Embed Size (px)
Citation preview

Harmful Algae 1 (2002) 351–360
Immunolocalization of theca antigens onAlexandrium catenellaand their use to isolate cells from fixed phytoplankton samples
José Luis Córdovaa,∗, Adolfo Jamettb, Georgina Susana Lembeyec
a Fundación Ciencia para la Vida, Millennium Institute for Fundamental and Applied Biology, Av. Marathón 1943, Ñuñoa, Santiago, Chileb Bios Chile I.G.S.A., Av. Marathón 1943, Ñuñoa, Santiago, Chile
c Subsecretar´ıa de Pesca, Bellavista 168, Piso 17, Valparaiso, Chile
Received 30 August 2002; received in revised form 22 September 2002; accepted 28 September 2002
Abstract
Murine monoclonal antibodies were generated and selected for their ability to specifically recognize theca antigens ofAlexandrium catenella(Whedon et Kafoid) Balech cells. The specificity of the monoclonal antibodies for theca antigens wasshown by indirect immunofluorescence and by confocal microscopic analysis. Using these antibodies we demonstrate, forthe first time, the presence of different theca antigens on the cell surface. The fluorescent signal analysis suggests that theseantigens differ in their distribution and quantities in the theca.
Also, using the antibodies we developed a rapid method to isolateA. catenellacells from a lugol-fixed phytoplanktonicsample. The method uses a mixture of different monoclonal antibodies to bind the cells, which then are pulled off from thesample by means of a second anti-mouse antibody coupled to 0.8�m magnetic beads.© 2002 Elsevier Science B.V. All rights reserved.
Keywords: Alexandrium catenella; Theca antigens; Cell immune separation
1. Introduction
Some marine phytoplanktonic species can bloomunexpectedly at certain times of the year. The prolif-erative organisms often produce toxins and are com-monly known as “red tide” or Harmful Algal Blooms(HABs). In recent years, HABs have been affectingnegatively the shellfish and aquaculture industriesaround the world (Anderson, 1989). This situationis becoming critical since there is evidence that thefrequency and distribution of HABs has increasedover the last decades (Hallegraeff, 1993). Therefore,accurate and early identification of toxic phytoplank-
∗ Corresponding author. Tel.:+56-2-239-8969;fax: +56-2-237-3259.
E-mail address:[email protected] (J.L. Cordova).
tonic species is crucial to take appropriate preventivemeasures in shellfish harvesting and other aquacul-ture commercial activities to overcome the negativeimpacts of HABs on human health.
Proper identification of toxic dinoflagellates presentin the water is important, in order to correlate its pres-ence with the nature of the toxin complex detected inshellfish as well as to provide an explanation for deadfish. The isolation and identification of toxic micro-algae from fixed phytoplanktonic samples is currentlya time consuming operation since it is performedmanually under the optical microscope. Furthermore,the time required increases when the number of thecells of interest in the sample is low.
In this paper, we report the generation of murinemonoclonal antibodies against a Chilean strain ofA.catenella. These monoclonal antibodies were selected
1568-9883/02/$ – see front matter © 2002 Elsevier Science B.V. All rights reserved.PII: S1568-9883(02)00052-5

352 J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360
for their ability to recognize specific surface thecaantigens as was demonstrated by immunofluorescenceand confocal microscopic analysis. Using these mon-oclonal antibodies, a method was developed to iso-lateA. catenellacells from lugol-fixed phytoplanktonsamples which takes only 1 h.
2. Materials and methods
2.1. Organism
The Chilean ACC-07 clone ofA. catenellawas iso-lated from phytoplankton samples collected on April1994 in Costa channel (45◦32′S; 73◦34′W), XI regionof Chile. The clone, which was established by the mi-cropipeting method, was kindly provided by MiriamSeguel from CERAN, Universidad Austral de Chile,Puerto Montt, Chile. Cells were cultured in glasstubes and maintained at 10◦C with constant light. Theculture media consisted of Guillard f/2 salts (Sigma,St. Louis, MO) diluted in seawater sterilized by filtra-tion with 0.2�m filtration units (Nalgene, Rochester,NY) as described byGuillard (1975). Dinoflagellateswere harvested when the cell chain was made of 4–6very motile cells, which under our growth conditionsoccurs at day 5 after the culture passage. A 100 mlsample of cell culture was concentrated by centrifu-gation at 1000 rpm for 5 min at room temperatureand counted with an hemocytometer. The pellet wasthen resuspended in PBS to give a final concentrationof 106 cells/ml and aliquoted in 0.5 ml portions thatwere kept frozen until use. The lugol-fixed sampleswere derived from the Chilean monitoring programin the XI region of Chile (Lembeye et al., 1994).
