Embed Size (px)
Citation preview

Acta Neuropathol (Berl) (1986)70:209-219 Acta Neuropathologica �9 Springer-Verlag 1986
Immunohistoehemical Demonstration of Vimentin in Human Cerebral Tumors*
D. Schiffer, M. T. Giordana, A. Mauro, A. Migheli, I. Germano, and G. Giaccone
II Neurological Clinic, University of Turin, Via Cherasco, 15, 1-10126 Turin, Italy
Summary. The distribution of vimentin (VIM) has been histochemically investigated in 53 cerebral tumors and compared in gliomas to that of glial fibrillary acidic protein (GFAP). In gliomas VIM is less positive than GFAP, but shows the same distribu- tion. It cannot be considered as indicating immaturity of glial tumor cells. VIM is also positive in glial processes of cerebellar pilocytic astrocytomas, in Schwann cells of neurinomas and in endothelial cells of all oncotypes. In medulloblastomas, VIM decorates reactive glia cells. A diffuse positive reaction has been observed in meningiomas. In hemangioblastomas, ~ besides intervascular and endothelial cells, groups of polygonal cells are intensely positive for both VIM and GFAP. The interpretati6n of VIM in cerebral tumors is largely based on the distribution patterns of this intermediate filament in the developing CNS of rodents.
Key words: Vimentin - Cerebral tumors - Immuno- histochemistry
Introduction
Vimentin (VIM) is an intermediate filament (IF) of 57,000 daltons in molecular weight, present in fibroblasts and other cells of mesenchymal origin (Franke et al. 1979), and also in glia cells (Chiu et al. 1981; Osborn et al. 1981). VIM and glial fibrillary acidic protein (GFAP) coexist in mature astrocytes (Dahl et al. 1981 a) but during development VIM is expressed much earlier than GFAP (Schnitzer et al. 1981; Bignami et al. 1982; Fedoroffe t al. 1983).
* Supported by a grant of the Italian National Research Council, Special Project "Oncology", contract number 84.00796.44. and by the Italian Association for Cancer Research (A.I.R.C.)
Offprint requests to: Prof. D. Schiffer (address see above)
Pixley and De Vellis (1984) demonstrated in the rat that at the end of intrauterine life radial glia is VIM-positive, and that in the first 2 weeks of extra- uterine life the reaction for VIM diminishes, while that for GFAP increases. This transition, electro- phoretically demonstrated by Dahl (1981) in the rat, takes place between the 2nd and 3rd week of extra- uterine life, and is considered to be an important marker of differentiation (Pixley and De Vellis 1984).
The distribution of VIM in the nervous system of adult rats (Dahl ct al. 1981 b; Shaw et al. 1981 ; Pixley et al. 1984) and adult mouse (Schnitzer et al. 1981) has been described.
To our knowledge, the only observation of VIM in the normal human nervous system was made by Bj6rklund etal . (1984); on smear preparations of CNS, the number of astrocytes stained for VIM was lower than for GFAP.
Very few studies have been done on cerebral tumors. Glioma cell lines are VIM-positive in cultures (Franke etal . 1978; Paetau et al. 1979). On frozen sections, Roessmann et al. (1983) found that neo- plastic astrocytes express both types of IF, but VIM was the predominant type. On formalin fixed tissue, Yung et al. (1985) observed the presence of VIM in neoplastic astrocytes, vessel and mesenchymal cells. Observations in meningiomas are also available (Kar- tenbeck et al. 1981 ; Yung et al. 1985).
This paper deals with the systematic investigation of the immunohistochemical distribution of VIM, compared to that of GFAP, in cerebral tumors.
