Embed Size (px)
Citation preview

OPHELIA, 7: 101-112 (December 1969)
IRRIGATION IN THE SIPUNCULID
PHASCOLION STROll1BI (MONT.)
J. HYLLEBERG KRISTENSEN
Kristineberg zoologiska Station, Fiskebackskil, SwedenPresent address: Marine Biological Laboratory, DK-3000 Hdsing0r, Denmark
ABSTRACT
The species irrigates its habitation by muscular means. The frequency of irrigation waves, thevolume of water transported per wave and the capacity of irrigation per gram per hour are calculated from kymograph recordings. Two types of irrigation (continuous and discontinuous) aredistinguished. The total irrigation is estimated at 15 ml/g/h at temperatures between 15.0 and22.2°C.
Generation of over- and under-pressures in P. strombi connected with a vertical capillary isdescribed. The experiments demonstrate that an eventual resistance yielded by the sedimentmay be overcome by the sipunculid.
A decapitated specimen was studied for thirteen days during which period the irrigationwas continuous.
The significance of the irrigation is discussed. The irrigtation was found mainly to serve arespiratory purpose.
INTRODUCTION
Little work has been done on irrigation in sipunculids in contrast to the polychaetes. Observations on Phascolion strombi were published by Perez (1925) andHampson (1964), but the authors only stated that the sipunculid irrigates itshabitation.
The present investigation describes the mechanism of irrigation and its capacity, as these are of ecological interest. Several animals are known to cohabit withP.strombi inside its shelter. Descriptions are given by Brumpt (1898), Perez (1924,1925), Knudsen (1944), Hampson (1964), and Nielsen (1964). Own observationson the fauna will be presented in near future.
P.strombi blocks, as described by Perez (1925, p.75), the aperture of inhabitedshells or tubes with sediment cemented together with secreted mucus. A mainchannel through the cement mass is left for protrusion and retraction of the introvert. In empty gastropod shells used by the sipunculid, a secondary channelmay be found near the columella, or in the siphonal channel in shells providedwith this structure. Irrigation of the shelter varies according to presence or absence of a secondary aperture as pointed out by Perez (1925, p.76).

102 J. HYLLEHERG KRISTENSEN
The present work was carried out at the Kristineberg Zoological Station, Sweden, and I havereceived financial support from the Nordic Council for Marine Biology. I express my bestthanks for tnis grant. Furthermore, I give my sincere thanks to the head of the station dr.B. Swedmark for eminent working conditions at the Kristineberg laboratory.
MATERIAL AND METHODS
Phascolion strombi in different types of shelters were dredged in Gasa Riinna in thevicinity of the Kristineberg Station. The bottom of the sampling locality consists ofsilty clay and clay inhabited by an Amphiura community. The depth is 25 to 35 m.
Animals were placed in glass tubes and allowed to establish themselves from fiveto thirty days. Such animals proved especially valuable as the movements insidethe shelter could be studied. In addition animals naturally sheltered in Dentaliumor Turritella shells were used. Turritella shells were used with the apex cut off, andshells without secondary apertures in the clay plug were selected, as a secondaryaperture makes them unusable due to the technique applied.
The apex end, or its analogue in glass tubes, was connected by a piece ofrubber tube with a 75 cm capillary bent to a narrow S-form for practical reasons(Fig. 1). The capillary was filled with sea water, and while immersed horizontallyin a shallow vessel an air bubble was placed in the capillary by a syringe. Thesipunculid was then connected with the capillary. The air bubble served as anindex of water movements. These movements were followed manually with a twoarmed indicator on a level writer. The trace was recorded on a smoked kymograph drum. A friction from 0.2 to 4 mm water column (Poiseuille's law) hasto be overcome within the measured range of irrigation speeds in the horizontalcapillary. The friction is insignificant compared with the force of Phascolion (seepp.l07-109). The loss of kinetic energy caused by the rubber connection mentionedabove is negligible.
Experiments were also made in a system where the components are placedvertically. The capillary was replaced by a 100 cm L-shaped capillary which, aftermounting of P.strombi, was put into a 30 cm high jar with sea water.
The capillary surface served as index of water movements produced by theanimal. However, in the vertical set-up, physical forces playa role. (Capillaryforce, friction and hydrostatic pressure influence the movement of the water inthe capillary).