2.2. Mice immunization
The immunization procedure was similar to thatreported bySako et al. (1993a). Briefly, A. catenellafrozen cells were thawed and warmed up quickly toroom temperature and immediately mixed at 1:1 ratiowith Complete Freund Adjuvant (Gibco Long Island,NY). This mixture was uniformly homogenizedbefore immunization using two syringes joined by athree-way Luer lock (Nipro Medical Industries Ltd.,Japan). Three 10-week old female BALB/c mice wereinjected subcutaneously in several places of their
skin in the back. Animals were boosted four timesat 2-week intervals using Incomplete Freund Adju-vant. Ten days after the last boost, serum sampleswere drawn from the immunized mice and sera titersdetermined by ELISA. The mouse with the highesttiter againstA. catenellawas used to generate thehybridoma.
2.3. Monoclonal antibody generation
Spleen cells from the mouse with the highest anti-body titer were harvested 4 days after the last boost.Cells were fused with 2.5 × 107 NSO/two mousemyeloma cells using 50% polyethylene glycol 4000(Merck, Rahway, NJ) as described byKöhler andMilstein (1975). Cell hybrids were resuspended with100�l/well of HAT selecting medium on 96-well tis-sue culture plates (Nunc, Roskilde, Denmark). HATmedium consisted in Dulbecco’s medium supple-mented with 0.1 mM nonessential amino acids, 10%fetal bovine serum (HyClone Laboratories, Logan,UT), penicillin (100 IU/ml) streptomycin (100�g/ml),2 mM l-glutamine, 10−4 M hypoxanthine, 4×10−7 Maminopterin, and 1.6 × 10−5 M thymidine (Sigma).Plates were incubated at 37◦C in a moist atmospherewith 10% CO2. Ten days after fusion, hybridomasupernatants were screened for secretion of specificantibodies by ELISA as described below. All posi-tive cultures were expanded onto 24-well plates andhybrid samples were frozen in liquid nitrogen witha medium containing 10% dimethylsulfoxide and10% fetal bovine serum. Positive hybridomas werere-cloned by the limiting dilution technique.
2.4. Immunoglobulin isotyping
Subclass typing of monoclonal antibodies fromtissue culture supernatants was done using the com-mercial kit, ISOTYPE Ab-SATTM (Menlo Park, CA).Isotyping was performed following the procedurerecommended by the manufacturer.
2.5. Selection of monoclonal antibodies by ELISA
The ELISA protocol used to screen the monoclonalantibodies was described byVoller et al. (1978).Briefly, each well of a 96-well ELISA plate was coatedwith a 50�l/well suspension of glutaraldehyde-fixed

J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360 353
1× 106 A. catenellacells/ml and incubated overnightat room temperature until dry. The well surface wasblocked with 1% BSA in PBS solution for 1 h atroom temperature. Undiluted supematants from theplated hybridoma and serial dilutions of the pre- andimmune serum were added into the wells and incu-bated overnight at 4◦C. The plates were washed threetimes with 0.02% Tween 20 in PBS, then incubatedfor 1 h at room temperature with goat anti-mouse IgGantibody conjugated with alkaline phosphatase andincubated for 1 h at room temperature. After washingoff the unbound second antibody, the activity was de-veloped by adding 50�l/well of p-nitro-phenyl phos-phate (1 mg/ml, Sigma) in 0.1 M Tris–HCl buffer, pH9.5 containing 0.1 M NaCl, 5 mM MgCl2 and incu-bating for 30 min at 37◦C. The reaction was stoppedby adding 50�l/well of 3N NaOH and the plateswere read at A405 nm using an ELISA reader, modelMKII (Titertek Multiskan Plus, Labsystem, Finland).An absorbance value of 0.1 or higher was consideredas positive.
2.6. Indirect immunofluorescence assayof the whole cell
The identification of monoclonal antibodies thatrecognize only external surface antigens was per-formed by indirect immunofluorescence as describedby Johnson et al. (1982). Briefly, 100 ml of 5 dayold culture was concentrated by centrifugation as de-scribed above and adjusted to approximately 106A.catenella cells/ml. Cells were fixed overnight with2% glutaraldehyde, at 4◦C under dim light. Underthis condition cells sank to the bottom of the tubewhich allowed removal of the fixing solution. Thencells were fixed again with 2% of glutaraldehydein 100 mM cacodylate buffer, pH 7.0, for 24 h at4◦C under dim light. A 100�l aliquot of fixed cellswas placed in an Eppendorftube and 500�l of 5%BSA-PBS was added to block the non-specific bind-ing sites. An additional 500�l of fresh 5% BSA-PBSwas added into the tube and incubated by rotation for1 h at room temperature. Cells were allowed to settledown, the 5% BSA-PBS solution was removed andcells resuspended with 200�l of diluted ascites fluid(1:1000) in 5% BSA-PBS and incubated with slow ro-tation for 1 h at room temperature. The first antibodywas removed by quick centrifugation using a bench
top microcentrifuge. Then, cells were subjected totwo washes in 1 ml of 5% BSA-PBS and resuspendedin 200�l per tube of goat anti-mouse conjugatedwith fluorescein diluted 1:250 in 5% BSA-PBS. Thetube was incubated with slow rotation for 1 h at roomtemperature, cells washed as described above andresuspended in 100�l of PBS. A 15�l aliquot wasplaced over a glass slide, covered and analyzed usinga Orthoplan fluorescent microscope (Leitz, Wetzlar,
Fig. 1. Procedure to isolate fixedAlexandriumcells from a phyto-planktonic sample. The method developed uses a mixture of mon-oclonal antibodies during the first incubation. Then, after washingunbound antibodies, the cells–antibody complex is incubated withanti-mouse IgG coupled to a magnetic latex particles. Finally, thewhole complex is isolated using a magnet. The whole proceduretakes 60 min.
