Embed Size (px)
Citation preview

European Journal of Neuroscience. Vol. 8, pp. 1061-1071, 1996 @I European Neuroscience Association
lmmunocytochemical Visualization of the mGluR1 a Metabotropic Glutamate Receptor at Synapses of Corticothalamic Terminals Originating from Area 17 of the Rat
Zoltan Vidnyanszky’, Tarnas J. Gorcsl, Laszlo Negyessyl, Zsolt Borostyankbi’, Rainer Kuhn2, Thomas Knopfe12 and Jozsef Harnori’ ’Laboratory of Neurobiology, 1 st Department of Anatomy, Semmelweis University Medical School, TOzolto u. 58, 1094 Budapest, Hungary 2Laboratory of Molecular and Cellular Biology, CNS, Ciba, Basel, Switzerland
Keywords: lateral geniculate nucleus, lateral posterior nucleus, Phaseolus vulgaris leucoagglutinin
Abstract
Pre-embedding immunogold histochemistry was combined with Phaseolus vulgaris leucoagglutinin anterograde tract tracing in order to analyse the relationship between the subcellular localization of the mGluRla metabotropic glutamate receptors and the distribution of corticothalamic synapses in the dorsal lateral geniculate nucleus (dLGN) and the lateral posterior nucleus (LP) of the rat. The injection of the tracer into area 17 labelled two types of corticothalamic terminals: (i) the small boutons constituting the majority of the labelled fibres which form asymmetrical synapses both in the dLGN and LP; and (ii) the giant terminals typically participating in glomerulus-like synaptic arrangements and found exclusively in the lateral posterior nucleus. The small corticothalamic terminals often established synapses with mGluR1 a-immunopositive dendrites, with immunometal particles concentrated at the periphery of their postsynaptic membranes. In contrast, the synapses formed by giant boutons in the lateral posterior nucleus were always mGluRl a-immunonegative. We conclude that the corticothalamic fibres forming the small synaptic terminals are the most likely candidates for the postulated mGluR-mediated modulation of visual information flow by corticothalamic feedback mechanisms.
Introduction
Transfer of visual information through the thalamus to the cerebral cortex is modulated by extraretinal afferents in a state-dependent manner so that, during arousal and attention, thalamic responsiveness is enhanced (for reviews see Sherman and Koch, 1986; Steriade and Llinas, 1988; McCormick, 1992). It has been proposed that this modulation is produced, in part, by corticothalamic feedback mechan- isms (McCormick, 1992; McCormick and von Krosigk, 1992) and, indeed, visual cortical projections provide numerous afferents to the thalamus (Wilson et al., 1984; Montero, 1991). In the rat, these projections originate mainly from the areas 17 and 18 and to a lesser extent from other extrastriatal areas (for reviews see Nauta and Bucher, 1954; Guillery, 1971; Szentigothai, 1973; Jones, 1985). All of these inputs are thought to use glutamate as their neurotransmitter (Baughman and Gilbert, 1980; Fonnum et al., 1981; Fosse and Fonnum, 1987; Montero, 1990).
Molecular biological, pharmacological and electrophysiological studies have shown that glutamate acts on two major classes of receptors: the ionotropic glutamate receptors, forming cation-specific ion channels, and the metabotropic glutamate receptors (mGluRs),
which couple to G proteins and modulate the concentration of intracellular messengers (for reviews see Nakanishi, 1992; Pin and Duvoisin, 1995).
The activation of corticothalamic fibres causes monosynaptic excita- tion of thalamic relay neurons through ionotropic receptors followed by a prolonged excitatory postsynaptic potential and increased excit- ability in the dorsal lateral geniculate nucleus (dLGN) (Deschtnes and Hu, 1990; McCormick and von Krosigk, 1992). The secondary, slow response can switch the firing of thalamic relay neurons from burst-firing mode to single-spike activity, a state associated with enhanced sensory transmission, arousal and attention. Electrophysio- logical studies by McCormick and von Krosigk (1992) have indicated that this slow depolarization and increase in excitability of the thalamic neurons is mediated by mGluRs.
Eight subtypes of the mGluR family (mGluR1-8) have been cloned and pharmacdlogically characterized in mammalian cell lines expressing cloned receptors. Based on their sequence similarities, intracellular signal transduction and sensitivity to agonists, mGluRs have been classified into three groups (for reviews see Nakanishi,
Correspondence to: T. J. Gorcs, as above
Received 23 December 1994, revised I3 April 1995, accepted 29 November 1995

1062 mGluR1 a at corticothalamic synapses
1992; Pin and Duvoisin, 1995). Group I includes mGluR1 and -5, which couple to phospholipase C and subsequent release of Ca’+ from intracellular stores. The members of group I1 (mGluR2 and -3) and group I11 (mGluR4, -6, -7 and -8) negatively couple to adenylate cyclase and thus slow down the production of CAMP.
Each of these mGluRs has a unique distribution in the central nervous system. In the rat thalamus, strong expression of the mRNAs coding for mGluRl was demonstrated by in situ hybridization (Masu et al., 1991; Shigemoto er al., 1992) and prominent expression of mGluRIa, a splice variant of mGluR1, was found in this region by immunohistochemistry (Martin ef al., 1992; Baude et al., 1993; Vidnyinszky et al., 1994). A recent study by Salt and Eaton (1994) suggested that mGluR1 is involved in the nociceptive responses of ventrobasal neurons of the rat thalamus.