Experiments in the horizontal system were carried out with stagnant water,while the longer lasting experiments in the vertical system were performed withrunning sea-water. Salinities ranged between 31 and 34 %0'
The vertical distance recorded on the smoked paper was a measure of thepumping activity of P.strombi, 1 cm equalling 0.01 ml sea water. (The volume ofthe bore was estimated by measuring the length of the water column after thefilling of the capillary with a known quantity of sea-water).

IRRIGATION IN PHASCOLION STROMBI to3
FIG. I. Horizontal set up used for manual registration of irrigation in Phascolion strombi
Over-flow technique was used in two experiments.The conclusions on irrigation in P. strombi are based on 20 recordings (355
min.) in the horizontal system and 70 recordings (3500 min.) in the verticalsystem.
RESULTS
The mechanism of irrigation
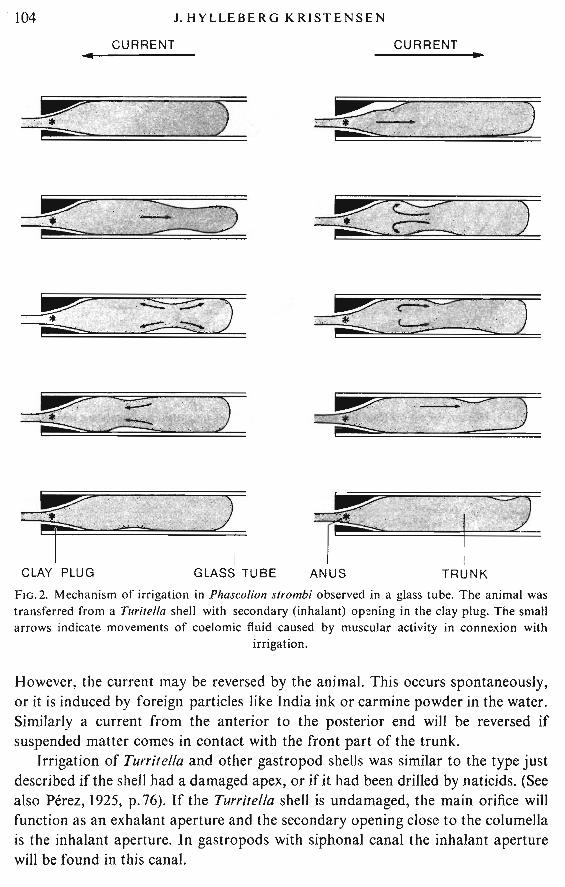
Examination of animals in glass tubes demonstrated that P. strombi belongs tothe type of worms in which the irrigation is produced by muscular means (Fig. 2).Piston-like swellings similar to peristaltic and antiperistaltic waves of the intestinetravel along the trunk.
The analogy with peristalsis of the gut is especially obvious when water istransported from the posterior end of the trunk and forwards. When water istransported in the opposite direction it is usually possible to distinguish a depression on the right side and one on the left side; the latter is the first to reach theposterior end of the trunk (Fig.2). The result is in both directions a transport of afinite volume of water, and no qualitative difference is seen on kymograph recordigs.
Animals in tubes open at both ends principally move the water over the bodyfrom the posterior end and forwards. The aperture for protrusion of the introvertfunctions as exhalant aperture while the apex opening is the inhalant aperture.
104 J. HYLLEBERG KRISTENSEN
CURRENT CURRENTII
CLAY PLUG GLASS TUBE ANUSI
TRUNKFIG.2. Mechanism of irrigation in Phaseo/ion strombi observed in a glass tube. The animal wastransferred from a Tllritella shell with secondary (inhalant) opening in the clay plug. The smallarrows indicate movements of coelomic fluid caused by muscular activity inconnexion with
irrigation.
However, the current may be reversed by the animal. This occurs spontaneously,or it is induced by foreign particles like India ink or carmine powder in the water.Similarly a current from the anterior to the posterior end will be reversed ifsuspended matter comes in contact with the front part of the trunk.
