Embed Size (px)
Citation preview

Metabotropic Glutamate Receptor Expression inOlfactory Receptor Neurons from the ChannelCatfish, Ictalurus punctatus
Kathryn F. Medler, Hang N. Tran, James M. Parker, John Caprio, Richard C. Bruch
Department of Biological Sciences, Louisiana State University, Baton Rouge, Louisiana 70803
Received 20 August 1997; accepted 1 December 1997
(S-4CPG), a specific antagonist to mGluR1, each sig-ABSTRACT: Metabotropic glutamate receptorsnificantly reduced olfactory receptor responses to L-(mGluRs) were identified in olfactory receptor neu-glutamate. S-4CPG and MCCG reduced the gluta-rons of the channel catfish, Ictalurus punctatus, bymate response to 54% and 56% of control, respec-polymerase chain reaction. DNA sequence analysistively, which was significantly greater than their effectconfirmed the presence of two subtypes, mGluR1 andon a neutral amino acid odorant, methionine. ThesemGluR3, that were coexpressed with each other andsignificant reductions of odorant response by the an-with the putative odorant receptors within single ol-tagonists, taken with the expression of these receptorsfactory receptor neurons. Immunocytochemical datathroughout the dendritic and ciliated portions of someshowed that both mGluR subtypes were expressed inolfactory receptor neurons, suggest that these mGluRsthe apical dendrites and some cilia of olfactory neu-may be involved in olfactory reception and signalrons. Pharmacological analysis showed that antago-transduction. q 1998 John Wiley & Sons, Inc. J Neurobiol 35:nists to each mGluR subtype significantly decreased94–104, 1998the electrophysiological response to odorant aminoKeywords: electrophysiology; immunocytochemistry;acids. a-Methyl-L-CCG1/(2S,3S,4S)-2-methyl-2-metabotropic glutamate receptors; olfactory recep-(carboxycyclopropyl)glycine (MCCG), a known an-tors; PCRtagonist to mGluR3, and (S)-4-carboxyphenylglycine
INTRODUCTION tus) found that odorant amino acids interacted withspecific receptor sites in isolated olfactory cilia andthat these receptors were coupled to G-proteinsOlfactory transduction in vertebrates is initiated by(Bruch and Kalinoski, 1987; Bruch and Rulli,the binding of stimuli to olfactory receptor proteins1988). Interaction of the receptors with stimuli re-located in the dendritic cilia of olfactory receptorsults in the activation of either adenylate cyclaseneurons. Molecular cloning techniques have shownor phospholipase C by G-protein subunits. Thesethat the putative odorant receptors belong to aactions cause the formation of either cAMP or phos-multigene family of the G-protein coupled receptorpholipid-derived second messengers (Breer et al.,superfamily (Ngai et al., 1993b). Previous ligand-1994; Bruch, 1996). These second messengers ulti-binding studies in channel catfish (Ictalurus puncta-mately control the gating of ion channels involvedin membrane depolarization, which leads to the pro-duction of action potentials and synaptic transmis-Correspondence to: K. Medler
Contract grant sponsor: NIH; contract grant number: sion to the central nervous system. These secondDC01500 messengers are also involved in the termination ofContract grant sponsor: NSF; contract grant number: IBN-
the olfactory signal by stimulating second-messen-9221891q 1998 John Wiley & Sons, Inc. CCC 0022-3034/98/010094-11 ger dependent protein kinases (Bruch et al., 1997).
94
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

mGluRs in Olfactory Neurons 95
based on amino acid homology and pharmacologicalprofile. Group I receptors comprise mGluR1 andmGluR5 subtypes which stimulate phospholipase Cactivity. Group II includes mGluR2 and mGluR3,which are negatively coupled to adenylate cyclase.Group III receptors include the subtypes mGluR4,mGluR6, mGluR7, and mGluR8, which are also neg-atively coupled to adenylate cyclase. Members of thisfamily of G-protein coupled receptors have a seven-transmembrane domain region, but have no sequencehomology to other known G-protein coupled recep-tors. These receptors are large (871–1199 aminoacids) compared to other G-protein receptors andpossess a large extracellular N-terminal domain(Schoepp, 1994). Splice variants have been foundin some of these receptors and primarily affect the
Figure 1 Sequence of mGluR PCR products amplified intracellular carboxyl tail after the seven-transmem-from olfactory epithelium. Two products were found that brane region of the protein. To date, splice variantsshared a high degree of sequence similarity to the rat have been found in the mGluR1, mGluR4, andmGluRs (mGluR1 was 83% identical to rat and mGluR3
mGluR5 subtypes (Pin and Duvoisin, 1995). Therewas 72% identical to rat) . Differences between the se-are several unique features of the mGluR family thatquences are indicated with boldface type. Roman numer-distinguish it from the rest of the G-protein coupledals indicate membrane-spanning regions.receptor superfamily. One feature conserved acrossthe family is an area in the extracellular domain andextracellular loops that is cysteine rich. AnotherAmino acids, bile acids, and steroid hormones
are potent olfactory stimuli in the channel catfish unique feature of this receptor family is that the gluta-mate binding site is located in the extracellular do-as in other aquatic organisms. It is known that the
amino acid L-glutamate is an olfactory stimulus in main instead of within the transmembrane domain(Conn and Pin, 1997). Finally, there is evidence fromcatfish (Caprio et al., 1989) as well as an important
neurotransmitter in the central nervous system of recent studies that at least some mGluRs exist in vivoas disulfide-linked dimers (Romano et al., 1996).vertebrates (Pin and Duvoisin, 1995). Recent stud-
ies indicate that glutamate may also be acting as While the family of putative odorant G-proteincoupled receptors has been characterized in sev-the primary neurotransmitter in the olfactory bulb
(Berkowicz et al., 1994) and the vomeronasal organ eral vertebrate species, its presence in the olfac-tory system does not exclude the possibility that(Dudley and Moss, 1995). In the Atlantic salmon,
Pang et al. (1994) described the ligand-binding other families of G-protein linked receptors mayalso have important functions in olfaction. In thischaracteristics of a glutamate receptor found in ol-
factory epithelium and found that L-glutamate study, we investigated the expression of mGluRsin the olfactory rosette and in olfactory receptorbinding was specific and was inhibited by known
agonists and antagonists of metabotropic glutamate neurons of the channel catfish. We were inter-ested in characterizing which subtypes were ex-receptors. Pang et al. (1994) also showed that gluta-
mate stimulated IP3 formation in a manner consis- pressed in the olfactory rosette and where thesereceptors were localized within olfactory recep-tent with G-protein linked activation of phospholi-
pase C (PLC). Northern blot analysis of olfactory tor neurons. Based on the salmon study (Pang etal., 1994 ) showing that activation of a glutamatetissue RNA resulted in cross-hybridization with me-
tabotropic glutamate receptor (mGluR) probes to receptor in the olfactory system led to PIP2 hy-drolysis, we wanted to know whether these recep-transcripts of 1.9 and 3.0 kb, which indicated that
a receptor belonging to the metabotropic glutamate tors were expressed in channel catfish in whichthe primary response to olfactory stimuli is thereceptor family was present in the olfactory epithe-
lium of the Atlantic salmon (Pang et al., 1994). activation of PLC (Bruch, 1996 ) . Molecularcloning analysis of PCR products indicated thatMetabotropic glutamate receptors are a multigene
family of G-protein linked receptors that consist of mGluR1 and mGluR3 subtypes were expressedin olfactory receptor neurons. Immunocytochem-eight subtypes, which are divided into three groups
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