354 J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360
Germany) or an Axiovert 135M Zeiss Confocal Mi-croscope (Germany). The images were analyzed usingthe LSM 3.9 program (Zeiss).
2.7. Isolation of A. catenella cells from lugol-fixedmonitoring samples
The method developed to isolate fixed cells ispresented inFig. 1. Briefly, 1 ml from a lugol-fixedmonitoring samples was placed in an Eppendorf-tube and subjected to quick centrifugation using abenchtop microcentrifuge (Eppendorf, Germany).The supernatant was removed and the pellet wascarefully resuspended with 1 ml of 5% BSA-PBSand centrifuged again. The washing procedure wasrepeated twice and the cellular pellet mixed with500�l of 1:500 dilution of the anti-theca monoclonalantibody mixture diluted in 5% BSA-PBS and incu-bated for 30 min at room temperature. The cells werecentrifuged, the supernatant removed and washed asdescribed above. The cell pellet was resuspended in1 ml of a 0.1% (w/v) solution of 0.8�m magneticlatex particles (Bangs Laboratories, Fishes, Indiana)coupled with anti-mouse IgG antibody. Then, theA.catenellacells were separated from other cells by the
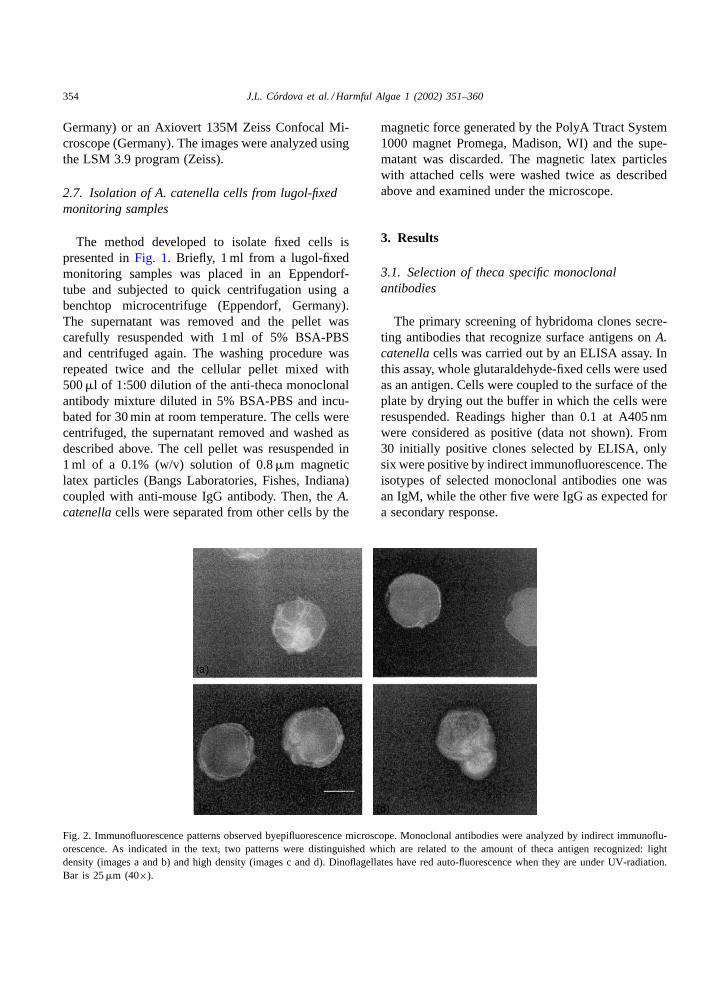
Fig. 2. Immunofluorescence patterns observed byepifluorescence microscope. Monoclonal antibodies were analyzed by indirect immunoflu-orescence. As indicated in the text, two patterns were distinguished which are related to the amount of theca antigen recognized: lightdensity (images a and b) and high density (images c and d). Dinoflagellates have red auto-fluorescence when they are under UV-radiation.Bar is 25�m (40×).
magnetic force generated by the PolyA Ttract System1000 magnet Promega, Madison, WI) and the supe-matant was discarded. The magnetic latex particleswith attached cells were washed twice as describedabove and examined under the microscope.
