Embed Size (px)
Citation preview

atd
w1sdaalob
Developmental Biology 210, 44–55 (1999)Article ID dbio.1999.9267, available online at http://www.idealibrary.com on
Interferon Regulatory Transcription Factors AreConstitutively Expressed and SpatiallyRegulated in the Mouse Lens
Wenmei Li,* Chandrasekharam N. Nagineni,* Bassey Efiok,†Ana B. Chepelinsky,‡ and Charles E. Egwuagu*,1
*Laboratory of Immunology and ‡Laboratory of Molecular & Developmental Biology,National Eye Institute, and †Laboratory of Molecular Hematology, National Heart,Lung, and Blood Institute, National Institutes of Health, Bethesda, Maryland 20892
Interferon regulatory factors (IRFs) are a family of transcription factors involved in regulation of cell growth andimmunological responses. Nine IRFs have been described and they are expressed in a variety of cells, except for ICSBP andLSIRF/Pip, which are thought to be expressed exclusively in immune cells. Here, we show that IRF-1, IRF-2, ICSBP, andLSIRF/Pip are constitutively expressed in the mouse lens. These IRFs are present in both the cytoplasm and the nuclei oflens cells. However, the nuclear and cytoplasmic proteins exhibit distinct mobilities on SDS/PAGE. We further show thatin the developing mouse lens, IRF-1 and IRF-2 are expressed at high levels in differentiated lens fiber cells with very low andbarely detectable levels in undifferentiated lens epithelial cells. Although the level of ICSBP expression is very low in thenormal mouse lens, in transgenic mice with constitutive expression of interferon g in the lens, its level is markedly elevatednd ICSBP expression is detected exclusively in the nuclei of undifferentiated lens cells. Taken together, our data suggesthat expression of IRF transcription factors is spatially regulated in the lens and that distinct IRFs may contribute toifferential gene regulation in the epithelial and fiber compartments of the vertebrate lens.
Key Words: IFNg; ICSBP; LSIRF/Pip; IRF-1; IRF-2; STAT1; lens differentiation.cshwGrife
IaIIIdit
INTRODUCTION
Interferons (IFNs) are multifunctional cytokines inducedin almost all vertebrate cells in response to infection orimmunological stimuli. There are two main types: Type 1IFN comprises leukocyte (IFNa) and fibroblast (IFNb) IFNs
hile Type 2 or immune IFN is IFNg (Vilcek and Sen,996). Although the two types of IFNs differ in the cellurface receptors they bind to and their physiological in-ucers, their biological activities are mediated throughctivation of the JAK/STAT signaling pathway (Schindlernd Darnell, 1995). Interaction of IFNs with their receptorseads to activation of protein tyrosine kinases, JAK1, JAK2,r Tyk2, which in turn phosphorylate and activate mem-ers of a family of latent cytoplasmic transcription factors
1 To whom correspondence should be addressed at the Labora-tory of Immunology, National Eye Institute, National Institutes of
s(
Health, 10/10N116, 10 Center Drive MCS 1858, Bethesda, MD20892. Fax: (301) 480-3914. E-mail: [email protected].
44
alled STATs (signal transducers and activators of tran-cription) (Darnell, 1997). Phosphorylated STATs formomo- or heterodimers that translocate to the nucleushere they bind to well-defined DNA sequences calledAS (gamma IFN activation site) or ISREs (IFN-stimulated
esponse elements) and activate transcription of genes cod-ng for members of the interferon regulatory factors (IRFs)amily of transcription factors (Decker et al., 1997; Boehmt al., 1997).IRFs interact with ISRE motifs in the promoters of
FN-regulatable genes and they mediate transcriptionalctivation or repression of these genes. Ten members of theRF family have been identified and they include ICSBP,SGF3g/p48, IRF-1, IRF-2, IRF-3, IRF-4/LSIRF/Pip/ICSAT,RF-5, IRF-6, IRF-7, and vIRF (Nguyen et al., 1997). IRFsiffer in the range of cell types they are normally expressedn, their physiological inducers, and the biological processeshey affect (Nguyen et al., 1997). With the recent demon-
tration of a virally encoded homologue of cellular IRFsMoore et al., 1996), it is likely that more IRFs will be0012-1606/99

u
glttsm
c
RrtDia4esaaPGG5CmAmAI5
45Differential Expression of IRFs in the Lens
identified and previously described members would befound to possess new functions.
IRF-1 and IRF-2 are the best characterized members ofthis family and were initially identified as regulators of theIFN system (Miyamoto et al., 1988; Harada et al., 1989).They have subsequently been shown to be key factors in theregulation of cell growth through their effects on the cellcycle (Taniguchi et al., 1995; Vaughan et al., 1997). IRF-1 isthought to function in a manner analogous to the tumorsuppressor p53, activating a set of genes whose products arerequired for negative regulation of cell growth. On the otherhand, IRF-2, which shares significant sequence similarity toIRF-1 within the DNA binding domain, represses IRF1-regulatable genes (Taniguchi, 1997). In contrast to IRF-1 andIRF-2 that are expressed in a variety of cell types, two IRFmembers, ICSBP (interferon consensus sequence bindingprotein) (Driggers et al., 1990) and LSIRF/Pip (lymphoid-specific IRF or Pu.1 interaction partner) (Eisenbeis et al.,1995; Matsuyama et al., 1995; Yamagata et al., 1996) arethought to be expressed exclusively in cells of macrophageand lymphocyte lineages. Constitutive expression of ICSBPis thought to be limited to B lymphocytes (Driggers et al.,1990; Politis et al., 1994; Nelson et al., 1996). However,transcription of the ICSBP gene can be induced in T cellsand macrophages by either IFNg or antigenic stimulation(Politis et al., 1994; Nelson et al., 1996). LSIRF/Pip expres-sion is thought to be restricted to cells of the T and Blymphocyte lineages. Mice with a null mutation for theICSBP or LSIRF/Pip gene develop myelogenous leukemia-like syndrome (Holtschke et al., 1996) or developmentaldefects in lymphocyte maturation (Mittrucker et al., 1997),respectively. Taken together, these observations have pro-vided support for the notion that LSIRF/Pip and ICSBP arelymphoid-specific IRFs.
We have previously reported the generation of transgenicmice with targeted ectopic expression of IFNg in the lens
nder the direction of the aA-crystallin promoter (Egwuaguet al., 1994a,b). In these mice, eye development is inhibitedand the developmental fate of cells destined to become lensfiber cells is altered. In the present study, we show thatabnormal lens differentiation observed in the IFNg trans-enic mice may derive in part from perturbations in theevels of IRF factors in the lens. We also show for the firstime that ICSBP, LSIRF/Pip, IRF-1, and IRF-2 are constitu-ively expressed in the normal mouse lens and that expres-ion of these IRFs is spatially regulated in the developingouse lens.
MATERIALS AND METHODS
Animals
FVB/N and BALB/c wild-type (WT) and the generation of theIFNg transgenic (TR) mice have previously been described
(Egwuagu et al., 1994a). All animal procedures conformed toinstitutional guidelines and the Association for Research in VisionCopyright © 1999 by Academic Press. All right
and Ophthalmology Resolution on the Use of Animals in Re-search.
Cell Culture and IFNg Treatment
The murine lens epithelial cell line, aTN4-1 (Yamada et al.,1990), kindly provided by Dr. Paul Russell (NEI, NIH, Bethesda,MD), was grown in Dulbecco’s modified Eagle’s medium supple-mented with 10% fetal bovine serum, 2 mM glutamine, penicillin(100 U/ml), and streptomycin (100 mg/ml). The CRLE 2 and 1AMLE6 mouse epithelial cell lines (Sax et al., 1995), kind gifts from Dr.Christina M. Sax (NEI, NIH, Bethesda, MD), were propagated inminimum essential medium supplemented with 5% rabbit serum,5% fetal bovine serum, 2 mM glutamine, penicillin (100 U/ml) andstreptomycin (100 mg/ml). The cells were treated with murinerecombinant IFNg (Life Technologies, Gaithersburg, MD) at aoncentration of 100 U/ml for 2 h at 37°C, 5% CO2. Some cells
were treated with medium containing the protein synthesis inhib-itor, cycloheximide (CHX) (Sigma, St. Louis, MO) at 35 mg/ml for30 min followed by addition of IFNg and incubation for 2 h.
