Embed Size (px)
Citation preview

Behavioural Brain Research 107 (2000) 21–33
Research report
Intracranial self-stimulation and sucrose intake differ as hedonicmeasures following chronic mild stress: interstrain and
interindividual differences
Christina Kurre Nielsen *, Jørn Arnt, Connie SanchezPharmacological Research, H. Lundbeck A/S, Ottilia6ej 9, DK-2500 Valby, Copenhagen, Denmark
Received 20 April 1999; received in revised form 22 June 1999; accepted 22 June 1999
Abstract
The present study was designed to assess the utility of sucrose intake and intracranial self-stimulation (ICSS) as hedonicmeasures for chronic mild stress (CMS) induced behavioural deficits. Wistar and PVG hooded rats were exposed to a variety ofmild stressors, e.g. periods of food and/or water deprivation, soiled cage, light/dark reversal, confinement to small cages andpairing, during 6–9 weeks. The intake of 1% sucrose solution was significantly reduced in stressed PVG hooded rats comparedto control animals. The sucrose intake in stressed Wistar rats remained unaltered, indicating that CMS-induced decreases insucrose intake are strain dependent. However, sucrose intake has in our experience been shown to be unreliable as the observeddecreases following CMS were inconsistent over time. ICSS behaviour was evaluated from rate/frequency functions bydetermining the frequency that supported 50% of maximal response rate. Neither the Wistar nor the PVG hooded rats showedan overall decrease in ICSS behaviour following CMS. However, the ICSS measures revealed interindividual differences in bothrat strains. In the stress groups a subgroup (1492.4%) of rats progressively exhibited an attenuated ICSS behaviour. Thesefindings may reflect the interindividual variability observed in humans as stress does not invariably lead to depression. The modelmay in its present form be used to study the pathophysiology of depressive disorders. However, the utility of the CMS model tostudy antidepressant drug actions has to be questioned. Our results show there is a need for rat strains in which there is a greatersensitivity for detecting stress effects. It emphasises the fact that replication of CMS-induced decreases in ICSS behaviour can beas problematic as inducing decreases in sucrose intake. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Chronic mild stress; Reward; Intracranial self-stimulation; Sucrose intake; Rat strain
www.elsevier.com/locate/bbr
1. Introduction
The chronic mild stress (CMS) model of depressionin rats originally described by Willner and co-workers[58], is hypothesised to model anhedonia through adecrease in responsiveness to reward. Anhedonia,defined as a loss of interest or pleasure, is one of thetwo core symptoms required for diagnosis of a majordepressive episode (DSM-IV, 1994). In addition, the
reversal of CMS-induced anhedonia typically requires2–5 weeks of treatment, which mimics the clinical timecourse of antidepressant action [57].
CMS involves sequential exposures to a variety ofmild stressors resulting in hedonic deficits to rats, suchas decrease in preference and/or consumption of palat-able solutions of sucrose or saccharin [34,35,45,50,58],reduction in place preference conditioning [40,45,53]and decrease in intracranial self-stimulation (ICSS) be-haviour [32]. The term mild stressors should be consid-ered as relative to the stressors used in earlier findings,in which rats sequentially exposed to a variety of severestressors failed to increase fluid intake when saccharinor sucrose was added to drinking water [20].
* Corresponding author. Tel.: +45-36-30-13-11; fax: +45-36-30-52-67.
E-mail address: [email protected] (C. Kurre Nielsen)
0166-4328/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 6 -4328 (99 )00110 -2

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–3322
Sucrose or saccharin intake or preference is the hedo-nic measure that has been most widely adopted and ithas been established in several laboratories[4,25,41,42,48,49]. However, recent papers question thevalidity of the model [13,16,17,26] and as reviewed byWillner [54] there are a number of laboratories in whicheffects of CMS, measured as a decrease in consumptionor preference for sweet solutions, are less reliablyobserved.
In the first attempt to adopt the CMS model we usedsucrose intake as hedonic measure, but in several exper-iments we were unable to find consistent changes fol-lowing CMS. The results of two of those experimentsare presented in the first part of the present paper.Realising the limitation of sucrose intake as a validmeasure of sensitivity to reward we chose to use ICSSas an alternative behavioural endpoint. Even thoughsucrose intake/preference is the far most used hedonicmeasure following CMS, ICSS is a direct measure ofreward and is often used in the discussion of thevalidation of the CMS paradigm, and therefore de-serves greater attention.
Since Olds and Milner [37] found electrical stimula-tion in certain brain areas to be rewarding in animalsthe ICSS paradigm has been used in the study ofreward, anhedonia and motivation (for review see [60]).Self-stimulation threshold has proven useful as a validindex of the hedonic–anhedonic state of an animal [23].
Exposure to severe stress such as uncontrollable foot-shock reduces responding for ICSS in various brainregions in rats [19] and in mice [5,61–64]. Adapting theCMS paradigm, Moreau and co-workers [28–33] haveshown a decreased ICSS behaviour following CMSusing the ventral tegmental area as stimulation site.
The first ICSS experiment in the present study, wasdesigned to replicate the findings of Moreau et al. [32].This was subsequently followed by an experiment in-cluding comparison of different rat strains and stressregimes, aiming at assessing whether ICSS is a morereliable hedonic measure than sucrose intake andwhether similar strain differences were obtained for thetwo methods.
Thus, the present study assesses the utility of sucroseintake and ICSS as behavioural measures for CMS-in-duced behavioural deficits applying different stressregimes and rat strains. An overview of the experimentsis shown in Table 1.
2. Materials and methods
2.1. Subjects
Male Wistar rats and male Pieball Virol Glaxo(PVG) hooded rats (Møllegard, Denmark) weighing250–300 and 220–240 g, respectively, at the beginning
Table 1Overview of experiments
Rat strain Stress regimena MeasurebStudy
CMS-1 SucroseWistar1Sucrose2 PVG hooded CMS-1ICSSWistar3 CMS-2
4 Wistar ICSSCMS-1 vs. CMS-24 PVG hooded CMS-2 ICSS
a CMS-1 adapted from Willner et al. [58] and Papp et al. [38];CMS-2 adapted from Moreau et al. [32].
b Sucrose, consumption of 1% sucrose; ICSS, sensitivity to intracra-nial selfstimulation.
of the study, were used. The rats were housed individu-ally or in pairs in Macrolon type III cages and main-tained on a 12 h light/dark cycle (lights on 06:00).Except when required by the test the animals had freeaccess to food and water. Temperature (2291°C),relative humidity (5595%) and air exchanges (16 timesper h) were automatically controlled. Body weight wasmeasured weekly during all experiments.
2.2. Stress regimes
Two different stress regimes were used. The protocolsare summarised in Table 2. CMS-1 was adapted fromthe procedure described by Willner et al. [58] and Pappet al. [38]. Each week consisted of: two periods (8- and19-h) of food deprivation; three periods (6-, 7- and19-h) of water deprivation; 2 periods (7- and 16-h) ofcage tilt (45°); two 17-h periods of paired housing; one17-h period in a soiled cage (200 ml water in 100 g
Table 2Chronic mild stress proceduresa
CMS-1Stress regimen CMS-2(study 1+2+4) (study 3+4)
Access to nutrientsFood deprivation (h) 8+19 17
6+7+19 2×17Water deprivation (h)1Empty water bottle (h)2Restricted food access (h)
Proximal en6ironment7+16Cage tilting (h)
Grouped housing (h) 172×1717 17Soiled cage
8×1Restraint (h)
Distal en6ironmentStrobe (h) 6
12Light on at night (h)48 64Ligh vs. dark reversal (h)
a The Table summarizes the stressors applied during each week ofthe CMS procedure. The duration and number of occasions areshown. For precise description of the stressors see Section 2.