Materials and Methods
We studied specimens from two cases of normal and reactive nervous tissue as well as from 53 cerebral tumors (six astrocytomas, five anaplastic asrocytomas, seven glioblastomas, four pilocytic astrocytomas of the cerebellum, three glio- sarcomas, four ependymomas, six oligodendrogliomas, five

210 D. Schiffer et al. : Vimentin in Brain Tumors
Diaminobenzidine (DAB) (Bionetics) was used as the sub- strate for the peroxidase. Double staining for GFAP and VIM was performed in selected cases as follows: (a) demonstration of VIM with ABC method using DAB as the substrate for peroxidase, followed by the demonstration of GFAP by PAP
�9
method and l-naphthol-baslc-dye (I.NBD) system for identification of peroxidase (Mauro et al. 1985). The two anti- bodies used were of different animal source; (b) demonstration of VIM by PAP and 1 .NBD system; photography, decoloration and visualization of GFAP by ABC method with DAB (Mauro et al., in preparation).
Results
In the immunoblot t ing experiments, the ant i -VIM antibody demonstrated one single band of immuno- reactivity against cytoskeletal preparat ions (Fig. 1).
Fig. 1. Sodium dodecylsulphate polyacrylamide gel electro- phoresis of a adult rat brain; b glioblastoma; c mol. wt. standards, d Immunostaining of b, transferred to nitrocellulose paper, with monoclonal antibody for VIM: one single band is evident (approximately 57 kDa mol. wt.)
Normal and Reactive Nervous Tissue
VIM was positive in endothelial cells (Fig. 2b), in other cells of cerebral and meningeal vessels as well as in ependymocytes (Fig. 2a). A very weak reaction could be observed in processes of Bergmann's glia and of some astrocytes of the white matter. In the same structures, G F A P showed the already very well- known pat tern distribution. In hypertrophic reactive astrocytes around tumors VIM occurred in the cytoplasm and large processes (Fig. 2c). This finding, however, was limited to the cells at the tumor edges, whereas G F A P was also positive in those at a distance f rom them.
medulloblastomas, seven meningiomas, three hemangio- blastomas, three neurinomas).
The neurosurgical specimens were fixed in Carnoy at 0 -4~ and embedded in paraffin. The histological sections were 5 Ixm thick.
GFAP was demonstrated by the peroxidase-antiperoxidase (PAP) method utilizing rabbit anti-GFAP serum Dako) diluted 1:600 in phosphate-buffered saline (PBS). The normal swine serum, swine anti-rabbit immunoglobulins and the PAP complex were those of Dako Co.
VIM was demonstrated by the avidin-biotin complex (ABC) method utilizing the ABC Vectastain kit (mouse) from Vector. Mouse monoclonal antibody to VIM (Labsystems) was diluted 1 : 100 in PBS. Sections were pre-digested with pepsin (Sigma) 2% in 0.01 N HC1 for 30-40 s at room temperature.
Production and specificity of the monoclonal murine anti- body PK-Vt against porcine VIM have been described (Lehtonen et al. 1983).
Triton insoluble cytoskeletal preparation was obtained by the procedure reported by Dahl et al. (1981 b) from two adult rat brains and two human glioblastomas. Electrophoresis on 8 % sodium dodecylsulphate x polyacrylamide gels was performed according to Laemmli (1970). The electrophoretic transfer from the gels to nitrocellulose paper was performed according to Towbin et al. (1979). Anti-vimentin monoclonal antibodies (Lab- systems) were diluted 1:750. 125I-labeled S. aureus Protein A was utilized for the identification of immunoreactive products�9
Astrocytic Gliomas
In the fibrillary variety of astrocytoma, VIM showed a diffuse positive reaction in the processes, as a fine positive network, whereas in the protoplasmic and gemistocytic varieties it gave an intense reaction in the cytoplasms and processes (Fig. 3). In many cells the reaction was less intense in the central par t of the cytoplasm. The staining was more evident in cell bodies and processes accumulating around the vessels. In anaplastic as t rocytomas the cells of the essentially as t rocytomatous areas were intensely stained, whereas the reaction was variably positive to negative in anaplastic areas.
