone J,
Meyer MP. 2010. Synaptic activity and activity-depend-
ent competition regulates axon arbor maturation, growth
arrest, and territory in the retinotectal projection. J Neu-
rosci 30:10939–10951.
Bishop DL, Misgeld T, Walsh MK, Gan WB, Lichtman
JW. 2004. Axon branch removal at developing synapses
by axosome shedding. Neuron 44:651–661.
Bresler T, Shapira M, Boeckers T, Dresbach T, Futter M,
Garner CC, Rosenblum K, et al. 2004. Postsynaptic
density assembly is fundamentally different from
presynaptic active zone assembly. J Neurosci 24:1507–
1520.
Campbell DS, Stringham SA, Timm A, Xiao T, Law MY,
Baier H, Nonet ML, et al. 2007. Slit1a inhibits retinal
ganglion cell arborization and synaptogenesis via Robo2-
dependent and -independent pathways. Neuron 55:231–
245.
Collins MO, Husi H, Yu L, Brandon JM, Anderson CN,
Blackstock WP, Choudhary JS, et al. 2006. Molecular
characterization and comparison of the components and
multiprotein complexes in the postsynaptic proteome.
J Neurochem 97(Suppl 1):16–23.
Douglass AD, Kraves S, Deisseroth K, Schier AF, Engert F.
2008. Escape behavior elicited by single, channelrho-
dopsin-2-evoked spikes in zebrafish somatosensory neu-
rons. Curr Biol 18:1133–1137.
Doyon Y, McCammon JM, Miller JC, Faraji F, Ngo C,
Katibah GE, Amora R, et al. 2008. Heritable targeted
gene disruption in zebrafish using designed zinc-finger
nucleases. Nat Biotechnol 26:702–708.
Dreosti E, Odermatt B, Dorostkar MM, Lagnado L. 2009.
A genetically encoded reporter of synaptic activity in
vivo. Nat Methods 6:883–889.
Easter SS Jr, Nicola GN. 1996. The development of vision
in the zebrafish (Danio rerio). Dev Biol 180:646–663.
Eisen JS, Smith JC. 2008. Controlling morpholino experi-
ments: Don’t stop making antisense. Development
135:1735–1743.
Engert F, Tao HW, Zhang LI, Poo MM. 2002. Moving vis-
ual stimuli rapidly induce direction sensitivity of devel-
oping tectal neurons. Nature 419:470–475.
Flanagan-Steet H, Fox MA, Meyer D, Sanes JR. 2005. Neu-
romuscular synapses can form in vivo by incorporation
of initially aneural postsynaptic specializations. Develop-
ment 132:4471–4481.
Fricke C, Lee JS, Geiger-Rudolph S, Bonhoeffer F, Chien
CB. 2001. Astray, a zebrafish roundabout homolog
required for retinal axon guidance. Science 292:507–510.
Godinho L, Mumm JS, Williams PR, Schroeter EH,
Koerber A, Park SW, Leach SD, et al. 2005. Targeting of
amacrine cell neurites to appropriate synaptic laminae in
the developing zebrafish retina. Development 132:5069–
5079.
Haas K, Li J, Cline HT. 2006. AMPA receptors regulate
experience-dependent dendritic arbor growth in vivo.
Proc Natl Acad Sci USA 103:12127–12131.
Hua JY, Smear MC, Baier H, Smith SJ. 2005. Regulation of
axon growth in vivo by activity-based competition.
Nature 434:1022–1026.
Huberman AD, Clandinin TR, Baier H. 2010. Molecular
and cellular mechanisms of lamina-specific axon target-
ing. Cold Spring Harb Perspect Biol 2:a001743.
Huberman AD, Feller MB, Chapman B. 2008. Mechanisms
underlying development of visual maps and receptive
fields. Annu Rev Neurosci 31:479–509.
Jan YN, Jan LY. 2010. Branching out: Mechanisms of den-
dritic arborization. Nat Rev Neurosci 11:316–328.
Jontes JD, Buchanan J, Smith SJ. 2000. Growth cone and
dendrite dynamics in zebrafish embryos: Early events in
synaptogenesis imaged in vivo. Nat Neurosci 3:231–237.
Jontes JD, Emond MR, Smith SJ. 2004. In vivo trafficking
and targeting of N-cadherin to nascent presynaptic termi-
nals. J Neurosci 24:9027–9034.
Kaethner RJ, Stuermer CA. 1992. Dynamics of terminal
arbor formation and target approach of retinotectal axons
in living zebrafish embryos: A time-lapse study of single
axons. J Neurosci 12:3257–3271.
Kaethner RJ, Stuermer CA. 1997. Dynamics of process
formation during differentiation of tectal neurons in
embryonic zebrafish. J Neurobiol 32:627–639.
Kay JN, Roeser T, Mumm JS, Godinho L, Mrejeru A,
Wong RO, Baier H. 2004. Transient requirement for
ganglion cells during assembly of retinal synaptic layers.
Development 131:1331–1342.
Keck T, Mrsic-Flogel TD, Vaz AM, Eysel UT, Bonhoeffer
T, Hubener M. 2008. Massive restructuring of neuronal
circuits during functional reorganization of adult visual
cortex. Nat Neurosci 11:1162–1167.
Knogler LD, Liao M, Drapeau P. 2010. Synaptic scaling
and the development of a motor network. J Neurosci
30:8871–8881.
Latefi NS, Pedraza L, Schohl A, Li Z, Ruthazer ES. 2009.
N-cadherin prodomain cleavage regulates synapse forma-
tion in vivo. Dev Neurobiol 69:518–529.
356 Nikolaou and Meyer
Developmental Neurobiology

Lichtman JW, Smith SJ. 2008. Seeing circuits assemble.