Based on the above data, we have assumed that the corticothalamic activation of the dLGN and the lateral posterior nucleus (LP) neurons is mediated by mGluRla. To test this hypothesis, we have investigated the relationship between the subcellular localization of the mGluRla metabotropic glutamate receptor and the distribution of cortico- thalamic synapses in the dLGN and the LP of the rat using pre- embedding immunogold histochemistry combined with Phaseolus vulgaris leucoagglutinin (PHAL) anterograde tract tracing. The tracer was injected into area 17, the main source of the visual corticothalamic input. The corticothalamic projections from area 17 consist of two distinct populations of the fibres of different origin and termination regions (Mason and Groos, 1981; Jones, 1985; DeschCnes et al., 1994; Bourassa et al., 1995). The first population of fibres is a projection from the layer VI pyramidal cells terminating both in the dLGN and in the LP. The fibres of the second population are collaterals of the corticotectal projections of the layer V pyramidal cells ending exclusively in the LP (DeschCnes e l al., 1994; Bourassa er al., 1995), the thalamic relay of the extrageniculate pathway.
Materials and methods
Injections and surgery Four male Wistar rats weighing 250-300 g received unilateral injections of the anterograde tracer PHAL into the visual cortical area 17. The animals were deeply anaesthetized with an intramuscular injection of a mixture of ketamine hydrochloride (Ketavet, Bayer, 100 mg/kg body wt), xylazine (Rompun, Bayer, 2 mgkg body wt) and chlorpromazine (Hibernal, Egis, Hungary, 10 mgkg body wt) and then placed in a stereotaxic apparatus. The injection sites were determined according to the stereotaxic coordinates of the atlas of Paxinos and Watson (1986). The PHAL solution (2.5% in 0.05 M sodium phosphate buffer, pH 7.4) was iontophoretically injected, using a pulsed 5 pA positive current (7 s on-off cycle) for 15-20 min at each site.
Perfusion and immunohistochemistry Seven to 10 days after surgery. the four PHAL-injected animals and the four unoperated male rats were deeply anaesthetized with sodium pentobarbital ( 150 mgkg, i.p.) and transcardially perfused first with 50 ml saline (0.9% NaCI) then with fixative (two PHAL-injected and two unoperated rats with 4% paraformaldehyde, 0.1 % glutaraldehyde and 0.2% picric acid in 0.1 M phosphate buffer, pH 7.4, and the other four rats with 2% paraformaldehyde, 4% acrolein, 0.1 % glutaraldehyde and 0.2% picric acid in 0.1 M phosphate buffer) for 30 min. After perfusion, the brains were removed from the skull, and blocks of the regions investigated were dissected out. The blocks were rinsed several times in phosphate buffer, cryoprotected with 10
and 20% sucrose solution, quickly frozen in liquid nitrogen, and thawed. Coronal Vibratome sections (50 pm thick) were cut and washed with phosphate buffer. The sections from the animals perfused with acrolein-containing fixative were treated with 1% NaBH4 in distilled water for 30 min. Following extensive washes in phosphate buffer, sections from PHAL-injected and from control rats were sequentially incubated in 20% normal goat serum (Vector Laborat- ories) in phosphate buffer for 1 h at room temperature, and then in the mGluRla affinity-purified antibody (0.2 Fg IgG/ml) for 2 days at 4°C. The specificity of the antibodies used has been reported previously (Grandes et al., 1994). Sections from rats without PHAL injections were processed either with the avidin-biotin-peroxidase complex method (Vectastain ABC kit) using 3,3’-diaminobenzidine (DAB) as chromogen, or for pre-embedding immunogold labelling using anti-rabbit IgG coupled to 1 nm colloidal gold particles (AuroProbe One, Amersham) at dilutions ranging from 1 :25 to I 5 0 followed by silver intensification for 15-20 min, using the Intense I1 kit (Amersham), both as recommended by the manufacturer.
Sections from the PHAL-injected animals were exclusively pro- cessed with the immunogold technique for the mGluRla immunohisto- chemistry. For the double-labelling experiments, the mGluR 1 a- immunoreacted sections were incubated after extensive washes in biotinylated anti-PHAL (1:200; Vector) solution for 2 days at 4°C followed by the standard ABC protocol described above, using the DAB chromogen.
For light microscopy, sections were mounted onto gelatinized slides, dehydrated and coverslipped with DePex. For electron microscopy, sections were osmicated, dehydrated in ethanol, block-stained with 1 % uranyl acetate in 70% ethanol and embedded in Durcupan ACM (Fluka). For control experiments, 1 ml working dilution of antiserum was absorbed with 20 pg of synthetic peptide overnight at 4°C. and used for immunohistochemistry. In additional parallel experiments, incubation with primary antiserum was either omitted or replaced by preimmune rabbit serum.
Analysis of the distribution of mGluR 1 a immunoreactivity in postsynaptic dendrites of PHAL-labelled cortical terminals The distribution of mGluRla immunoreactivity (ir) at the synapses formed by PHAL-labelled cortical boutons and different morpho- logically identified terminals in the neuropil of the dLGN and the LP was evaluated on ultrathin sections cut from the surface of blocks, double-immunostained for mGluRla and PHAL. One pair of blocks containing dLGN or LP from three rats was used for analysis. Based on their ultrastructural features, different types of boutons were identified in the neuropil of the dLGN and LP (for review see Jones, 1985). In the dLGN, the RSD terminals are small in size, characterized by tightly packed and round synaptic vesicles and dark mitochondria and are mainly of cortical or midbrain origin. The RLP terminals originate from the retina and are characterized by round synaptic vesicles, large size and pale mitochondria. In the LP, small RS and large RL boutons, both containing round vesicles and originating mainly from the visual cortex, were identified. In the dLGN, the synaptic contacts of RLP, PHAL-labelled corticogeniculate RSD and PHAL-immunonegative RSD boutons were analysed. In the LP, the synapses formed by PHAL-labelled and unlabelled RS and RL terminals were analysed. The PHAL-labelled cortical boutons estab- lishing synaptic contacts were followed in serial sections and the immunogold particles overlying postsynaptic dendritic profiles were counted. The synapses formed by the PHAL-labelled boutons were considered mGluR1a-immunopositive if at least two immunogold particles were found perisynaptically, when the total surface of the postsynaptic membrane was examined in consecutive serial sections.