Endothelial cells, especially those of vascular glomeruli, and other elements of the vessel walls were strongly positive. In pilocytic as t rocytomas of the cerebellum, the bundles of processes and the fibrous perivascular bushes were positive (Fig. 4 c). Rosenthal fibres were negative except in a peripheral rim. Also in these tumors the vascular glomeruli showed strongly positive endothelial cells (Fig. 4d).

D. Schiffer et al.: Vimentin in Brain Tumors 211
Fig. 2. a Normal ependyma showing positive reaction to vimentin in cytoplasm; x 200. b Endothelial cells positive for vimentin; x 400. e Reactive astrocytes positive for vimentin; x400
O Iig odendr o gliomas
Tumor cells were negative. A positive staining could be observed only in reactive astrocytes or in tumor astrocytes (Fig. 4a, b). The cells of the vessels were positive.
Glioblastoma
There was a positive reaction in cytoplasm and pro- cesses of cells of different kinds: small or elongat-
ed cells, gemistocytes or astroblastic elements (Fig. 5a, b), etc. However, the reaction was variable from region to region and also in the same region, being even completely negative. In the vascular component ofglioblastomas and in gliosarcomas VIM was clearly positive in the proliferating endothelial cells of vascular buds and glomeruli (Fig. 5c) and also in cells of other nature in the vessel walls. In mesodermal areas of gliosarcomas the reaction was stronger in periluminal cells than in those of fibroblastic aspect far from the vessel lumina.

212 D. Schiller et al. : Vimentin in Brain Tumors
Fig. 3. a Astrocytoma showing scattered and perivascular astrocytes positive for GFAP; x 400. b A lower number ol astrocytes positive for vimentin; x 400. e Gemistocytic astrocytoma; GFAP, x 400. d Gemistocytic astrocytoma; vimentin, x 400

Fig. 4. a Oligo-astrocytoma showing GFAP-positive astrocytes; x 400. b With vimentin astrocytes are less positive; x 400. e Pilocytic astrocytoma of the cerebellum with vimentin-positive bundles of processes, both in longitudinal and transverse section; x 400. d Vimentin-positive endothelial cells of glomeruli in a pilocytic astrocytoma; x400
