Embed Size (px)
Citation preview

Research in Veterinary Science 1996, 60, 82-87
Immunocytochemical detection of growth factors (PDGF and TGF in equine chronic pneumonia
A. RODR[GUEZ, M. CASTAIgqO, L. PElqA, M. A. SANCHEZ, A. NIETO, M. RODR.[GUEZ, Departamento Patologfa Animal II, Facultad de Veterinaria, Universidad Complutense, 28040 - Madrid, Spain
SUMMARY
The roles played by platelet-derived growth factor (PDGF) and transforming growth factor 13 (TGF ~) in the development of pulmonary fibrosis in equine chronic pneumonia varied greatly. Macrophages, epithelial cells and fihroblasts were identified as a source of PDGF, the principal role of which was related to its mitotic effect on epithelial cells, and particularly on fibroblasts in the final phase of the disease. TGF ~ was detected in epithelial cells in all three phases of the disease and in fibroblasts in the last phase. However, the role of TGF [~ in this pulmonary disease is not clear because its expression in the cytoplasm of fibroblasts in areas of strong collagenation during the last phase was weak. Nonetheless, it was responsible for the production and release of collagen during the stage of total fibrosis.
PULMONARY fibrosis is a chronic response of the parenchyma to various disorders, independent of the animal species, which may progress to a final stage (Spencer 1975, Hara et al 1989, Barrios and Selman 1991, Harrison and Laurent 1991). Fibrosis is a significant lesion in many human diseases (atherosclerosis, interstitial lung disease, hepatic cirrhosis, etc), and in recent years numerous studies have focused on one major interstitial lung affection of unknown aetiology which has been called idiopathic pul- monary fibrosis (IPF) (Liebow 1975, Spencer 1975, Crystal 1987, Selman 1990, Selman and Barrios 1991).
Research has demonstrated that the equine lung can develop various degrees of fibrosis (Cagny and Gobert 1880, Fr6hner and Zwick 1932, Nieberle and Cohrs 1967, Buergelt et al 1986, Dungworth 1992); the most severe manifestation occurs in chronic pneumonia, in which the histological findings are similar to those of human 'honey- comb' lung. However, most of the literature on the subject concerns human and experimental pulmonary fibrosis (Adamson and Bowden 1979, Selman et al 1985, Adamson et al 1988, Hara et al 1989, Selman et al 1989) and many phenomena which occur during the development of human and animal fibrosis are not completely understood. Davidson (1990) and Kumar et al (1993) have stated that fibrosis is due to the accumulation of collagens and other extracellular matrix proteins, such as fibronectin and elastin, in the alveolar region, which cause severe dysfunc- tions of the alveolar-capillary unit. Fibrosis is considered to be an exaggeration of the normal process of healing after a chronic inflammation caused by an imbalance in the home- ostasis of collagen, with excessive synthesis and insuffi- cient degradation (Lanrent 1986, Davidson 1990, Selman and Barrios 1991, Kumar et al 1993).
Recently, Barrios and Selman (1991) and Kumar et al (1993) have demonstrated that the increased deposits of collagen in the lungs are the result of the abnormal prolifer- ation and accumulation of fibroblasts in the presence of inflammatory cells which release cytokines or growth fac- tors that are mitogenic for fibroblasts, or that promote the synthesis of collagen (Wahl et al 1987, Selman and Barrios 1991, Kumar et al 1993). Some of these mitotic growth factors have been found in patients with interstitial lung
disorders (Antoniades et al 1991), and at least five families are known: the platelet-derived growth factor (PDGF), epidermal growth factor/transforming growth factor (x (EGF/TGF ~), fibroblast growth factor (FGF), transforming growth factor 13 (TGF [3) and insulin-like growth factors (IGF-I and II) (Benito and Lorenzo 1993, Pusztai et al 1993). Activated human alveolar macrophages have been shown to synthesise PDGF (Sugarman et al 1985, Diegelman et al 1986, Osornio-Vargas et al 1991), which is a platelet peptide found in alpha granules, and is chemotac- tic and mitogenic for fibroblasts and smooth muscle fibres of blood vessels (Bitterman et al 1986, Richardson and Campion 1988, Mendoza et al 1990). Alveolar macro- phages are the predominant inflammatory cells found in cases of desquamative pneumonia (Crystal et al 1984) and during the process of healing after parenchymal tissue dam- age (Mornex et al 1986), and their content of PDGF would partially explain the mechanism of human pulmonary fibro- sis (Martinet et al 1987, Raghu and Kavanagh 1991).
Similar studies have been done in vivo by Selman et al (1989) and Antoniades et al (1991), who have demonstrat- ed the presence of PDGF proteins in the alveolar mono- nuclear cells and cubic epithelial cells of individuals with a high degree of pulmonary cell proliferation. PDGF and ANN- m were also detected in lung biopsies of human beings with IPF by means of immunohistochemical techniques and in situ hybridisation, respectively. These investigations con- firmed the presence of PDGF in epithelial cells of individu- als with pulmonary fibrosis (Antoniades et al 1991). After a severe pulmonary lesion, this growth factor has the effect of inducing the proliferation of mesenchymal cells. The continuous proliferative response leads to an extensive intra-alveolar fibrosis, even when the inflammatory process had ended (Chen et al 1992, Fabisiak et al 1992).