mGluRla at corticothalamic synapses 1063
FIG. I . Distribution of the mGluRla-ir and PHAL-labelled corticothalamic projections from cortical area 17 in the dLGN and LP at the light microscopic level. (A, B) Strong, predominantly neuropil, immunoperoxidase labelling of mGluRla-ir in the LGN and LP with the exception of the vLGPC and intergeniculate leaflet (IGL), where immunoreactivity is significantly weaker. (C) Frontal section through a representative PHAL injection placed in area 17. Arrowhead indicates the approximate border between areas 17 and 18. (D) Double labelling showing mCluRla-ir, as demonstrated by the pre-embedding immunogold technique, and PHAL tracing of corticothalamic fibres. PHAL-labelled dense axon arborization is seen in the mGluR1a-immunolabelled LGN and LP neuropil (arrows). At higher magnification, PHAL-labelled fibres with several small collateral boutons (arrowheads) in the LGN (E) and with a giant bouton (arrow) and also small terminals (arrowheads) in the LP (F) can be seen in the neuropil, which is labelled by dense immunometal particles representing mGluRla-ir. Scale bars I mm in A and C, 500 pn in D (also applies to B) and 10 pm in F (also applies to E).
Synaptic contacts of randomly selected, morphologically identified PHAL-immunonegative RSD and RLP boutons in the dLGN, as well as RS and RL terminals in the LP, were similarly analysed in the same sections.
The distribution of postsynaptic dendrites of the examined boutons based on their mGluRla-ir content was expressed as the sum of immunogold particles overlying the target profiles in three consecutive serial sections.
Results In the control experiments, both the preabsorption of the mGluRla antiserum with the synthetic peptide used for immunization and the
omission of the primary antibody resulted in the absence of specific labelling in immunoperoxidase, as well as in immunogold experi- ments, confirming the specificity of the immunocytochemical methods.
The distribution of the mGluRla-ir in the rat brain was similar to that described previously (Martin et al., 1992; Baude et al., 1993; Gorcs et al., 1993). Accordingly, the highest density of labelling was found in the molecular layer of the cerebellum and in the olfactory bulb. Both the LGN and LP exhibit strong mGluRla-ir. However, in contrast to other parts of the LGN, the ventral LGN parvocellular (vLGPC) and the intergeniculate leaflet were only very weakly labelled for rnGluRla (Fig. IA, B). In the dLGN and LP, a strong diffuse neuropil staining pattern was characteristic of mGluR1 a-ir (Fig. 1B).
1064 mGluRl a at corticothalamic synapses
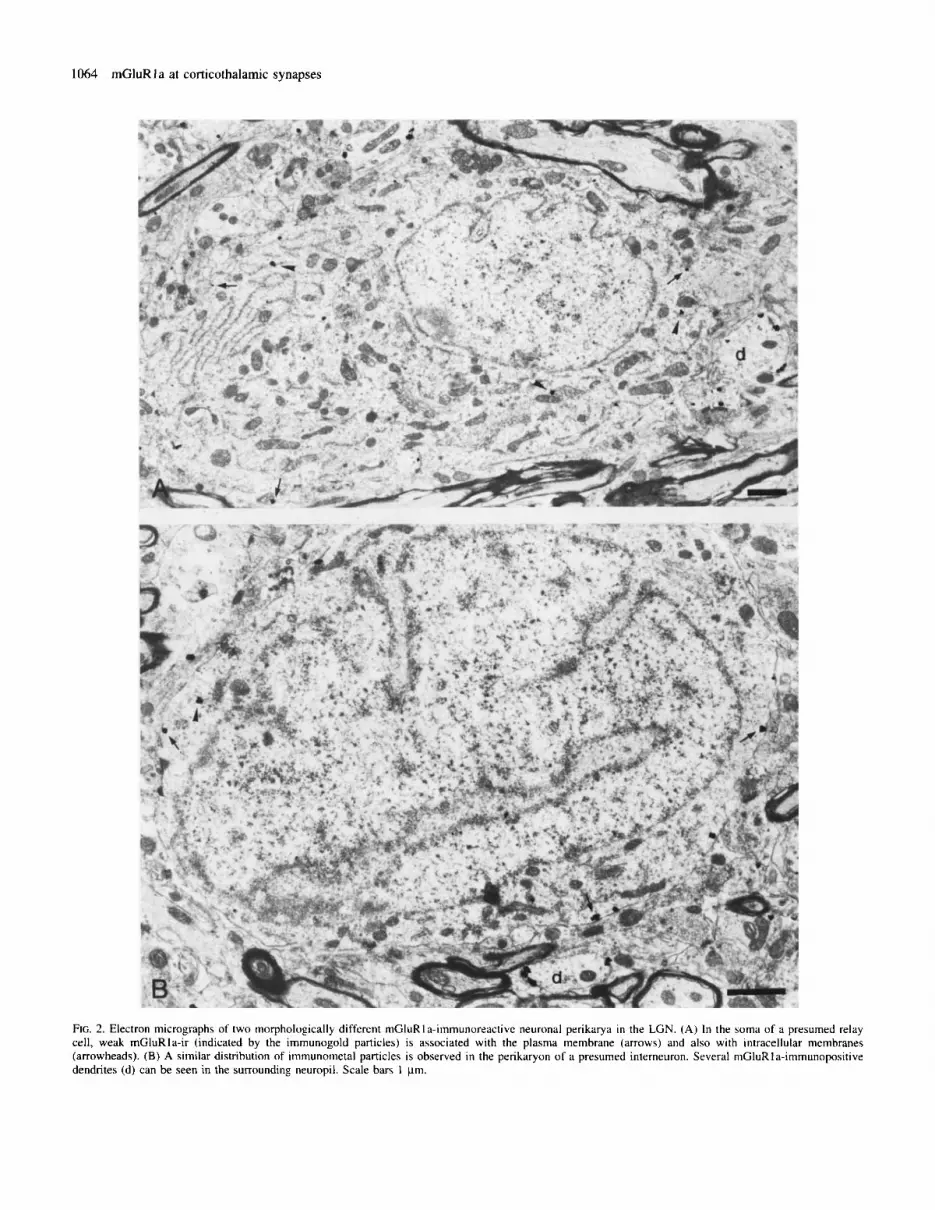
FIG. 2. Electron micrographs of two morphologically different mGluRIa-iinmunoreactive neuronal perikarya in the LGN. (A) In the soma of a presumed relay cell, weak rnGluRIa-ir (indicated by the immunogold particles) is associated with the plasma membrane (arrows) and also with intracellular membranes (arrowheads). (B) A similar distribution of immunometal particles is observed in the perikaryon of a presumed interneuron. Several niGluRla-immunopositive dendrites (d) can be seen in the surrounding neuropil. Scale bars I pm.

mGluRla at corticothalamic synapses 1065