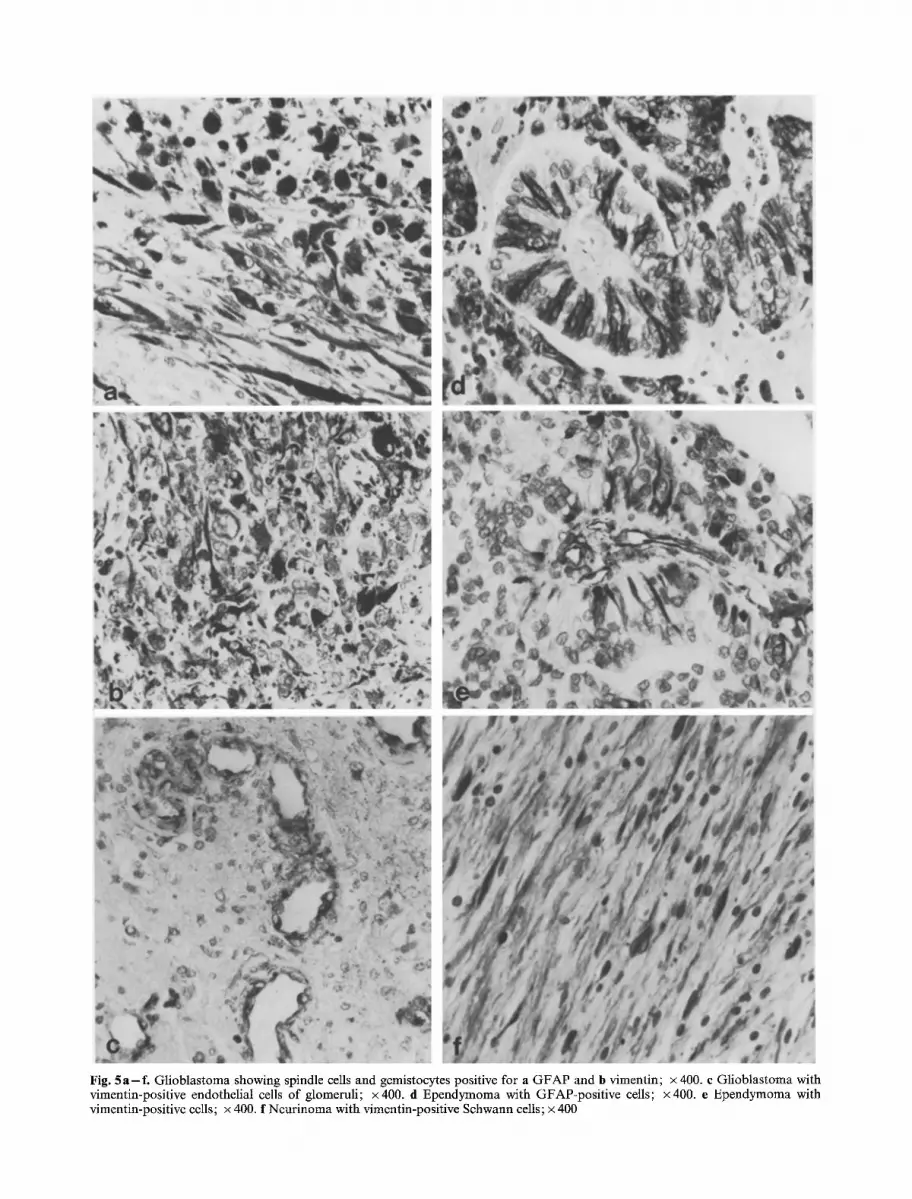
Fig. 5 a - f . Glioblastoma showing spindle cells and gemistocytes positive for a GFAP and b vimentin; • 400. c Glioblastoma with vimentin-positive endothelial cells of glomeruli; x400. d Ependymoma with GFAP-positive cells; • e Ependymoma with vimentin-positive cells; x 400. f Neurinoma with vimentin-positive Schwann cells; x 400

D. Schiffer et al.: Vimentin in Brain Tumors 215
Ependymomas
The cytoplasm of many radiated crowns showed a positive reaction (Fig. 5 d, e). On the contrary, in cana- liculi and rosettes the part of the cytoplasm abutting on the center was negative. Endothelial cells were posi- tive.
In conclusion, in all the tumors considered until now, VIM showed the same distribution as GFAP, with a lower reaction intensity and a smaller number of positive cells.
Neurinomas
Both in longitudinal and transverse section, the posi- tive reaction of the elongated Schwann cells was evident (Fig. 5 f). The astrocyte-like cells of B areas were negative. Vessel cells were positive.
Medulloblastomas
VIM as well as GFAP were positive in reactive astrocytes at the periphery of the tumor (Fig. 6 a) and also in some cells and processes, unevenly distributed in the tumor (Fig. 6 b). In loose mesodermic areas, rich in reticulin, the cells were VIM-positive and GFAP- negative. Endothelial (Fig. 6c) and vessel cells (Fig. 6 d) were VIM-positive.
Meningiomas
The reaction for VIM was intense in the cells of syncitia and whorls (Fig. 7a). The reaction dis- appeared in the latter when hyaline degeneration and calcification developed (Fig. 7b). The vessel cells were only slightly positive.
Hemangioblastomas
A diffuse positive staining for VIM was found in endothelial cells and in the majority of intervascular cells (Fig. 7c). When the latter were large and filled with lipid vacuoles, they were negative. In some peri- pheral or central areas, cells polygonal in shape were clearly positive for both VIM and GFAP (Fig. 7d, e). Reactive astrocytes positive for both markers were found in peritumoral tissue.