96 Medler et al.
MATERIALS AND METHODS
Reverse-Transcriptase PolymeraseChain Reaction (RT-PCR) Analysis ofmGluR Expression in Olfactory Tissue
Initial experiments were done on total RNA isolated fromolfactory rosettes using Trizol Reagent (Gibco, GrandIsland, NY) per the reagent’s protocol. Random primedRT was performed as previously described (Bruch andMedler, 1996). After termination of the reaction, thecDNA was used for PCR amplification as described byAbe et al. (1992). Degenerate primers were made to theamino acid sequence of the transmembrane segments IIand V of the mGluR family. The 5* primer was 5*-TCI-AGICGIGA(A/G)CTITG(C/T)TA(C/T)AT and the3* primer was 5*-TTICGIGT(C/T)TT(A/G)AA(A/C/G/T)GC(A/G)TA. All PCR analyses were initiatedwith Taq polymerase at 907C. The cDNA was then ampli-fied according to the following schedule: 947C for 1 min,427C for 2 min, and 727C for 2 min for 50 cycles, fol-lowed by 727C for 15 min with termination at 947C for15 min. The samples were slowly cooled to 47C. Negativecontrols were performed which lacked RT.
An aliquot of the PCR product was electrophoresedon a 2% agarose gel and the amplified DNA (approxi-mately 450 bp) was excised and gel-purified using theFigure 2 Sequence of PCR products of putative odorantQIAEX II Gel Extraction kit (Qiagen, Chatsworth, CA).receptors amplified from olfactory receptor neurons thatThis product was ligated into pCR2.1 (Invitrogen, Sanwere coexpressed with the mGluRs. The italicized se-Diego, CA), transformed into DH5a-competent cells,quence is the published sequence of a putative odorantand plated on LB plates with 50 mg/mL ampicillin andreceptor (Ngai et al., 1993b). The next six sequences1 mg/plate Xgal. Plasmids with inserts were identifiedwere from a single olfactory receptor neuron and wereby restriction digestion with EcoRI. Positive plasmidscoexpressed with both mGluR1 and mGluR3 subtypes.were grown in LB broth cultures and purified using col-The last three sequences are products expressed in an-umns from Qiagen (Chatsworth, CA) per the vendor’sother olfactory receptor neuron that also expressedprotocol. Products were sequenced on both strands usingmGluR1. Roman numerals indicate membrane-spanningthe Fidelity DNA Sequencing system (Oncor, Gaithers-regions. Differences found between the sequences areburg, MD) and were identified by comparison to the se-indicated with boldface type. All sequences were ú90%quences in the databanks using the BLAST program (Na-identical with the published putative odorant receptor se-tional Center for Biotechnology Information, Bethesda,quence.MD). The Gene Runner program was used to obtainthe predicted amino acid sequences (Hastings Software,Hastings, NY).
istry showed that both mGluR1 and mGluR3 sub-types were localized in the apical dendrites and Olfactory Neuron Isolationsome cilia of olfactory receptor neurons. Thesereceptors were molecularly characterized in sin- Channel catfish, 15–20 cm in length, were deeply anes-
thetized in MS-222 (1:5000) and sacrificed. Olfactorygle isolated olfactory receptor neurons and wererosettes were removed and placed in fish Ringer’s solu-found to be coexpressed with odorant receptorstion in a Sylgard-coated 35-mm plastic dish. Olfactoryin the same cells. We also report the novel obser-neurons were obtained following the methodology of Re-vation that antagonists specific to mGluR1 andstrepo and Teeter (1990). Rosettes were bathed in cal-
mGluR3 subtypes individually reduced olfactorycium and magnesium-free fish Ringer’s solution and were
receptor neural activity to the odorant L-gluta- subsequently dissociated in calcium and magnesium-freemate, to 55% of normal. Taken together, these fish Ringer’s solution containing 30 U/mL activated pa-results suggest that mGluRs may have a chemo- pain and 100 mg/mL gentamicin. The tissue was bathed
with this solution every 90 s for 15 min. Dissociationsensory function in the catfish olfactory system.
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

mGluRs in Olfactory Neurons 97
was terminated using fish Ringer’s solution with 2 mMCaCl2 , 10 mg/mL leupeptin, and 100 mg/mL gentamicin.Olfactory receptor neurons were visualized by phase-con-trast microscopy and were identified by their morphologi-cal characteristics (Bruch and Medler, 1996). Each cellwas individually removed using suction attached to a 20-mm-bore siliconized (Sigmacote, Sigma Chemical Co.,St. Louis, MO) microcapillary. Single neurons wereplaced into 100 mL of lysis buffer [1% Igepal, 10 mMTris/HCl, pH 8, 140 mM NaCl, 5 mM KCl, 5 mM dithio-threitol (DTT) and 40 U RNAsin] and frozen on dry ice.Immediately prior to analysis, tubes were centrifuged at12,000 1 g for 2 min to remove cellular and nucleardebris. The supernatant was removed to a fresh tube andused for RT-PCR.