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 23
sawdust bedding); one 6-h period of low intensity stro-boscopic illumination (150 flashes/min); and 48 h oflight/dark reversal.
CMS-2 was adapted from Moreau et al. [32]. Eachweek consisted of: One 17-h period of food and waterdeprivation immediately followed by 2 h of restrictedaccess to food; one 17-h period of water deprivationimmediately followed by 1-h exposure to an emptybottle; one 17-h periods of paired housing in a soiledcage (200 ml water in 100 g sawdust bedding); eight 1-hperiods of confinement to small cages (25×10×10cm); one period of overnight illumination; and 64-h ofreversed light/dark cycle during the weekend.
2.3. Hedonic measures
2.3.1. Sucrose intake (experiments 1 and 2)Rats were acquired twice weekly (Tuesday and Fri-
day) to consume a 1% sucrose solution. The acquisitionconsisted of 8–10 1 h baseline tests in which sucrosesolution was presented, in their home cage, following a20 h period of food and water deprivation. Sucroseintake was measured by weighing the bottles before andafter the test. Following stable baseline levels sucroseconsumption was measured during the CMS protocol,under similar conditions, at weekly intervals (Wednes-day), throughout the experiments. The food and waterdeprivation prior to the sucrose tests is a stressor uponwhich subsequent stressors included in the CMS regimeis imposed, a prior stressor history which is not consid-ered in any subsequent analysis.
2.3.2. Intracranial self-stimulation (experiments 3 and4)
All rats were housed in pairs and allowed 2 weeks toacclimatise before surgery. They were anesthetized with0.2 ml/100 g Hypnorm®/Dormicum® (fentanyl–fluanisone/midazolam) and mounted in a stereotaxicframe (David Kopf) with the incisor bar set 2.7 mmbelow the interaural line. Stainless-steel bipolar elec-trodes (MS 303/1, Plastic Products, Roanoke, VA) wereimplanted unilaterally into the ventral tegmental area(VTA) with the electrode tips separated by 0.5 mm. Thecoordinates used were 3.2 mm anterior to the interauralline, 0.6 mm lateral from the midline and 8.2 mmventral from the skull surface according to Paxinos andWatson [46]. Five stainless-steel screws and dental ce-ment were used to anchor the electrode assembly to theskull.
Two weeks postoperatively each rat was shaped tomake a nose-poke for rewarding intracranial electricalstimulation in a ENV-008CT operant test chamber(MED-Associates) placed in a sound-attenuated box.Poking the nose into a hole (2.5 cm in diameter) locatedin the side wall 1 cm above the floor interrupted aconvergent light beam, thereby initiating brain stimula-
tion delivered through a constant current stimulator(PHM-150B, MED-Associates). Each nose-poke pro-duced a 0.5 s train of monophasic square-wave pulses,each of 0.1 ms in duration. Stimulation was deliveredunder a fixed-interval 1 s schedule of reinforcements inorder to avoid extreme density of stimulation. Electricalstimulation and data recording were controlled througha computer and interface (MED-Associates).
The rats were initially trained at a fixed stimulationfrequency of 70 Hz and an individually selected currentintensity (70–300 mA) in order to maintain the highestpossible response rate without inducing motor impair-ment. They were trained daily in 40 min sessions forapproximately 3 weeks.
Subsequently, rate–frequency curves were establishedby stepwise changing the frequency while maintainingthe current intensity constant. At 2 min intervals thefrequency was decreased and then increased in 0.05–0.2log unit steps in the range 1.1–1.9 log Hz (10–85 Hz).Each session comprised 15 discrete 2-min trials for eachfrequency with 1 priming stimulation at the beginningof each trial. The decreasing series of frequencies wasused as a warm up phase, that is only data obtainedfrom the ascending phase was used in the subsequentcalculations. The increasing series of frequencies wereindividually selected so as to yield an ascending curvewith at least two adjacent points at maximum andminimum response rate, respectively.
This study employed the rate–frequency version ofthe curve-shift method [24] to assess CMS effect onreward. The properties of the rate-frequency function,in particular with regard to separating ICSS rewardeffects on operant motor/performance capacity, havebeen extensively validated and investigated (e.g. Refs.[7,10,18,22,27]). We evaluated ICSS behaviour by deter-mining the frequency that will support 50% of maximalresponse rate, the effective frequency (EF50, nomencla-ture adopted from Kling-Petersen and Svensson [22]).Based on the ‘‘broken-line’’ principle presented byCampbell et al. [7], we described a mathematical modelto estimate EF50. In our hands, this method givesreliable and stable EF50 estimations.
Each test day ascending rate–frequency curves weregenerated for each rat by modelling the observed re-sponse rate (nosepokes(NP)/2 min trial) as a functionof log10 of the imposed frequency (F) (SAS® version6.09). The function consists of three linear segments; ahorizontal lower asymptote through the minimum re-sponse (NPmin), a horizontal upper asymptote throughthe maximum response (NPmax) and a linear transitionbetween the asymptotes (a+b*F). Only data (M) be-tween the last NPmin and first NPmax is included. Thiscan be described by the following equations:
1. Consider M={(F, NP(F))�max{F �NP(F)=NPmin}
5F5min{F �NP(F)=NPmax}}

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–3324
Fig. 1. A representative example of the rate–frequency curve used todetermine EF50 (the frequency that will support 50% of maximalresponse rate). Response rate (nose-pokes/2 min) is plotted againstthe frequency expressed in log units. A lateral curve-shift is inter-preted to reflect changes in reward effectiveness of the stimulation.
Except when required by the stress protocol the animalswere individually housed. According to baseline sucroseintakes the animals were allocated to two matchedgroups. One group of rats was subjected to CMS-1 fora period of 9 weeks, while the other group was main-tained under standard laboratory conditions.
2.4.3. Experiment 3: the effects on ICSS in Wistar ratsexposed to the CMS-2 regime
Wistar rats (N=27) were allocated to one group,housed individually and submitted to 10 weeks ofCMS-2 and another group (control group), pairedhoused under standard laboratory conditions. TheirEF50 were determined twice weekly and comparisonswith the mean of the two last baseline tests wereperformed for each individual rat.
2.4.4. Experiment 4: the effects of rat strain and stressregime on ICSS beha6iour
Wistar rats (N=33) were allocated to three matchedgroups; a control group and two groups subjected to 9weeks of CMS-1 and CMS-2, respectively. PVG rats(N=21) were allocated to two matched groups; onegroup was submitted to 9 weeks of CMS-2 and theother group (control group) maintained under standardlaboratory conditions. The control groups were housedin pairs. Except where required by the stress protocolthe rats in the stress groups were singly housed. TheirEF50 were determined twice weekly and comparisonswith the mean of the two last baseline tests wereperformed for each individual rat.
2.5. Histology
After completion of ICSS testing, the animals werekilled and the brains removed and stored at −80°C.Sections of 12 mm were cut on a cryostat before beingstained with cresyl violet. Electrode tip placement wasdetermined on the frontal planes of the Paxinos andWatson stereotaxic atlas [46].