Irrigation of Turritella and other gastropod shells was similar to the type justdescribed if the shell had a damaged apex, or if it had been drilled by naticids. (Seealso Perez, 1925, p. 76). If the Turri/ella shell is undamaged, the main orifice willfunction as an exhalant aperture and the secondary opening close to the columellais the inhalant aperture. In gastropods with siphonal canal the inhalant aperturewill be found in this canal.
IRRIGATION IN PHASCOLION STROMBI
ml
105
0.15
zo~ 0.10Cl~II:
0.05
introvertinduced.activity
I
/?\, ~
~'-------'---,Jdiscontinuous
[
irrigation0.01 I0.00 -' ! ! ! ! , !, !
1 2 3 4 5 10
absence ofirrigation
! , , , , , 1
15
continuousirrigation
, ': " , ! , ! I "20 25 min
TIME
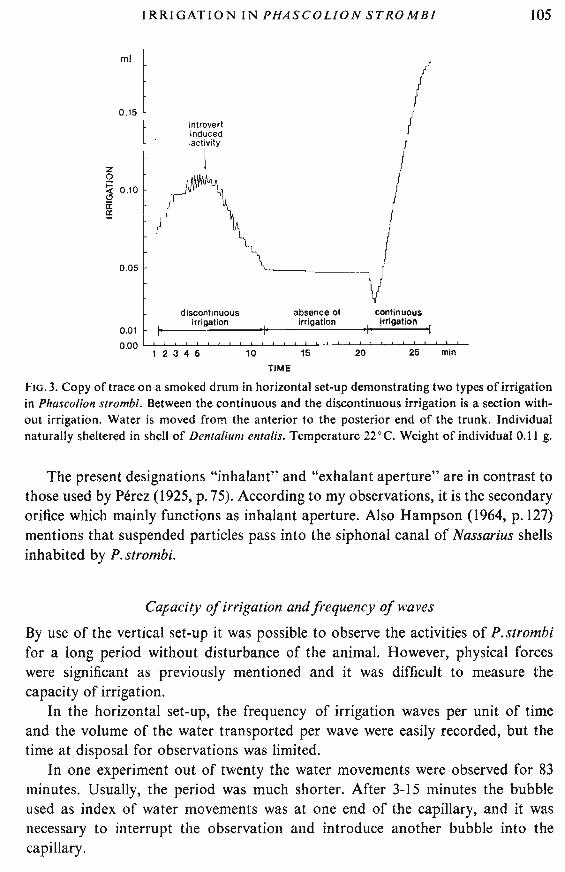
FIG. 3. Copy of trace on a smoked drum in horizontal set-up demonstrating two types of irrigationin Phascolion strombi. Between the continuous and the discontinuous irrigation is a section without irrigation. Water is moved from the anterior to the posterior end of the trunk. Individualnaturally sheltered in shell of Dentalium entalis. Temperature 22°C. Weight of individual 0.11 g.
The present designations "inhalant" and "exhalant aperture" are in contrast tothose used by Perez (1925, p. 75). According to my observations, it is the secondaryorifice which mainly functions as inhalant aperture. Also Hampson (1964, p.127)mentions that suspended particles pass into the siphonal canal of Nassarius shellsinhabited by P. strombi.
Capacity of irrigation andfrequency of waves
By use of the vertical set-up it was possible to observe the activities of P.strombifor a long period without disturbance of the animal. However, physical forceswere significant as previously mentioned and it was difficult to measure thecapacity of irrigation.
In the horizontal set-up, the frequency of irrigation waves per unit of timeand the volume of the water transported per wave were easily recorded, but thetime at disposal for observations was limited.
In one experiment out of twenty the water movements were observed for 83minutes. Usually, the period was much shorter. After 3-15 minutes the bubbleused as index of water movements was at one end of the capillary, and it wasnecessary to interrupt the observation and introduce another bubble into thecapillary.

106 J. HYLLEBERG KRISTENSEN
An extract of a record from the horizontal set-up is shown in Fig. 3. Twotypes of irrigation are distinguished in the example. In the discontinuous irrigation,movements of the introvert will move water in the habitation. Protrusions willinduce water to be drawn into the shelter while retractions causes ejection of water.These introvert induced movements are supplemented by peristaltic waves ofirrigation. In the continuous irrigation water is drawn rhythmically through thehabitation. The current was always in one direction during an observed periodof continuous irrigation, but recordings were obtained, where forward irrigationwas succeeded by discontinuous irrigation, and this again by continuous irrigationin the opposite direction.