However, the action of PDGF should not be studied alone because, for example, it is known that the increased colla- gen content of inflammatory or fibrotic lesions is the result of the combined action of eDGF and TGF ~ (Ooshima 1981, Morton and Barnes 1982, Narayanan et al 1983). TGF ~ is a polypeptide which regulates growth and differentiation (Assoian and Spom 1986, Assoian et al 1987, Spom et al 1987) and influences the synthesis and degradation of
82

PDGF and TGF )~ in equine chronic pneumonia 83
matrix components. More specifically, this growth factor stimulates the production of collagen by pulmonary fibro- blasts (Fine and Goldstein 1987), which results in the syn- thesis of fibrotic tissue in vivo (Spom et al 1987). In vitro studies with fibroblasts from normal and fibrotic human lungs have shown that this growth factor increases the rate of production of collagen types I, III and V by up to four times the normal rate, as well as increasing the rate of pro- duction of other extracellular matrix proteins such as fibronectin and glycosaminoglycans, with practically no proliferation of fibroblasts (Roberts et al 1985, 1986, Ignotz and Massagu6 1986, Ignotz et al 1987, Fine and Goldstein 1987, Antoniades et al 1991). Nevertheless, Bernard et al (1990) state that TGF ~ stimulates the production of certain mesenchymal cells, such as fibroblasts, but not of epithelial cells, while inhibiting the proliferation of neoplastic carci- nomatous cells (lung, breast, endometrium and liver carci- nomas) and sarcomatous cells (melanoma, fibrosarcoma and osteosarcoma) in vitro. The apparent contradiction between the well-documented in vitro inhibitory activity of TGF ~ and its ability to promote tumorous formations in vivo under certain circumstances (Torre-Amione et al 1990), underlines the difficulty of extrapolating from in vitro experimental results to in vivo situations in which many substances, some of which may be unknown, inter- vene. TGF ~ and other growth factors probably act as syner- gists or antagonists in the proliferation of fibroblastic cells in pulmonary parenchyma in vivo. Pulmonary fibrosis results from the interaction of various substances, including growth factors, which affect the rate of synthesis and depo- sition of collagen (Antoniades et al 1991).
There is no information concerning the detection of PDGF or TGF ~ in cases of equine pulmonary fibrosis. Relevant data would be of interest not only from the veterinary point of view, but also for its application in human medicine, owing to the numerous anatomical and physiopathological similarities between human and equine lungs. The aim of this study was to detect the growth factors PDGF and TGF [~ by using immunocytochemical methods, in order to identify the cells most responsible for their production and to study the role of the factors in equine pulmonary fibrosis.
TABLE 2: Immunofluorescent activity of PDGF in different types of lung cell in horses at different phases of chronic pneumonia
Phase of chronic pneumonia Type of cell Initial Intermediate Final
Smooth muscle cells - - + Alveolar macrophages +/++ - /+ - Fibroblasts Intra-alveolar formations - +/++
Alveolar septa - - /+ +++ Epithelial cells Bronchial - /+ +/++ ++
Bronchiolar + ++ +++
Immunocytochemical detection of PDGF: - Negative, + Slightly positive, ++ Positive, +++ Highly positive
TABLE 3: Immunofluorescent activity to TGF j~ in different types of lung cell in horses at different phases of chronic pneumonia
Phase of chronic pneumonia Type of cell Initial Intermediate Final
Epithelial cells Bronchial - - /+ + Bronchiolar - /+ + ++
Fibroblasts Intra-alveolar formations - +/++ Alveolar septa - - /+ +++
Immunocytochemical detection of TGF J~: - Negative, + Slightly positive, ++ Positive, +++ Highly positive
1/30, and anti-TGF ~ (from porcine platelets) developed in rabbits (British Biotechnology, BDA 5) and used at a work- ing dilution of 1/5. The secondary antibodies were conju- gated with fluorescein isothiocyanate (FITC), depending on the primary antisera: rabbit IgG anti-goat IgG F~TC (Sigma Immunochemicals F-7367, working dilution 1/64) and sheep IgG anti-rabbit IgG FITC (ICN Immunobiologicals 683081-0012) for PDGF and TGF ~, respectively (Table 1). Canine mammary turnouts with marked fibroplasia were used as positive controls for TGF ~ and PDGF, and negative controls were made by replacing the first antibody with phosphate buffered saline (PBS) pH 7.2.
According to the proportion of positive cells and the intensity of the fluorescence, the immunocytochernical detection of PDGF (Table 2) and TGF [3 (Table 3) was classi- fied as negative, slightly positive, positive or highly positive.