2.6. Statistical analysis
In the majority of comparisons between the controlgroup and stress group either normality or variancehomogeneity tests failed, both for sucrose intake andICSS data. Therefore, it was decided to evaluate alldata by non-parametric statistics. Two way analysis ofvariance (ANOVA) were performed on ranks, withdays and treatment (stress) as factors and with animalnested in treatment which implies that a set of parame-ters corresponds to only one observation. All observa-tions are, therefore, independent. In the ICSSexperiments there appeared to be a sub-classification ofthe stress group. If the total mean response of a stressedanimal exceeded the maximum mean response in the
2. Fit f(F)=min(max(a+b*F, NPmin), NPmax) to thedata in M by the least squares procedure. In case ofambiguity, the function with the smallest b is used.
The effective frequency (EF50), defined as the fre-quency that will support 50% of the maximal responserate was determined. A lateral curve-shift along the axisof simulation frequencies, i.e. a change in EF50, willreflect changes in reward effectiveness of the stimula-tion. A representative rate–frequency curve for oneanimal is shown in Fig. 1.
Rate–frequency testing were performed twice weekly(Tuesday and Friday). A stable response was achievedwhen maximum response showed no increasing or de-creasing trends and when EF50 varied by less than10–12%. The animals were subsequently allocated totwo matched groups.
2.4. Design of experiments
2.4.1. Experiment 1: the effects on sucrose intake inWistar rats exposed to the CMS-1 regime
Wistar rats (N=78) were brought into the labora-tory 4 weeks before the experiment was started. Exceptwhen required by the stress protocol the animals wereindividually housed. According to baseline sucrose in-takes the animals were allocated to two matchedgroups. One group of rats was subjected to CMS-1 fora period of 6 weeks, while the other group was main-tained under standard laboratory conditions.
2.4.2. Experiment 2: the effects on sucrose intake inPVG hooded rats exposed to the CMS-1 regime
PVG hooded rats (N=20) were brought into thelaboratory 3 weeks before the experiment was started.
C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 25
control group it was classified as belonging to a sub-group. Prior to comparisons between stress (sub- andmain group) and control group the response of thestress subgroup and the main group were compared bytwo way ANOVA on ranks, with days and group asfactors and with animal nested in groups. Significantmain effects (PB0.05) on treatment were followed byan unpaired t-test for each day (SAS® version 6.09).
For clarity it was decided to present both sucroseintake data and ICSS data as mean9SEM, still per-forming non-parametric analysis.
Body weight data, that passed normality and vari-ance homogeneity tests were analysed by one or twoway ANOVA. Multiple comparisons were performedby Tukey’s test. Otherwise, Kruskal–Wallis ANOVAon ranks were used (SigmaStat® version 2.0).
3. Results
3.1. Experiment 1: sucrose intake
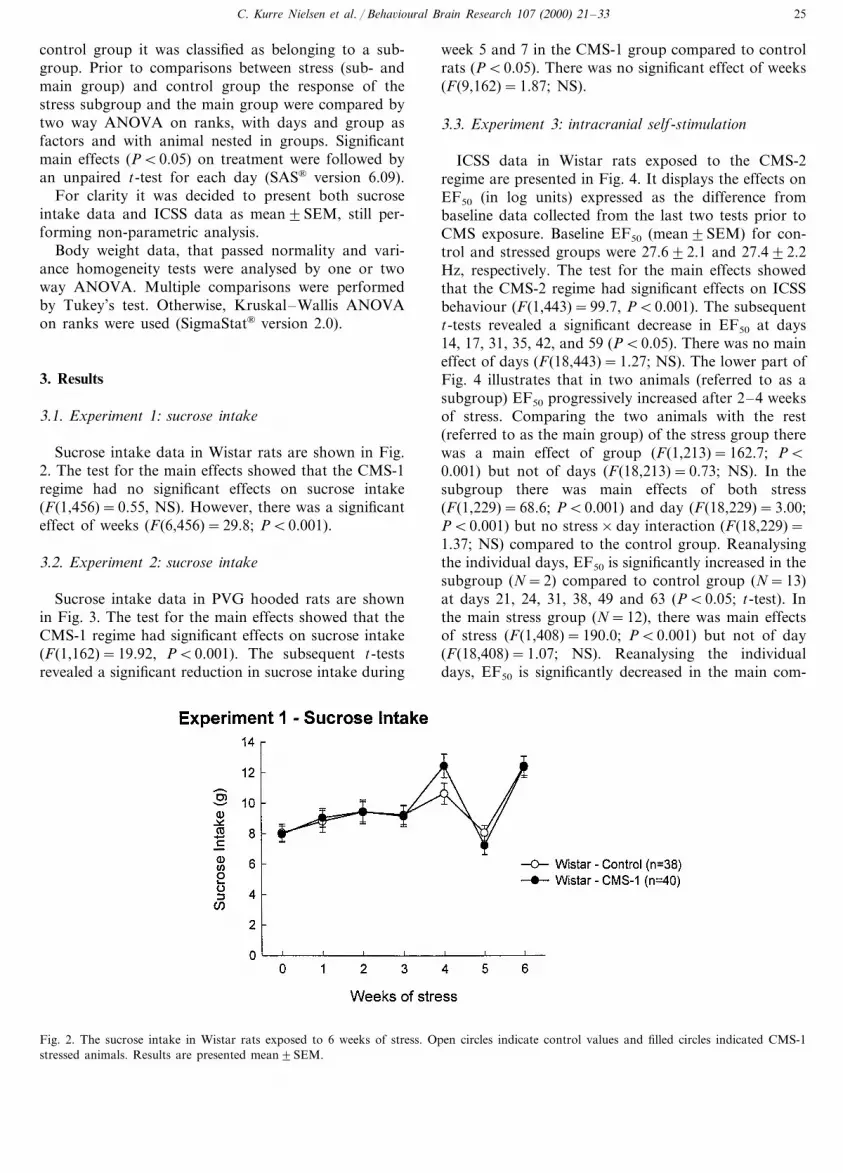
Sucrose intake data in Wistar rats are shown in Fig.2. The test for the main effects showed that the CMS-1regime had no significant effects on sucrose intake(F(1,456)=0.55, NS). However, there was a significanteffect of weeks (F(6,456)=29.8; PB0.001).
3.2. Experiment 2: sucrose intake
Sucrose intake data in PVG hooded rats are shownin Fig. 3. The test for the main effects showed that theCMS-1 regime had significant effects on sucrose intake(F(1,162)=19.92, PB0.001). The subsequent t-testsrevealed a significant reduction in sucrose intake during
week 5 and 7 in the CMS-1 group compared to controlrats (PB0.05). There was no significant effect of weeks(F(9,162)=1.87; NS).