The introvert is often, though not always, withdrawn during continuous irrigation.
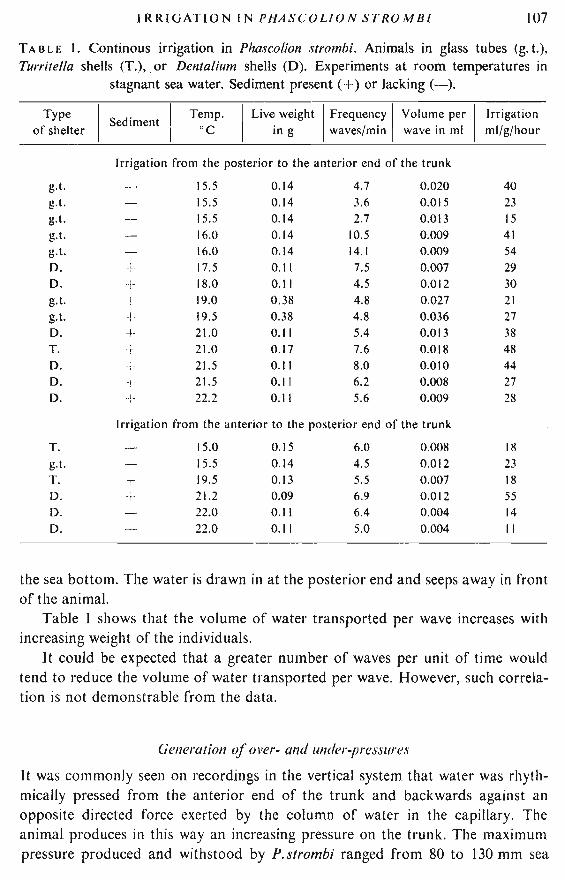
The results of measurements of continuous irrigation on the smoked paper aresummarized in Table1. The irrigation ranges from 15 to 54 ml/g/h when water ismoved forward from the posterior end. In the opposite direction irrigation rangesfrom lito 55 ml/g/h. The experimental temperatures were between 15.0 and 22.2 () C.An effect of temperature on the rhythm and on the capacity of irrigation is notevident from the present recordings. It was noted that irrigation varied significantly in the same individual at the same temperature, even when the recordings weremade with an interval of a few minutes.
Irrigation calculated at temperatures from 15.0 to I9.0°C was 29.4 ml/g/h, andbetween 19.1 and 22.2°C, it was 31.0 ml/g/h, both directions taken into consideration.
Continuous irrigation has been observed for 153 minutes in twenty experiments. Discontinuous irrigation or an absence of irrigation has been observedfor 203 minutes in the same experiments. In view of this finding, the totalirrigation must be reduced to about half when calculated from the continuousirrigation. A value of 15 ml/g/h at 15.0-22.2°C is thus considered representativefor irrigation of Dentalium-type shelters for longer periods. Similar figures, 14.1and 14.6 ml/g/h, were found in two experiments by use of over-flow techniqueduring periods of 90 and 250 minutes, respectively (16°C).
Table I shows that only in two experiments were frequencies of peristalticwaves found to be higher than 10 waves per minute. The highest frequency wasrecorded in an individual which also showed the lowest frequency (2.7 waves perminute). Maximum and minimum values were recorded at about the sametemperature. In the other experiments, frequencies were found to range from 4.5to 8.0 waves per minute at tempeartures between 15.0 and 22.2°C.
A quantitative difference is evident when irrigation in the two directions iscompared (Table 1). The frequency of peristaltic waves and the resulting irrigationwas greater from the posterior end and forward compared with the opposite direction. An exception was the smallest individual weighing 0.09 g. The forwarddirection represents the inhalant current in the natural position of the shelter on
I RRIGATION IN PHASCOLION STROMBI 107
TABLE J. Continous irrigation in Phascolion strombi. Animals in glass tubes <g. t.),Turritella shells (T.),. or Dentalium shells (D). Experiments at room temperatures in
stagnant sea water. Sediment present (+) or Jacking (-).