MATERIALS AND METHODS
The study used the lungs of 68 crossbred horses, on aver- age eight years of age (Table 1), which were slaughtered in the Madrid municipal abattoir. Eight pulmonary samples were obtained from each animal by the method established by Viel (1983), fixed in buffered formalin and Bouin's fluid and embedded in paraffin wax. The sections were stained with haematoxylin and eosin, Masson's trichrome and Schiff' s periodic acid.
Indirect immunofluorescence was used to detect the growth factors. The primary antibodies were anti-human PDGF (A and B chains) developed in goats (British Biotechnology, BDA 16) and used at a working dilution of
TABLE 1: Numbers and age ranges of the horses in different phases of chronic pneumonia
Phase of Age (years) chronic pneumonia 0-5 5-15 >15
Initial 10 3 1 Intermediate 16 16 4 Final 3 9 6
RESULTS
There were two types of gross lesions in the equine lungs: one group of lungs had a characteristic diffuse tan- coloured induration, most evident in the hilar region, which usually corresponded with interstitial pneumonia; the other group of lungs exhibited wide whitish masses of connective tissue which extended, in some cases, from the cranial lobe along the dorsal side of the caudal lobe.
All the histological samples showed signs of chronic interstitial pneumonia, which varied according to the degree of the accumulation of inflammatory cells and fibro- sis in the interstitium. According to the severity of the lesions and the degree of fibrosis, the samples were divided into three phases of disease: an initial phase which affected 14 of the horses (20.6 per cent), an intermediate phase which affected 36 of the horses (53 per cent) and a final phase which affected 18 of the horses (26.4 per cent).
Initial phase
This phase was characterised by oedematous septa with scars, inflammatory cells (lymphocytes and neutrophils), capillary congestion and proliferation. In the alveolar
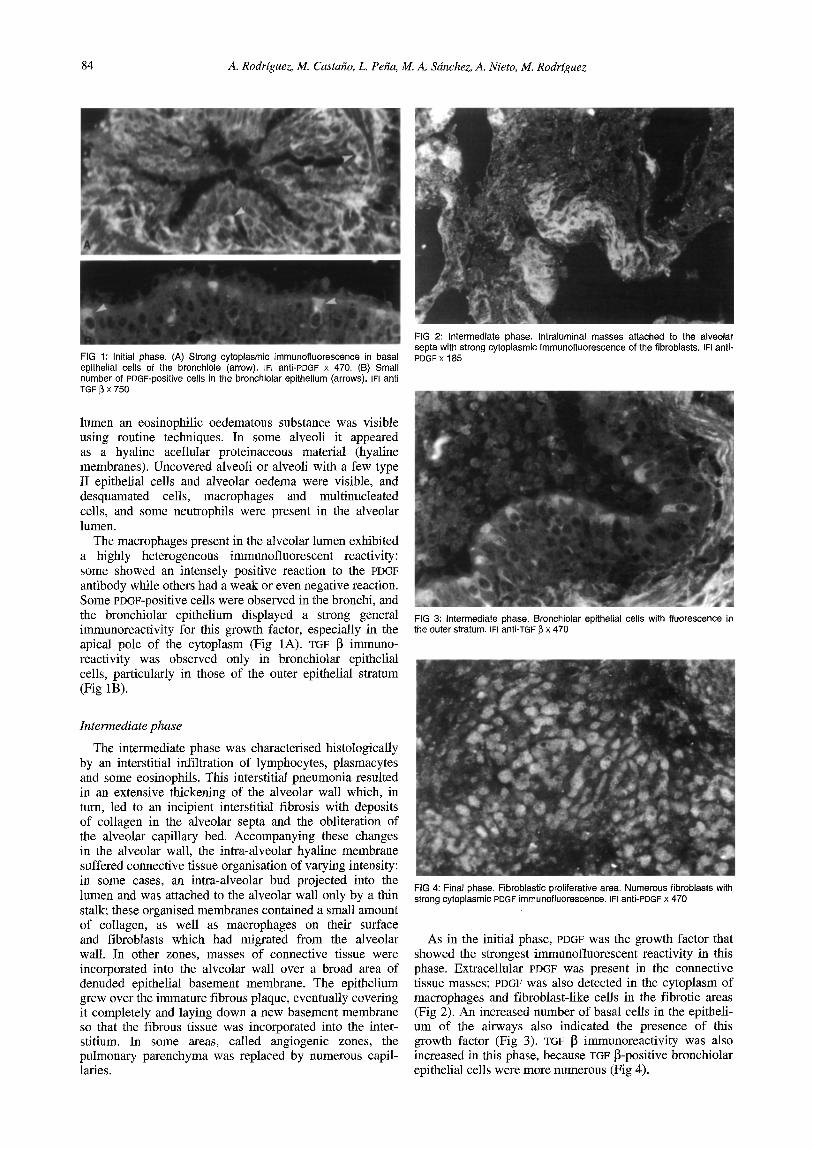
84 A. Rodrfguez, M. Casta~o, L. Pe~a, M. A. Sdnchez, A. Nieto, M. Rodrfguez
FIG 1: Initial phase. (A) Strong cytoplasmic immunofluorescence in basal epithelial cells of the bronchiole (arrow), IFI anti-PDGF x 470. (B) Small number of PDGF-positive cells in the bronchiolar epithelium (arrows). IFI anti TGF ~ X 750
lumen an eosinophilic oedematous substance was visible using routine techniques. In some alveoli it appeared as a hyaline aceUular proteinaceous material (hyaline membranes). Uncovered alveoli or alveoli with a few type II epithelial cells and alveolar oedema were visible, and desquamated cells, macrophages and multinucleated cells, and some neutrophils were present in the alveolar lumen.