3.3. Experiment 3: intracranial self-stimulation
ICSS data in Wistar rats exposed to the CMS-2regime are presented in Fig. 4. It displays the effects onEF50 (in log units) expressed as the difference frombaseline data collected from the last two tests prior toCMS exposure. Baseline EF50 (mean9SEM) for con-trol and stressed groups were 27.692.1 and 27.492.2Hz, respectively. The test for the main effects showedthat the CMS-2 regime had significant effects on ICSSbehaviour (F(1,443)=99.7, PB0.001). The subsequentt-tests revealed a significant decrease in EF50 at days14, 17, 31, 35, 42, and 59 (PB0.05). There was no maineffect of days (F(18,443)=1.27; NS). The lower part ofFig. 4 illustrates that in two animals (referred to as asubgroup) EF50 progressively increased after 2–4 weeksof stress. Comparing the two animals with the rest(referred to as the main group) of the stress group therewas a main effect of group (F(1,213)=162.7; PB0.001) but not of days (F(18,213)=0.73; NS). In thesubgroup there was main effects of both stress(F(1,229)=68.6; PB0.001) and day (F(18,229)=3.00;PB0.001) but no stress×day interaction (F(18,229)=1.37; NS) compared to the control group. Reanalysingthe individual days, EF50 is significantly increased in thesubgroup (N=2) compared to control group (N=13)at days 21, 24, 31, 38, 49 and 63 (PB0.05; t-test). Inthe main stress group (N=12), there was main effectsof stress (F(1,408)=190.0; PB0.001) but not of day(F(18,408)=1.07; NS). Reanalysing the individualdays, EF50 is significantly decreased in the main com-
Fig. 2. The sucrose intake in Wistar rats exposed to 6 weeks of stress. Open circles indicate control values and filled circles indicated CMS-1stressed animals. Results are presented mean9SEM.

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–3326
Fig. 3. The sucrose intake in PVG hooded rats exposed to 9 weeks of stress. Open circles indicate control values and filled circles indicated CMS-1stressed animals. Results are presented mean9SEM. * PB0.05 (see Section 3 for details).
to control group (F(1,303)=7.12; PB0.01) but not ofday (F(18,303)=0.81; NS). Though, reanalysing theindividual days by t-test, revealed no significant differ-ences between the main compared to control group.
In PVG hooded rats, baseline EF50 (mean9SEM)for control and stressed groups were 30.891.5 and31.991.8 Hz, respectively. The test for the main effectsshowed that the CMS-2 regime had significant effectson ICSS behaviour (F(1,296)=90.16, PB0.001) whilethere was no main effect of days (F(18,296)=1.49;NS). The subsequent t-tests revealed a significant de-crease in EF50 at days 3–10, 17, 24, 28, 35, 38, 45, 52and 59 (PB0.05). The lower part of Fig. 5 illustratesthat one animal fell into the subgroup. This rat differedsignificantly from the rest (referred to as the maingroup) of the stress group (F(1,131)=26.67; PB0.001)and from the control group (F(1,158)=11.31; P=0.001). Unfortunately, this rat had to be omitted after 4weeks, as its metal dust-cap got stuck in the cage lidcausing loosening of the electrode assembly. In themain stress group, there was main effects of stresscompared to control group (F(1,289)=144.0; PB0.001) and of days (F(18,289)=1.91; PB0.05) but nostress×day interaction (F(18,289)=0.91; NS). Re-analysing the individual days, EF50 is significantly de-creased in the main compared to control group at days3–10, 17–28, 35, 38, 45, 52 and 59 (PB0.05; t-test).
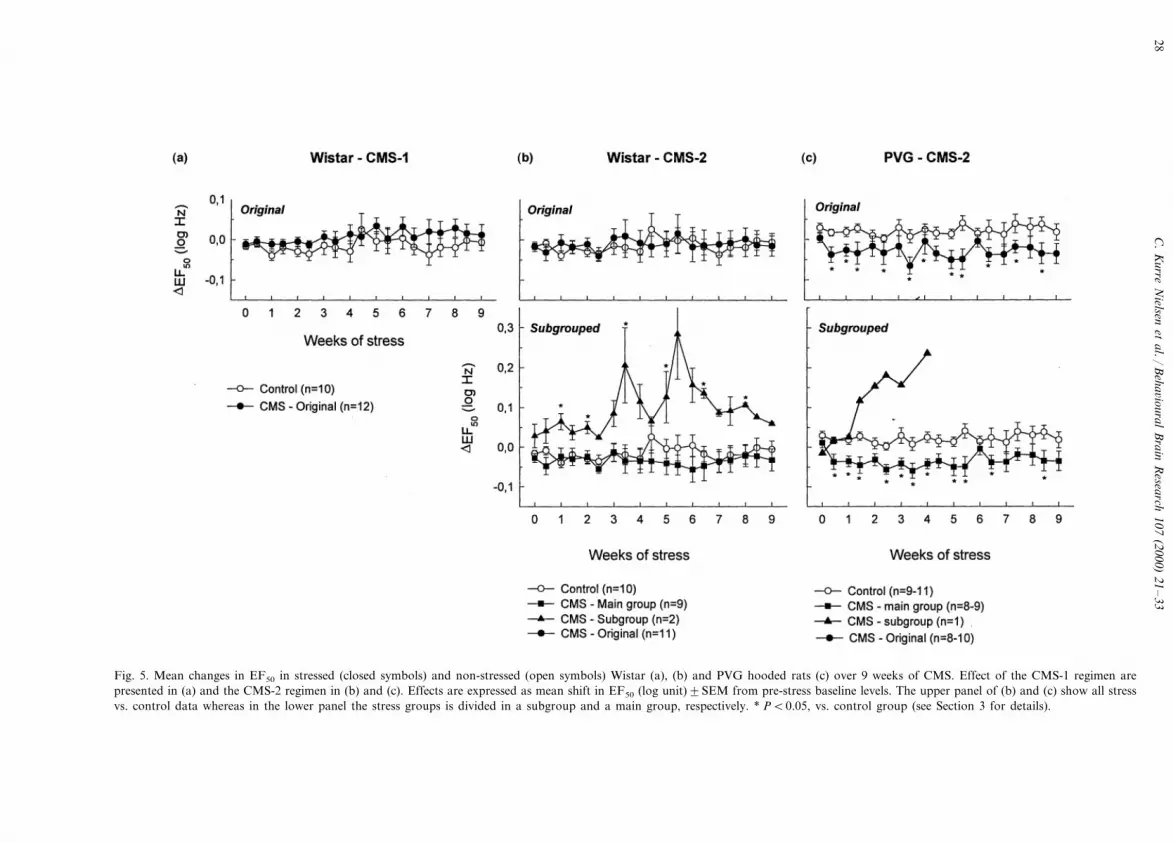
Histologic analysis indicated that the electrode tipswere located in the VTA (Fig. 6.). There did not appearto be any differences between the electrode locations ofcontrol and stress animals. Moreover, the apparentsubgroups in the Wistar rats following CMS did notappear to be outliers with regard to electrodeplacements.
pared to control group at days 10–17, 24, 31–45 and52–59 (PB0.05; t-test).
3.4. Experiment 4: intracranial self-stimulation
ICSS data in Wistar rats exposed to the CMS-1 andCMS-2 regime plus data in PVG hooded rats exposedto the CMS-2 regime are presented in Fig. 5. It displaysthe effects on EF50 (in log units) expressed as thedifference from baseline data collected from the lasttwo tests prior to CMS exposure.