Reverse-Transcribed Polymerase Chain Reaction(RT-PCR)
Total RNA was isolated from 6-week-old WT or TR mouse lensesor from lens cell lines by a modification of the phenol–guanidineisothiocyanate single-step method (Chomczynski and Sacchi, 1987) asdescribed for the TRIzol Reagent (Life Technologies). Lenses werecarefully dissected and washed before RNA isolation to avoid anypossible contamination by other tissues. All RNA samples weredigested with RNase-free DNase 1 (Life Technologies) for 30 min andpurified by phenol–chloroform extraction and precipitation in 0.4 MLiCl. cDNA synthesis was performed at 42°C for 1 h with 10 mg oftotal RNA, 0.3 mg oligo(dT)(12-16) and 1000 U Superscript ReverseTranscriptase II (Life Technologies) in a final volume of 50 ml. For each
NA preparation, a negative control reaction was performed withouteverse transcriptase. After purification of the cDNA by LiCl precipi-ation, hot-start PCR assays were performed with AmpliTaq GoldNA polymerase (Perkin–Elmer, Foster City, CA). Samples were
ncubated at 95°C for 10 min to activate the AmpliTaq Gold andmplification was carried out for 25 cycles at 94°C for 45 s, 63°C for5 s, and 72°C for 1 min. This was followed by a final 10-minxtension at 72°C. All the primer pairs used for PCR amplificationspanned at least one intron, making it possible to distinguish betweenmplification products derived from cDNA and those resulting fromny contaminating genomic DNA templates. The sequences of theCR primers used are for mouse b-actin, 59-GTGGGCC-CTCTAGGCACCAA-39 and 59-TCGTTGCCAATAGT-ATGACTTGGC-39 (Alonso et al., 1986); for mouse G3PDH,9-TGAAGGTCGGTGTGAACGGATTTGGC-3 9 and 5 9-ATGTAGGCCATGAGGTCCACCAC-39 (Sabath et al., 1990); forouse IRF-1, 59-TGAGACCCTGGCTAGAGATGC-39 and 59-CTCAGAGAGACTGCTGCTGACGAC-39 (Pine et al., 1990); forouse IRF-2, 59-AGCATCAACCAGGAATAGATAAAC-39 and 59-TAGGTGTTCCGTGTCCCCAT-39 (Harada et al., 1989); for mouse
CSBP, 59-GCTGCGGCAGTGGCTGATCGAACAGATCG-39 and9-AGTGGCAGGCC TGCACTGGGCTGCTG-39 (Driggers et al.,
1990). For Southern blot analysis, the amplified fragments wereelectrophoresed in agarose gels, transferred onto Hybond N1 nylonmembrane (Amersham, Arlington Heights, IL), and probed with
fluorescein–dUTP 39-end-labeled oligonucleotide probe, internal tothe corresponding PCR primers. Probe labeling and signal detections of reproduction in any form reserved.

(
f
thwA
0
Un
Crc(ShE
fiumBiw
mtr(
10
tG
iGSP
sds(
46 Li et al.
were performed with the ECL 39-oligolabeling and detection systemAmersham).
Northern Blot Analysis
Total RNA (30 mg) was fractionated on a 0.8% agarose-ormaldehyde gel, transferred to Hybond N1 membrane (Amer-
sham), and hybridized for 12 h at 65°C in hybridization solutioncontaining 5 3 106 cpm/ml probe as described (Ausubel et al.,1995). IRF-1, IRF-2, ICSBP, or b-actin-specific cDNA fragmentswere labeled to high specific activity (.108 cpm/mg) with[a-32P]dCTP by random priming (oligolabeling kit; Pharmacia Bio-ech, Piscataway, NJ) and used as hybridization probes. After twoigh-stringency washes in 0.13 SSC, 0.1% SDS at 65°C, signalsere detected by autoradiography at 270°C with Kodak X-OmatR film and Cronex intensifying screens.
Western Blot and Immunoprecipitation Analysis
Six-week-old mouse lenses (derived from WT BALB/c and IFNgBALB/c transgenic litter mates) or cultured lens cells were dis-rupted in 50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1% Nonidet-P40, 0.5% sodium deoxycholate, 2 mM leupeptin, 2 mM pepstatin,.1 mM aprotinin, 1 mM AEBSF, 0.5 mM phenylmethylsulfonyl
fluoride, and 1 mM E-64 on ice. Extracts were clarified by centrif-ugation and protein levels determined by Coomassie blue-dyebinding method as recommended for Coomassie Plus Protein AssayReagent kit (Pierce, Rockford, IL). For immunoprecipitation, WT (5mg protein) or IFNg transgenic (0.5 mg protein) mouse lens extractwas incubated with protein G agarose (Pharmacia) and anti-STAT1antibody for 1 h at 4°C and immunoprecipitates were washed fourtimes in lysis buffer. For analysis of the lens epithelial cell lines,cells were cultured for 2 h in medium or medium containing 100
/ml IFNg. The cells were lysed and fractionated to cytosolic anduclear fractions as previously described (Politis et al., 1994). All
samples were heated for 10 min at 95°C in 13 sample buffer andelectrophoresed in 10% SDS/PAGE. Gels were electroblotted ontopolyvinylidene fluoride membranes, blocked with 5% nonfat milk,and probed with one of the following antibodies: IRF-1 (M-20),IRF-2 (C-19), ICSBP (C-19), ICSAT (M-17) (also designated LSIRF/Pip), or Stat1a p91 (C-111) from Santa Cruz Biotech (Santa Cruz,
A). Rabbit polyclonal anti-mouse ICSBP from Zymed Laborato-ies, Inc. (San Francisco, CA). were also utilized. Mouse aA-rystallin-specific antibody was kindly provided by Sam ZiglerNEI/NIH). Preimmune serum was also used in parallel as controls.ignals were detected with anti-rabbit-, anti-mouse-, or anti-goatorseradish peroxidase conjugated-secondary antibodies using theCL system (Amersham).
Immunohistochemistry
Seventeen (E17)- or sixteen (E16)-day mouse embryos were fixedin 4% paraformaldehyde and embedded in Ameraffin tissue em-bedding medium (Baxter). Tissue sections (5 mm) were deparaf-
nized in xylene, rehydrated through a graded alcohol series, andsed for immunostaining by the avidin–biotin–peroxidase complexethod (Vectastain Elite ABC kit PK-6105; Vector Laboratories,
urlingame, CA.). After preincubation for 30 min with 2% block-ng serum, sections were incubated for 2 h at room temperatureith antibodies (2 mg/ml) specific to mouse ICSBP, IRF-1, or IRF-2
(Santa Cruz). Sections were subsequently incubated with biotinyl-ated secondary antibody for 30 min at room temperature and signal
ve
Copyright © 1999 by Academic Press. All right
was visualized with diaminobenzidine–H2O2 as recommended(Vector). In all our immunolocalization experiments, three serialsections were fixed onto the same glass slide; one section served asa negative control and was incubated with normal preimmuneserum, another section received the primary antibody, and thethird section received the primary antibody and 10-fold excess of aneutralizing peptide specific to immunogenic epitopes of therelevant IRF protein. More than three independent experimentswere performed for each antibody. The experimental sectionsshowed identical antibody-staining patterns while the negativecontrol section consistently showed no immunological reactivity.The section containing 10-fold excess of the peptide consistentlyshowed no significant immunoreactivity. The neutralizing effectsof the blocking peptides were found to be concentration-dependent.
Electrophoretic Mobility Shift Assay (EMSA)
Lens nuclear extracts were prepared either from 1- to 3-day-oldWT CD-1 mouse lenses or from cultured lens epithelial cells aspreviously described (Schreiber et al., 1989). Buffer used for nuclearprotein extraction contained the following protease inhibitors: 2mM leupeptin, 2 mM pepstatin, 0.1 mM aprotinin, 1 mM AEBSF, 0.5
M phenylmethylsulfonyl fluoride, 1 mM E-64. Protein concentra-ion was determined by the Coomassie blue dye-binding method asecommended for Coomassie Plus Protein Assay Reagent kitPierce) and extracts were stored at 270°C until use. Nuclearextract (10 mg) in binding buffer (20 mM Hepes, pH 7.9, 50 mM KCl,0% glycerol, 0.5 mM dithiothreitol, 0.1 mM EDTA) containing.14 mg/ml poly(dI–dC) was incubated on ice for 10 min. 32P-59-end-
labeled double-stranded DNA probe (50,000 cpm) was then addedand incubated for an additional 20 min on ice. Samples wereelectrophoresed in 4% polyacrylamide gel in 0.53 Tris–borate–EDTA buffer. For competition experiments, the nuclear extractwas preincubated with unlabeled probe and poly(dI–dC) for 20 minon ice prior to the addition of labeled probe. The sequences used forthe double-stranded probes or competitors are ICSBP pIRE/GAS,59-AGTGATTTCTCGGAAAGAGAGCGCTT C-39 (2175 to2149); ICSBP-IRE 1, 59-GTAAAGAGAGAAAAGGACTC-39 (2217o 2198) (Kanno et al., 1993); and ISRE, GATCGATCCTCGG-AAAGGGAAACCGAAACTGAAGCC (Decker et al., 1997). For
supershift assays, the indicated antibody was added to the bindingbuffer containing the nuclear extract and preincubated on ice for 10min. The 32P-labeled probe was then added and the entire mixturencubated for an additional 20 min on ice before electrophoresis.el-shift-grade anti-mouse ICSBP, IRF-1, or IRF-2 (Santa Cruz) andTAT1a polyclonal antibodies (Upstate Biotechnology, Inc., Lakelacid, NY) were used.