The macrophages present in the alveolar lumen exhibited a highly heterogeneous immunofluorescent reactivity: some showed an intensely positive reaction to the PDGF antibody while others had a weak or even negative reaction. Some PDGF-positive cells were observed in the bronchi, and the bronchiolar epithelium displayed a strong general immunoreactivity for this growth factor, especially in the apical pole of the cytoplasm (Fig 1A). TGF ~ immuno- reactivity was observed only in bronchiolar epithelial cells, particularly in those of the outer epithelial stratum (Fig 1B).
Intermediate phase
The intermediate phase was characterised histologically by an interstitial infiltration of lymphocytes, plasmacytes and some eosinophils. This interstitial pneumonia resulted in an extensive thickening of the alveolar wall which, in turn, led to an incipient interstitial fibrosis with deposits of collagen in the alveolar septa and the obliteration of the alveolar capillary bed. Accompanying these changes in the alveolar wall, the intra-alveolar hyaline membrane suffered connective tissue organisation of varying intensity: in some cases, an intra-alveolar bud projected into the lumen and was attached to the alveolar wall only by a thin stalk; these organised membranes contained a small amount of collagen, as well as macrophages on their surface and fibroblasts which had migrated from the alveolar wall. In other zones, masses of connective tissue were incorporated into the alveolar wall over a broad area of denuded epithelial basement membrane. The epithelium grew over the immature fibrous plaque, eventually covering it completely and laying down a new basement membrane so that the fibrous tissue was incorporated into the inter- stitium. In some areas, called angiogenic zones, the pulmonary parenchyma was replaced by numerous capil- laries.
FIG 2: Intermediate phase. Intraluminal masses attached to the alveolar septa with strong cytoplasmic immunofluorescence of the fibroblasts. IFI anti- PDGF x 185
FIG 3: Intermediate phase. Bronchiolar epithelial cells with fluorescence in the outer stratum. IFI anti-TGF ~ x 470
FIG 4: Final phase. Fibroblastic proliferative area. Numerous fibroblasts with strong cytoplasmic PDGF immunofluorescence. IFI anti-PDGF x 470
As in the initial phase, PDGF was the growth factor that showed the strongest immunofluorescent reactivity in this phase. Extracellular PDGF was present in the connective tissue masses; PDGF was also detected in the cytoplasm of macrophages and fibroblast-like cells in the fibrofic areas (Fig 2). An increased number of basal cells in the epitheli- um of the airways also indicated the presence of this growth factor (Fig 3). TGF ~ immunoreactivity was also increased in this phase, because TGF ~-positive bronchiolar epithelial cells were more numerous (Fig 4).

PDGF and TGF t~ in equine chronic pneumonia 85
Final phase
This phase corresponded with advanced lesions, and the histological changes in the lungs included severe intra- and inter-alveolar fibrosis. In addition, small cavities lined with cuboid or columnar cells were observed, indicating the overgrowth of the bronchiolar epithelium next to adjacent emphysematous areas. These histopathological changes are similar to those of the 'honeycomb lung' observed in human pathology. It was also possible to observe extensive masses of connective tissue without remodelled alveoli in the final phase of total fibrosis. Two different classes of fibrotic tissue could be differentiated: one of a more cellu- lar type (immature fibrous tissue), and another of a more connective tissue nature (mature fibrous tissue). Hyper- plasia of the intrapulmonary lymphoid follicles was some- times observed in the interstitium.
During this phase, immunofluorescence revealed the presence of the growth factors in the cytoplasm of mes- enchymal cells. In the cellular type of fibrous tissue (young fibrous tissue), with immature fibroblast proliferation and scarce deposits of fibrous extracellular matrix, the fibro- blasts displayed strong immunofluorescent reactivity (Fig 5). As the fibrotic tissue aged, creating areas of abundant fibrous extracellular matrix and some fibrocytes, a marked decrease of PDGF reactivity was evident. In this phase of equine chronic pneumonia with fibrosis, the hyperplastic epithelial cells of the airways exhibited intense PDGF fluo- rescence (Fig 6). PDGF was also detected in the smooth
FIG 5: Final phase. Bronchiolar epithelium. Strongest expression of PDGF in the epithelial cells. IFI anti-PDGF x 185
FiG 6: Final phase. Discrete cytoplasmic immunoreaction of the fibroblasts for TGF ~ in areas of strong collagenation (arrows). IFI anti-TGF ~ X 185
muscle cells of the Reisseisen muscle of the airways. TGF was also present in the hyperplastic epithelial cells of the airways, although the reaction was less intense than in the case of PDGF. In contrast with the findings in the other phases, the fibroblasts displayed weak TGF ~ immunoreac- tivity in the final phase in areas with a moderate amount of extracellular matrix. This reactivity remained limited even when additional fibrous components of extracellular matrix were deposited around the cells (Fig 7).