FIG. 3. Subcellular localization of mGluRla-ir in the LGN and LP neuropil, as revealed by the pre-embedding immunogold technique. Serial sections showing the mGluRIa-immunopositive dendrites in the LGN ( A X ) and LP (D-F). In a dendritic profile (d l ) establishing asymmetrical synapses with small terminals, the immunometal particles are predominantly localized at the periphery of the synaptic junctions (arrows) and also extrasynaptically (arrowheads) ( A X ) . In the LP (D-F) similar pensynaptic (arrows) and extrasynaptic (arrowhead) localization of immunometal particles is present in a dendrite (d2) synapsing with a small terminal containing round vesicles. Scale bar 0.3 pm in F (applies to A-E).
In the case of simultaneous double labelling for mGluRla and PHAL-the latter being injected into the deep layers of the visual cortical area 17 (Fig. lC)-dense arborization of the PHAL-labelled corticothalamic axons was found in addition to the general staining by mGluRla in the dLGN and LP (Fig. 1D). These arborizations appeared as dark patches superimposed on the mGluR la-immunoreac- tive neuropil. The PHAL-labelled corticothalamic axons from area 17 gave rise to two morphologically distinct types of terminals. The small terminals connected by short stalks to the main axon shaft constituted the majority of PHAL-labelled fibres, and were found in both the dLGN and the LP (Fig. IE, F). The second type of boutons
was found exclusively in the LP. These were giant boutons (Fig. IF) which occurred both as boutons terminaux and en passant.
At the electron microscopic level, using the pre-embedding silver- intensified immunogold method, mGluR 1 a-ir was associated with neuronal membranes. In the mGluR 1 a-immunopositive neuronal perikarya, the immunometal particles were mainly associated with plasma membranes as well as intracellular membranes, including the Golgi apparatus and the endoplasmic reticulum (Fig. 2A, B). Similar weak perikaryal labelling was observed in both types of neurons in the dLGN, identified on the basis of the cytoplasmic ultrastructure of the perikarya (Lieberman, 1973; Pasik er al.. 1973): (i) projection

1066 mGluR1 a at corticothalamic synapses
FIG. 4. Relationship between synapses formed by corticothalamic terminals from area 17 and the subcellular localization of the mGluR1a-ir in the LGN. A PHAL-labelled corticothalamic bouton establishing asymmetrical synapses with an mGluRl a-immunopositive dendrite (d I ) is shown in three consecutive serial sections (A-C). Immunometal particles are localized at the periphery of the cortical synapses (arrows). In two serial sections (D. F) the asymmetrical synapse between a PHAL-labelled cortical terminal and an mGluR I a-immunopositive dendrite (d2)+ontdining immunogold particles in extrasynaptic position-is devoid of immunoreactivity. A presumed retinal terminal (RT) establishes several puncta adherentes with an unlabelled large-calibre dendritic profile. Scale bar 0.3 pm in C (also applies to A, B) and 0.5 pm in E (also applies to D).
neurons, with well-developed, rough endoplasmic reticulum and large dark mitochondria (Fig. 2B). In the mGluR1a-labelled dendrites, pale mitochondria (Fig. 2A) and (ii) presumed intemeurons, with immunometal particles were predominantly concentrated at the pale cytoplasm, few cisterns of endoplasmic reticulum and small, periphery of postsynaptic membranes of asymmetrical synapses (Fig.