RESULTS
Effect of IFNg Transgene on Lens Development
We previously generated TR rats and mice with targetedexpression of IFNg in the lens to study the role of ectopicexpression of IFNg in enhanced susceptibility to organ-pecific autoimmunity (Egwuagu et al., 1999, 1994a). Theisruption of the lens differentiation program and theevere microphthalmia manifested by these rodentsEgwuagu et al., 1994b) provided the opportunity to study in
ivo the molecular mechanisms underlying the regulatoryffects of IFNg on the growth and differentiation of normals of reproduction in any form reserved.
leD
47Differential Expression of IRFs in the Lens
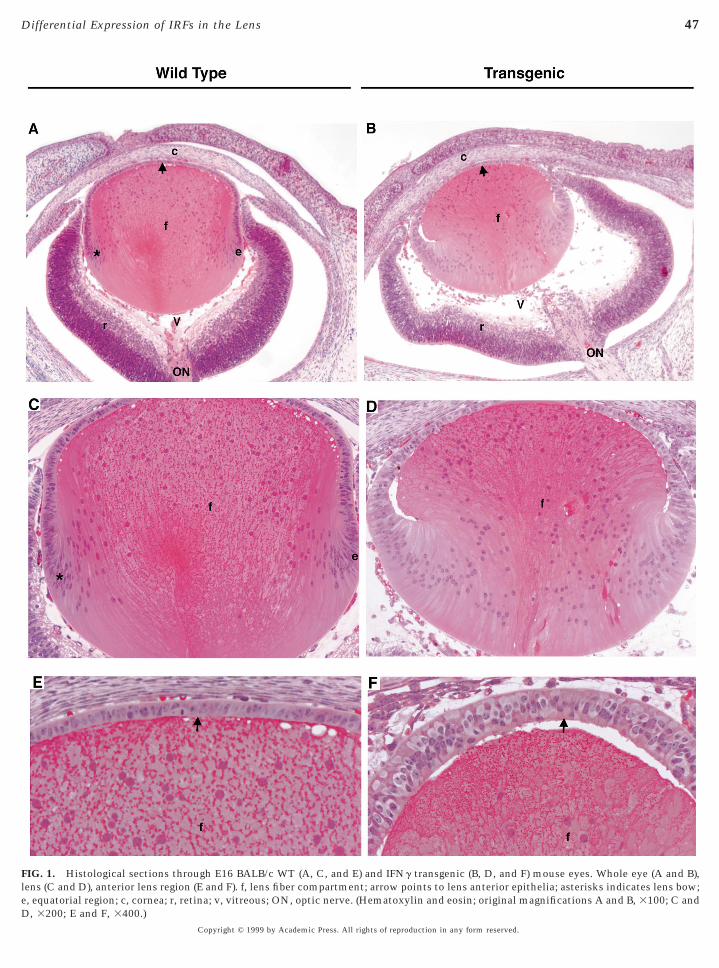
FIG. 1. Histological sections through E16 BALB/c WT (A, C, and E) and IFNg transgenic (B, D, and F) mouse eyes. Whole eye (A and B),ens (C and D), anterior lens region (E and F). f, lens fiber compartment; arrow points to lens anterior epithelia; asterisks indicates lens bow;
, equatorial region; c, cornea; r, retina; v, vitreous; ON, optic nerve. (Hematoxylin and eosin; original magnifications A and B, 3100; C and, 3200; E and F, 3400.)Copyright © 1999 by Academic Press. All rights of reproduction in any form reserved.

ctfedGbsatttaeweattda
nestacpil
eiaaI(tdimah
gc
waIsbltsWtLnthfidr
48 Li et al.
epithelial tissues (Saunders et al., 1996). The vertebrate lensomprises of epithelial cells at varying stages of differentia-ion (Piatigorsky, 1981; McAvoy, 1981). The anterior sur-ace is covered by a monolayer of undifferentiated cuboidalpithelial cells and the remainder of the lens is populated byifferentiating or terminally differentiated lens fiber cells.rowth of the normal mouse lens (Figs. 1A and 1C) occursy proliferation of anterior epithelial cells and their progres-ive displacement posteriorly toward the lens equator. Cellst the equatorial zone stop dividing and initiate a differen-iation program that culminates in the terminally differen-iated lens fibers. In the IFNg TR mouse, lens differentia-ion is perturbed during embryonic development (Figs. 1Bnd 1D). The anterior epithelial cells extend beyond thequatorial plate to the posterior pole, failing to form aell-defined lens bow characteristic of normal lens differ-
ntiation. Unlike the WT mouse lens with a monolayer ofnterior epithelial cells (Fig. 1E), the anterior epithelia ofhe TR lens is thicker and appears multilayered (Fig. 1F). Inhe adult TR mouse, the lens is disorganized and terminallyifferentiated denucleated fiber cells are replaced by nucle-ted gigantic balloon-like cells (Egwuagu et al., 1994a,b).
Constitutive and Inducible Expression of IRFFactors in the Mouse Lens
IFNg is a potent inducer of squamous differentiation inormal human epidermal keratinocytes and its effects arexerted through control of regulatory and differentiation-pecific genes (Saunders and Jetten, 1994). As transcrip-ional regulation by IFNg is mediated by activated STAT1nd IRFs, and some IRFs have been shown to be importantell cycle regulators, we addressed the possibility that theerturbation of the physiological levels of STAT1 and IRFsn the developing mouse lens may underlie the abnormalens differentiation in the IFNg TR mice. We therefore
examined the levels of IRFs and STAT1 expression in WTand TR mouse lenses. RT-PCR and Western blot analyses oflenses from 6-week-old BALB/c WT and TR mouse litter-mates are shown in Fig. 2. As indicated by immunoprecipi-tation and Western blotting using anti-STAT1a antibody(Fig. 2A), STAT1 is present in the lens and its level ismarkedly increased in the TR compared to the WT lens,providing direct evidence that lens cells are responsive toIFNg signaling. This is the first documented evidence of thexpression of a STAT protein in the normal lens. However,t should be emphasized that the level of STAT1 is very lownd can be detected only by immunoprecipitation. RT-PCRnalysis of RNAs from these mice show that ICSBP andRF-2 genes are constitutively expressed in the mouse lensFig. 2B). Although IRF-1 transcripts are not detectable inhe WT lens after 25 cycles of PCR amplification, they areetectable after 35 cycles of amplification (data not shown),ndicating that their levels are relatively low in the normal
ouse lens. In the IFNg TR mouse lens, the levels of IRF-1
nd ICSBP transcripts are significantly increased. The en-anced transcription of IRF-1 and ICSBP genes, two IRFCopyright © 1999 by Academic Press. All right
enes that contain GAS elements in their promoters, isonsistent with their activation by STAT1 in the IFNg TR
lens. These results have been confirmed by RT-PCR analy-ses of at least six independent RNA preparations and theauthenticity of the lens IRF transcripts has been verified bycDNA sequencing; the IRF-1, IRF-2, and ICSBP transcriptsisolated from the lens were found to be identical to pub-lished sequences reported for the corresponding IRFs inmouse hematopoietic cells.