Discussion
In normal human nervous tissue we found the same distribution of GFAP as recorded in the literature. The distribution of VIM is more limited than that reported in the mouse (Schnitzer et al. 1981) and in the rat (Dahl et al. 1981 b; Yen and Fields 1981 ; Shaw et al. 1981; Pixley et al. 1984). Very few astrocytic
processes could be observed, as already remarked by Bj6rklund et al. (1984). On the contrary, reactive astrocytes are strongly positive for VIM when located in the tissue immediately adjacent to the tumor, as already observed in stab wounds in the rat (Pixley and De Vellis 1984), and in other experimental models (Schiffer et al. 1984a; Giordana et al. 1984).
In astrocytic gliomas, the VIM and GFAP dis- tribution patterns are similar but the reaction of VIM is less intense, so that cells, which are weakly positive for GFAP, are negative for VIM. This is clearly demonstrated by the double staining, which shows that some elements are positive for GFAP and VIM and others for GFAP only. We have not observed glial elements positive for only VIM. These data are inconsistent with those of Roessmann et al. (1983) who reported that VIM-positive cells predominated over GFAP-positive cells. This discrepancy may be due to the difference in technique, i.e., predigestion with pepsin used by us and, more important, the dif- ferent fixatives and monoclonal antibodies employed. However, the use of pepsin can only favor the demon- stration of VIM; in fact, the clearly positive reaction observed in the vascular elements in the same slides containing VIM-negative glia cells demonstrated that the weaker response of astrocytes is not due to the technique.
The coexistence of two intermediate filaments in glioma cells has prompted conclusions as to their maturation stage. Based upon observations that VIM precedes GFAP in the maturation of glia in the rat and in the mouse (Dahl et al. 1981 b; Schnitzer et al. 1981), one might conclude that the presence of VIM in glioma cells indicates their immaturity (Yung et al. 1985). However, the significance of the two inter- mediate filaments with regard to maturation and differentiation has not yet been completely clarified. The appearance of GFAP does not coincide with the disappearance of VIM in the same cells (Schnitzer et al. 1981 ; Pixley and De Vellis 1984). Moreover, the observations of Houle and Fedoroff (1983) demon- strate that VIM appears only at a certain stage of maturity of neural tube, being thus a marker of matu- rity and differentiation.
Our results confirm the presence of VIM in both neoplastic and reactive astrocytes. In astrocytic tumors, its distribution undergoes the same variations as does GFAP, that is the variability of distribution increases with anaplasia. Therefore, our observations do not allow the conclusion that VIM indicates imma- turity of tumoral astrocytes. At most they might indi- cate the opposite. It may be then that also the ex- pression of VIM, as that of GFAP, has a genotypic basis, acknowledging policlonality as the basis of anaplasia (Bigner et al. 1981 ; Rubinstein et al. 1984).

216 D. Schiffer et al.: Vimentin in Brain Tumors
Fig. 6. Medulloblastoma. a Vimentin-positive peritumoral reactive astrocytes; x400. b Vlmentln-posmve glm celts scatterea m me tumor; x 400. c Vimentin-positive endothelial cells and d vessel cells; x 400
Also, in the ependymoma, the distribution of VIM follows that of GFAP. The positive reaction for VIM and the negative reaction for G F A P of adult ependymal cells do not contribute to the clarifica-
tion of the positive reaction to both markers in ependymomas. Maybe the positive staining of du- plicating neuroepithelial cells has some significance in this regard.

Fig. 7. a Meningioma. Most cells of syncitia and whorls are vimentin-positive; x 400. b Meningioma with vimentin-positive cells of a loose area and negative cells in hyaline vessels; x 400. e Hemangioblastoma showing vimentin-positive endothelial and vascular ceils; x 400, tl and e Hemangioblastoma with intratumoral polygonal GFAP-positive (d) and vimentin-positive (e) cells; • 400

218 D. Schiffer et al.: Vimentin in Brain Tumors
In medulloblastoma, VIM- and GFAP-positive cells represent reactive astrocytes, trapped in the tumor (Mannoji etal. 1981; Schiffer etal. 1983; Giordana et al. 1983; Coffin et al. 1983; Schindler and Gullotta 1983, Marsden et al. 1983). The non-vascular elements positive for VIM and negative for GFAP are mesodermal cells belonging to scar areas. The positive response of neurinoma cells to VIM is in line with the findings of Schachner (Schnitzer et al. 1981) and Shaw et al. (1981) on Schwann cells.