In the Wistar rats baseline EF50 (mean9SEM) forcontrol, CMS-1 and CMS-2 groups were 31.991.9,32.491.3 and 30.991.4 Hz, respectively. The test forthe main effects showed that the CMS-1 regime hadsignificant effects on ICSS behaviour (F(1,353)=28.46,PB0.001) and no effect of days (F(18,353)=1.17; NS).Though, analysing the individual days by t-test, therewere no significant differences between the control andCMS-1 group. With respect to the CMS-2 regime therewas no significant effect of stress (F(1,338)=1.93; NS)or days (F(18,338)=0.82; NS). However, in the CMS-2group two animals (referred to as the subgroup) dif-fered significantly from the rest (referred to as the maingroup) of the stress groups (F(1,159)=185.2; PB0.001) with no main effect of days (F(18,159)=0.73;NS). In the subgroup there were main effects of stress(F(1,178)=133.9; PB0.001) but not of days(F(18,178)=0.65; NS) compared to the control group.Reanalysing the individual days, EF50 is significantlyincreased in the subgroup compared to control group atdays 7, 14, 24, 35, 45, 56 (PB0.05; t-test). In the mainstress group, there were main effects of stress compared

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 27
3.5. Experiment 1–4: body weight
Changes in body weight from baseline to week 5 werecalculated for each experiment and illustrated in Fig. 7.Due to significant differences in baseline body weightbetween studies for Wistar rats (F(2,135)=231; PB0.001) and PVG hooded rats (PB0.001; Kruskal–Wal-lis ANOVA on ranks) as well as between rat strains(PB0.001; Kruskal–Wallis ANOVA on ranks) eachstudy is separately analysed.
3.5.1. Experiments 1–3CMS-1 significantly reduced weight gain in Wistar
rats (F(1,76)=9.65; PB0.01) and PVG hooded rats(F(1,18)=13.6; PB0.01). CMS-2 significantly reducedweight gain in Wistar rats (F(1,25)=38.5; PB0.001).
3.5.2. Experiment 4Stress had a significant effect on body weight gain
(F(2,30)=41.8; PB0.001). Multiple comparisons usingTukey’s test revealed significant reduced body weightgain in CMS-1 and CMS-2 groups compared to controlgroup (PB0.01) whereas the effect of CMS-1 andCMS-2 did not differ. The PVG hooded rats hadsimilarly reduced body weight gain following CMS-2exposure (F(1,16)=21.7; PB0.001).
4. Discussion
4.1. Chronic mild stress
Stressors induce behavioural disturbances and neuro-chemical changes in animals, some of which have asimilarity to those observed in depressive disorders [2].Based on the procedure by Katz [20] in which rats areexposed sequentially to a variety of severe stressorsWillner et al. [58] developed the chronic mild stress(CMS) procedure. The intentions were to provide amore realistic analogue of the strains encountered ineveryday life, by reducing the severity of the stressors[58]. The presentation of a variety of stressors appearedto be essential, as repeated presentation of single stres-sors resulted in rapid habituation to stress-inducedbehavioural changes [35].
In the studies using sucrose intake as hedonic mea-sure, the CMS-1 regime was adapted from Willner et al.[58] and Papp et al. [38]. However, as we changed to theICSS paradigm, we adopted the CMS procedure de-scribed by Moreau et al. [32], i.e. CMS-2. The stressregimes both consist of a variety of mild stressors, butdiffer in some of the stressors used. A review of theliterature, comparing CMS procedures between labora-tories does not make it possible to extract necessary
Fig. 4. Mean changes in EF50 in stressed (closed symbols) and non-stressed (open symbols) Wistar rats over 9 weeks of CMS-2. Effects areexpressed as mean shift in EF50 (log unit)9SEM from pre-stress baseline levels. The upper panel shows all stress vs. control data whereas in thelower panel the stress group is divided in a subgroup and a main group, respectively. * PB0.05 vs. control group (see Section 3 for details).
C.
Kurre
Nielsen
etal./
Beha6ioural
Brain
Research
107(2000)
21–
3328
Fig. 5. Mean changes in EF50 in stressed (closed symbols) and non-stressed (open symbols) Wistar (a), (b) and PVG hooded rats (c) over 9 weeks of CMS. Effect of the CMS-1 regimen arepresented in (a) and the CMS-2 regimen in (b) and (c). Effects are expressed as mean shift in EF50 (log unit)9SEM from pre-stress baseline levels. The upper panel of (b) and (c) show all stressvs. control data whereas in the lower panel the stress groups is divided in a subgroup and a main group, respectively. * PB0.05, vs. control group (see Section 3 for details).

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 29
Fig. 6. Location of the electrode tips (filled circle) for each animal in experiment 3 and 4. The placement of seven rats could not be verified.Coronal sections are redrawn from Paxinos and Watson atlas [46]. The numbers in the top indicate the distance from the interaural line. Thearrows identify the subgroups of animals decreasing their ICSS behaviour following CMS exposure.
and sufficient features of the CMS procedure [54]. Thecriticisms of the CMS model have concentrated on theinadequacy of the sucrose consumption method to con-sistently quantify stress-induced anhedonia, rather thanon the CMS procedure in itself. The question whethersucrose intake and ICSS may differ in their sensitivityto CMS has received limited attention. To address thisissue we included both stress regimes in the last experi-ment. Firstly, we were able to compare the sucroseintake procedure with ICSS as hedonic measures. Sec-ondly, it was possible to compare the influence of twowidely used stress regimes on ICSS behaviour, in thesame laboratory. We found that the PVG hooded rats,
which showed a decrease in sucrose intake did not showan impaired ICSS behaviour following CMS-1, indicat-ing that the rewarding effect of ICSS is not influencedin the same manner as sucrose intake. Overall, the twostress regimes (CMS-1 and CMS-2) affected the ICSSbehaviour of the Wistar rats in much the same manner,suggesting that differences in elements applied in theCMS procedures may be of minor importance whencomparing results between laboratories. However, al-though the influence of the CMS-1 regime on ICSSbehaviour only was tested once in the Wistar rats, itwas only in the CMS-2 group there appeared a sub-group which showed an impaired ICSS response, indi-

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–3330
cating that ICSS behaviour may be more susceptible tothe CMS-2 than to the CMS-1 regime.
Due to different test conditions, the experimentsusing ICSS and sucrose intake, respectively, differedwith respect to the housing conditions prior to theCMS exposure. In the sucrose experiments the rats weresingle housed. In the ICSS experiments, on the con-trary, the test conditions allowed paired housing. Thereare studies reporting altered circadian rhythms of thehypothalamic catecholaminergic system and hormonesas a consequence of individual housing [14]. The ICSScontrol groups we left paired to minimize stress,whereas the stress groups were individually housedimmediately after baseline determinations. Moreover,individual housing may contribute as a stressor, stillallowing the grouping as part of the stress regimes. Inthe present study the stress effect in the subgroups ofWistar rats developed 2–3 weeks following CMS-2.Moreau and co-workers [28–33] obtain stress effect5–9 days after onset of the similar CMS regime. Itindicates that individual housing prior to CMS expo-sure may influence the onset of the stress effect but isunlikely to be necessary to obtain the stress effect.However, it cannot be ruled out that the observeddifference in onset of stress effect simply is due todifferences between the Wistar rat strain used in thetwo laboratories as the rats are obtained from differentsuppliers.
4.2. Sucrose intake as a hedonic measure
Exposure to the CMS-1 regime did not alter thesucrose intake in Wistar rats whereas the PVG hoodedrats showed an overall reduction in their sucrose intakecompared to non-stressed control groups. This indicatesthat a decrease in sucrose intake following CMS isdependent upon the rat strain used. Sensitivity to stresshas been reported to vary between strains of rats [49]and mice [15,64]. However, the rat strains used in thepresent study have both shown CMS-induced decreasesin sucrose consumption in other laboratories [43,56].