RT-PCR Analysis of mGluR Expressionin Olfactory Receptor Neurons
To determine which glutamate subtypes were foundwithin olfactory receptor neurons, individual neuronswere analyzed. To further reduce the possibility of geno-mic contamination, the Dynabeads mRNA DIRECT kit(DYNAL, Oslo, Norway) was used to isolate poly(A)/
RNA from each neuron per the vendor’s protocol. Theresulting RNA on the Dynabeads was eluted in sterilewater and used for RT. After termination of the RT,the cDNA from each cell was divided equally for PCRanalysis. Negative controls lacking RT were also per-formed with the analysis. Half of the cDNA from eachcell was used to amplify glutamate receptors as describedabove, and the other half of the cDNA was used to am-plify odorant receptors (Ngai et al., 1993b), except thatthe 727C extension step was shortened to 1.5 min and atotal of 40 cycles was used. Products from two cells weresubcloned and analyzed by DNA sequence. Productsfrom subsequent analyses were characterized by Southernblot analysis using specific probes to identify glutamateand odorant receptor PCR products.
Southern Blotting
Figure 3 Southern blots of PCR products obtained from An aliquot of the receptor PCR products from isolatedindividual olfactory receptor neurons. (Upper) Metabo- neurons were electrophoresed on a 2% agarose gel andtropic glutamate receptor PCR products amplified from transferred to a nylon membrane overnight with 101 SSC.half the cDNA of a single neuron and hybridized with a The membrane was baked for 1 h at 807C and prehybridizedknown glutamate receptor probe. Lane 1-positive control; for 2 h at 427C with buffer [50% formamide, 0.02% sodiumlane 2-negative control; lanes 3 and 4-PCR products fromsingle olfactory receptor neurons. Truncated PCR prod-ucts are present in both sample lanes as well as the posi-tive control lane. The positive control was a cloned PCRproduct that had been sequenced and had previously been negative control; lanes 3 and 4-PCR products from singleshown to be an mGluR subtype. Film was exposed to olfactory receptor neurons. Film was exposed to the che-the chemiluminescent blot for 5 min. (Lower) Putative miluminescent blot for 3 min to develop the positiveodorant receptor PCR products amplified from the same control and for 15 min to develop lanes 2–4. The positiveneurons as the mGluRs and hybridized with a known control was a cloned PCR product that had been se-odorant receptor probe. Lane 1-positive control; lane 2- quenced and shown to be a putative odorant receptor.
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

98 Medler et al.
dodecyl sulfate (SDS), 0.1% N-lauroylsarcosine, 2% (Ohishi et al., 1994). This antibody was shown to reactwith mGluR2 and mGluR3 and was graciously providedblocking reagent, 51 SSC]. Hybridization was performed
overnight at 427C with digoxigenin-labeled cDNA probes by R. Shigemoto (Kyoto University) .(10 ng/mL in buffer) prepared from a glutamate receptoror odorant receptor PCR product obtained from olfactoryrosettes. The probes were gel-purified and labeled with
In Vivo Electrophysiologydigoxigenin overnight with the Klenow enzyme (Boeh-ringer Mannheim, Indianapolis, IN).
Each fish preparation was performed as previously de-After hybridization, the membranes were washedscribed (Caprio et al., 1989; Caprio, 1995). A metal-twice for 5 min each with 21 SSC/0.1% SDS at 427Cfilled glass micropipette was plated with platinum (balland twice for 15 min each with 0.51 SSC/0.1% SDS atdiameter Å 20 mm; impedance Å 30 KV) . The electrode,657C. Hybrids were detected using alkaline phosphatase-r.c.-coupled (220 pF capacitor, 20 MV resistor) to oneconjugated antibody to digoxigenin and chemilumines-input grid of a high impedance probe, was placed againstcent substrate CSPD (Boehringer Mannheim).the surface of the epithelium of a single olfactory lamella.The other active input was grounded and connected tothe reference electrode (hypodermic needle) embeddedmGluR Protein Localizationin the flank musculature. The multiunit activity was am-plified (bandpass 30–300 Hz), integrated (0.5 s riseMetabotropic glutamate receptor protein localization was
performed by immunocytochemistry. Olfactory rosettes time) and displayed on an oscilloscope and pen recorder.The response magnitude was measured in millimeterswere fixed by immersion in 4% paraformaldehyde in 0.1