Detection of ICSBP transcripts in the WT lens wasunexpected because transcription of the ICSBP gene isthought to be restricted to hematopoietic cells (Boehm etal., 1997; Driggers et al., 1990; Holtschke et al., 1996). It
as therefore necessary to establish that the ICSBP protein,s well as other IRF proteins, is expressed in the lens.ndeed, ICSBP and IRF-2 proteins are constitutively synthe-ized in the normal mouse lens as indicated by Westernlotting of lens extracts (Fig. 2C). We also analyzed theenses of another mouse strain to rule out the possibilityhat expression of ICSBP in the lens is an epiphenomenoneen only in the BALB/c mouse strain. As shown on theestern blot (Fig. 2D), the ICSBP protein is also present in
he FVB/N mouse lens. To our surprise we also detectedSIRF/Pip protein in the FVB/N (Fig. 2D) and BALB/c (dataot shown) mouse lenses. LSIRF/Pip is another member ofhe IRF family thought to be exclusively expressed inematopoietic cells (Matsuyama et al., 1995). This is therst time that expression of ICSBP or LSIRF/Pip has beenemonstrated in a tissue that is not directly involved inegulating immune responses.
Expression of IRF Transcription Factors IsSpatially Regulated in the Lens
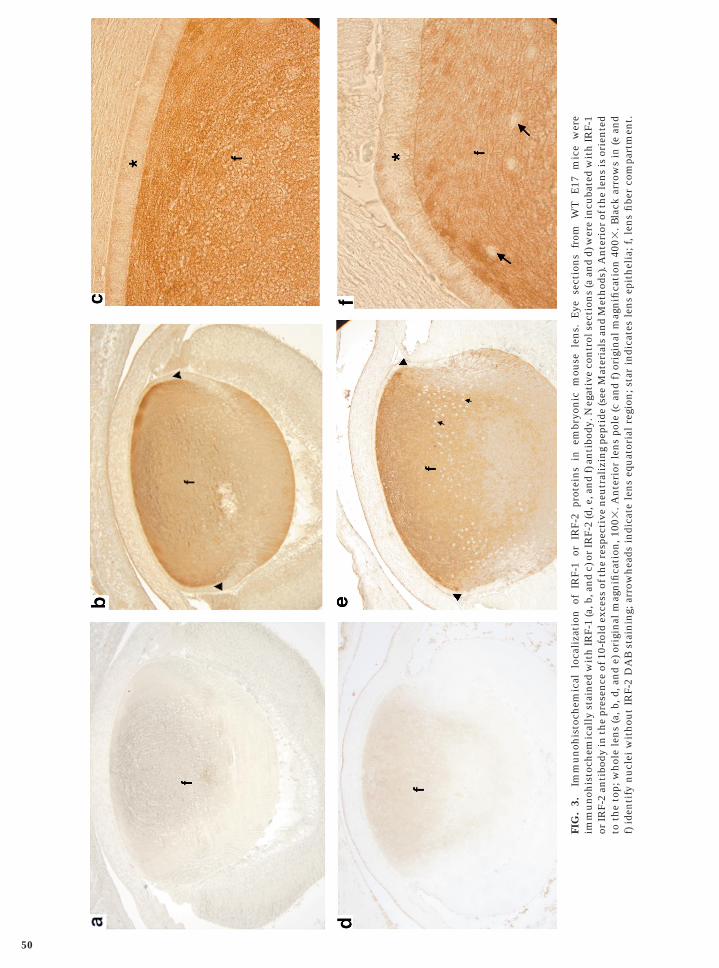
As proliferating and differentiated cells are spatially seg-regated in the lens, we examined whether expression ofIRF-1, IRF-2, and ICSBP is spatially regulated in the lens.Results of immunolocalization of IRF-1 and IRF-2 in E17mouse eye sections are shown in Fig. 3. Both IRF-1 andIRF-2 proteins are predominantly detected in the lens fibercell compartment. However, very little expression of eitherprotein is detectable in the anterior epithelia. Interestingly,the IRF-2 protein appears to be exclusively expressed in thecytoplasm as the lens cell nuclei are not stained by theantibodies (see arrows in Fig. 3f). On the other hand, theIRF-1 protein is present in both the cytoplasm and thenuclei of the fiber cells. However, not all of the cells shownuclear staining, particularly some of the more superficialnuclei (Fig. 3b).
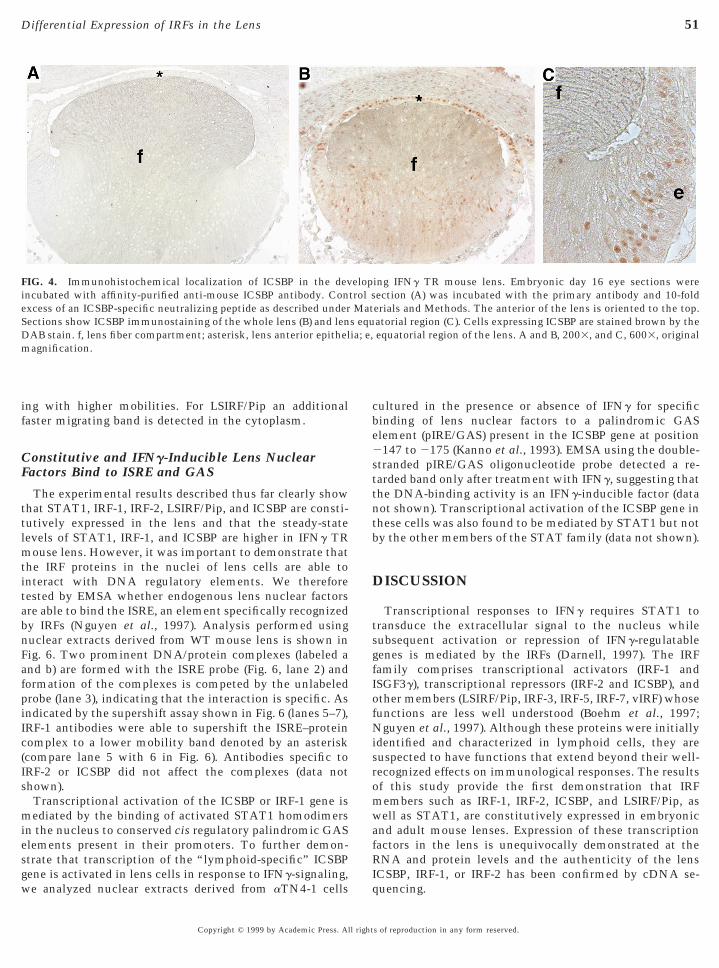
In the WT lens, the level of ICSBP is very low and barelydetectable. However, the IFNg TR mice with upregulatedexpression of ICSBP mRNA (Fig. 2B) and protein (Fig. 2C)allowed for delineation of ICSBP-expressing lens cells. Re-sults of ICSBP immunolocalization in E16 TR mouse eyesare shown in Fig. 4. As shown in Figs. 4B and 4C, ICSBP
localizes to the nuclei of cells at the anterior epithelia, atthe lens equator, and at the posterior pole. In situ hybrid-s of reproduction in any form reserved.

mired t
WbiasI
pI d by6
49Differential Expression of IRFs in the Lens
ization analysis of embryonic eye sections from IFNg TRice showed a similar pattern of ICSBP mRNA localization
n the lens (data not shown). Similar immunolocalizationesults of IRF-1, IRF-2, or ICSBP have been obtained usingye sections of mice at days 16 through 20 of embryonicevelopment (data not shown).