3. Results
3.1. Selection of theca specific monoclonalantibodies
The primary screening of hybridoma clones secre-ting antibodies that recognize surface antigens onA.catenellacells was carried out by an ELISA assay. Inthis assay, whole glutaraldehyde-fixed cells were usedas an antigen. Cells were coupled to the surface of theplate by drying out the buffer in which the cells wereresuspended. Readings higher than 0.1 at A405 nmwere considered as positive (data not shown). From30 initially positive clones selected by ELISA, onlysix were positive by indirect immunofluorescence. Theisotypes of selected monoclonal antibodies one wasan IgM, while the other five were IgG as expected fora secondary response.
J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360 355
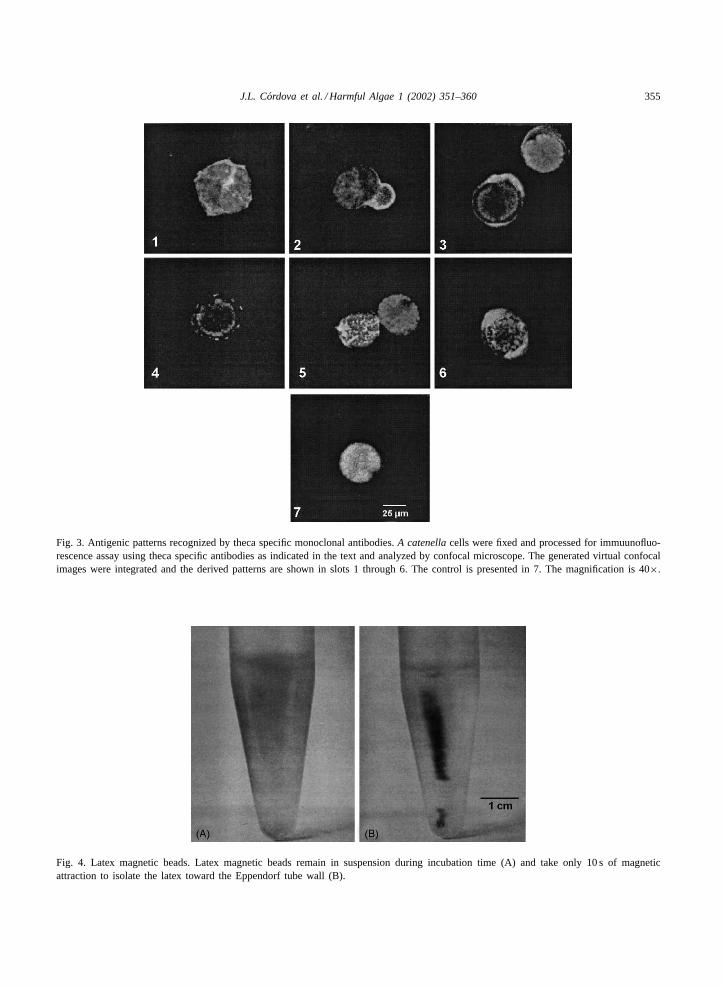
Fig. 3. Antigenic patterns recognized by theca specific monoclonal antibodies.A catenellacells were fixed and processed for immuunofluo-rescence assay using theca specific antibodies as indicated in the text and analyzed by confocal microscope. The generated virtual confocalimages were integrated and the derived patterns are shown in slots 1 through 6. The control is presented in 7. The magnification is 40×.
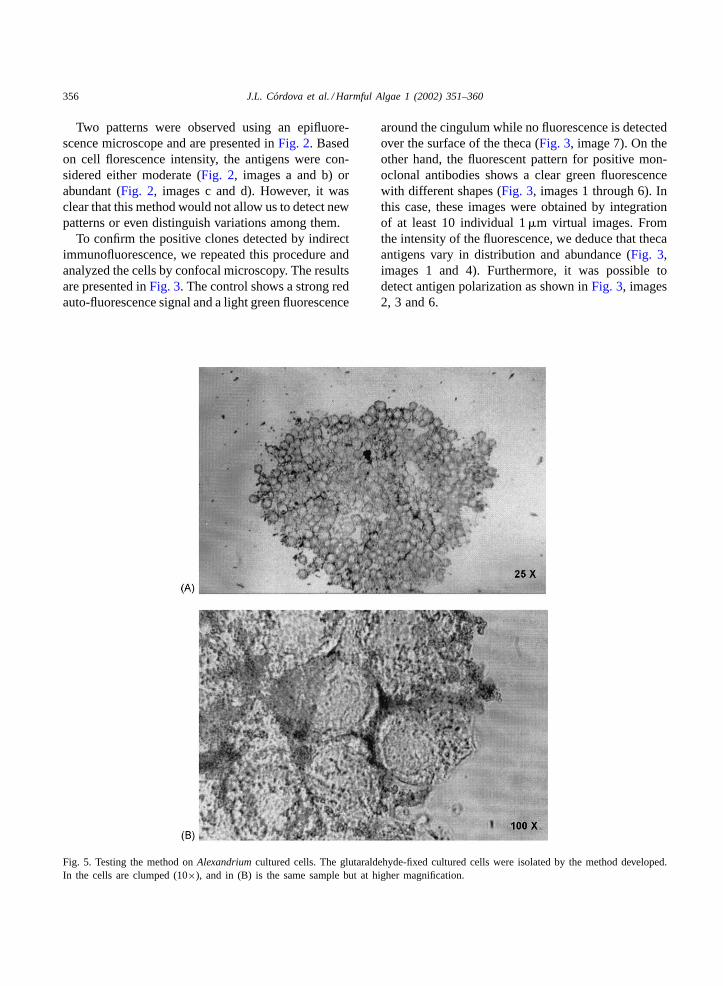
Fig. 4. Latex magnetic beads. Latex magnetic beads remain in suspension during incubation time (A) and take only 10 s of magneticattraction to isolate the latex toward the Eppendorf tube wall (B).