M phosphate-buffered saline (PBS) for 4 h at 47C. The of the integrated phasic displacement from baseline andstandardized to the response of the standard, either L-tissue was then washed three times each for 15 min in
PBS, quenched in 0.1 M glycine for 30 min at room glutamate or L-methionine.Charcoal-filtered artesian tapwater (pH 8.5) was directedtemperature, and cryoprotected with serial changes of
10%, 20%, and 25% sucrose in PBS for 1 h each at 47C. by polyethylene tubing to a glass capillary connected bypolyethylene tubing to the olfactory organ which continu-The tissue was embedded in OCT compound and 12-mm
horizontal sections were cut on a cryostat and mounted on ously bathed (10 mL/min) the olfactory mucosa. Stimulussolutions were drawn hydrostatically from a disposable bea-silanized slides for immunocytochemical staining. Slides
were placed in 0.1% SDS in PBS for 20 min at room ker and delivered through a second tube leading to thecapillary. During odorant testing, the flow of water to thetemperature and were then treated with 0.25% H2O2 in
PBS for 30 min at room temperature. Nonspecific protein- olfactory organ was instantly replaced with the test solutionfor 5 s by the activation of an electronic time switch. Atbinding sites were blocked with diluted normal goat se-
rum with 10 mg/mL avidin (Sigma) at 377C for 30 min. the end of 5 s, the flow of water to the capillary was instantlyresumed. This stimulus delivery system provided for theThe primary antibodies were diluted with this normal
serum and left on the slides overnight at 47C. The next presentation of the odorant without a change in either pres-sure or temperature and with no dilution. The pH (8.5) ofday, the diluted secondary antibody was placed on the
slides for 30 min at 377C, followed by a 45-min incuba- the gill irrigation water, the stimulus delivery water, andtest solutions were equilibrated to the pond water where thetion with newly made ABC reagent (Vector Laboratories,
Burlingame, CA) at 377C. All steps were followed by fish were obtained.The antagonist used for subtype mGluR3 was the groupthree 5-min washes with PBS except for the blocking
step. Binding was visualized using the DAB peroxidase II mGluR antagonist [a-methyl-L-CCG I/(2S,3S,4S)-2-methyl-2-(carboxycyclopropyl)glycine](MCCG)substrate kit (Vector Laboratories) as per the vendor’s
protocol for 5 min. (Tocris, Ballwin, MO). (S)-4-Carboxyphenylglycine[(S)-4CPG] was used as the antagonist for mGluR1 sub-The primary antibody to mGluR1 was obtained from
Upstate Biotechnology (Lake Placid, NY) and was pro- type (Tocris) . Glutamate antagonists were diluted to 0.1mM, pH balanced in charcoal-filtered tapwater to pH 8.5,duced to a 21-residue synthetic peptide (KPNVTYASV-
ILRDYKQSSSTL) corresponding to the C-terminus of and applied directly to the olfactory epithelium for 1min. Prior to the application of the antagonist, 1 mM L-mGluR1 with an additional lysine added at the N-termi-
nus. The primary antibody to mGluR5 was also from methionine and 1 mM L-glutamate were tested as stan-dards. Following standardization, one antagonist was pre-Upstate Biotechnology and was produced to a 21-residue
synthetic peptide (KSSPKYDTLIIRDYTNSSSSL) cor- sented continuously to the olfactory organ through thestimulus delivery system for 1 min followed immediatelyresponding to the C-terminus of mGluR5 with an addi-
tional lysine added to the N-terminus. The mGluR group by the test of L-glutamate for 5 s. The antagonist wasresumed for 10 s followed by a 5 s presentation of L-II antibody (Ohishi et al., 1994) was raised against a
fusion protein containing a C-terminal portion of mGluR2 methionine and then a resumption of the water flow with-out antagonist for 4 min. Following this, the test schemeand had 80% amino acid sequence homology with the
corresponding C-terminal residues 855–879 of mGluR3 was repeated for the second antagonist.
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio
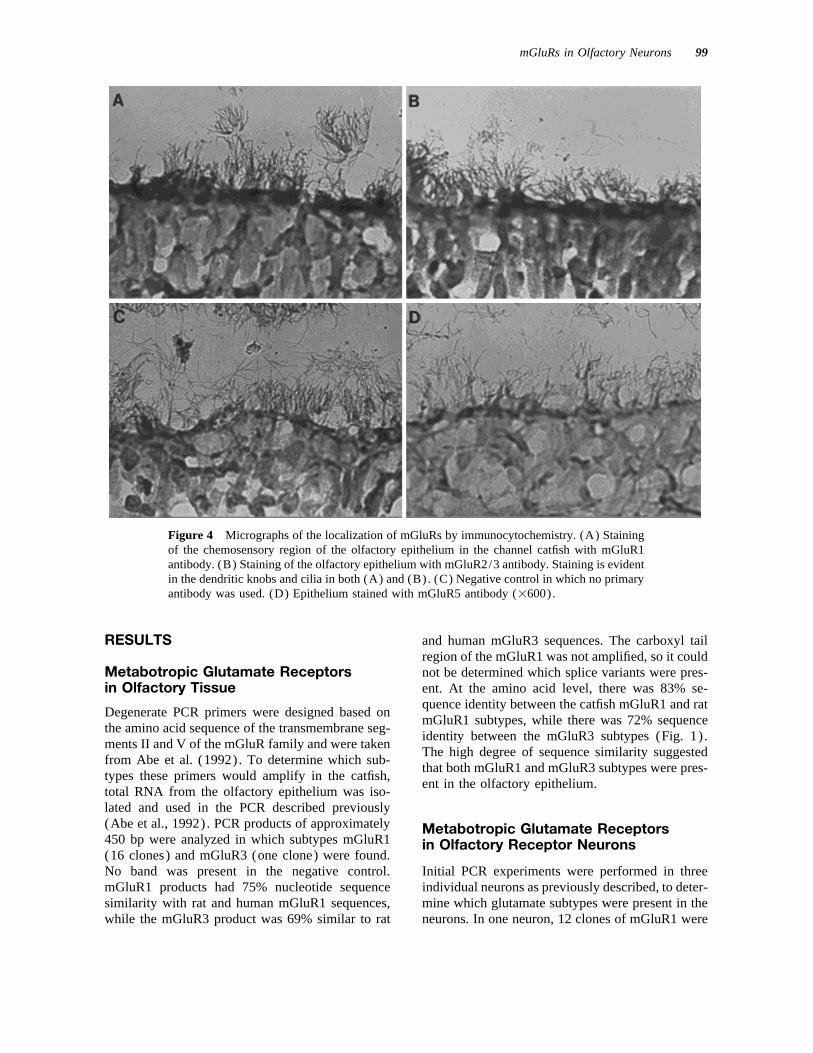
mGluRs in Olfactory Neurons 99
Figure 4 Micrographs of the localization of mGluRs by immunocytochemistry. (A) Stainingof the chemosensory region of the olfactory epithelium in the channel catfish with mGluR1antibody. (B) Staining of the olfactory epithelium with mGluR2/3 antibody. Staining is evidentin the dendritic knobs and cilia in both (A) and (B). (C) Negative control in which no primaryantibody was used. (D) Epithelium stained with mGluR5 antibody (1600).