Lens Epithelial Cells Are Able to Receive andTransduce IFNg Signals
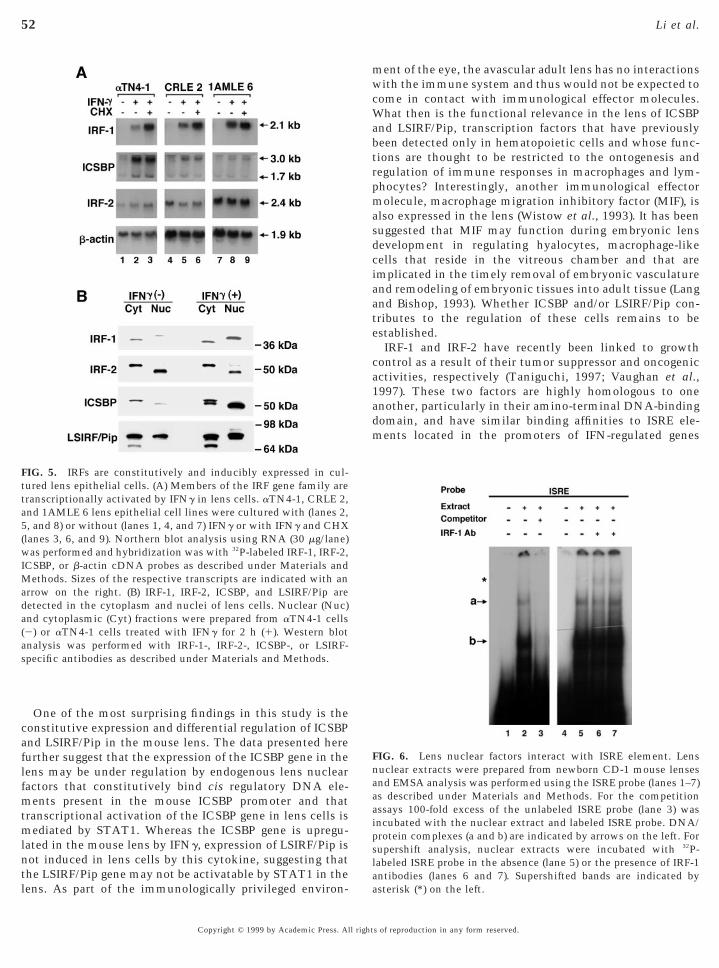
To rule out the possibility that the IRF mRNA transcriptsor proteins detected in the mouse lens derived from con-taminating lymphoid cells, it is important to reproduce ourfindings in lens cells in culture. Consequently, we have alsoanalyzed three well-characterized lens epithelial cell lines(see Materials and Methods) to confirm that IRFs areconstitutively and inducibly expressed in lens cells.aTN4-1, CRLE 2, and 1AMLE 6 lens cells were treated withmouse IFNg either in the presence or in the absence ofCHX. RNA was isolated from the various treatment groupsand used for Northern analyses. In each of the cell lines,two ICSBP mRNAs of 3.0 and 1.7 kb and a single IRF-2transcript of 2.4 kb are detected (Fig. 5A). The sizes of thesetranscripts are similar to the ones reported for these genesin mouse hematopoietic cells (Driggers et al., 1990; Kannoet al., 1993). Although IRF-1 transcripts are not detectablein the untreated cells in the Northern blot shown here, theyare detectable after longer exposure of the autoradiogram(data not shown). Detection of these transcripts in unstimu-lated lens cells confirms that these IRF transcription factors
FIG. 2. STAT1a, ICSBP, IRF-2, and LSIRF/Pip are constitutivelylenses were used to prepare whole lens extracts or poly(A) RNAs. (from 6-week-old WT (5 mg protein) or TR (0.5 mg protein) mouse litfor details). (B) RT-PCR and Southern blot analyses for expression ofragments are ICSBP, 627 bp; IRF-1, 563 bp; IRF-2, 626 bp; G3PDHrotein/lane) or IFNg TR (50 mg protein/lane) mouse litter-matesRF-2, or aA-crystallin. (D) ICSBP or LSIRF/Pip protein was detecte-week-old WT FVB/N mouse lens.
are constitutively expressed in lens cells. Furthermore, thelevels of IRF-1 and ICSBP transcripts are significantly
Copyright © 1999 by Academic Press. All right
increased in cells treated with IFNg, indicating that thesegenes are transcriptionally activated by IFNg. The lowabundance of the IRF-1 transcript could be attributed to itsrelatively short half-life (,30 min) (Taniguchi et al., 1995).This issue was examined by treatment of cells with IFNg inhe presence of the protein synthesis inhibitor CHX.
hereas lens ICSBP and IRF-2 mRNAs levels are unaffectedy CHX, the level of IRF-1 transcripts is significantlyncreased, indicating that lens IRF-1 mRNA is short-livednd suggesting that it is rapidly degraded by de novoynthesized nucleases that are concomitantly induced byFNg (Kalvakolanu and Borden, 1996). Similar effects of
CHX on the stability of IRF-1 mRNA have previously beenreported (Pine et al., 1990) It is important to note that theaTN4-1 line is a transformed cell line as the cells expressSV40 T antigen (Yamada et al., 1990). However, we ob-tained similar results with two nontransformed lens epithe-lial cell lines (CRLE 2 and 1AMLE6) (Sax et al., 1995),suggesting that these results are generally applicable to lensepithelial cells.
To further confirm that IRF-1, IRF-2, ICSBP, and LSIRF/Pip proteins are indeed synthesized in lens cells, lens cellsthat were cultured for 2 h in the presence or absence of IFNg
were fractionated into cytoplasmic and nuclear fractionsand analyzed by Western blotting. As indicated in Fig. 5B,each of these proteins is detected in both the cytoplasm andthe nucleus of the lens cells. However, the cytoplasmic andnuclear forms exhibit distinct mobilities in SDS/PAGE.Cytoplasmic IRF-1 exhibits a higher mobility compared to
ssed in mouse lens. Six-week-old BALB/c WT or IFNg TR mouseAT1 was immunoprecipitated from whole lens extracts prepared
ates using anti-mouse STAT1 antibody (see Materials and MethodsBP, IRF-1, IRF-2, or G3PDH mRNAs. Sizes of the amplified DNAbp. (C) Whole lens extracts from 6-week-old BALB/c WT (100 mg
immunoblotted with antibodies specific for mouse ICSBP, IRF-1,Western blotting of whole-cell extracts (100 mg protein/lane) from
expreA) STtermf ICS, 983
were
its nuclear counterparts. On the other hand, the reverse istrue for ICSBP and IRF-2 with the nuclear proteins migrat-
s of reproduction in any form reserved.
FIG
.3.
Imm
un
ohis
toch
emic
allo
cali
zati
onof
IRF-
1or
IRF-
2pr
otei
ns
inem
bryo
nic
mou
sele
ns.
Eye
sect
ion
sfr
omW
TE
17m
ice
wer
eim
mu
noh
isto
chem
ical
lyst
ain
edw
ith
IRF-
1(a
,b,a
nd
c)or
IRF-
2(d
,e,a
nd
f)an
tibo
dy.N
egat
ive
con
trol
sect
ion
s(a
and
d)w
ere
incu
bate
dw
ith
IRF-
1or
IRF-
2an
tibo
dyin
the
pres
ence
of10
-fol
dex
cess
ofth
ere
spec
tive
neu
tral
izin
gpe
ptid
e(s
eeM
ater
ials
and
Met
hod
s).A
nte
rior
ofth
ele
ns
isor
ien
ted
toth
eto
p;w
hol
ele
ns
(a,b
,d,a
nd
e)or
igin
alm
agn
ifica
tion
,100
3.A
nte
rior
len
spo
le(c
and
f)or
igin
alm
agn
ifica
tion
4003
.Bla
ckar
row
sin
(ean
df)
iden
tify
nu
clei
wit
hou
tIR
F-2
DA
Bst
ain
ing;
arro
wh
eads
indi
cate
len
seq
uat
oria
lre
gion
;sta
rin
dica
tes
len
sep
ith
elia
;f,l
ens
fibe
rco
mpa
rtm
ent.
50
mtitabnFafpiIc(Is
miesg
be2
t
ofNisromwafR
ia; e,
51Differential Expression of IRFs in the Lens
ing with higher mobilities. For LSIRF/Pip an additionalfaster migrating band is detected in the cytoplasm.
Constitutive and IFNg-Inducible Lens NuclearFactors Bind to ISRE and GAS
The experimental results described thus far clearly showthat STAT1, IRF-1, IRF-2, LSIRF/Pip, and ICSBP are consti-tutively expressed in the lens and that the steady-statelevels of STAT1, IRF-1, and ICSBP are higher in IFNg TR
ouse lens. However, it was important to demonstrate thathe IRF proteins in the nuclei of lens cells are able tonteract with DNA regulatory elements. We thereforeested by EMSA whether endogenous lens nuclear factorsre able to bind the ISRE, an element specifically recognizedy IRFs (Nguyen et al., 1997). Analysis performed usinguclear extracts derived from WT mouse lens is shown inig. 6. Two prominent DNA/protein complexes (labeled and b) are formed with the ISRE probe (Fig. 6, lane 2) andormation of the complexes is competed by the unlabeledrobe (lane 3), indicating that the interaction is specific. Asndicated by the supershift assay shown in Fig. 6 (lanes 5–7),RF-1 antibodies were able to supershift the ISRE–proteinomplex to a lower mobility band denoted by an asteriskcompare lane 5 with 6 in Fig. 6). Antibodies specific toRF-2 or ICSBP did not affect the complexes (data nothown).Transcriptional activation of the ICSBP or IRF-1 gene isediated by the binding of activated STAT1 homodimers
n the nucleus to conserved cis regulatory palindromic GASlements present in their promoters. To further demon-trate that transcription of the “lymphoid-specific” ICSBP
FIG. 4. Immunohistochemical localization of ICSBP in the devincubated with affinity-purified anti-mouse ICSBP antibody. Conexcess of an ICSBP-specific neutralizing peptide as described underSections show ICSBP immunostaining of the whole lens (B) and lenDAB stain. f, lens fiber compartment; asterisk, lens anterior epithelmagnification.