356 J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360
Two patterns were observed using an epifluore-scence microscope and are presented inFig. 2. Basedon cell florescence intensity, the antigens were con-sidered either moderate (Fig. 2, images a and b) orabundant (Fig. 2, images c and d). However, it wasclear that this method would not allow us to detect newpatterns or even distinguish variations among them.
To confirm the positive clones detected by indirectimmunofluorescence, we repeated this procedure andanalyzed the cells by confocal microscopy. The resultsare presented inFig. 3. The control shows a strong redauto-fluorescence signal and a light green fluorescence
Fig. 5. Testing the method onAlexandriumcultured cells. The glutaraldehyde-fixed cultured cells were isolated by the method developed.In the cells are clumped (10×), and in (B) is the same sample but at higher magnification.
around the cingulum while no fluorescence is detectedover the surface of the theca (Fig. 3, image 7). On theother hand, the fluorescent pattern for positive mon-oclonal antibodies shows a clear green fluorescencewith different shapes (Fig. 3, images 1 through 6). Inthis case, these images were obtained by integrationof at least 10 individual 1�m virtual images. Fromthe intensity of the fluorescence, we deduce that thecaantigens vary in distribution and abundance (Fig. 3,images 1 and 4). Furthermore, it was possible todetect antigen polarization as shown inFig. 3, images2, 3 and 6.
J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360 357
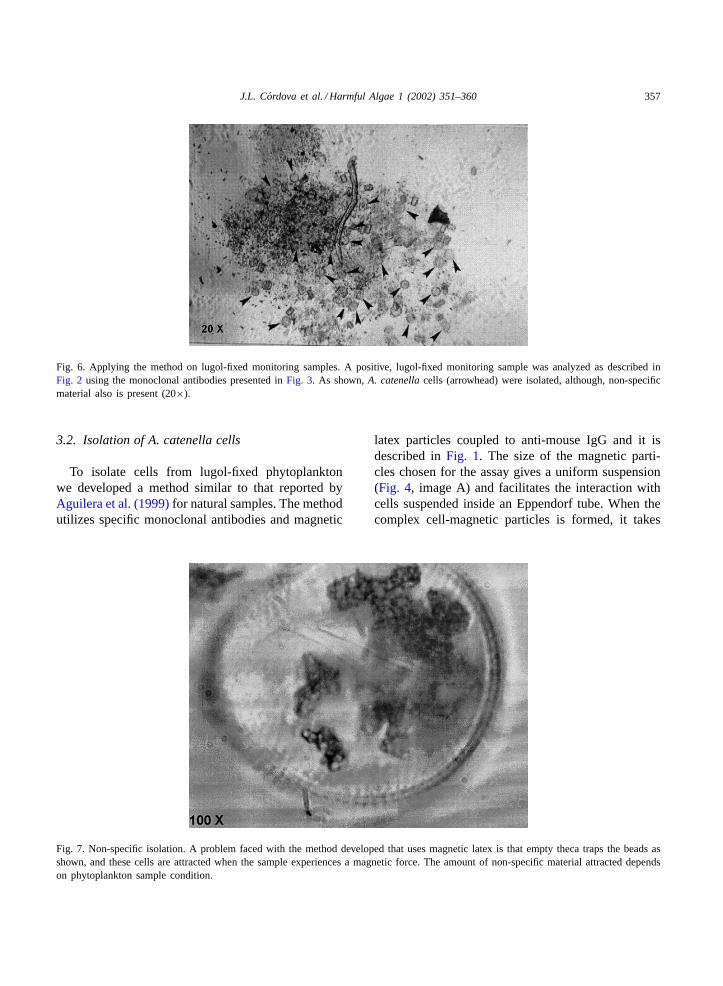
Fig. 6. Applying the method on lugol-fixed monitoring samples. A positive, lugol-fixed monitoring sample was analyzed as described inFig. 2 using the monoclonal antibodies presented inFig. 3. As shown,A. catenellacells (arrowhead) were isolated, although, non-specificmaterial also is present (20×).