Neuron 60:441–448.
McDearmid JR, Liao M, Drapeau P. 2006. Glycine recep-
tors regulate interneuron differentiation during spinal
network development. Proc Natl Acad Sci USA 103:
9679–9684.
McLean DL, Fan J, Higashijima S, Hale ME, Fetcho JR.
2007. A topographic map of recruitment in spinal cord.
Nature 446:71–75.
Meyer MP, Smith SJ. 2006. Evidence from in vivo imaging
that synaptogenesis guides the growth and branching of
axonal arbors by two distinct mechanisms. J Neurosci
26:3604–3614.
Mumm JS, Williams PR, Godinho L, Koerber A, Pittman
AJ, Roeser T, Chien CB, et al. 2006. In vivo imaging
reveals dendritic targeting of laminated afferents by
zebrafish retinal ganglion cells. Neuron 52:609–621.
Nevin LM, Taylor MR, Baier H. 2008. Hardwiring of fine
synaptic layers in the zebrafish visual pathway. Neural
Dev 3:36.
Niell CM. 2006. Theoretical analysis of a synaptotropic
dendrite growth mechanism. J Theor Biol 241:39–48.
Niell CM, Meyer MP, Smith SJ. 2004. In vivo imaging of
synapse formation on a growing dendritic arbor. Nat
Neurosci 7:254–260.
Niell CM, Smith SJ. 2004. Live optical imaging of nervous
system development. Annu Rev Physiol 66:771–798.
O’Rourke NA, Cline HT, Fraser SE. 1994. Rapid remodel-
ing of retinal arbors in the tectum with and without
blockade of synaptic transmission. Neuron 12:921–934.
Panzer JA, Gibbs SM, Dosch R, Wagner D, Mullins MC,
Granato M, Balice-Gordon RJ. 2005. Neuromuscular
synaptogenesis in wild-type and mutant zebrafish. Dev
Biol 285:340–357.
Panzer JA, Song Y, Balice-Gordon RJ. 2006. In vivo
imaging of preferential motor axon outgrowth to and
synaptogenesis at prepatterned acetylcholine receptor
clusters in embryonic zebrafish skeletal muscle. J Neuro-
sci 26:934–947.
Prange O, Murphy TH. 2001. Modular transport of postsy-
naptic density-95 clusters and association with stable
spine precursors during early development of cortical
neurons. J Neurosci 21:9325–9333.
Ramdya P, Reiter B, Engert F. 2006. Reverse correlation of
rapid calcium signals in the zebrafish optic tectum in
vivo. J Neurosci Methods 157:230–237.
Ruthazer ES, Cline HT. 2004. Insights into activity-depend-
ent map formation from the retinotectal system: A mid-
dle-of-the-brain perspective. J Neurobiol 59:134–146.
Ruthazer ES, Li J, Cline HT. 2006. Stabilization of axon
branch dynamics by synaptic maturation. J Neurosci
26:3594–3603.
Schoonheim PJ, Arrenberg AB, Del BF, Baier H. 2010.
Optogenetic localization and genetic perturbation of sac-
cade-generating neurons in zebrafish. J Neurosci 30:
7111–7120.
Schroeter EH, Wong RO, Gregg RG. 2006. In vivo devel-
opment of retinal ON-bipolar cell axonal terminals
visualized in nyx::MYFP transgenic zebrafish. Vis Neu-
rosci 23:833–843.
Scott EK, Mason L, Arrenberg AB, Ziv L, Gosse NJ, Xiao
T, Chi NC, et al. 2007. Targeting neural circuitry in
zebrafish using GAL4 enhancer trapping. Nat Methods
4:323–326.
Shapira M, Zhai RG, Dresbach T, Bresler T, Torres VI,
Gundelfinger ED, Ziv NE, et al. 2003. Unitary assembly
of presynaptic active zones from Piccolo-Bassoon trans-
port vesicles. Neuron 38:237–252.
Smear MC, Tao HW, Staub W, Orger MB, Gosse NJ, Liu
Y, Takahashi K, et al. 2007. Vesicular glutamate trans-
port at a central synapse limits the acuity of visual per-
ception in zebrafish. Neuron 53:65–77.
Sorensen JB, Matti U, Wei SH, Nehring RB, Voets T, Ash-
ery U, Binz T, et al. 2002. The SNARE protein SNAP-25
is linked to fast calcium triggering of exocytosis. Proc
Natl Acad Sci USA 99:1627–1632.
Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR,
Welker E, Svoboda K. 2002. Long-term in vivo imaging
of experience-dependent synaptic plasticity in adult cor-
tex. Nature 420:788–794.
Waites CL, Craig AM, Garner CC. 2005. Mechanisms of
vertebrate synaptogenesis. Annu Rev Neurosci 28:251–
274.
Xiao T, Baier H. 2007. Lamina-specific axonal projections
in the zebrafish tectum require the type IV collagen
Dragnet. Nat Neurosci 10:1529–1537.
Xiao T, Roeser T, Staub W, Baier H. 2005. A GFP-based
genetic screen reveals mutations that disrupt the architec-
ture of the zebrafish retinotectal projection. Development
132:2955–2967.
Yang G, Pan F, Gan WB. 2009. Stably maintained dendritic
spines are associated with lifelong memories. Nature
462:920–924.
Yoshimura Y, Yamauchi Y, Shinkawa T, Taoka M, Donai H,
Takahashi N, Isobe T, et al. 2004. Molecular constituents
of the postsynaptic density fraction revealed by proteomic
analysis using multidimensional liquid chromatography-
tandem mass spectrometry. J Neurochem 88:759–768.
Imaging Circuit Formation in Zebrafish 357
Developmental Neurobiology





