mGluRla at corticothalamic synapses 1067
FIG. 5. Relationship between synapses formed by the corticothalamic terminals from area 17 and the subcellular localization of the mGluR1a-ir in the LP. (A, B) A small PHAL-labelled corticothalamic terminal synapsing with an mCluRIa-immunoreactive dendrite (d l ) is shown in two consecutive serial sections. Immunometal particles are associated with the periphery of the subsynaptic membrane (arrows) of the labelled bouton. On serial sections (C, D) a PHAL- labelled giant bouton establishes synaptic contact with an mGluRla-immunonegative dendrite (d2). In an adjacent position, an unlabelled small terminal forms a synapse with an mGluRla-immunopositive dendrite (d3) in which immunometal particles are localized both pen- and extrasynaptically. Scale bars 0.5 pn.

1068 mGluRla at corticothalamic synapses
TABLE 1. Proportion of the mGluRla-immunopositive synaptic contacts formed by different boutons in the dLGN neuropil
Synaptic terminals Synaptic contacts formed by different boutons in:
block id. F94.5.1 block id. P94.7.1 block id.-P94.8.2 Total
n mGluRIa-ip (Yo) n mGluR1a-ip (Yo) n mCluR1a-ip (%) n mGluRla-ip ('3)
RSD 25 5 (20) 25 4 (16) 25 5 (20) 7s 14 (18.6) RSD (PHAL-labelled) 22 4 (18.2) 21 3 (14.3) 16 3 ( 18.75) 59 10 (16.9) RLP 15 0 (0) 15 0 (0) 15 0 (0) 4 s 0 (0)
n, number of synapses examined; mGluR1a-ip, mGluR1a-immunopositive synapses
TABLE 2. Proportion of the mGluRIa-immunopositive synaptic contacts formed by different boutons in the LP neuropil
Synaptic terminals Synaptic contacts formed by different boutons in:
block id. P94.5.3 block id. P94.7.5 block id. P94.8.4 Totdl
n mGluR1a-ip (96) n rnGluR1a-ip (%) n rnGluRla-ip (8) n mGluRla-rp ('3)
RS 25 5 (20) 25 3 (12) 25 4 (16) 75 12 (16) RS (PHAL-labelled) 17 3 (17.6) 14 2 (14.3) 20 3 (15) 51 8 (15.7) RL 15 0 (0) 15 0 (0) 15 0 (0) 45 0 (0) RL (PHAL-labelled) 12 0 (0) 11 0 (0) 14 0 (0) 31 0 (0)
n, number of synapses examined; mGluR la-ip, mGIuR1a-immunopositive synapses.
3A-F). No immunometal particles were found in association with the main body of postsynaptic densities. In the mGluRl a-immunoreactive dendrites, staining of the extrasynaptic membranes was also observed in cases where synapses were not labelled or no synapses were found in the vicinity.
Terminals of different morphological features form synapses with the mGluRla-immunoreactive dendrites. However, a specific peri- synaptic localization of the immunometal particles was observed only at the asyinmetrical synapses formed by small-calibre axon terminals containing round vesicles (Fig. 3A-F). These boutons in the dLGN and LP exhibited the ultrastructural features of the RSD and RS terminals respectively.
The quantitative analysis of the distribution of the mGluRla-ir at the synapses formed by different types of the PHAL-labelled and unlabelled boutons in the double-stained material supported the above observation (Tables 1 and 2). It appeared that 16.9% of synaptic contacts of the PHAL-labelled corticogeniculate RSD boutons in the dLGN and 15.4% of the synapses formed by PHAL-labelled small RS corticothalamic terminals in the LP were mGluRla-irnmuno- positive, i.e. immunometal particles were found at the periphery of their postsynaptic densities (Figs 4A-C and SA, B). In some cases, the dendrites postsynaptic to the small PHAL-labelled corticothalamic terminals contained immunometal particles exclusively extra- synaptically, while the cortical synapses were devoid of labelling (Fig. 4D, E). The proportions of the mGluRla-immunopositive synapses as a percentage of the total synaptic contacts established by the PHAL-unlabelled, morphologically identified RSD terminals in the dLGN and as RS terminals in the LP, were 18.6 and 16% respectively. These ratios are similar to those found in the case of PHAL-labelled small cortical boutons.
In the LP, the PHAL-labelled giant terminals formed asymmetrical synapses, often in glomerulus-like synaptic arrangements, with large dendritic profiles (Fig. 5C, D). The immunometal particles indicating mGluR1a-ir, however, were never observed at the subsynaptic membranes and only very rarely at the extrasynaptic membranes of
the dendrites targeted by giant cortical boutons in the LP (Tables 1 and 2). Similarly, no perisynaptic labelling and, in most cases, not even extrasynaptic labelling for the mGluRla were observed in the dendrites postsynaptic to the retinal RLP terminals in the dLGN (Fig. 4D, E).
The analysis of the number of immunogold particles in the dendrites postsynaptic to the PHAL-labelled and unlabelled RSD terminals and in the target dendrites of the RLP retinal terminals in the dLGN revealed that significantly more mGluRla-ir is associated with dendrites that receive input from the small cortical RSD terminals than with dendrites that receive RLP retinal boutons (Fig. 6A). Similar distribution of the mGluRla was found in the LP, where immunogold particles were associated mainly with the dendrites postsynaptic to the small RS terminals, while the target dendrites of the giant RL cortical terminals were, in most cases, devoid of silver particles (Fig. 6B). What makes the observed preferential localization of the mGluRla-ir in the postsynaptic dendrites of the small cortical terminals even more conspicuous is the fact that the mean diameter of the distal, corticorecipient dendrites, postsynaptic to the small cortical terminals (RSD, RS) is significantly smaller than that of those mainly proximal dendrites participating in synaptic connections with the retinal RLP terminals in the dLGN or with the large RL corticothalamic boutons in the LP (Jones, 1985; Sherman and Koch, 1986).