3.2. Isolation of A. catenella cells
To isolate cells from lugol-fixed phytoplanktonwe developed a method similar to that reported byAguilera et al. (1999)for natural samples. The methodutilizes specific monoclonal antibodies and magnetic
Fig. 7. Non-specific isolation. A problem faced with the method developed that uses magnetic latex is that empty theca traps the beads asshown, and these cells are attracted when the sample experiences a magnetic force. The amount of non-specific material attracted dependson phytoplankton sample condition.
latex particles coupled to anti-mouse IgG and it isdescribed inFig. 1. The size of the magnetic parti-cles chosen for the assay gives a uniform suspension(Fig. 4, image A) and facilitates the interaction withcells suspended inside an Eppendorf tube. When thecomplex cell-magnetic particles is formed, it takes
358 J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360
only 10 s for magnetic latex particles to sedimentinto the Eppendorf tube walls as observed inFig. 4,image B.
To evaluate the efficiency of this method to isolateA. catenellacells from a complex cell mixture, weused glutaraldehyde-fixed cell from unialgal cultures.As shown inFig. 5, cells were clumped by the mag-netic beads (Fig. 5, image A) and the magnetic par-ticles were associated to the theca (Fig. 5, image B).
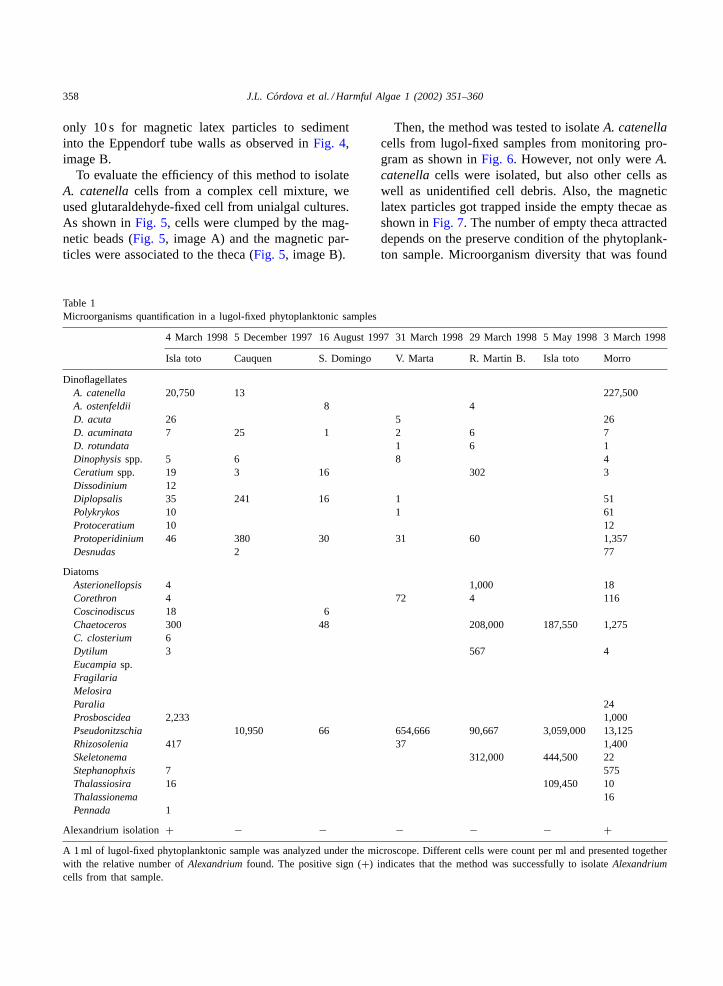
Table 1Microorganisms quantification in a lugol-fixed phytoplanktonic samples
4 March 1998 5 December 1997 16 August 1997 31 March 1998 29 March 1998 5 May 1998 3 March 1998
Isla toto Cauquen S. Domingo V. Marta R. Martin B. Isla toto Morro
DinoflagellatesA. catenella 20,750 13 227,500A. ostenfeldii 8 4D. acuta 26 5 26D. acuminata 7 25 1 2 6 7D. rotundata 1 6 1Dinophysisspp. 5 6 8 4Ceratiumspp. 19 3 16 302 3Dissodinium 12Diplopsalis 35 241 16 1 51Polykrykos 10 1 61Protoceratium 10 12Protoperidinium 46 380 30 31 60 1,357Desnudas 2 77
DiatomsAsterionellopsis 4 1,000 18Corethron 4 72 4 116Coscinodiscus 18 6Chaetoceros 300 48 208,000 187,550 1,275C. closterium 6Dytilum 3 567 4Eucampiasp.FragilariaMelosiraParalia 24Prosboscidea 2,233 1,000Pseudonitzschia 10,950 66 654,666 90,667 3,059,000 13,125Rhizosolenia 417 37 1,400Skeletonema 312,000 444,500 22Stephanophxis 7 575Thalassiosira 16 109,450 10Thalassionema 16Pennada 1
Alexandrium isolation+ − − − − − +A 1 ml of lugol-fixed phytoplanktonic sample was analyzed under the microscope. Different cells were count per ml and presented togetherwith the relative number ofAlexandriumfound. The positive sign (+) indicates that the method was successfully to isolateAlexandriumcells from that sample.