ene is activated in lens cells in response to IFNg-signaling,we analyzed nuclear extracts derived from aTN4-1 cells
Iq
Copyright © 1999 by Academic Press. All right
cultured in the presence or absence of IFNg for specificinding of lens nuclear factors to a palindromic GASlement (pIRE/GAS) present in the ICSBP gene at position147 to 2175 (Kanno et al., 1993). EMSA using the double-
stranded pIRE/GAS oligonucleotide probe detected a re-tarded band only after treatment with IFNg, suggesting thathe DNA-binding activity is an IFNg-inducible factor (data
not shown). Transcriptional activation of the ICSBP gene inthese cells was also found to be mediated by STAT1 but notby the other members of the STAT family (data not shown).
DISCUSSION
Transcriptional responses to IFNg requires STAT1 totransduce the extracellular signal to the nucleus whilesubsequent activation or repression of IFNg-regulatablegenes is mediated by the IRFs (Darnell, 1997). The IRFfamily comprises transcriptional activators (IRF-1 andISGF3g), transcriptional repressors (IRF-2 and ICSBP), andther members (LSIRF/Pip, IRF-3, IRF-5, IRF-7, vIRF) whoseunctions are less well understood (Boehm et al., 1997;guyen et al., 1997). Although these proteins were initially
dentified and characterized in lymphoid cells, they areuspected to have functions that extend beyond their well-ecognized effects on immunological responses. The resultsf this study provide the first demonstration that IRFembers such as IRF-1, IRF-2, ICSBP, and LSIRF/Pip, asell as STAT1, are constitutively expressed in embryonic
nd adult mouse lenses. Expression of these transcriptionactors in the lens is unequivocally demonstrated at theNA and protein levels and the authenticity of the lens
ing IFNg TR mouse lens. Embryonic day 16 eye sections wereection (A) was incubated with the primary antibody and 10-folderials and Methods. The anterior of the lens is oriented to the top.atorial region (C). Cells expressing ICSBP are stained brown by theequatorial region of the lens. A and B, 2003, and C, 6003, original
eloptrol sMat
s equ
CSBP, IRF-1, or IRF-2 has been confirmed by cDNA se-uencing.
s of reproduction in any form reserved.
ntl
mwcWabtrpmasdciaate
ca1adm
a5
w
Mada(
52 Li et al.
One of the most surprising findings in this study is theconstitutive expression and differential regulation of ICSBPand LSIRF/Pip in the mouse lens. The data presented herefurther suggest that the expression of the ICSBP gene in thelens may be under regulation by endogenous lens nuclearfactors that constitutively bind cis regulatory DNA ele-ments present in the mouse ICSBP promoter and thattranscriptional activation of the ICSBP gene in lens cells ismediated by STAT1. Whereas the ICSBP gene is upregu-lated in the mouse lens by IFNg, expression of LSIRF/Pip is
ot induced in lens cells by this cytokine, suggesting that
FIG. 5. IRFs are constitutively and inducibly expressed in cul-tured lens epithelial cells. (A) Members of the IRF gene family aretranscriptionally activated by IFNg in lens cells. aTN4-1, CRLE 2,nd 1AMLE 6 lens epithelial cell lines were cultured with (lanes 2,, and 8) or without (lanes 1, 4, and 7) IFNg or with IFNg and CHX
(lanes 3, 6, and 9). Northern blot analysis using RNA (30 mg/lane)as performed and hybridization was with 32P-labeled IRF-1, IRF-2,
ICSBP, or b-actin cDNA probes as described under Materials andethods. Sizes of the respective transcripts are indicated with an
rrow on the right. (B) IRF-1, IRF-2, ICSBP, and LSIRF/Pip areetected in the cytoplasm and nuclei of lens cells. Nuclear (Nuc)nd cytoplasmic (Cyt) fractions were prepared from aTN4-1 cells2) or aTN4-1 cells treated with IFNg for 2 h (1). Western blotanalysis was performed with IRF-1-, IRF-2-, ICSBP-, or LSIRF-specific antibodies as described under Materials and Methods.
he LSIRF/Pip gene may not be activatable by STAT1 in theens. As part of the immunologically privileged environ-
Copyright © 1999 by Academic Press. All right
ent of the eye, the avascular adult lens has no interactionsith the immune system and thus would not be expected to
ome in contact with immunological effector molecules.hat then is the functional relevance in the lens of ICSBP
nd LSIRF/Pip, transcription factors that have previouslyeen detected only in hematopoietic cells and whose func-ions are thought to be restricted to the ontogenesis andegulation of immune responses in macrophages and lym-hocytes? Interestingly, another immunological effectorolecule, macrophage migration inhibitory factor (MIF), is
lso expressed in the lens (Wistow et al., 1993). It has beenuggested that MIF may function during embryonic lensevelopment in regulating hyalocytes, macrophage-likeells that reside in the vitreous chamber and that aremplicated in the timely removal of embryonic vasculaturend remodeling of embryonic tissues into adult tissue (Langnd Bishop, 1993). Whether ICSBP and/or LSIRF/Pip con-ributes to the regulation of these cells remains to bestablished.IRF-1 and IRF-2 have recently been linked to growth
ontrol as a result of their tumor suppressor and oncogenicctivities, respectively (Taniguchi, 1997; Vaughan et al.,997). These two factors are highly homologous to onenother, particularly in their amino-terminal DNA-bindingomain, and have similar binding affinities to ISRE ele-ents located in the promoters of IFN-regulated genes
FIG. 6. Lens nuclear factors interact with ISRE element. Lensnuclear extracts were prepared from newborn CD-1 mouse lensesand EMSA analysis was performed using the ISRE probe (lanes 1–7)as described under Materials and Methods. For the competitionassays 100-fold excess of the unlabeled ISRE probe (lane 3) wasincubated with the nuclear extract and labeled ISRE probe. DNA/protein complexes (a and b) are indicated by arrows on the left. Forsupershift analysis, nuclear extracts were incubated with 32P-labeled ISRE probe in the absence (lane 5) or the presence of IRF-1
antibodies (lanes 6 and 7). Supershifted bands are indicated byasterisk (*) on the left.s of reproduction in any form reserved.

Itplfinotpt
ocntmItokdcatlBelp
fss
tletpt(1fiInIbb(fidIedlepI
rst
53Differential Expression of IRFs in the Lens
(Miyamoto et al., 1988; Harada et al., 1989). Both IRF-1 andRF-2 are expressed at low constitutive levels in many cellypes. In the embryonic mouse lens, the level of IRF-1 inroliferating cells of the lens epithelia compartment is veryow. In contrast, IRF-1 levels are relatively higher in theber compartment that contains differentiating and termi-ally differentiated lens cells. This result is consistent withbserved increase in IRF-1 levels in other differentiated cellypes (Harada et al., 1990). However, similar to otherroteins that are preferentially expressed by fiber cells (e.g.,he major intrinsic protein of the lens (MIP) and
g-crystallins), IRF-1 mRNA levels decrease significantlyafter birth. This may account for the very low level of IRF-1mRNA in 6-week-old mouse lenses compared to the levelsin embryonic lenses. IRF-2 is also expressed in the embry-onic mouse lens and its spatial distribution is similar tothat of IRF-1. However, the level of IRF-2 is significantlyhigher than that of IRF-1 in the lens as indicated by ourNorthern and Western blot analyses. In contrast to IRF-1,IRF-2 appears to localize to the cytoplasm of lens cells inthe embryonic mouse lens.