Discussion
The distribution of mGluRla immunoreactivity in the rat LGN and LP
Using immunohistochemistry, it is demonstrated in the present study that the two main visual thalamic nuclei, the dLGN and LP, express high levels of mGluRla. Strong, diffuse neuropil labelling for mGluRla was found in both the LGN and the LP, with the exception of the vLGPC and intergeniculate leaflet, while in the perikarya

mGluR1 a at corticothalamic synapses 1069
A
W RSD HRSD (PHAL
ORLP labelled)
2s
g 20
g 15
: 10
- ._ L '0
U r, 0
a
2 5 E,
0 0 1 2 3 4 5 6 1
Number of immunogold particles
B
25 T RS mRS (PHAL-
labelled)
labelled) 0 RL (PHAL-
0 1 2 3 4 5 6 7
Number of immunogold particles
FIG. 6. Distribution of dendritic profiles postsynaptic to different types of synaptic terminals based on their content of immunogold particles representing mCluRla-ir in the dLGN and LP of the rat. No significant differences were found between the distribution of the dendrites postsynaptic to PHAL-labelled and unlabelled RSD terminals in the dLGN (x2 = 0.72, P > 0.99) and the distribution of PHAL-labelled and unlabelled RS boutons in the LP (x2 = 1.848, P > 0.975). Similarly, the distribution of dendrites postsynaptic to the PHAL-labelled RSD terminals in the dLGN and the target dendrites of the PHAL-labelled RS terminals in the LP are not different ( x 2 = 2.565, P > 0.95). However, the distribution of the dendrites postsynaptic to RLP terminals in the dLGN and PHAL-labelled RL cortical boutons in the LP is different from the distribution of the dendrites postsynaptic to PHAL-labelled small cortical terminals, RSD and RS ( x 2 = 20.627, P < 0.01 in the dLGN and x2 = 22.512, P < 0.01 in the LP).
moderate cytoplasmic staining could be detected. In previous immuno- histochemical studies, similar, predominantly neuropil labelling for mGluRla was found in several brain regions, including the thalamus (Martin et al., 1992) and for mGluR5 in the striatum and hippocampus (Shigemoto et al., 1993). However, in other brain regions, such as the hippocampus and cerebellar cortex, strong somatic and proximal dendritic labelling for mGluRla was also reported (Martin et al., 1992; Baude et al., 1993; Grandes et al., 1994). The functional significance of these differences in the subcellular distribution of mGluRl a in different brain regions remains to be elucidated.
At the electron microscopic level, the pre-embedding silver- intensified immunogold technique revealed an exclusively neuronal membrane-associated localization of mGluR 1 a-ir in the dLGN and LP. Immunoreactivity for mGluRl a was predominantly found in small-calibre dendrites, while axons were always immunonegative. The subcellular localization of mGluR1a-ir in the dLGN and LP was similar to those described previously in the hippocampus and cerebel- lar cortex (Baude et al., 1993). Immunometal particles were localized at the periphery of asymmetrical synapses, and also extrasynaptically. Specific labelling at the postsynaptic dendritic membranes was only present in the asymmetrical synapses formed by the small RSD and RS terminals. About 15-20% of the synapses formed by the RSD terminals in the dLGN and RS terminals in the LP were mGluR1a- immunopositive. The proportion of the mGluR1a-immunopositive synapses might be significantly underestimated due to the technical limitations of the pre-embedding silver-intensified immunogold method, such as the penetration of the antibody, the accessibility of the antigenic sites and the unpredictable loss of silver particles during osmication. The postsynaptic dendrites targeted by large, presumably primary afferent, terminals, often forming a glomerular (RLP terminals in the dLGN) or glomerulus-like (RL terminals in the LP) synaptic arrangement, were found to be mostly mGluR1a-immunonegative. The postsynaptic membranes of the primary afferent synapses were always devoid of mGluRla-ir. This observation is in agreement with the electrophysiological recordings, which demonstrate that, unlike corticothalamic fibre stimulation, optic tract stimulation did not induce mGluR-mediated responses in the dLGN (McCormick and von Krosigk, 1992).
The localization of mGluRla at the periphery of the identified corticothalamic synapses in the LGN and LP McCormick and von Krosigk (1 992) proposed that the prolonged excitatory postsynaptic potential in guinea-pig dLGN neurons evoked by activation of corticogeniculate afferents is mediated by mGluRs. Their study did not address questions about the mGluR subtype and the morphological identity of the corticothalamic projections mediating this response. According to previous immunohistochemical studies with different mGluR antibodies (Vidnyhszky et al., 1994), the most likely candidate for visual thalamic nuclei is mGluRla. Indeed, the cellular and subcellular localization of mGluRla in the dLGN and LP, as demonstrated in the present study, suggests that this receptor is predominantly localized in the dendrites postsynaptic to an identified population of the PHAL-labelled corticothalamic ter- minals.
In fact, after the injection of PHAL into layer V-VI of area 17, two morphologically distinct types of corticothalamic terminals were labelled. The majority of fibres originate in the cortical layer VI pyramidal cells and end in the dLGN and LP as small boutons (Mason and Groos, 1981; Jones, 1985; Deschtnes et al., 1994). These small terminals establish asymmetrical synapses, the small-calibre dendrites showing a high level of mGluRla-ir. We found that immunometal particles were most abundant at the periphery of the postsynaptic densities of these synapses.
Possible functional significance
We have found that two morphologically distinct corticothalamic projections exhibit a clear difference in their set of postsynaptic glutamate receptors, suggesting different functional properties.