Then, the method was tested to isolateA. catenellacells from lugol-fixed samples from monitoring pro-gram as shown inFig. 6. However, not only wereA.catenellacells were isolated, but also other cells aswell as unidentified cell debris. Also, the magneticlatex particles got trapped inside the empty thecae asshown inFig. 7. The number of empty theca attracteddepends on the preserve condition of the phytoplank-ton sample. Microorganism diversity that was found

J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360 359
in the monitoring samples used to isolateA. catenellacells is presented inTable 1.
4. Discussion
To develop a method for isolatingA. catenellacells from monitoring samples, it was necessary togenerateA. catenellatheca specific monoclonal anti-bodies, since intracellular antigens would be hiddenwhen cells are fixed and preserved. The generationof the monoclonal antibodies capable of recognizingA. catenellatheca was difficult because several micedied during the last intravenous boost before spleencells fusion. We believe that the mice deaths can-not be attributed to cell toxicity because of the lownumber of cells used. This problem was overcomeby injecting the mice using small amounts of cellssubcutaneously at different places under the skin.
The selection of monoclonal antibodies that onlyrecognized theca antigens was done in a two stepprocedure. First, the initial screening was performedby ELISA using as antigen glutaraldehyde-fixed cul-tured cells. Fixed cells were necessary since non-fixedcells seemed to wash off easily from the well. Also,non-fixed cells could expose non-theca antigens andthe assay may not discriminate internal from externalantigen. In the second step, the positive ELBA cloneswere re-screened using indirect immunofluorescenceassay that allowed the visual detection of cell integrityand fluorescence as shown inFig. 2. Thus, theca anti-gens were easily recognized facilitating monoclonalantibody selection. Only two fluorescence patternswere distinguished by indirect immunofluorescence,either discreet or abundant, as shown inFig. 2. How-ever, it was not possible to discriminate if thesepatterns correspond to a uniform antigen distributionover the theca of if there is any variation in antigenpattern distribution throughout the cell cycle, since weonly used cells from one stage of the growth curve.However, it has been shown that there is variability inantigen expression depending on the cell metabolismas reported byAnderson et al. (1997).
This reported variability was the reason why we de-cided to use a mixture of all available monoclonal an-tibodies. Also, we found that monoclonal antibodiesthat recognize theca antigens with low distributions(Fig. 3, image 4) requires more time to bind cells and
was less efficient to pull off cells from the phytoplank-tonic sample.
To improve the resolution of the fluorescencepattern observed inFig. 2, cells prepared for indi-rect immunofluorescence were analyzed by confocalmicroscopy as shown inFig. 3, an unique antigendistribution pattern is observed. Two important as-pects should be pointed out, since this is the first timethat specific theca antigens are described. One is theexistence of different theca antigens, and second, thehigh variability in their quantities and distributionor possible re-arrangement (polarization). This lastobservation could also, indicate that theca antigenscan be freely rearranged by the cell, for example dur-ing cell division as it could be interpreted fromFig. 3,image 2.
With the help of theca specific monoclonal antibod-ies a method to isolate fixedA. catenellacells fromlugol-fixed monitoring sample was developed (Fig. 1).Initially, we used only one monoclonal antibody, butthe results were considered no good enough, becausethe assay takes more time than ideal (1 h assay). Then,we decided to use a mixture, of all of them, sincethe possibility exist that antigen variation or antigendestruction due to fixation or handling would affectrecognition and thus cell isolation. The size of thebeads to use was also investigated. Among the dif-ferent sizes of beads tested we selected the 0.8�msize beads because they keep a uniform distributionin 1 ml samples as shown inFig. 4. Also, sample ro-tation to facilitate the interaction was not require us-ing this size beads. Rotation increase collision amongthe cells and beads affecting their morphology, andlarger or smaller bead sizes tend to sink or float, re-spectively. To evaluate the effectiveness of the methodfor isolated A. catenellacells, glutaraldehyde-fixedcells from unialgal cultures were used and as shownin Fig. 5, they were effectively isolated. In addition,the method was evaluated to isolateA. catenellacellsfrom a lugol-fixed cells and again cells were effec-tively isolated, indicating that these theca antigenswere not substantially modified during lugol fixation(Fig. 6).