DISCUSSION
The study of fibrosis in human medicine and laboratory animals is generally directed to furthering the understand- ing of the causes of two of the fundamental phenomena underlying the process: fibroblastic proliferation and the deposition of collagen (Liebow 1975, Spencer 1975, Adamson and Bowden 1988, Barrios and Selman 1991, Harrison and Lanrent 1991).
The recent discovery of the growth factors and their effect on cellular proliferation and the deposition of extra- cellular matrix components (Davidson 1990, Selman and Barrios 1991, Kumar et al 1993, Puztai et al 1993) led the authors to consider their role in equine pulmonary fibrosis. It was considered that PDGF and TGF ~ were the growth factors which would probably have the greatest role in this disorder. However, little work has been done on growth factors and pulmonary fibrosis, and at times the results have been contradictory. It is probable that the diversity of the results is due to the fact that most studies of human lung fibrosis have been done in Vitro (Pantazis et al 1985, Roberts et al 1985, Assoian and Spore 1986, Bitterman et al 1986, Diegelmann et al 1986, Momex et al 1986, Assoian et al 1987, Martinet et al 1987, Osornio-Vargas et al 1991), and there has been little investigation of growth factors in situ in human lung samples by means of immuno- histochemJcal techniques (Richardson and Campion 1988, Antoniades et al 1991, Fabisiak et al 1992), and none at all in horses.
In this study, the samples of lung from horses with chronic pneumonia were divided into three phases on the basis of histological criteria. The usefulness of this classifi- cation was demonstrated later by the use of immunohisto- chemical techniques to detect PDGF and TGF ~, because these growth factors had a different distribution in each of the phases. In the first phase, the alveolar macrophages dis- played PDGF immunoreactivity. PDGF production by alveo- lar macrophages has been observed in vitro by several authors (Assoian et al 1987, Martinet et al 1987, Osornio- Vargas et al 1991) and confirmed by immunohistochemical techniques in human lung tissue (Mornex et al 1986, Antoniades et al 1991 ). In this study, the expression of PDGF by the macrophages was extremely heterogeneous, with very positive cells found next to cells which displayed hardly any reaction. Antoniades et al (1991) suggested that maximum positivity may be observed only in active macrophages, but Momex et al (1986) and Martinet et al (1987) reported that non-active macrophages also liberate significant quantities of PDGF.
The observation of fluorescence in the epithelial cells of the bronchi and bronchioles treated with anti-PDGF antibod- ies agrees with the investigations of Mornex et al (1986) and Martinet et al (1987) in human patients with idiopathic pulmonary fibrosis, and with the investigations of Antoniades et al (1991), who were the first to observe PDGF

86 A. Rodrfguez, M. Casta~o, L. Pe~a, M. A. Sdnchez, A. Nieto, M. Rodrfguez
in non-tumorous human epithelial cells. PDGF was expressed by respiratory epithelial cells in all three phases, although there was clearly an increase as the pulmonary fibrosis developed. The liberation of PDGF by the macro- phages in the earliest stages of equine interstitial pneu- monia induces fibroplastic proliferation, as Sugarman et al (1985) and Diegelmann et al (1986) indicated after study- ing alveolar macrophages in cultures from individuals with idiopathic pulmonary fibrosis. In turn, the fibroplastic pro- liferation causes increased deposits of proteins, chiefly fibronectin and type III collagen (Davidson 1990), from the extracellular matrix.
TGF 15 also appeared in the epithelial cells, although in the initial phase only in a small number of superficial epithelial cells of bronchioles with pronounced cellular hyperplasia. As in the case of PDGF, the number of discrete epithelial cells positive for TGF 13 in the bronchi and bron- chioles increased during the intermediate and final phases. The expression of TGF 13 was always weaker than that of PDGF, and its increases in the later phases were less evident. This finding has apparently not been previously recorded, and may be the result either of a pH reduction in vivo, as Richardson and Campion (1988) observed in pig fetuses, or of an imbalance in the epithelium/matrix relationship as a result of the mitogenic effect of PDGF on the fihroblasts, in agreement with the investigations of Mornex et al (1986) and Assoian et al (1987). This phenomenon leads to the expression of TGF ]3 by epithelial cells, which stimulates the production and accumulation of collagen; Bernard et al (1990) and Antoniades et al (1991) have made similar observations in pathological human cases.
The intermediate phase was characterised histologically by considerable proliferative activity, some degree of fibro- sis and the presence of intra-alveolar masses of connective tissue. By using indirect immunofluorence, it has been shown that the fibroblasts present in the alveolar masses adherent to the alveolar septa were PDGF-positive. PDGF production has been detected in fibroblasts in cell cultures (Bitterman et al 1986) and in fibrotic human lung tissue (Martinet et al 1987). The strong positive reactivity of the fibroblasts in the intra-alveolar masses can be explained as a consequence of the paracrine action of the PDGF in the macrophages on the fibroblasts, which induces them to pro- liferate (Chen et al 1992). Subsequently, they continue to proliferate owing to autocrine stimuli, as the fibroblasts themselves synthesise more PDGF.