Type
ISediment I Temp. I Liv~ weight IF;equen~y I Volu~e per I Irrigation
of shelter I cc In g Iwaves/mm wave 10 ml ml/g/hour
Irrigation from the posterior to the anterior end of the trunk
g.t. 15.5 0.14 4.7 0.020 40g.t. 15.5 0.14 3.6 0.015 23g.t. 15.5 0.14 2.7 0.013 15g.t. 16.0 0.14 10.5 0.009 41g.t. 16.0 0.14 14.1 0.009 54D. + 17.5 0.11 7.5 0.007 29D. -I- 18.0 0.11 4.5 0.012 30g.t. + 19.0 0.38 4.8 0.027 21g.t. -I 19.5 0.38 4.8 0.036 27D. -+- 21.0 0.11 5.4 0.013 38T. -+ 21.0 0.17 7.6 0.018 48D. -+- 21.5 0.11 8.0 0.010 44D. + 21.5 0.11 6.2 0.008 27D. -l- 22.2 0.11 5.6 0.009 28
Irrigation from the anterior to the posterior end of the trunk
T. 15.0 0.15 6.0 0.008 18g.t. 15.5 0.14 4.5 0.012 23T. T 19.5 0.13 5.5 0.007 18D. -r" 21.2 0.09 6.9 0.012 55D. 22.0 0.11 6.4 0.004 14D. 22.0 0.11 5.0 0.004 II
the sea bottom. The water is drawn in at the posterior end and seeps away in frontof the animal.
Table I shows that the volume of water transported per wave increases withincreasing weight of the individuals.
It could be expected that a greater number of waves per unit of time wouldtend to reduce the volume of water transported per wave. However, such correlation is not demonstrable from the data.
Generation of over- and under-pressures
It was commonly seen on recordings in the vertical system that water was rhythmically pressed from the anterior end of the trunk and backwards against anopposite directed force exerted by the column of water in the capillary. Theanimal produces in this wayan increasing pressure on the trunk. The maximumpressure produced and withstood by P. strombi ranged from 80 to 130 mm sea
lOS J. HYLLEBERG KRISTENSEN
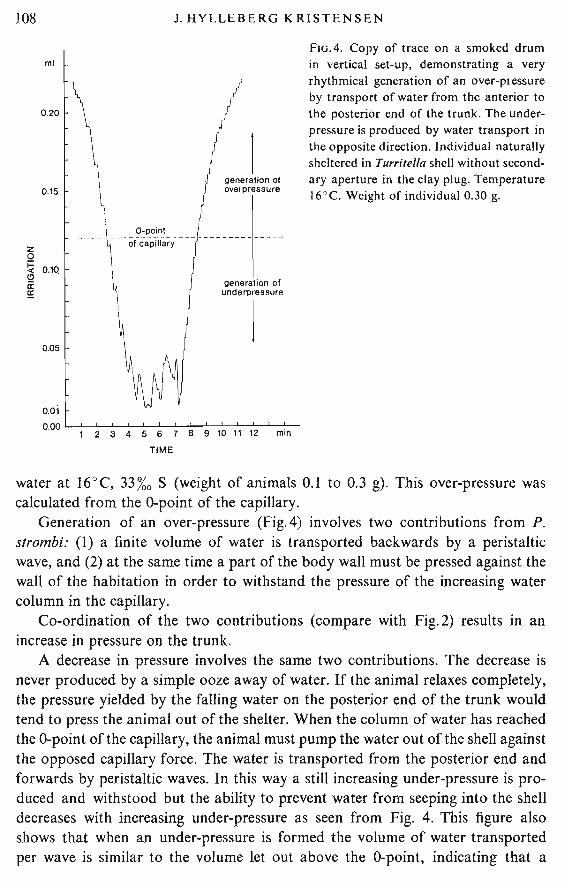
FIG.4. Copy of trace on a smoked drumin vertical set-up, demonstrating a veryrhythmical generation of an over-ptessureby transport of water from the anterior tothe posterior end of the trunk. The underpressure is produced by water transport inthe opposite direction. Individual naturallysheltered in Turritel/a shell without secondary aperture in the clay plug. Temperature16°C. Weight of individual 0.30 g.
generation ofoverpressure
generation ofunderpressure
O-point---ofcapD~ri-----------------
zo~ O.lQCloc!E
0.15
0.20
ml
0.05
0.01
0.00 l--.L.-.L.-L.--~L.......JL.-Jc.......:c.......:~~:-:':;-.l.---L-1 2 3 4 5 6 7 8 9 10 11 12 min
TIME
water at 16°C, 33%0 S (weight of animals 0.1 to 0.3 g). This over-pressure wascalculated from the O-point of the capillary.