The use of small magnetic latex particles (0.8�m)coupled with anti-mouse IgG antibody as a “pullingagent”, allowed us to isolate cells from complexsamples with low amount of contaminating mate-rial as shown inFig. 6. Although, in some instances

360 J.L. Cordova et al. / Harmful Algae 1 (2002) 351–360
the latex appears to enter the empty theca, draggingnon-specifically other cells as shown onFig. 7. Thisnon-specific binding reaction reduces the specificityand sensibility of the method.
Finally, this method could be useful to analyze largenumber of monitoring samples to determine the pres-ence ofA. catenellaas shown inTable 1. However, wehave not determined yet the minimum number ofA.catenellacells that this assay is able to isolate. Basedon our preliminary results, the assay sensitivity willdepend not only of the number ofA. catenellacellspresent in the sample (2×104 cells/ml), but also uponthe presence and relative abundance of other phyto-planktonic organisms. Similar methods could also beused to isolate other microorganisms of interest bygenerating their respective monoclonal antibodies andusing the same second antibody-latex particles.
Acknowledgements
We would like to thank Mr. Alvaro Morales fortaken care of the immunized animals. This work waspartially financed by the P. Valenzuela and B. MendezFoundation and the Millennium Institute for Funda-mental and Applied Biology.
References
Aguilera, A., Gonzalez-Gil, S., Keafer, B.A., Anderson,D.M., 1999. Immunomagnetic separation cells of the toxicdinoflagellate Alexandrium fundvensefrom natural samples.Mar. Ecol. Prog. Ser. 143, 255–269.
Anderson, D.M., 1989. Toxic algal blooms and red tides: a globalperspective. In: Okaichi, T., Anderson, D.M. (Eds.), Red TidesBiology, Environmental Science and Toxicology. Elsevier, NewYork, pp. 11–16, 489.
Anderson, D.M., Kulis, D.M., Keafer, B.A., Berdlet, E., 1997.Detection of the toxic dinoflagellateAlexandrium fundvensewith rRNA and antibody probes: variability in labeling intensity
with physiological condition (Abstract, ASLO). In: Proceedingsof the Aquatic Sciences Meeting, Santa Fe, February 1996.
Guillard, R.R.L., 1975. Culture of phytoplankton for feedingmarine invertebrates. In: Smith, W.L., Chanley, M.H. (Eds.),Culture of Marine Invertebrate Animals, pp. 29–60.
Hallegraeff, G.M., 1993. A review of harmful algal blooms andtheir apparent global increase. Phycology 32, 79–99.
Johnson, G.D., Davison, R.S., McNamee, K.C., Goodwin, D.,Holborow, E.J., 1982. Fading the immunofluorescence duringmicroscopy: a study of the phenomenon and its remedy. J.Immunol. Methods 55, 215–242.
Köhler, B., Milstein, C., 1975. Continuos cultures of fused cellssecreting antibody of predefined specificity. Nature 256, 495–498.
Lembeye, G., Loyola, H., Seguel, M., Ñúñez, M., 1994. Presenciade Alexandriumen la XI Región. Un llamado de alerta. XIVJornadas de Ciencias de1 Mar y I Jornada de Chilena deSalmonicultura, 23–25 de Mayo 1994, p. 83.
Sako, Y., Adachi, M., Ishida, Y., 1993. Preparation and charac-terization of monoclonal antibodies to Alexandrium species. In:Smayda, T.J., Shimizu, Y. (Eds.), Toxic Phytoplankton Bloomsin the Sea. Elsevier, New York, pp. 87–93.
Voller, A., Bartlett, A., Bidwell, D.E., 1978. Enzyme immuno-assays with special reference to ELISA techniques. J. Clin.Pathol. 31, 507–520.
Further reading
Adachi, M., Sako, Y., Ishida, Y., 1993a. The identification ofspecific dinoflagellatesAlexandrium tamarensefrom Japan andThailand by monoclonal antibodies. Nippon Siusan Gakkaishi59, 327–332.
Adachi, M., Sako, Y., Ishida, Y., 1993b. Application of monoclonalantibodies to field samples ofAlexandrium species. NipponSiusan Gakkaishi 59, 1171–1175.
Sako, Y., Adachi, M., Ishida, Y., 1995. Specific monoclonalantibodies and DNA probes for the identification of thetoxic dinoflagellate genus Alexandrium. In: Lassus, P., Arzul,G., Erard, E., Gentien, P., Marcaillou, C. (Eds.), HarmfulMarine Algal Blooms, Technique et documentation—Lavoisier.Intercept Ltd., pp. 77–82.
Seguel, M., 1995. Evaluatión de aspectos biológicos deA.catenellaen cultivo. ler. Seminario Taller sobre Mareas RojasTóxicas de1 Extremo sur y Austral de Chile. Punta Arenas,24 pp.





