The methodological issues in the sucrose intake testhas been thoroughly discussed and evaluated by Will-ner [54]. As the main purpose of the present study is tocontribute with data using ICSS as hedonic measurefollowing CMS, the reliability of sucrose measurementswill only be discussed in brief.
It has been suggested that reduction in body weightgain following CMS contribute to a decrease in sucroseintake [26]. Comparison of data from a number oflaboratories, including our own, showed that decreasedhedonic responsiveness following CMS could not besolely attributed to loss of body weight [56]. In fact, thepresent study confirms that body weight is not corre-lated to sucrose intake, as in experiment 1, body weightis decreased while sucrose intake remains unaffected.
The apparent problems in detecting an impaired con-sumption of sweet solutions following CMS in a num-ber of laboratories have lead to various alterations inthe test procedure. A prolonged test duration has beenreported to increase the sensitivity of the test [4]whereas others suggest testing during the dark phase ofthe light/dark cycle [8]. CMS-induced decreases in su-crose/saccharin intake are in some laboratories foundto be accompanied by decreases in preference for su-crose/saccharin solutions [4,8,58]. Others have founddecreases in sucrose intake while sucrose preferenceremained unaltered [13,26]. Decreases in sucrose prefer-ence may even be detected in the absence of a decreasein sucrose intake [9].
It cannot be ruled out that by changing the testprocedure we would have obtained a greater sensitivityto CMS. However, as seen in the present study usingthe PVG hooded rats, decreases in sucrose intake fol-lowing CMS was not consistent from week to week,indicating that sucrose intake is not a reliable measure.Our main concerns were the variability within as well asbetween experiment (unpublished observations), leadingus to ICSS as an alternative hedonic measure.
4.3. ICSS as a hedonic measure
Neither the Wistar nor the PVG hooded rats showedan overall decrease in ICSS behaviour following CMS.These findings do not support the work by Moreau and
Fig. 7. Mean body weight changes9SEM for Wistar (W) and PVGhooded (P) rats following 5 weeks of CMS and the baseline weights(upper right) for experiments 1–4. ** PB0.01, *** PB0.001 vs.control group (ANOVA).

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 31
co-workers [28–33]. Due to reproducibility, the ICSStechnique contrary to sucrose measure, offered thechance to look at individual differences. In all thegroups of CMS-2 stressed rats, a subgroup (N=1–2,1492.4%) of rats progressively exhibited an attenuatedICSS behaviour as indicated by an elevation of EF50
value. This was not the case for the CMS-1 stressedWistar rats.
It could be argued that our method lack sensitivityfor detecting lateral curve-shifts. However, prior to thestudy we showed that haloperidol dose-dependentlyincreased the EF50 value (MED=0.027 mmol/kg (0.01mg/kg), unpublished data). This agrees with the litera-ture data using ICSS from either VTA [52] or MFB(medial forebrain bundle; [21]).
In the present study, ICSS behaviour is establishedby placing electrodes in the VTA, the origin of themesolimbic dopamine system [36]. The mesolimbic pro-jections from VTA are a major source of the braindopaminergic innervation and play a major role inreward processes and/or motivation [11,47,59,60]. ICSSestablished from VTA is known to increase extracellu-lar concentrations of dopamine (DA) and its metabo-lites in the nucleus accumbens [12]. Exposure to CMShas shown to increase DA levels in the nucleus accum-bens [51,55]. This is suggested to lead to a decrease insensitivity of postsynaptic DA D2 receptors within thelimbic forebrain, which is believed to be responsible forCMS-induced anhedonia. [39,44].
Exposure to severe stress such as uncontrollable foot-shock is shown to reduce responding for ICSS in vari-ous brain regions in rats [19] and in mice [5,61–64].Studies by Moreau and co-workers show that chronicexposure to unpredictable mild stressors reduce re-sponding for ICSS from VTA in rats [32]. However,studies in mice have indicated that the VTA is notuniformly affected by stressors [63]. ICSS from thedorsal VTA was reduced following uncontrollable foot-shock while ICSS from the ventral VTA was unaffectedby the stressor. In the present study the subgroup ofrats in which CMS-2 caused an increase in EF50 value,there was no apparent difference in their electrodeplacements compared to the rest of the stress group.Regarding the CMS-1 exposed Wistar rats, even thoughelectrode placements varied within the VTA, no in-crease in EF50 was seen in the stress group. In spite ofthe relatively small number of rats, it therefore seemsunlikely that regional variations in electrode placementscould account for the failure to detect decreased ICSSbehaviour following CMS.
As the hypothesis was that CMS induced a decreasedsensitivity to reward, measured either as a decrease insucrose intake or ICSS behaviour, it was unexpected tofind that CMS-2 increased ICSS behaviour in the Wis-tar rats (experiment 3) and PVG hooded rats (experi-ment 4). As the finding was not consistent between
experiments 3 and 4 (Wistar rats) and as we are dealingwith less than a 13% decrease in EF50, the findings mustbe interpreted with caution. It may be speculated as anon-specific effect on the rewarding effect of ICSS,caused by an conditioned behaviour in an environment(test chamber) the animals associate with reward andnot with stress, as in the sucrose intake test which isperformed in the home cage. Others have observed anincreased sensitivity to reward following stress expo-sure. Zacharko et al. [64] found an increased ICSSbehaviour in one mouse strain following exposure toinescapable footshock. Likewise Willner et al. [55]found an initial increase in sucrose intake following 1week of CMS, where the sucrose intake decreasedafterwards. However, according to the hypothesis weexpected that a prolonged exposure to CMS (up to 9weeks) eventually would lead to a motivational impair-ment (for review see Ref. [6]).
4.4. Interstrain and interindi6idual differences
The reason for including the PVG hooded rats in theICSS experiments was the apparent greater vulnerabil-ity to the rewarding effect of sucrose compared toWistar rats. Moreover, the PVG hooded rat is aninbred strain compared to the widely used outbredWistar rat. However, both strains showed similar ICSSbehaviour following exposure to the CMS-2 regime.That is, strain differences in CMS-induced decrease insucrose intake cannot predict a similar decrease in ICSSbehaviour. Thus, instead of considering one strain to beparticularly vulnerable to CMS, it may be more appro-priate to conclude that the effects of CMS in differentstrains of rats vary across the different tasks in whichthe rats are tested.
Taking the apparent sub-classification of the ICSSbehaviour in Wistar and PVG rats exposed to theCMS-2 regime into consideration, the present studyconfirms the need for considering interindividual differ-ences in behavioural response to stress [2]. In humans,stress does not invariably lead to depression and not allcases of depression are a consequence of stressful expe-riences [3]. In an epidemiological study life-event lossesand perceived strain were positively correlated to de-pressive symptomatology [1]. Depending on the level ofstress the prevalence of depression ranged from 15 to 28and 5 to 40% related to loss events and perceived strain,respectively. Thus, our results are consistent with theclinical situation.
4.5. Summary
The present study suggests that ICSS and sucroseintake are differentially affected by CMS, as the twomeasures do not correspond within the same strain ofrat. The rat strain which appeared vulnerable to the

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–3332
rewarding effect of sucrose did not show a decrease inICSS behaviour following CMS.