Generation of an over-pressure (Fig.4) involves two contributions from P.strombi: (I) a finite volume of water is transported backwards by a peristalticwave, and (2) at the same time a part of the body wall must be pressed against thewall of the habitation in order to withstand the pressure of the increasing watercolumn in the capillary.
Co-ordination of the two contributions (compare with Fig.2) results in anincrease in pressure on the trunk.
A decrease in pressure involves the same two contributions. The decrease isnever produced by a simple ooze away of water. If the animal relaxes completely,the pressure yielded by the falling water on the posterior end of the trunk wouldtend to press the animal out of the shelter. When the column of water has reachedthe O-point of the capillary, the animal must pump the water out of the shell againstthe opposed capillary force. The water is transported from the posterior end andforwards by peristaltic waves. In this way a still increasing under-pressure is produced and withstood but the ability to prevent water from seeping into the shelldecreases with increasing under-pressure as seen from Fig. 4. This figure alsoshows that when an under-pressure is formed the volume of water transportedper wave is similar to the volume let out above the O-point, indicating that a

IRRIGATION IN PHASCOLION STROMBI 109
decrease in pressure is not generated by simple seeping, but involves transport ofafinite volume per wave. The time used to let out the water and produce an underpressure is similar to the time used to let in the water and generate an over-pressure.
Generation of over- and under-pressures in P. strombi must be seen in relationto circumstances in nature. When the shelter is dragged through the sediment bythe animal, the inhalant orifice may be completely buried in the bottom material.Irrigation under these conditions will call for a pressure which will overcomeresitance against movement of water.
A comparison of the traces of rise and fall in Fig.4 shows a difference intransport of water from the front to the rear and vice versa. The animal used inthis experiment lived in a Turritella shell with a perforated apex. Before theexperiment started, it was contr:olled that the holes in the apex were used inirrigation under natural conditions. The falling part of the trace reflects the inhalant current, and the rise reflects the situation when irrigation has been reversedas previously described.
Continuous irrigation (i. e. rise and fall of the recording), discontinuous irrigation and an absence of irrigation were observed both in the vertical and in thehorizontal set-up.
Extirpation of the introvert
In order to investigate the influence of the introvert on the irrigation, an animal(0.15 g) in a Dentalium shell was decapitated and observed for thirteen days in thevertical system.
The protruded introvert was seized in the middle region with a pair of tweezersand the tip of the introvert cut off. The remaining part of the introvert was evidently too damaged to allow the normal regeneration from the wound described bySchleip (1934). Three days later the introvert was autotomized along a predestinedline on the transition between trunk and introvert.
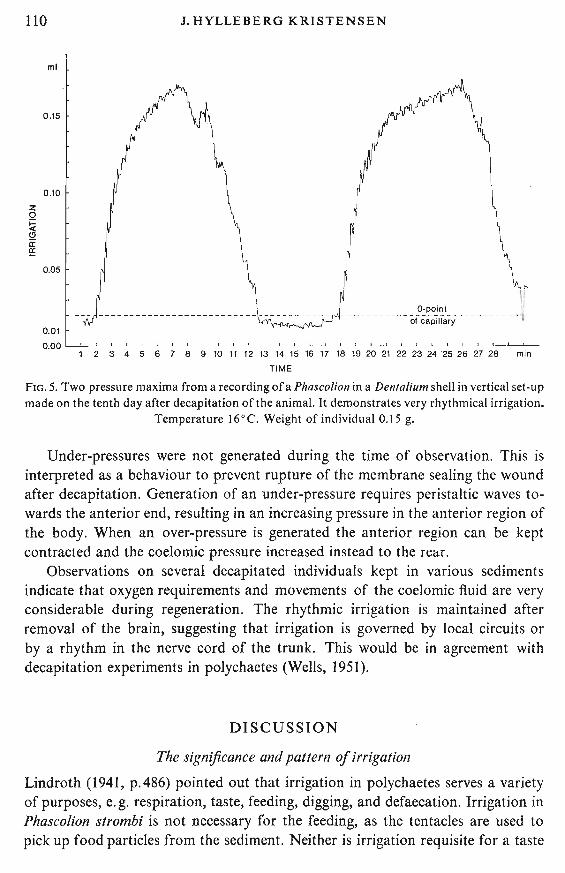
In the decapitated animal an unusually high pressure maximum was sustainedon the first day after decapitation. It ranged between 200 and 250 mm sea-water(160C, 33 %0 S). No regular rhythm was found in the irrigation, but water movements were never interrupted. The frequency ofperistaltic waves was about 180 perhour and did not change significantly during the observed period of the regeneration. Rhythmical pressure maxima were produced three to four times every hourfrom the eighth day after decapitation (Fig. 5). Between two pressure maxima wasa horizontal part, usually with rise and fall. This rhythm was maintained for thenext five days. Then the experiment was terminated.