In the final phase, characterised by total pulmonary fibrosis, the most significant finding was the production and release of PDGF and TGF I~ by fibroblasts in areas in dif- ferent phases of fibrotic evolution. Thus, in areas of young or immature fibrous tissue, with numerous active fibro- blasts, the fibroblastic production of PDGF was intense. In other more mature fihrotic areas, rich in fibre and with a limited number of old fibroblasts, the number and intensity of PDGF-positive cells decreased. In the authors' opinion, this could have been due to the fact thatproliferation is reduced in mature fibrotic areas and the large-scale produc- tion or deposition of collagen is not necessary. This sugges- tion agrees with the observations of Fabisiak et al (1992) in cultures of human pulmonary fibroblasts, which demon- strated the role of fibroblasts as producers not only of collagen but also of PDGF.
During this final phase, TGF 13 expression was also observed in fibroblasts, which was not the case in the earli- er phases. In this phase the fibroblasts in the connective tissue masses showed a discrete cytoplasmic fluorescence,
possibly as a result of an increased sensitivity towards the stimulus of numerous cytokines, in agreement with the findings of Davidson (1990).
TGF ~ was detected in epithelial cells in all three phases of the disease and in fihroblasts in the final phase, but it should be noted that Assoian et al (1987), when using immunohistochemical techniques in cell cultures, observed that platelets, macrophages and lymphocytes were the only producers of TGF ~. According to Roberts et al (1985), TGF 13 is a negative growth factor, the absence of which brings about an uncontrolled proliferation of fibroblasts. This phe- nomenon was observed in the present study in samples of lung in the first two phases of interstitial pneumonia; how- ever, in the final phase only a slight fluorescence was observed in the areas of fibroblastic proliferation, suggest- ing that, during this period, TGF ~ limits fibroblastic prolif- eration, while stimulating collagen production, as has been observed in in vitro studies (Ignotz and Massagu6 1986, Fine and Goldstein 1987, Ignotz et al 1987).
Finally, the results of this study indicate that the presence of PDGF favours the fibroblastic proliferation which takes place in cases of pulmonary fibrosis in horses. Deposits of collagen increase indirectly, as a result of the increase in the numbers of cells which produce it, and directly during the last phase, as a result of the stimulus of TGF ~ on each producing cell. These findings are in accordance with the results of human studies by Antoniades et al (1991) and Fabisiak et al (1992).
ACKNOWLEDGEMENT
This research was supported in part by the Complutense University of Madrid (Proyecto Multidisciplinar n ° 4835).
REFERENCES ADAMSON, I, Y. R. & BOWDEN, D, H. (1979) Bleomycin-indnced injury and
metaplasia of alveolar type 2 cells: Relationship of cellular responses to drug pres- ence in the lung. American Journal of Pathology 96, 531-542
ADAMSON, I. Y. R., YOUNG, L. & BOWDEN, D. H. (1988) Relationship of alve- olar epithelial injury and repair to the induction of pulmonary fibrosis. American Journal of Pathology 130~ 377-383
ANTONIADES, H. N., BRAVO, M. A., GALANOPOULOS, T. & SELMAN, M. (1991) Molecular basis for the role of platelet-derived growth factor in pulmonary fibrosis. In Interstitial Pulmonary Diseases: Selected Topics. Eds M. Selman and R. Barrios. Boca Rat6n (Florida), CRC Press. pp 35-46
ASSOIAN, R. K., FLEURDELYS, B. E., STEVENSON, H. C., MILLER, P. J., MADTER, D. K., RAINES, E. W., ROSS, R. & SPORN, M. B. (1987) Expression and secretion of type beta transforming growth factor by activated human macrophages. Proceedings of the National Academy of Science, USA 84, 6020-6024
ASSOIAN, R. K. & SPORN, M. B. (1986) Type [3 transforming growth factor in human platelets. Release during platelet degranulation and action on vascular smooth muscle cells. Journal of Cell Biology 102, 1217-1223
BARRIOS, R. & SELMAN, M. (1991) Morphological aspects of chronic interstitial lung diseases. In Interstitial Pulmonary Diseases: Selected Topics. Eds M. Selman and R. Barrios. Boca Rat6n (Florida), CRC Press. pp 197-228