RESULTS and human mGluR3 sequences. The carboxyl tailregion of the mGluR1 was not amplified, so it couldnot be determined which splice variants were pres-Metabotropic Glutamate Receptors
in Olfactory Tissue ent. At the amino acid level, there was 83% se-quence identity between the catfish mGluR1 and rat
Degenerate PCR primers were designed based onmGluR1 subtypes, while there was 72% sequence
the amino acid sequence of the transmembrane seg-identity between the mGluR3 subtypes (Fig. 1) .
ments II and V of the mGluR family and were takenThe high degree of sequence similarity suggested
from Abe et al. (1992). To determine which sub-that both mGluR1 and mGluR3 subtypes were pres-
types these primers would amplify in the catfish,ent in the olfactory epithelium.
total RNA from the olfactory epithelium was iso-lated and used in the PCR described previously(Abe et al., 1992). PCR products of approximately Metabotropic Glutamate Receptors450 bp were analyzed in which subtypes mGluR1 in Olfactory Receptor Neurons(16 clones) and mGluR3 (one clone) were found.No band was present in the negative control. Initial PCR experiments were performed in three
individual neurons as previously described, to deter-mGluR1 products had 75% nucleotide sequencesimilarity with rat and human mGluR1 sequences, mine which glutamate subtypes were present in the
neurons. In one neuron, 12 clones of mGluR1 werewhile the mGluR3 product was 69% similar to rat
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

100 Medler et al.
sequenced, another neuron had seven mGluR1 and of an antibody specific for mGluR5, specific stain-ing was also absent [Fig. 4(D)] .three mGluR3 clones identified, while in the third
neuron one mGluR3 and four mGluR1 subtypeswere sequenced. Results indicated that both ElectrophysiologymGluR1 and mGluR3 subtypes were found in theolfactory receptor neurons and that both subtypes Multiunit recordings within the olfactory rosette of
channel catfish were performed to determine ifcan be expressed within a single cell. Negative con-trol lanes showed no bands present. mGluRs are involved in olfactory responses. In this
technique, electrical responses are recorded fromSubsequent experiments were performed to de-termine if glutamate receptors were coexpressed the surface of several olfactory neurons simultane-
ously. Antagonists specific to certain mGluR sub-with the putative odorant receptors within individualneurons. After RT on individual neurons was com- types were used. S-4CPG is a known antagonist for
mGluR1 subtypes (Pin and Duvoisin, 1995), whileplete, the cDNA was divided into two parts. Onehalf of the cDNA was used to amplify the odorant in the rat spinal cord, MCCG has been shown to
act as an antagonist to Group II mGluR subtypesreceptors, and the other half was used to amplifythe glutamate receptors found in the same cell. In (Jane et al., 1994). Recent studies have shown that
MCCG is a more potent antagonist for mGluR3 astwo neurons, the PCR products were subcloned andanalyzed by sequence to determine which receptors compared to mGluR2 (McCool et al., 1996). Each
antagonist was individually applied to the olfactorywere present. In one neuron, three odorant receptorclones and four mGluR1 clones were identified. In epithelium and its effect on the L-glutamate induced
response was measured. These experiments werethe second neuron, six odorant receptor clones, fivemGluR1 clones, and one mGluR3 clone were identi- duplicated with an L-methionine standard as a con-
trol for nonspecific effects of the antagonists. Re-fied (Fig. 2) . Subsequent PCR products obtainedfrom 10 additional individual neurons were ana- sponses to glutamate in the presence of antagonist
were first compared to the standard L-glutamate re-lyzed by Southern blotting to determine if the PCRproducts were odorant or glutamate receptors (Fig. sponse in the absence of antagonist and were ana-
lyzed by Wilcoxon signed rank test. Both antago-3). Southern blotting was performed under stringentconditions using dioxigenin-labeled odorant recep- nists significantly reduced the glutamate response
(p Å 0.02 for both) and also significantly reducedtor or glutamate receptor probes. Previous experi-ments indicated that the probes did not cross-hybrid- the methionine response (p Å 0.02 for both) as
compared to the methionine standard. Representa-ize to each other (data not shown). The PCR prod-ucts from each neuron hybridized to their respective tive recordings are shown in Figure 5.
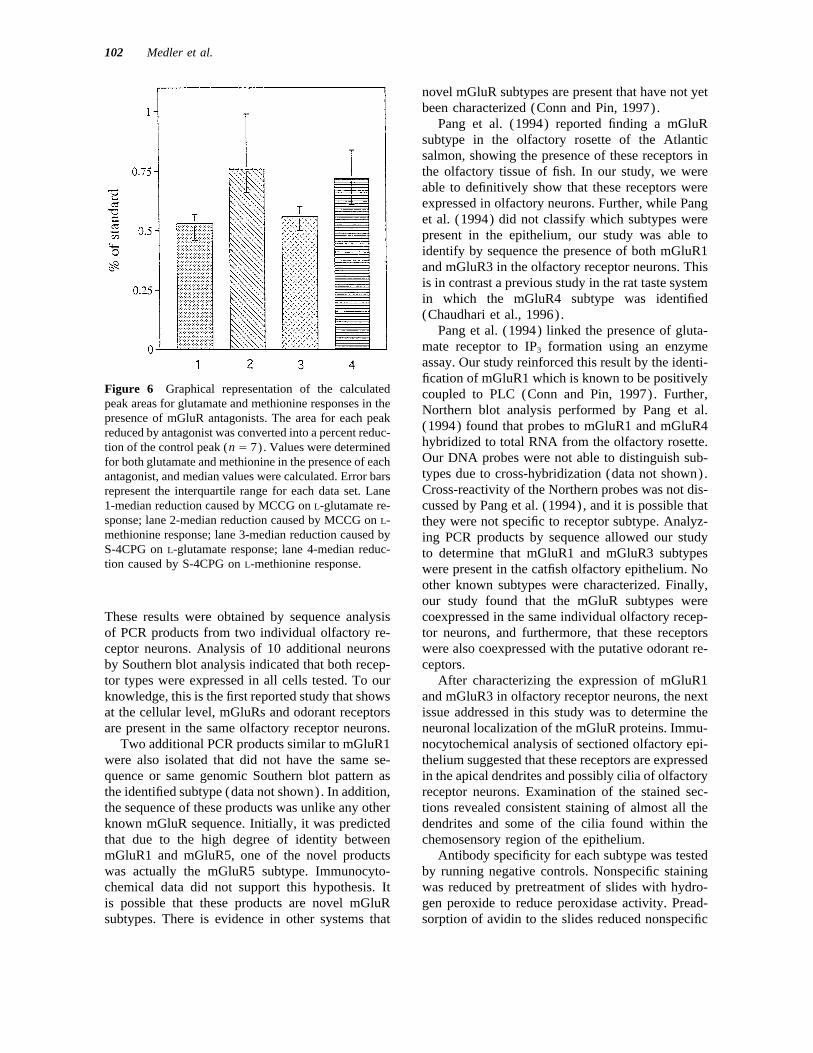
Glutamate and methionine responses with antag-probes, indicating that both the putative odorant re-ceptors and glutamate receptors were amplified onist were then expressed as percent reductions of