The first type of PHAL-labelled terminals originated from the collaterals of the corticotectal axons of layer V pyramidal cells of the area 17 (Deschtnes et al., 1994; Bourassa et al., 1995). and is

1070 mGluRla at corticothalamic synapses
confined to the LP. They are giant boutons, typically establishing glomerulus-like synaptic arrangements with large-calibre (probably proximal) dendrites. Their postsynaptic dendrites were usually devoid of mGluR1a-ir, at both the subsynaptic and the extrasynaptic mem- brane. Previously, a feed-forward, primary afferent function of this cortical giant terminal projection to non-specific thalamic nuclei has been suggested in visual (Robson and Hall, 1977), somatosensory (Hoogland et al., 1991) and auditory (Rouiller and Welker, 1991) systems. This model might be extended by the present data showing that postsynaptic membranes to giant terminals in the LP, similar to retinal terminals in the LGN and in contrast to the corticothalamic projections forming small terminals, are mGluRl a-immunonegative. The giant cortical terminals, similar to retinal endings, might release their transmitter exclusively at the ionotropic receptors producing fast synaptic transmission. We cannot, however, exclude the expression of other mGluR subtypes and splice variants not yet identified at these synapses.
The second type of terminals originated from the corticothalamic fibres of the layer VI pyramidal cells (Mason and Groos, 1981; Jones, 1985; Deschtnes er al., 1994) and are of smaller diameter. These boutons were found in two thalamic visual centres with distinct functions in processing sensory input (Jones, 1985) but exhibited very similar morphological features and a common pattern of postsynaptic mGluRla localization. An attractive hypothesis is that these projec- tions are the substrate of an mGluR 1 a-mediated, state-dependent modulation of thalamic information flow (McCormick and von Krosigk, 1992). The mGluR-mediated slow depolarization of thalamic relay neurons described by McCormick and von Krosigk (1992) occurs only with the delivery of several electrical corticothalamic stimuli at intervals as short as 20 ms. Thus, repeated high-frequency stimulation might be required for released glutamate to overcome uptake mechanisms and to reach the perisynaptically localized mGluRl a.
Acknowledgements We thank Mrs Zsuzsa Szab6 for excellent technical assistance. This work was supported by National Scientific Research Fund (OTKA) grants F-6066, 2617, 1107 andT016164, and National Health Council Fund (ETT) grant T-O4491/93.
Abbreviations ABC DAB dLGN ir LGN LP mGluR PHAL RSD
RLP
RS
RL
vLGPC
avidin-biotin-peroxidase complex 3.3‘-diaminobenzidine dorsal lateral geniculate nucleus immunoreactivity lateral geniculate nucleus lateral posterior nucleus metabotropic glutamate receptor Phaseolus vulgaris leucoagglutinin boutons in the dLGN neuropil characterized by round synaptic vesicles, small size and dark mitochondria boutons in the dLGN neuropil characterized by round synaptic vesicles, large size and pale mitochondria boutons in the LP neuropil characterized by round synaptic vesicles and small size boutons in the LP neuropil characterized by round synaptic vesicles and large size ventral LGN, parvocellular part
References Baude, A,, Nusser, Z., Roberts, J. D. B., Muhihill, E., McIlhinney, R. A.
J. and Somogyi, P. (1993) The metabotropic glutamate receptor (mGluRla) is concentrated at perisynaptic membrane of neuronal subpopulations as detected by immunogold reaction. Neuron, 11, 771-787.
Baughman, R. W. and Gilbert. C. D. (1980) Aspartate and glutamate as possible neurotransmitters of cells in layer 6 of the visual cortex. Ntrrure.
Bourassa, J.. Pinaullt, D. and Deschenes. M. (1995) Corticothalamic projections from the cortical barrel field to the somatosensory thalamus in rats: a single-fibre study using biocytin as an anterograde tracer. Eur. J. Neurosci., 7, 19-35.
Deschenes, M. and Hu, B. (1990) Electrophysiology and pharmacology of the corticothalamic input to lateral thalamic nuclei: an intracellular study in the cat. Eur. J. Neurosci.. 2, 140-152.
Deschenes., M., Bourassa, J. and Pinault. D. (1994) Corticothalamic projections from layer V cells in rat are collaterals of long-range corticofugal axons. Brain Res.. 664, 215-219.
Fonnum, F., Storm-Mathisen, J. and Divac, I. (1981) Biochemical evidence for glutamate as neurotransmitter in corticostriatal and corticothalamic fibres in rat brain. Neuroscience, 6 , 863-873.
Fosse, V. M. and Fonnum, F. (1987) Biochemical evidence for glutamate and/or aspartate as neurotransmitter in fibres from the visual cortex to the lateral posterior thalamic nucleus (pulvinar) in rats. Brain Res.. 400. 2 19-224.
Gorcs, T. J., Penke, B., Bdti, Z., Katarova, Z. and Hamori. J. (1993) Immunohistochemical visualization of a metabotropic glutamate receptor. NeuroReport, 4, 283-286.
Grandes, P., Mateos, J. M., Riiegg, D., Kuhn, R. and Knopfel. T. (1994) Differential cellular localization of three splice variants of the mGluRl metabotropic glutamate receptor in rat cerebellum. NeuroReport. 5. 2249-2252.
Guillery, R. W. (1971) Patterns of synaptic interactions on the dorsal lateral geniculate nucleus of cat and monkey: a brief review. Vision Res., 3(Suppl.), 21 1-227.