BEN/TO, M. & LORENZO, M. (1993) Platelet derived growth factor/tyrosine kinase receptor mediated proliferation. Growth Regulation 3, 172-179
BERNARD, J. A., LYONS, R. M. & MOSES, H. L. (1990) The cell biology of transforming growth factor [k Biochemistry and Biophysics acta 1032, 79-87
BITTERMAN, P. B., WEWERS, M. D., RENNARD, S. I., ADELBERG, S. & CRYSTAL, R. G. (1986) Modulation of alveolar macrophage driven fibroblast proliferation by alternative macrophage mediators. Journal of Clinical Investigation 77, 700-705
BUBRGELT, C. D., HINES, S. A., CANTOR, G., STIRK, A. & WILSON, J. H. (1986) A retrospective study of proliferative interstitial long disease of horses in Florida. Veterinary Pathology 23, 750-756
CAGNY, P. & GOBERT, H.-J. (1880) Veterinary Dictionary IU. Madrid, I&P CHEN, L. B., POLUNOVSKY, V. WHITE, J., BLAZAR, B., NAKHLEH, R.,
JESSURON, J., PETERSON, M. & BtTI'bRMAN, P. (1992) Mesenchymal cells isolated after lung injury manifest as enhanced proliferative phenotype. Journal of Clinical Investigation 90, 1778-1785
CRYSTAL, R. G. (1987) Interstitial lung disorders. In Harrison's Principles of Internal Medicine. 1 lth edn. New York, Brannwold, McGraw-Hill. p 1095
CRYSTAL, R. G., BITTERMAN, P. B., RENNARD, S. I., HANCE, A. J. & KEOGH, B. A. (1984) Interstitial lung disease of unknown cause: Disorders char-

PDGF and TGF fl in equine chronic pneumon ia 87
acterized by chronic inflammation of the lower respiratory tract. New England Journal of Medicine 310, 235-244
DAVIDSON, J. M. (1990) Biochemisla-y and turnover of lung interstitium. European Respiratory Journal 3, 1048-1068
D1EGELMANN, R. F., SCHULLER-LEVIS, G., COHEN, I. K. & KAPLAN, A. M. (1986) Identification of a low molecular weight macrophage-derived chemotactic factor for fibroblasts. Clinical Immunology and Immunopathology 41, 331
DUNGWORTH, D. L. (1992) The Respiratory System. In Pathology of Domestic Animals. 4th edn. Eds K. V. F. Jubb, P. C. Kennedy and N. Palmer. San Diego, Academic Press. pp 539-699
FABISIAK, J. P., ABSHER, M., EVANS, J. N. & KELLEY, J. (1992) Spontaneous production of PDGF A chain homodimer by rat lung fibmblasts in vitro. American Journal of Physiology 263, 185-193
FINE, A. & GOLDSTEIN, R. H. (1987) The effect of transforming growth factor [~ on cell proliferation and collagen formation by lung fibroblasts. Journal of Biological Chemistry 262, 3897-3902
FROHNER, E. & ZWICK, G. (1932) Patolog/a y Terapeutica Veterinarias. 2nd edn. Ed Gustavo Girl. Barcelona
HARA, H., MANABE, T. & HAYASHI, T. (1989) An immunohistochemical study of the fibrosing process in paraquat lung injury. Virchows Archives A (Pathology and Anatomy) 415, 357-366
HARRISON, N. K. & LAURENT, G. J. (1991) Lung collagen metabolism: The link between inflammation and pulmonary fibrosis. In Interstitial Pulmonary Diseases: Selected Topics. Eds M. Selman and R. Barrios. Boca Rat6n (Florida), CRC Press. pp 47-74
IGNOTZ, R. A., ENDO, T. & MASSAGUI~, J. (1987) Regulation of fibronectin and type I collagen mRNA levels by transforming growth factor ~. Journal of Biological Chemistry 262, 6443-6446
IGNOTZ, R. A. & MASSAGUE. J. (1986) Transforming growth factor beta stimu- lates the expression of fibronectin and collagen and their incorporation into the extracellular matrix. Journal of Biological Chemistry 261, 4337-4345
KUMAR, R. K., O'GRADY, R., LI, W. & RAJKOVIC, 1. (1993) Secretion of epi- dermal growth factor-like molecular species by lung parenchymal macrophages: Induction by Interferon- 7. Growth Factors 9, 223-230
LAURENT, G. J. (1986) Lung eonagen: More than scaffolding. Thorax 41, 418 LIEBOW, A. A. (1975) Definition and classification of interstitial pneumonia in
human pathology. Progress in Respiratory Research 8, 1-33 MARTINET, Y., ROME, W. N., GROTENDORST, G. R., MARTIN, G. R. &
CRYSTAL, R. G. (1987) Exaggerated spontaneous release of platelet-derived growth factor by alveolar macrophages from patients with idiopathic pulmonary fibrosis. New England Journal of Medicine 317, 202
MENDOZA, A. E., YOUNG, R., ORKIN, S. H. & TUCKER, C. (1990) Increased platelet-derived growth factor A-chain expression in human uterine smooth mus- cle cells during the physiologic hypertrophy of pregnancy. Clinical Research 87, 2177-2181