VIM filaments are characteristics of endothelial cells (Franke et al. 1979), fibroblasts, macrophages, chondrocytes, lymphocytes and other mesenchymal elements (Bennet etal. 1978; Franke etal. 1978; Osborn and Weber 1983), as well as smooth muscle cells in the wall of blood vessels (Franke and Warren 1981; Gabbiani et al. 1981; Schmid et al. 1982). The diffuse positive pattern of VIM observed in tumors of mesodermal origin can thus be explained. In meningiomas, positive reaction for VIM was already found in vitro by Lolait et al. (1984) and in vitro, as well in tissue sections, by Kartenbeck et al. (1981).
Of particular interest are the findings in the mesodermal component of gliosarcomas and in the vascular stroma of gliomas, where VIM is intensely positive in the cells of the vessel wall and especially in endothelial cells. This is particularly evident in the endothelial proliferations of the glomeruli of glio- blastomas, cerebellar astrocytomas and in the fibro- sarcomatous proliferations of gliosarcoma, which derive from endothelial proliferation of glioblastomas (Feigin et al. 1958; McComb et al. 1982a; Schiffer et al. 1984b). In hemangioblastoma, both endothelial as well as intervascular elements were VIM-positive provided they did not undergo degenerative processes. The problem of the nature of GFAP-positive stromal cells (De Armond et al. 1980; Deck and Rubinstein 1981; McComb et al. 1982b; Schiffer et al. 1983) is not clarified by the positive reaction for VIM. VIM staining, in fact, could indicate either the mesodermal nature of the cells or their origin from reactive astrocytes.
References
Bennet GS, Fellini SA, Croop JM, Otto J J, Bryan J, Holtzer H (1978) Differences among 100 A filament subunits from different celI types. Proc Natl Acad Sci USA 75: 4364-- 4368
Bignami A, Raju T, Dahl D (1982) Localization of vimentin, the nonspecific intermediate filament protein in embryonal glia and in early differentiating neurons. Dev Bio191:286- 295
Bigner DD, Bigner SH, Pont6n JN, Westermark B, Mahaley MS, Ruoslathi E, Herschman H, Eng CF, Wilkstrand CJ (1981) Heterogeneity of genotypic and phenotypic characteristics of permanent cell lines derived from human gliomas. J Neuropathol Exp Neurol 40:201 --229
BjSrklund H, Eriksdotter-Nilsson M, Dahl D, Olson L (1984) Astrocytes in smears of CNS tissues as visualized by GFA and vimentin immunofluorescence. Med Biol 62 : 34 - 48
Chiu FC, Norton WT, Fields KL (1981) The cytoskeleton of primary astrocytes in culture contains actin, glial fibrillary acidic protein and the fibroblast-type filament protein, vimentin. J Neurochem 37:147-155
Coffin CM, Mukai K, Dehner LP (1983) Glial differentiation in medulloblastomas. Histogenetic insight, glial reaction or invasion of brain? Am J Surg Pathol 7 : 555 - 565
Dahl D (1981) The vimentin-GFAP protein transition in rat neuroglia eytoskeleton occurs at the time of myelination. J Neurosci Res 6: 741 - 748
Dahl D, Bignami A, Weber K, Osborn M (1981a) Filament proteins in rat optic nerves undergoing Wallerian degener- ation; localization of vimentin, the fibroblastic 100 A fila- ment protein, in normal and reactive astrocytes. Exp Neurol 13:496--506
Dahl D, Rueger DC, Bignami A (1981 b) Vimentin, the 57,000 molecular weight protein offibroblast filaments, is the major cytoskeletal component in immature glia. Eur J Cell Biol 24:191 - 196