Hoogland, P. V., Wouterlood, F. G.. Welker, E. and Van der Loos, H. (1991) Ultrastructure of giant and small thalamic terminals of cortical origin: a study of the projections from the barrel cortex in mice using Phaseolus vulgaris leuco-agglutinin (PHAL). Exp. Brain Res.. 87. 159-1 72.
Jones, E. G. (1985) The Thalamus. Plenum Press, New York. Lieberman. A. R. (1973) Neurons with presynaptic perikarya and presynaptic
dendrites in the rat lateral geniculate nucleus. Brain Res.. 59. 35-60. Martin, L. J., Blackstone, C. D., Huganir, R. L. and Price, D. L. (1992)
Cellular localization of a metabotropic glutamate receptor in rat brain. Neuron, 9, 259-270.
Mason, R. and Groos, G. A. (1981) Cortico-recipient and tecto-recipient visual zones in the rat’s lateral posterior (pulvinar) nucleus: an anatomical study. Neurosci. Left., 25, 107-1 12.
Masu, M., Tanabe, Y., Tsuchida, K., Shigemoto, R. and Nakanishi, S. (1991) Sequence and expression of a metabotropic glutamate receptor. Nature, 349, 760-765.
McCormick, D. A. (1992) Neurotransmitter actions in the thalamus and cerebral cortex and their role in neuromodulation of thalamocortical activity. Prog. Neurobiol., 39, 337-388.
McCormick, D. A. and Von Krosigk, M. (1992) Corticothalamic activation modulates thalamic firing through glutamate ‘metabotropic’ receptors. Proc. Natl Acad. Sci. USA, 89. 2774-2778.
Montero, V. M. (1990) Quantitative immunogold analysis reveals high glutamate levels in synaptic terminals of retino-geniculate, cortico- geniculate and geniculo-cortical axons in the cat. Ks. Neurosci.. 4, 437443.
Montero, V. M. (1991) A quantitative study of synaptic contacts on interneurons and relay cells of the cat lateral geniculate nucleus. Exp. Brain Res., 86, 257-270.
Nakanishi, S. (1992) Molecular diversity of glutamate receptors and implications for brain function. Science. 258, 597-603.
Nauta, W. J. H. and Bucher, V. M. (1954) Efferent connections of the striate cortex in the albino rat. J. Comp. Neurol., 100, 257-285.
Pasik, P., Pasik, T., Hamori, J. and Szentagothai, J . (1973) Golgi type I1 interneurons in the neuronal circuit of the monkey lateral geniculate nucleus. Exp. Brain Res. , 17, 18-34.
Paxinos, G. and Watson, C. (1986) The Rat Brain in Sfereotaxic Coordinafes. Academic Press, New York.
Pin, J.-P. and Duvoisin, R. (1995) Review: neurotransmitter receptors. I. The metabotropic glutamate receptors: structure and functions. Neuropharmacology, 34, 1-26.
Robson, J. A. and Hall, W. C. (1977) The organization of the pulvinar in the grey squirrel (Sciurus carolinensis). 11. Synaptic organization and comparisons with the dorsal lateral geniculate nucleus. J. Comp. Neurol.. 173, 389416.
287, 848-850.

Rouiller. E. M. and Welker. E. (1991) Morphology of corticothalamic terminals arising from the auditory cortex of the rat: a P h a s e o h i,u/garis-leucoagglutinin (PHAL) tracing study. Hearing Res., 56, 179-190.
Salt. T. E. and Eaton. S. A. (1994) The function of metabotropic excitatory amino acid receptors in synaptic transmission in the thalamus: studies with novel phenylglycine antagonists. Neurochem. Int., 24, 451-458.
Sherman, S. M. and Koch, C. (1986) The control of retinogeniculate transmission in the mammalian lateral geniculate nucleus. Exp. Brain Res., 63, 1-20.
Shigemoto, R., Nakanishi, S. and Mizuno. N. (1992) Distribution of the mRNA for a metabotropic glutamate receptor (mGluRI) in the central nervous system: an in situ hybridization study in adult and developing rat. J . Comp. Neurof., 322, 121-135.
mGluR1 a at corticothalamic synapses 1071
Shigemoto, R., Nomura, S., Ohishi. H., Sugihara, H., Nakanishi, S. and Mizuno, N. (1993) Imrnunohistochemical localization of a metabotropic glutamate receptor, mGluR5, in the rat brain. Neurosci. Lett., 163, 53-57.
Steriade. M. and Llinb. R. R. (1988) The functional states of the thalamus and the associated neuronal interplay. Physiol. Rev., 68, 649-742.
Szentigothai, J. (1973) Neuronal and synaptic architecture of the lateral geniculate nucleus. In Jung, R. (ed.), Handbook of Sensory Physiology, Vol. VW3, Part B. Springer, Berlin, pp. 141-176.
Vidnyanszky, Z., Himori, J., Kuhn, R., Ntgyessy, L. and Gorcs, T. J. (1994) Immunocytochemistry of rnetabotropic glutamate receptors in the visual system of the rat. Eu,: J . Neurosci. (Suppl.), 7 , 193.
Wilson, J. R., Friedlander, M. J. and Sherman, S. M. (1984) Fine structural morphology of identified X- and Y-cells in the cat’s lateral geniculate nucleus. Proc. Natl Acad. Sci. USA, 221, 411436.


















![[2018] SGHC 61 Originating Summons No 1002 of 2017](https://img.pdfslide.net/doc/110x75/6321d127aa6c954bc7076d05/2018-sghc-61-originating-summons-no-1002-of-2017-.jpg)