We found IRF-1, IRF-2, ICSBP, and LSIRF/Pip to bepresent in cytosolic and nuclear fractions of cultured lenscells. However, the IRF proteins present in the nucleusexhibit electrophoretic mobilities in SDS/PAGE that aredistinct from their counterparts in the cytoplasm. Thesechanges are possibly due to posttranslational modifications;thus nuclear localization of IRF-1 may be associated withphosphorylation while nuclear localization of ICSBP orIRF-2 may derive from dephosphorylation events. Thesepredictions remain to be verified. To our knowledge, this isthe first study of subcellular localization of IRFs. Thepresence of IRFs in the nucleus and cytoplasm suggests thepossibility that IRFs may have distinct functions in thesesubcellular compartments. Thus, nuclear ICSBP, IRF-1,IRF-2, and LSIRF/Pip may function as transcription factors,while their cytoplasmic counterparts may function to se-quester other regulatory proteins in the cytoplasm throughprotein–protein interactions (Drew et al., 1995a,b). On thether hand, interactions of IRFs with other proteins in theytoplasm may also serve to regulate their transport to theucleus. Further studies are therefore needed to elucidatehe functions of IRFs in the cytoplasm and the regulatoryechanisms that control their translocation to the nucleus.
t is important to note that selective localization to eitherhe cytoplasm or the nucleus has also been observed forther lens proteins. For example, the cyclin-dependentinase 5 (cdk5), a protein associated with elongation andenucleation of differentiating fiber cells, is present in theytoplasm of epithelial and fiber cells of the rat lens (Gao etl., 1997). However, in the final stages of lens differentia-ion, immediately prior to denucleation, the cdk5 proteinocalizes to the nuclei of fiber cells (Gao et al., 1997). Cyclin2 has also been shown to localize to the cytoplasm in thepithelia and in the nuclei of primary fibers of the chicken
ens (Gao et al., 1995). Differential localization of lensroteins to distinct subcellular compartments may there-Copyright © 1999 by Academic Press. All right
ore be an important regulatory mechanism by whichpecific biochemical pathways are temporally and spatiallyegregated in the lens.The anatomical segregation of undifferentiated, differen-
iating, and terminally differentiated cells in the vertebrateens suggests that differentiation-specific genes are differ-ntially activated or repressed in different compartments ofhe lens. Characterization of regulatory elements in theromoters of lens-specific genes have led to the identifica-ion of several transcription factors such as maf-1, maf-2Yoshida et al., 1997), and AP2a (Ohtaka-Maruyama et al.,998) that are differentially expressed in the epithelia andber compartments of the lens. Constitutive binding of
RF-1 to the ISRE element in lens nuclear extracts ofewborn mouse lenses provides the first evidence thatRF-1 may regulate gene transcription in the lens. It woulde of interest to know whether other endogenous ISRE-inding factors regulate gene transcription in the epitheliawhere they might regulate cell proliferation) or in lensbers (where they would more likely to be involved in cellifferentiation). The predominant expression of IRF-1 andRF-2 in lens fibers and detection of ICSBP in anteriorpithelial cells of the TR mouse lens suggest that theistribution of IRFs may be spatially regulated in the mouseens. It would therefore be of interest to examine genesxpressed in the lens that contain ISRE elements in theirromoters and determine whether they are regulated byCSBP, IRF-1, or IRF-2.
Recent characterization of the mouse MIP gene promoterevealed the presence of a GAS element proximal to an ISREite (Chepelinsky et al., unpublished data). It is interestinghat in the IFNg TR mouse lens with perturbed expression
of IRFs, expression of the gene coding for the lens fiber-specific MIP protein is completely repressed (Egwuagu etal., 1994b). Studies in progress will allow us to determinewhether transcription of the MIP gene is regulated bytranscription factors of the IRF family. It is also interestingto note that several extracellular signaling molecules (EGF,PDGF, c-fos, and retinoic acid) that have been implicated inlens differentiation utilize the JAK/STAT signaling path-way. In view of the fact that IRFs are key players in thisimportant biochemical pathway, it is important to investi-gate gene regulatory elements and transcription factors thatinteract with IRFs in the lens and examine their roles inactivation or repression of genes expressed during lensdifferentiation.
ACKNOWLEDGMENTS
We are grateful to Drs. Peggy Zelenka, Graeme Wistow, andJen-Yue Tsai for critically reading the manuscript; Drs. PaulRussell and Christina M. Sax for providing the lens epithelial celllines; Dr. Samuel Zigler for providing the mouse aA-crystallin-specific antibody; Nicole Newman for preparation of the histolog-
ical sections; Ricardo Dreyfus, Shauna Everett, Wayne Randolph,John Ward, Gary Best, Chris Vargas, and Hassan Ennaciri fors of reproduction in any form reserved.

A
B
C
D
D
D
D
D
E
E
E
H
H
K
K
L
M
M
M
N
N
O
54 Li et al.
photographic assistance; and Rashid M. Mahdi for assistance inDNA and RT-PCR analysis.
REFERENCES
Alonso, S., Minty, A., Bourlet, Y., and Buckingham, M. (1986).Comparison of three actin-coding sequences in the mouse:Evolutionary relationships between the actin genes of warmblooded vertebrates. J. Mol. Evol. 23, 11–22.usubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman,J. G., Smith, J. A., and Stuhl, K. (1995). “Current Protocols inMolecular Biology.” Wiley, New York.
oehm, U., Klamp, T., and Howard, J. C. (1997). Cellular responsesto interferon-g. Annu. Rev. Immunol. 15, 749–795.homczynski, P., and Sacchi, N. (1987). Single-step method ofRNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 162, 156–159.arnell, J. E., Jr. (1997). STATs and gene regulation. Science 277,1630–1635.ecker, T., Kovarik, P., and Meinke, A. (1997). GAS elements: Afew nucleotides with a major impact on cytokine-induced geneexpression. J. Interferon Cytol. Res. 17, 121–134.rew, P. D., Franzoso, G., Becker, K. G., Bours, V., Carlson, L. M.,Siebenlist, U., and Ozato, K. (1995a). NF kappa B and interferonregulatory factor 1 physically interact and synergistically inducemajor histocompatibility class I gene expression. J. InterferonCytol. Res. 15, 1037–1045rew, P. D., Franzoso, G., Carlson, L. M., Biddison, W. E., Sieben-list, U., and Ozato, K. (1995b). Interferon regulatory factor-2physically interacts with NF-kappa B in vitro and inhibitsNF-kappa B induction of major histocompatibility class I andbeta 2-microglobulin gene expression in transfected human neu-roblastoma cells. J. Neuroimmunol. 63, 157–62.riggers, P. H., Ennist, D. L., Gleason, S. L., Mak, W.-H., Marks,M. S., Levi, B.-Z., Flanagan, J. R., Appella, E., and Ozato, K.(1990). An interferon g-regulated protein that binds theinterferon-inducible enhancer element of major histocompatibil-ity complex class I genes. Proc. Natl. Acad. Sci. USA 87,3743–3747.
gwuagu, C. E., Sztein, J., Chan, C. C., Reid, W., Mahdi, R.,Nussenblatt, R. B., and Chepelinsky, A. B. (1994a). Ectopicexpression of gamma interferon in the eyes of transgenic miceinduces ocular pathology and MHC class II gene expression.Invest. Ophthalmol. Visual Sci. 35, 332–341.
gwuagu, C. E., Sztein, J., Chan, C. C., Mahdi, R., Nussenblatt,R. B., and Chepelinsky, A. B. (1994b). gInterferon expressiondisrupts lens and retinal differentiation in transgenic mice. Dev.Biol. 166, 557–568.
gwuagu, C. E., Sztein, J., Mahdi, R. M., Li, W., Chan, C. C., Smith,J. A., Charukamnoetkanok, P., and Chepelinsky, A. B. (1999).Interferon-g (IFNg) increases the severity and accelerates theonset of experimental autoimmune uveitis in transgenic rats.J. Immunol. 162, 510–517.
Eisenbeis, C. F., Singh, H., and Storb, U. (1995). Pip, a novel IRFfamily member, is a lymphoid-specific, PU.1-dependent tran-scriptional activator. Genes Dev. 9, 1377–1387.
Gao, C. Y., Bassnett, S., and Zelenka, P. S. (1995). Cyclin B, p34cdc2,
and H1-kinase activity in terminally differentiating lens fibercells. Dev. Biol. 169, 185–194.Copyright © 1999 by Academic Press. All right
Gao, C. Y., Zakeri, Z., Zhu, Y., He, H., and Zelenka, P. S. (1997).Expression of cdk5, p35, and cdk5-associated kinases activity inthe developing rat lens. Dev. Genet. 20, 267–275.