the standard response for each stimulus for bothfrom a single neuron in all neurons tested.antagonists. Methionine was reduced to 73% of thestandard response by S-4CPG and to 76% of stan-dard response by MCCG, while the glutamate re-mGluR Protein Localizationsponse was reduced to 54% of standard by S-4CPGand 56% by MCCG (Fig. 6) . Wilcoxon signed rankTo localize the area of the neuron that contained the
glutamate receptor protein, immunocytochemistry tests for each antagonist were performed betweenglutamate and methionine. Significant differenceswas performed using antibodies specific to the
mGluR1 and Group II subtypes. Specific labeling between odorant receptor responses to glutamateand to methionine were found for each antagonistwas observed in the dendritic knobs and some cilia
in the chemosensory region for both subtypes 1 and (p Å 0.02 for both) .3 [Fig. 4(A,B)] . Staining for both subtypes wasapproximately equal, although the staining was lessintense in the cilia. Nonspecific staining was mini- DISCUSSIONmized by pretreatment of the slides with both hydro-gen peroxide to remove endogenous peroxidase ac- Degenerate PCR primers were designed based on
the amino acid sequence of the transmembrane seg-tivity and avidin to bind to endogenous biotin pres-ent. In the absence of primary antibody, no specific ments II and V of the mGluR family (Abe et al.,
1992). PCR products of approximately 450 bp werestaining was evident [Fig. 4(C)] . In the presence
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

mGluRs in Olfactory Neurons 101
Figure 5 Effect of the glutamate antagonists S-4CPG (left) and MCCG (right) on integratedolfactory receptor responses to L-glutamate (top) and L-methionine (bottom). C-water control;A-L-glutamate standard; B-L-glutamate with S-4CPG; D-L-glutamate with MCCG; A *-L-methi-onine standard; B*-L-methionine with S-4CPG; D*-L-methionine with MCCG.
obtained from olfactory rosettes that corresponded in the same neurons. In each RT-PCR reaction doneon individual cells, precautions were taken to mini-to subtypes mGluR1 (16 clones) and mGluR3 (one
clone). The carboxyl tail region of the mGluR1 was mize genomic DNA contamination. First, prior tothe analysis, each tube containing the lysed neuronnot amplified, so it could not be determined which
splice variants were present. At the amino acid was centrifuged to remove the nucleus. Second,poly-A/ RNA was used as a template in RT-PCR.level, there was 83% sequence identity between the
catfish mGluR1 and rat mGluR1 subtypes, while Negative controls lacking RT did not produce PCRproducts, indicating a lack of genomic DNA con-there was 72% sequence identity between the
mGluR3 subtypes. The high degree of sequence tamination.Another aspect of this study was the novel deter-similarity suggested that both mGluR1 and mGluR3
subtypes were present in the olfactory epithelium. mination that the mGluRs were coexpressed in thesame olfactory receptor neurons with the putativePCR products of the appropriate size were not ob-
tained from negative controls lacking RT, sug- odorant receptors. Individual neurons were isolatedand underwent RT. The cDNA was then split intogesting that mGluR PCR products were not ampli-
fied from genomic DNA. two tubes and PCR analysis was performed on each.One tube was used to amplify the putative odorantTo characterize the cell types expressing the
mGluR subtypes in the olfactory epithelium, indi- receptors, while the other tube was used to amplifymGluRs. This analysis indicated that mGluRs andvidual olfactory receptor neurons were analyzed.
RT-PCR analysis of individual cells indicated that odorant receptors were expressed in the same cells.Furthermore, both mGluR1 and mGluR3 were ex-both subtypes mGluR1 and mGluR3 were present
in olfactory receptor neurons and were coexpressed pressed in the same cells as the odorant receptors.
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio
102 Medler et al.
novel mGluR subtypes are present that have not yetbeen characterized (Conn and Pin, 1997).
Pang et al. (1994) reported finding a mGluRsubtype in the olfactory rosette of the Atlanticsalmon, showing the presence of these receptors inthe olfactory tissue of fish. In our study, we wereable to definitively show that these receptors wereexpressed in olfactory neurons. Further, while Panget al. (1994) did not classify which subtypes werepresent in the epithelium, our study was able toidentify by sequence the presence of both mGluR1and mGluR3 in the olfactory receptor neurons. Thisis in contrast a previous study in the rat taste systemin which the mGluR4 subtype was identified(Chaudhari et al., 1996).