De Armond S J, Eug LF, Rubinstein LJ (1980) The application of glial fibrillary acidic (GFA) protein immunohisto- chemistry in neurooncology: a present report. Pathol Res Pract 168:374- 394
Deck JHN, Rubinstein LJ (1981) Glial fibrillary acidic protein in stromal ceils of some capillary hemangiobtastomas: sig- nificance and possible implications of an immunoperoxidase study. Acta Neuropathol (Berl) 54:173-181
Fedoroff A, White R, Neal J, Subrahmanian L, Kalnins VI (1983) Astrocyte cell lineage. II. Mouse fibrous astrocytes and reactive astrocytes in cultures have vimentin- and GFAP-containing intermediate filaments. Dev Brain Res 7:303-315
Feigin I, Allen LB, Lipkin L, Gross SW (1958) The endothelial hyperplasia of cerebral blood vessels with brain tumors, and its sarcomatous transformation. Cancer 11 : 264- 277
Franke ED, Warren L (1981) Aortic smooth muscle cells contain vimentin instead of desmin. Proc Natl Acad Sci USA 78 : 3020- 3024
Franke WW, Schmid E, Osborn M, Weber K (1978) Different intermediate-sized filaments distinguished by immuno- fluorescence microscopy. Proc Natl Acad Sci USA 75: 5034- 5038
Franke WW, Schmid E, Osborn M, Weber K (1979) In- termediate-sized filaments of human endothelial cells. J Cell Bioi 81 : 570-- 580
Gabbiani G, Schmid E, Winter S, Chaponnier C, De Chastonay C, Vandererckhove J, Weber K, Franke WW (1981) Vascular smooth muscle cells differ from other smooth muscle cells: predominance of vimentin filaments and a specific alfa-type actin. Proc Natl Acad Sci USA 78: 298 - 302
Giordana NIT, Mauro A, Migheli A, Schiffer D (1983) Contribu- tion of immunohistochemistry to the problem of differentia- tion in medulloblastomas. Ital J Neurol Sci 4: 411 - 415
Giordana MT, Mauro A, Germano I, Giaecone G, Migheli A, Schiffer D (1984) Transplacental ENU tumors of the rat: immunohistochemical contribution to the recognition of cell types. J Neurooncol 2:270
Houle J, Fedoroff S (1983) Temporal relationship between the appearance of vimentin and neural tube development. Dev Brain Res 9:189-195
Kartenbeck J, Schwechheimer K, Moll R, Franke WW (1981) Attachment of vimentin filaments to desmosomal plaques in human meningiomal cells and arachnoidal tissue. J Cell Biol 88:115-126

D. Schiffer et al.: Vimentin in Brain Tumors 219
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680- 685
Lehtonen E, Lehto LP, Paasivuo R, Virtanen I (1983) Parietal and visceral endoderm differ in their expression of inter- mediate filaments. EMBO J 7 :1023- :1028
Lolait S J, Underwood JR, Mu FT, Alderuccio F, Dow CA, Pedersen TS, Chalmers PJ, Toh BH (1984) Vimentin inter- mediate filaments in cultures of human meningiomas. Neuropathol Appl Neurobiol 10:321 - 331
Mannoji H, Takeshita I, Fukui M, Ohta M, Kitamuri K (1981) Glial fibrillary acidic protein in medulloblastoma. Acta Neuropathol (Berl) 55: 63-- 69
Marsden HB, Kumar S, Kahn J, Anderton BJ (1983) A study of glial fibrillary acidic protein (GFAP) in childhood brain tumors. Int J Cancer 31:439-445
Mauro A, Germano I, Giaccone G, Giordana MT, Schiffer D (1985) 1-naphthol-basic-dye (1.NBD): an alternative to diaminobenzidine in immunoperoxidase techniques. Histo- chemistry 83 : 97-102
McComb RD, Trevor PJ, Pizzo SV, Bigner DD (1982a) Speci- ficity and sensitivity of immunohistochemical detection of F V I I I / R A g in formalin-fixed paraffin-embedded tissue. J Histochem Cytochem 30: 371 - 377