Sucrose intake as a hedonic measure has in ourhands proved to be unreliable as decreases followingCMS are inconsistent over time. Moreover, CMS-in-duced decrease in sucrose intake has shown to be straindependent.
The ICSS technique appeared to be reliable, butoverall it was not possible to decrease ICSS behaviourfollowing CMS. The ICSS measures revealed interindi-vidual differences in both rat strains. However, thisappeared to be dependent upon the CMS regime.
Given that the theoretical rationale for the CMSmodel is that the procedure simulates anhedonia, ourresults may suggest that CMS-induced anhedonia is atrait rather than a state phenomenon. The model doesindeed mimic the interindividual variability observed inhumans as stress does not invariably lead to depression.Thus the model may in its present form be used tostudy the pathophysiology of depressive disorders.However, the utility of the CMS model to study antide-pressant drug actions has to be questioned.
ICSS has only been used as a hedonic measure in afew CMS studies and the present studies contribute toincrease knowledge about how ICSS behaviour is af-fected by different stress regimes and rat strains. Al-though ICSS may be suitable as a hedonic measure, ourresults show there is a need for rat strains in whichthere is a greater sensitivity for detecting stress effects.It emphasises the fact that replication of CMS-induceddecreases in ICSS behaviour can be as problematic asinducing decreases in sucrose intake.
Acknowledgements
We thank Søren Hansen, Annette Jøns, MariannePetersen, Jesper Rasmussen and Lotte Nielsen for theirexcellent technical assistance in conducting these exper-iments, and Carsten Rasmussen for his valuable helpwith the statistical analysis.
References
[1] Aneshensel CS, Stone JD. Stress and depression: a test ofbuffering model of social support. Arch Gen Psychiatry1982;39:1392–6.
[2] Anisman H, Zacharko RM. Multiple neurochemical and behav-ioral consequences of stressors: implications for depression.Pharmacol Ther 1990;46:119–36.
[3] Anisman H, Zacharko RM. Depression as a consequence ofinadequate neurochemical adaptation in response to stressors. BrJ Psychiatry Suppl 1992;36–43.
[4] Ayensu WK, Pucilowski O, Mason GA, Overstreet DH, RezvaniAH, Janowsky DS. Effects of chronic mild stress on serumcomplement activity, saccharin preference, and corticosteronelevels in Flinders lines of rats. Physiol Behav 1995;57:165–9.
[5] Bowers WJ, Zacharko RM, Anisman H. Evaluation of stressoreffects on intracranial self-stimulation from the nucleus accum-bens and the substantia nigra in a current intensity paradigm.Behav Brain Res 1987;23:85–93.
[6] Cabib S, Puglisi-Allegra S. Stress, depression and the mesolimbicdopamine system. Psychopharmacology (Berl) 1996;128:331–42.
[7] Campbell KA, Evans G, Gallistel CR. A microcomputer-basedmethod for physiologically interpretable measurement of therewarding efficacy of brain stimulation. Physiol Behav1985;35:395–403.
[8] D’Aquila PS, Newton J, Willner P. Diurnal variation in theeffect of chronic mild stress on sucrose intake and preference.Physiol Behav 1997;62:421–6.
[9] De Vry J, Schreiber R. The chronic mild stress depression model:future developments from a drug discovery perspective. Psy-chopharmacology (Berl) 1997;134:349–50.
[10] Edmonds DE, Stellar JR, Gallistel CR. Parametric analysis ofbrain stimulation reward in the rat: II. Temporal summation inthe reward system. J Comp Physiol Psychol 1974;87:860–9.
[11] Fibiger HC, LePiane FG, Jakubovic A, Phillips AG. The role ofdopamine in intracranial self-stimulation of the ventral tegmen-tal area. J Neurosci 1987;7:3888–96.
[12] Fiorino DF, Coury A, Fibiger HC, Phillips AG. Electricalstimulation of reward sites in the ventral tegmental area in-creases dopamine transmission in the nucleus accumbens of therat. Behav Brain Res 1993;55:131–41.
[13] Forbes NF, Stewart CA, Matthews K, Reid IC. Chronic mildstress and sucrose consumption: validity as a model of depres-sion. Physiol Behav 1996;60:1481–4.
[14] Gambardella P, Greco AM, Sticchi R, Bellotti R, Di Renzo G.Individual housing modulates daily rhythms of hypothalamiccatecholaminergic system and circulating hormones in adult malerats. Chronobiol Int 1994;11:213–21.
[15] Griffiths J, Shanks N, Anisman H. Strain-specific alterations inconsumption of a palatable diet following repeated stressorexposure. Pharmacol Biochem Behav 1992;42:219–27.
[16] Harris RB, Zhou J, Youngblood BD, Smagin GN, Ryan DH.Failure to change exploration or saccharin preference in ratsexposed to chronic mild stress. Physiol Behav 1997;63:91–100.
[17] Hatcher JP, Bell DJ, Reed TJ, Hagan JJ. Chronic mild stress-in-duced reductions in saccharin intake depend upon feeding status.J Psychopharmacol 1997;11:331–8.
[18] Hunt GE, Atrens DM. Parametric manipulations and fixed-in-terval self-stimulation. Physiol Behav 1992;51:1009–20.
[19] Kamata K, Yoshida S, Kameyama T. Antagonism of footshockstress-induced inhibition of intracranial self-stimulation bynaloxone or methamphetamine. Brain Res 1986;371:197–200.
[20] Katz RJ. Animal model of depression: pharmacological sensitiv-ity of a hedonic deficit. Pharmacol Biochem Behav 1982;16:965–8.
[21] Kling-Petersen T, Ljung E, Svensson K. The preferential do-pamine autoreceptor antagonist (+ )-UH232 antagonizes thepositive reinforcing effects of cocaine and d-amphetamine in theICSS paradigm. Pharmacol Biochem Behav 1994;49:345–51.
[22] Kling-Petersen T, Svensson K. A simple computer-based methodfor performing and analyzing intracranial self-stimulation exper-iments in rats. J Neurosci Methods 1993;47:215–25.
[23] Markou A, Koob GF. Postcocaine anhedonia. An animal modelof cocaine withdrawal. Neuropsychopharmacology 1991;4:17–26.
[24] Markou A, Koob GF. Intracranial self-stimulation thresholds asa measure of reward. In: Sahgal A, editor. Behavioural Neuro-science: a Practical Approach. Oxford: IRL Press, 1993:93–115.
[25] Marona-Lewicka D, Nichols DE. The effect of selective sero-tonin releasing agents in the chronic mild stress model of depres-sion in rats. Stress 1997;2:91–100.

C. Kurre Nielsen et al. / Beha6ioural Brain Research 107 (2000) 21–33 33
[26] Matthews K, Forbes N, Reid IC. Sucrose consumption as anhedonic measure following chronic unpredictable mild stress.Physiol Behav 1995;57:241–8.
[27] Miliaressis E, Rompre PP, Laviolette P, Philippe L, CoulombeD. The curve-shift paradigm in self-stimulation. Physiol Behav1986;37:85–91.
[28] Moreau JL, Borgulya J, Jenck F, Martin JR. Tolcapone: apotential new antidepressant detected in a novel animal model ofdepression. Behav Pharmacol 1994;5:344–50.