Harada, H., Fujita, T., Miyamoto, M., Kimura, Y., Maruyama, M.,Furia, A., Miyata, T., and Taniguchi, T. (1989). Structurallysimilar but functionally distinct factors, IRF-1 and IRF-2, bind tothe same regulatory elements of IFN and IFN-inducible genes.Cell 58, 729–739.arada, H., Willison, K., Sakakibara, J., Miyamoto, M., Fujita, T.,and Taniguchi, T. (1990). Absence of the type I IFN system in ECcells: Transcriptional activator (IRF-1) and repressor (IRF-2) genesare developmentally regulated. Cell 63, 303–312.oltschke, E., Lohler, J., Kanno, Y., Fehr, T., Giese, N., Rosenbauer,F., Lou, J., Knobeloch, K., Gabriele, L., Waring, J. F., Bachmann,M. F., Zinkernagel, R. M., Morse, H. C., III, Ozato, K., and Horak,I.. (1996). Immunodeficiency and chronic myelogenousleukemia-like syndrome in mice with a targeted mutation in theICSBP gene. Cell 87, 307–317.alvakolanu, D. V., and Borden, E. C. (1996). An overview of theinterferon system: Signal transduction and mechanisms of ac-tion. Cancer Invest. 14, 25–53.
anno, Y., Kozak, C. A., Schindler, C., Driggers, P. H., Ennist, D. L.,Gleason, S. L., Darnell, J. E., Jr., and Ozato, K. (1993). The genomicstructure of the murine ICSBP gene reveals the presence of thegamma interferon-responsive element, to which an ISGF3a subunit(or similar) molecule binds. Mol. Cell. Biol. 3, 3951–3963.
ang, R. A., and Bishop, M. J. (1993). Macrophages are required forcell death and tissue remodeling in the developing mouse eye.Cell 74, 453–452.atsuyama, T., Grossman, A., Mittrucker, H.-W., Siderovski,D. P., Kiefer, F., Kawakami, T., Richardson, C. D., Taniguchi, T.,Yoshinaga, S. K., and Mak, T. W. (1995). Molecular cloning ofLSIRF, a lymphoid-specific member of the interferon regulatoryfactor family that binds the interferon-stimulated response ele-ment (ISRE). Nucleic Acids Res. 23, 2127–2136.cAvoy, J. W. (1981). Developmental biology of the lens. In“Mechanisms of Cataract Formation in the Human Lens” (G.Duncan, Ed.), pp. 7–46. Academic Press, New York.
Mittrucker, H. W., Matsuyama, T., Grossman, A., Kundig, T. M.,Potter, J., Shahinian, A., Wakeham, A., Patterson, B., Ohashi,P. S., and Mak, T. W. (1997). Requirement for the transcriptionfactor LSIRF/IRF4 for mature B and T lymphocyte function.Science 275, 540–543.
Miyamoto, M., Fujita, T., Kimura, Y., Maruyama, M., Harada, H.,Sudo, Y., Miyata, T., and Taniguchi, T. (1988). Regulated expres-sion of a gene encoding a nuclear factor, IRF-1, that specificallybinds to the IFN-beta gene regulatory elements. Cell 54, 903–913.oore, P. S., Boshoff, C., Weiss, R. A., and Chang, Y. (1996).Molecular mimicry of human cytokine and cytokine responsepathway genes by KSHV. Science 274, 1739–1744.elson, N., Kanno, Y., Hong, C., Cantursi, C., Fujita, T., Fowlkes,B. J., O’Connell, E., Hu-li, J., Paul, W. E., Jankovic, D., Sher, A. F.,Coligan, J. E., Thorton, A., Appella, E., Yang, Y., and Ozato, K.(1996). Expression of IFN regulatory factor family proteins inlymphocytes. J. Immunol. 156, 3711–3720.guyen, H., Hiscott, J., and Pitha, P. M. (1997). The growing familyof interferon regulatory factors. Cytokine Growth Factor Rev. 8,293–312.htaka-Maruyama, C., Hanaoka, F., and Chepelinsky, A. B. (1998).A novel alternative spliced variant of the transcription factor
AP2alpha is expressed in the murine ocular lens. Dev. Biol. 202,125–135.s of reproduction in any form reserved.

P
S
S
S
S
T
T
V
V
W
Y
Y
Y
55Differential Expression of IRFs in the Lens
Piatigorsky, J. (1981). Lens differentiation in vertebrates: A reviewof cellular and molecular features. Differentiation 19, 134–153.
Pine, R., Decker, T., Kessler, D. S., Levy, D. E., and Darnell, J. E., Jr.(1990). Purification and cloning of interferon-stimulated genefactor 2 (ISGF2): ISGF2 (IRF-1) can bind to promoters of bothinterferon- and interferon-stimulated genes but is not a primarytranscriptional activator. Mol. Cell. Biol. 10, 2448–2457.
olitis, A. D., Ozato, K., Coligan, J. E., and Vogel, S. N. (1994).Regulation of IFNg-induced nuclear expression of IFN consensussequence binding protein in murine macrophages. J. Immunol.152, 2270–2278.
Sabath, D. E., Broome, H. E., and Prystowsky, M. B. (1990).Glyceraldehyde-3-phosphate dehydrogenase mRNA is a majorinterleukin 2-induced transcript in a cloned T-helper lympho-cyte. Gene 91, 185–191.
Saunders, N., Dahler, A., Jones, S., Smith, R., and Jetten, A. (1996).Interferon-g as a regulator of squamous differentiation. J. Derma-tol. Sci. 13, 98–106.
aunders, N. A., and Jetter, A. M. (1994). Control of growthregulatory and differentiation-specific genes in human epidermalkeratinocytes by interferon-g. J. Biol. Chem. 269, 2016–2022.
ax, C. M., Dziedzic, D. C., Piatigorsky, J., and Reddan, J. R. (1995).Analysis of a-crystallin expression in cultured mouse and rabbitlens cells. Exp. Eye Res. 61, 125–127.
chindler, C., and Darnell, J. E., Jr. (1995). Transcriptional re-sponses to polypeptide ligands: The JAK-STAT pathway. Annu.Rev. Biochem. 64, 621–651.
chreiber, E., Matthias, P., Muller, M. M., and Schaffner, W. (1989).Rapid detection of octamer binding proteins with mini-extracts,prepared from a small number of cells. Nucleic Acids Res. 17, 6419.
aniguchi, T. (1997). Transcription factors IRF1 and IRF2: Linking
the immune responses and tumor suppression. J. Cell. Physiol.173, 128–130.Copyright © 1999 by Academic Press. All right
aniguchi, T., Harada, H., and Lamphier, M. (1995). Regulation ofthe interferon system and cell growth by the IRF transcriptionfactors. J. Cancer Res. Clin. Oncol. 121, 516–520.aughan, P. S., van Wijnen, A. J., Stein, J. L., and Stein, G. S. (1997).Interferon regulatory factors: Growth control and histone generegulation—It’s not just interferon anymore. J. Mol. Med. 75,348–359.ilcek, J., and Sen, G. C. (1996). Interferons and other cytokines. In“Fields Virology” (B. N. Fields et al., Eds.), pp. 375–398.Lippincott–Raven, Philadelphia.istow, G. J., Shaughnessy, M. P., Lee, D. C., Hodin, J., andZelenka, P. S. (1993). A macrophage migration inhibitory factoris expressed in the differentiating cells of the eye lens. Proc. Natl.Acad. Sci. USA 90, 1772–1775.
amada, T., Nakamura, T., Westphal, H., and Russell, P. (1990).Synthesis of a-crystallin by a cell line derived from the lens of atransgenic animal. Curr. Eye Res. 9, 31–37.
amagata, T., Nishida, J., Tanaka, T., Sakai, R., Mitani, K., Yo-shida, M., Taniguchi, T., Yazaki, Y., and Hirai, H. (1996). A novelinterferon regulatory factor family transcription factor, ICSAT/Pip/LSIRF, that negatively regulates the activity of interferon-regulated genes. Mol. Cell. Biol. 16, 1283–1294.oshida, K., Imaki, J., Koyama, Y., Harada, T., Shinmei, Y., Oishi,C., Matsushima-Hibiya, Y., Matsuda, A., Nishi, S., Matsuda, H.,and Sakai, M. (1997). Differential expression of maf-1 and maf-2genes in the developing rat lens. Invest. Ophthalmol Visual Sci.38, 2679–83
Received for publication June 26, 1998
Revised March 5, 1999Accepted March 5, 1999
s of reproduction in any form reserved.





