McComb RD, Jones TR, Pizzo SV, Bigner DD (1982b) Localization of Factor VIII/von Willebrand Factor and glial fibrillary acidic protein in hemangioblastoma: implications for stromal cells histogenesis. Acta Neuropathol (Berl) 56:207-213
Osborn M, Ludwig-Festl M, Weber K, Bignami A, Dahl D, Bayreuther K (1981) Expression of glial and vimentin type intermediate filaments in cultures derived from human glial material. Differentiation 19 : 161 - 167
Osborn J, Weber K (1983) Tumor diagnosis by intermediate filament typing: a novel tool for surgical pathology. Lab Invest 48 : 372-- 385
Paetau A, Virtanen I, Stemman S, Kurki P, Linder E, Vaheri A, Westermark B, Dahl D, Haltia M (1979) Glial fibrillary acidic protein and intermediate filaments in human glioma cells. Acta Neuropathol (Berl) 47 : 71 - 74
Pixley SK, De Vellis J (1984) Transition between immature radial glia and mature astrocytes studied with a monoclonal anti- body to vimentin. Dev Brain Res 15 : 201 - 209
Pixley SK, Kobayashi H, De Vellis J (1984) Monoclonal anti- body to intermediate filament protein in astrocytes. J Neurosci Res 12: 525-- 541
Roessmann U, Velasco ME, Gambetti P, Autilio-Gambetti L (1983) Vimentin intermediate filaments are increased in
human neoplastic astrocytes. J Neuropathol Exp Neurol 42:309
Rubinstein LJ, Herman MM, VandenBerg SR (1984) Differentiation and anaplasia in central neuroepithelial tumors. Progr Exper Tumor Res 27:32-48
Schiffer D, Giordana MT, Mauro A, Migheli A (1983) Glial fibrillary acidic protein (GFAP) in human cerebral tumors. An immunohistochemical study. Tumori 69: 95 - 104
Schiffer D, Bertolotto A, Giordana MT, Mauro A (1984a) In- duction of brain tumors by transplacental ENU: correlation between neurocytogenesis and tumor development. In: Caciagli F, Giacobini E, Paoletti R (eds) Developmental Neuroscience: Physiological, Pharmacological and Clinical Aspects. Elsevier, Amsterdam, pp 267-272
Schiffer D, Giordana MT, Mauro A, Migheli A (1984b) GFAP, F VIII/RAg laminin and fibronectin in gliosarcomas: an immunohistochemical study. Acta Neuropathol (Berl) 63:108-116
Schindler E, Gullotta F (1983) Glial fibrillary acidic protein in medulloblastomas and other embryonic CNS tumors of children. Virchows Arch [A] 398:263-275
Schmid E, Osborn M, Rungger-Br/indle E, Gabbiani G, Weber K, Franke WW (1982) Immunohistochemical demonstra- tion of vimentin in astrocytes and ependymal cells of developing and adult mouse nervous system. J Cell Biol 90:435-447
Schnitzer J, Franke WW, Schachner M (198:1) Immunohisto- chemical demonstration of vimentin in astrocytes and ependymal cells of developing and adult nervous system. J Cell Biol 90:435- 447
Shaw G, Osborn M, Weber K (1981) An immunofluorescence microscopical study of the naurofilament triplet proteins, vimentin and glial fibrillary acidic protein within the adult rat brain. Eur J Cell Biol 26:68--82
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets; procedure and applications. Proc Natl Acad Sci USA 76:4350-4354
Yen SH, Fields KL (1981) Antibodies to neurofilament, glial filament, and fibroblast intermediate filament proteins bind to different cell types of the nervous system. J Cell Biol 88:115-126
Yung WKA, Luna M, Borit A (1985) Vimentin and glial fibrillary acidic protein (GFAP) in human brain tumors. J Neurooncol 3 : 35 - 38
Received July 2, 1985/Accepted January 8, 1986





