MORNEX, J. F., MARTINET, Y. & YAMAMUCHI, K. (1986) Spontaneous expres- sion of the c-sis gene and release of a platelet-derived growth factor-like molecule by human alveolar rnacrophages. Journal of Clinical Investigation 78, 61-66
MORTON, L. F. & BARNES, M. J. (1982) Collagen polymorphism in the normal and diseased blood vessel wall. Investigation of collagens type I, III and V. Atherosclerosis 42, 41
NARAYANAN, A. S., ENGEL, L. D. & PAGE, R. C. (1983) The effect of chronic inflammation on the composition of collagen types in human connective tissue. Collagen and Related Research 3, 323 -330
NIEBERLE, L. & COHRS, P. (1967) Respiratory System. In Special Pathological Anatomy of Domestic Animals. Oxford, Pergamon Press. pp 169-288
OOSHIMA, A. (1981) Collagen ct B chains increased proportion in human
atherosclerosis. Science 213, 666 OSORNIO-VARGAS, A., BONNER, J. C. & BRODY, A. R, (1991) Rat lung
fibroblast (RFL) chemotaxis to PDGF-AA, AB y BB directs. International Conference Suppl. Abs. 143, A254
PANTAZIS, P., PELICCI, P. A., DALLA-FAVERA, R. & ANTONIADES, H. N. (1985) Synthesis and secretion of proteins resembling platelet-derived growth fac- tor by human glioblastoma and fibrosarcoma cells in culture. Proceedings of the National Academy of Science, USA 82, 2404
PUSZTAI, L., LEWIS, C. E., LORENZEN, J. & McGEE, J. O'd. (1993) Growth factors: Regulation of normal and neoplastic growth. Journal of Pathology 169, 191-201
RAGHU, G. & KAVANAGH, T. (1991) The human lung fibroblast: A multifaceted target and effector ceil. In Interstitial Pulmonary Diseases: Selected Topics. Eds M. Selman and R. Barrios. Boca Rat6n (Florida), CRC Press. pp 1-34
RICHARDSON, R. L. & CAMP/ON, D. R. (1988) Immunohistochemical locarlza- tion of transforming growth factor type ~ (TGF-b) in fetal pig subcutaneous adi- pose tissue. Journal of Animal Science 66 Supplement, 286-296
ROBERTS, A. B., ANZANO, M. A., WAKEFIELD, L. M., ROCHE, N. S., STERN, D. F. & SPORN, M. B. (1985) Type ~ transforming growth factor: A hiinnctional regulator of cellular growth. Proceedings of the National Academy of Science, USA 82, 119-123
ROBERTS, A., SPORN, M. B, ASSOIAN, R. K., SMITH, J. M., ROCHE, N. S., WAKEFIELD, L. M., LIOTTA, L. A., FALANGA, V., KEHRL, J. H. & FAUCI, A. S. (1986) Transforming growth factor ~: Rapid induction of fibrosis and angio- genesis in vivo and stimulation of collagen formation in vitro. Proceedbzgs of the National Academy of Science, USA 83, 4167-4171
SELMAN, M. (1990) Pulmonary fibrosis. Experimental and lung disease in connec- tive tissue. In Health and Disease. Boca Rat6n (Florida), CRS Press. Ch 5
SELMAN, M. & BARRIOS, R. (1991) Interstitial Pulmonary Diseases: Selected Topics. Boca Rat6n, (Florida), CRS Press. Chs 1, 2 and 3
SELMAN, M., MONTA2qO, M., MONFORT, L & PEREZ-TAMAYO, R. (1985) A new model of diffuse interstitial pulmonary fibrosis in rat. Experimental Molecular Pathology 43, 375
SELMAN, M., MONTAIqO, M., RAMOS, C., BARRIOS, R. & PEREZ-TAMAYO, R. (1989) Experimental pulmonary fibrosis induced by paraquat plus oxygen in rats: a morphologic and biochemical sequential study. Experimental Molecular Pathology 50, 147
SPENCER, H. (1975) Pathogcnesis of interstitial fibrosis of the lung. Progress in Respiratory Research 8, 34-44
SPORN, M. B., ROBERTS, A. B., WAKEFIELD, L. M. & DE COMBRUGGHE, B. (1987) Some recent advances in the chemistry and biology of transforming growth factor ~. Journal of Cell Biology 105, 1039-t045
SUGARMAN, B. J., ASGARWAL, B. B., HASS, P. E., FIGARI, I. S., PALLA- DINO, M. A. & SHEPARD, H. M. (1985) Recombinant human tumor necrosis factor ~: Effects on proliferation of normal and transformed cells in vitro. Science 230, 943
TORRE-AMIONE, G., GEAUCHAMP, R. D. & KOEPPEN, H. (1990) A highly immnnogenic tumor transfected with a murine transforming growth factor type 1 cDNA escapes immune surveillance. Proceedings of the National Academy of Science, USA 87, 1486-1490
VIEL, L. (1983) Structural-functional correlations of the lung in horses with small airway disease. Doctoral Thesis, Guelph (Canada)
WAHL, S. M., WAHL, L. M. & McCARTHY, J. B. (1987) Lymphocyte-mediated activation of fibroblast proliferation and collagen production. Journal of Immunology 121, 942
Received June 21, 1994 Accepted August 1, 1995





