Pang et al. (1994) linked the presence of gluta-mate receptor to IP3 formation using an enzymeassay. Our study reinforced this result by the identi-fication of mGluR1 which is known to be positively
Figure 6 Graphical representation of the calculated coupled to PLC (Conn and Pin, 1997). Further,peak areas for glutamate and methionine responses in the Northern blot analysis performed by Pang et al.presence of mGluR antagonists. The area for each peak
(1994) found that probes to mGluR1 and mGluR4reduced by antagonist was converted into a percent reduc-hybridized to total RNA from the olfactory rosette.tion of the control peak (n Å 7). Values were determinedOur DNA probes were not able to distinguish sub-for both glutamate and methionine in the presence of eachtypes due to cross-hybridization (data not shown).antagonist, and median values were calculated. Error barsCross-reactivity of the Northern probes was not dis-represent the interquartile range for each data set. Lane
1-median reduction caused by MCCG on L-glutamate re- cussed by Pang et al. (1994), and it is possible thatsponse; lane 2-median reduction caused by MCCG on L- they were not specific to receptor subtype. Analyz-methionine response; lane 3-median reduction caused by ing PCR products by sequence allowed our studyS-4CPG on L-glutamate response; lane 4-median reduc- to determine that mGluR1 and mGluR3 subtypestion caused by S-4CPG on L-methionine response. were present in the catfish olfactory epithelium. No
other known subtypes were characterized. Finally,our study found that the mGluR subtypes werecoexpressed in the same individual olfactory recep-These results were obtained by sequence analysis
of PCR products from two individual olfactory re- tor neurons, and furthermore, that these receptorswere also coexpressed with the putative odorant re-ceptor neurons. Analysis of 10 additional neurons
by Southern blot analysis indicated that both recep- ceptors.After characterizing the expression of mGluR1tor types were expressed in all cells tested. To our
knowledge, this is the first reported study that shows and mGluR3 in olfactory receptor neurons, the nextissue addressed in this study was to determine theat the cellular level, mGluRs and odorant receptors
are present in the same olfactory receptor neurons. neuronal localization of the mGluR proteins. Immu-nocytochemical analysis of sectioned olfactory epi-Two additional PCR products similar to mGluR1
were also isolated that did not have the same se- thelium suggested that these receptors are expressedin the apical dendrites and possibly cilia of olfactoryquence or same genomic Southern blot pattern as
the identified subtype (data not shown). In addition, receptor neurons. Examination of the stained sec-tions revealed consistent staining of almost all thethe sequence of these products was unlike any other
known mGluR sequence. Initially, it was predicted dendrites and some of the cilia found within thechemosensory region of the epithelium.that due to the high degree of identity between
mGluR1 and mGluR5, one of the novel products Antibody specificity for each subtype was testedby running negative controls. Nonspecific stainingwas actually the mGluR5 subtype. Immunocyto-
chemical data did not support this hypothesis. It was reduced by pretreatment of slides with hydro-gen peroxide to reduce peroxidase activity. Pread-is possible that these products are novel mGluR
subtypes. There is evidence in other systems that sorption of avidin to the slides reduced nonspecific
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

mGluRs in Olfactory Neurons 103
binding to biotin. Negative controls indicated that present in the olfactory epithelium. This study didstaining was not due to nonspecific effects. Further, not address the presence of these receptors in theslides stained with mGluR5 antibody showed no olfactory tissue; however, a study in rat (Thukralspecific immunoreactivity. These results further et al., 1997) found ionotropic glutamate receptorconfirm antibody specificity, because mGluR5 is ap- subunits in the olfactory epithelium and hasproximately 45% identical to Group II receptors shown that the proteins were primarily localizedand 65% identical to mGluR1 in the rat (Pin and in the dendritic knobs of the olfactory receptorDuvoisin, 1995). neurons. Finally, there may be an odorant receptor
Pharmacological studies were performed to de- belonging to another G-protein coupled family oftermine if these receptors affect odorant signaling. receptors that responds to glutamate. Any of theseS-4CPG is a specific antagonist to mGluR1 (Conn receptors may be contributing to the glutamateand Pin, 1997; Pin and Duvoisin, 1995; Batchelor response and prevent complete reduction of theet al., 1997) , while MCCG has been shown to be olfactory response by mGluR antagonists.a selective antagonist for Group II subtypes This study presents the first evidence linking me-(Conn and Pin, 1997; Jane et al., 1994) . Re- tabotropic glutamate receptors to olfaction in thesponses of known odorants were first recorded in channel catfish. PCR analysis of individual olfac-the absence of antagonists to be used as standards tory receptor neurons found that mGluR1 andand were then recorded after antagonists had been mGluR3 subtypes are expressed in the neurons,applied to the epithelium through the bathing me- while immunocytochemistry showed the proteinsdium. Each antagonist significantly reduced the were localized in the dendritic knobs and some cilia,signal caused by odorant application. The gluta- the site of olfactory transduction. Electrophysiologi-mate response was reduced to 54% of standard
cal experiments found that antagonists to theseby S-4CPG and reduced to 56% of standard by
mGluR subtypes inhibited the response to the odor-MCCG, while methionine was reduced to 73% of
ant glutamate significantly greater than their effectsstandard by S-4CPG and to 76% of standard byon methionine. The combined data are consistentMCCG. The intensity of the antagonist effect waswith the conclusion that mGluRs are localized insignificantly greater for the glutamate response asthe cilia and may, at least partially, act as glutamatecompared to the methionine response. The methi-odorant receptors. This is in agreement with anotheronine response may have been affected by thestudy showing that mGluRs can act as sensory re-glutamate antagonists, because these antagonistsceptors. Chaudhari et al. (1996) found that in theare glycine derivatives. Glycine has been shownrat taste system, mGluR4 seems to act in part as theto cross-react to methionine receptors by abouttaste receptor for monosodium glutamate (MSG).30% (Caprio and Byrd, 1984) , and it is possibleHowever, in the channel catfish, the widespread dis-that these antagonists may be able to competetribution of the mGluRs throughout the epitheliumwith methionine for their binding sites. Anotherdoes not agree with a previous study indicating thatpossible reason that methionine response can beindividual putative odorant receptors are not highlyreduced by these glutamate antagonists is thatexpressed throughout the epithelium (Ngai et al.,these mGluRs are functioning in a general modu-1993a). While it is possible that mGluRs may belatory role within olfaction. Further studies areacting as glutamate odorant receptors, their highneeded to test this hypothesis.density in the epithelium suggests that these recep-These data further support a function of metab-tors may have additional, perhaps modulatory, func-otropic glutamate receptors in olfaction and showtions in olfactory receptor neurons. However, therefor the first time that antagonists to these subtypes
reduce the odorant response. The remainder of the are no behavioral or electrophysiological data cur-glutamate response may be due to other mGluRs rently available showing that glutamate modulatespresent in the tissue. There were two PCR prod- olfactory responses to other odorants.ucts similar to mGluR1 that did not have the samesequence as a known mGluR subtype. These novelproducts may represent additional mGluR sub- This research was supported by NIH DC01500 (totypes that have not yet been characterized but may RCB) and NSF IBN-9221891 (to JC). The authors thankbe responsible for some of the glutamate odorant A. I. Farbman and J. Buchholz for sectioning tissue usedresponse. The glutamate response may also be for immunocytochemistry, and S. Medler for photo-
graphic assistance.partially due to ionotropic glutamate receptors
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio

104 Medler et al.
CONN, P. and PIN, J. (1997). Pharmacology and functionsREFERENCESof metabotropic glutamate receptors. Annu. Rev. Phar-macol. Toxicol. 37:205–237.