[29] Moreau JL, Bourson A, Jenck F, Martin JR, Mortas P. Curativeeffects of the atypical antidepressant mianserin in the chronicmild stress-induced anhedonia model of depression. J PsychiatryNeurosci 1994;19:51–6.
[30] Moreau JL, Bos B, Jenck F, Martin JR, Mortas P, Wichmann J.5-HT2C receptor agonists exhibit antidepressant-like propertiesin the anhedonia model of depression in rats. Eur Neuropsy-chopharmacol 1996;6:169–75.
[31] Moreau JL, Jenck F, Martin JR, Mortas P, Haefely W. Effectsof moclobemide, a new generation reversible Mao-A inhibitor, ina novel animal model of depression. Pharmacopsychiatry1993;26:30–3.
[32] Moreau JL, Jenck F, Martin JR, Mortas P, Haefely WE. An-tidepressant treatment prevents chronic unpredictable mildstress-induced anhedonia as assessed by ventral tegmentum self-stimulation behavior in rats. Eur Neuropsychopharmacol1992;2:43–9.
[33] Moreau JL, Scherschlicht R, Jenck F, Martin JR. Chronic mildstress-induced anhedonia model of depression: sleep abnormali-ties and curative effects of electroshock treatment. Behav Phar-macol 1995;6:682–7.
[34] Muscat R, Papp M, Willner P. Reversal of stress-induced an-hedonia by the atypical antidepressants, fluoxetine and maproti-line. Psychopharmacology (Berl) 1992;109:433–8.
[35] Muscat R, Willner P. Suppression of sucrose drinking by chronicmild unpredictable stress: a methodological analysis. NeurosciBiobehav Rev 1992;16:507–17.
[36] Oades RD, Halliday GM. Ventral tegmental (A10) system: neu-robiology. 1. Anatomy and connectivity. Brain Res1987;434:117–65.
[37] Olds J, Milner P. Positive reinforcement produced by electricalself-stimulation of septal area and other regions of rat brain. JComp Physiol Psychol 1954;47:419–27.
[38] Papp M, Klimek V, Willner P. Effects of imipramine on sero-tonergic and beta-adrenergic receptor binding in a realistic ani-mal model of depression. Psychopharmacology (Berl)1994;114:309–14.
[39] Papp M, Klimek V, Willner P. Parallel changes in dopamine D2receptor binding in limbic forebrain associated with chronic mildstress-induced anhedonia and its reversal by imipramine. Psy-chopharmacology (Berl) 1994;115:441–6.
[40] Papp M, Lappas S, Muscat R, Willner P. Attenuation of placepreference conditioning but not place aversion conditioning bychronic mild stress. J Psychopharmacol 1992;6:352–6.
[41] Papp M, Moryl E. Antidepressant activity of non-competitiveand competitive NMDA receptor antagonists in a chronic mildstress model of depression. Eur J Pharmacol 1994;263:1–7.
[42] Papp M, Moryl E. Antidepressant-like effects of 1-aminocyclo-propanecarboxylic acid and D-cycloserine in an animal model ofdepression. Eur J Pharmacol 1996;316:145–51.
[43] Papp M, Moryl E, Willner P. Pharmacological validation of thechronic mild stress model of depression. Eur J Pharmacol1996;296:129–36.
[44] Papp M, Muscat R, Willner P. Subsensitivity to rewarding andlocomotor stimulant effects of a dopamine agonist following
chronic mild stress. Psychopharmacology (Berl) 1993;110:152–8.[45] Papp M, Willner P, Muscat R. An animal model of anhedonia:
attenuation of sucrose consumption and place preference condi-tioning by chronic unpredictable mild stress. Psychopharmacol-ogy (Berl) 1991;104:255–9.
[46] Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordi-nates. New York: Academic Press, 1986.
[47] Phillips AG. Brain reward circuitry: a case for separate systems.Brain Res Bull 1984;12:195–201.
[48] Przegalinski E, Moryl E, Papp M. The effect of 5-HT1A recep-tor ligands in a chronic mild stress model of depression. Neu-ropharmacology 1995;34:1305–10.
[49] Pucilowski O, Overstreet DH, Rezvani AH, Janowsky DS.Chronic mild stress-induced anhedonia: greater effect in a ge-netic rat model of depression. Physiol Behav 1993;54:1215–20.
[50] Sampson D, Willner P, Muscat R. Reversal of antidepressantaction by dopamine antagonists in an animal model of depres-sion. Psychopharmacology (Berl) 1991;104:491–5.
[51] Stamford JA, Muscat R, O’Connor JJ, et al. Voltammetricevidence that subsensitivity to reward following chronic mildstress is associated with increased release of mesolimbic do-pamine. Psychopharmacology (Berl) 1991;105:275–82.
[52] Tsibulsky VL, Grocki S, Dashevsky BA, et al. Mixed D2/5-HT2A antagonism of cocaine-induced facilitation of brain stimu-lation reward. Pharmacol Biochem Behav 1998;59:275–80.
[53] Valverde O, Smadja C, Roques BP, Maldonado R. The attenua-tion of morphine-conditioned place preference following chronicmild stress is reversed by a CCKB receptor antagonist. Psy-chopharmacology (Berl) 1997;131:79–85.
[54] Willner P. Validity, reliability and utility of the chronic mildstress model of depression: a 10-year review and evaluation.Psychopharmacology (Berl) 1997;134:319–29.
[55] Willner P, Golembiowska K, Klimek V, Muscat R. Changes inthe mesolimbic dopamine may explain stress-induced anhedonia.Psychobiology 1991;19:79–84.
[56] Willner P, Moreau JL, Nielsen CK, Papp M, Sluzewska A.Decreased hedonic responsiveness following chronic mild stressis not secondary to loss of body weight. Physiol Behav1996;60:129–34.
[57] Willner P, Muscat R, Papp M. Chronic mild stress-inducedanhedonia: a realistic animal model of depression. NeurosciBiobehav Rev 1992;16:525–34.
[58] Willner P, Towell A, Sampson D, Sophokleous S, Muscat R.Reduction of sucrose preference by chronic unpredictable mildstress, and its restoration by a tricyclic antidepressant. Psy-chopharmacology (Berl) 1987;93:358–64.
[59] Wise RA. Catecholamine theories of reward: a critical review.Brain Res 1978;152:215–47.
[60] Wise RA. Neuroleptics and operant behavior: the anhedoniahypothesis. Behav Brain Sci 1982;5:39–87.
[61] Zacharko RM, Bowers WJ, Kelley MS, Anisman H. Preventionof stressor-induced disturbances of self-stimulation bydesmethylimipramine. Brain Res 1984;321:175–9.
[62] Zacharko RM, Bowers WJ, Kokkinidis L, Anisman H. Region-specific reductions of intracranial self-stimulation after uncon-trollable stress: possible effects on reward processes. Behav BrainRes 1983;9:129–41.
[63] Zacharko RM, Kasian M, MacNeil G, Anisman H. Stressor-in-duced behavioral alterations in intracranial self- stimulationfrom the ventral tegmental area: evidence for regional variations.Brain Res Bull 1990;25:617–21.
[64] Zacharko RM, Lalonde GT, Kasian M, Anisman H. Strain-spe-cific effects of inescapable shock on intracranial self-stimulationfrom the nucleus accumbens. Brain Res 1987;426:164–8.
.





