ABE, T., SUGIHARA, H., NAWA, H., SHIGEMOTO, R., MI- DUDLEY, C. and MOSS, R. (1995). ElectrophysiologicalZUNO, N., and NAKANISHI, S. (1992). Molecular char- evidence for glutamate as a vomeronasal receptor cellacterization of a novel metabotropic glutamate receptor neurotransmitter. Brain Res. 675:208–214.mGluR5 coupled to inositol phosphate/Ca2/ signal JANE, D., JONES, P., POOK, P., TSE, H., and WATKINS,transduction. J. Biol. Chem. 267:13361–13368. J. (1994). Actions of two new antagonists showing
BATCHELOR, A., KNOPFEL, T., GASPARINI, F., and selectivity for different sub-types of metabotropic glu-GARTHWAITE, J. (1997). Pharmacological character- tamate receptor in the neonatal rat spinal cord. Br. J.ization of synaptic transmission through mGluRs in rat Pharmacol. 112:809–816.cerebellar slices. Neuropharmacology 36:401–403. MCCOOL, B., PIN, J., BRUST, P., HARPOLD, M., and LOV-
BERKOWICZ, D., TROMBLEY, P., and SHEPHERD, G. INGER, D. (1996). Heterologous expression of rat(1994). Evidence for glutamate as the olfactory recep- group II metabotropic glutamate receptors (mGluR2tor cell neurotransmitter. J. Neurophysiol. 71:1–5. and 3) in HEK 293 cells: functional coupling to a
BREER, H., RAMING, K., and KRIEGER, J. (1994). Signal stably expressed v-conotoxin GVIA-sensitive calciumrecognition and transduction in olfactory neurons. Bio- channel. Mol. Pharmacol. 50:912–922.chim. Biophys. Acta 1224:277–287. NGAI, J., CHESS, A., DOWLING, M., NECLES, N., MA-
BRUCH, R. (1996). Phosphoinositide second messengers CAGNO, E., and AXEL, R. (1993a). Coding of olfactoryin olfaction. Comp. Biochem. Physiol. 113B:451–459. information: topography of odorant receptor expression
BRUCH, R. and KALINOSKI, D. (1987). Interaction of in the catfish olfactory epithelium. Cell 72:667–680.GTP-binding regulatory proteins with chemosensory NGAI, J., DOWLING, M., BUCK, L., AXEL, R., and CHESS,receptors. J. Biol. Chem. 262:2401–2404. A. (1993b). The family of genes encoding odorant
BRUCH, R., KANG, J., MOORE, M., and MEDLER, K. receptors in the channel catfish. Cell 72:657–666.(1997). Protein kinase C and receptor kinase gene ex- OHISHI, H., OGAWA-MEGURO, R., SHIGEMOTO, R., KA-pression in olfactory receptor neurons. J. Neurobiol. NEKO, T., NAKANISHI, S., and MIZUNO, N. (1994). Im-33:387–394. munohistochemical localization of metabotropic gluta-
BRUCH, R. and MEDLER, K. (1996). A regulator of G- mate receptors, mGluR2 and mGluR3, in rat cerebellarprotein signaling in olfactory receptor neurons. Neuro- cortex. Neuron 13:55–66.report 7:2941–2944. PANG, J., LO, Y., CHANDLEE, J., and RHOADS, D. (1994).
BRUCH, R. and RULLI, R. (1988). Ligand binding speci- A subtype of the metabotropic glutamate receptor fam-ficity of a neutral L-amino acid olfactory receptor. ily in the olfactory system of Atlantic salmon. FEBSComp. Biochem. Physiol. 91B:535–540. Lett. 354:301–304.
CAPRIO, J. (1995). In vivo olfactory and taste recordings PIN, J. and DUVOISIN, R. (1995). Review: neurotransmit-in fish. In: Experimental Cell Biology of Taste and ter receptors. I. The metabotropic glutamate receptors:Olfaction. CRC Press, Boca Raton, FL, pp. 251–261. structure and functions. Neuropharmacology 34:1–26.
CAPRIO, J. and BYRD, R. (1984). Electrophysiological RESTREPO, D. and TEETER, J. (1990). Olfactory neuronsevidence for acidic, basic, and neutral amino acid olfac- exhibit heterogeneity in depolarization-induced cal-tory receptor sites in the catfish. J. Gen. Physiol. cium changes. Am. J. Physiol. 258:C1051–C1061.84:403–422. ROMANO, C., YANG, W., and O’MALLEY, K. (1996). Me-
CAPRIO, J., DUDEK, J., and ROBINSON, J. (1989). Electro- tabotropic glutamate receptor 5 is a disulfide-linkedolfactogram and multiunit olfactory receptor responses dimer. J. Biol. Chem. 271:28612–28616.to binary and trinary mixtures of amino acids in the SCHOEPP, D. (1994). Glutamate receptors. In: Handbookchannel catfish, Ictalurus punctatus. J. Gen. Physiol. of Receptors and Channels: G Protein Coupled Recep-93:245–262. tors. CRC Press, Boca Raton, FL, pp. 129–140.
CHAUDHARI, N., YANG, H., LAMP, C., DELAY, E., CART- THUKRAL, V., CHIKARAISHI, D., HUNTER, D., and WANG,FORD, C., THAN, T., and ROPER, S. (1996). The taste J. (1997). Expression of non– N-methyl-D-aspartateof monosodium glutamate: membrane receptors in taste glutamate receptor subunits in the olfactory epithelium.
Neuroscience 79:411–424.buds. J. Neurosci. 16:3817–3826.
8p3a 1938/ 8P3A$$1938 02-20-98 13:09:52 nbioas W: Neurobio





