Fig.5 shows that when an over-pressure is produced it is difficult for the decapitated animal to prevent water from oozing away at the end of a peristalticwave. At a low over-pressure the loss is about 20 % and at the maximum it isabout 50 %. This loss together with the decreasing volume transported per waveresults in an S-form of the rise of the curve.
110
ml
J. HYLLEBERG KRISTENSEN
0.15
0.10
0.05
0.01
0.00 L-..J'_'__L____!.__'_----'-____'_---'-----'-----'-----'--'--'---'--'---'--""---'---'---'--"----~'___''_'__L____!.__'__.l.____'__
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 '25 26 27 28 - min
TIME
FIG. 5. Two pressure maxima from a recording of a Phascolion in a Denfalium shell in vertical set-upmade on the tenth day after decapitation of the animal. It demonstrates very rhythmical irrigation.
Temperature 16°C. Weight of individual 0.15 g.
Under-pressures were not generated during the time of observation. This isinterpreted as a behaviour to prevent rupture of the membrane sealing the woundafter decapitation. Generation of an under-pressure requires peristaltic waves towards the anterior end, resulting in an increasing pressure in the anterior region ofthe body. When an over-pressure is generated the anterior region can be keptcontracted and the coelomic pressure increased instead to the rear.
Observations on several decapitated individuals kept in various sedimentsindicate that oxygen requirements and movements of the coelomic fluid are veryconsiderable during regeneration. The rhythmic irrigation is maintained afterremoval of the brain, suggesting that irrigation is governed by local circuits orby a rhythm in the nerve cord of the trunk. This would be in agreement withdecapitation experiments in polychactes (Wells, 1951).
DISCUSSION
The significance and pattern ofirrigation
Lindroth (1941, p.486) pointed out that irrigation in polychaetes serves a varietyof purposes, e. g. respiration, taste, feeding, digging, and defaecation. Irrigation inPhascolion strombi is not necessary for the feeding, as the tentacles are used topick up food particles from the sediment. Neither is irrigation requisite for a taste

IRRIGATION IN PHASCOLION STROMBl III
function. According to Akesson (1958, p. 179), the nuchal organ at the dorsal base ofthe tentacles is most probably a chemoreceptory organ. The introvert is used indigging as well as dragging of the shelter, but these functions do not depend onirrigation. On the other hand, feeding, digging and dragging always cause displacement of water in the habitation, resulting in what I have termed discontinuousirrigation, which evidently has a certain respiratory significance. However, measure.ments of oxygen concentrations of the inhalant and exhalant currents areneeded to estimate the exact relations of the two types of irrigation to respiration.The significance of irrigation for defaecation and removal of excretory products isunknown.
A relation of irrigation to respiration is indicated by nitrogen bubbling experiments (vertical set-up), as the pattern of irrigation is altered when the oxygen tension is lowered. On one occasion bubbling was continued for six hours. During thefirst hour of bubbling irrigation was increased and continuous. Next, irrigationgradually decreased and became intermittent. Pauses with no other activity thanoccasional protrusions of the introvert alternated with short-lasting bursts ofirrigation at the end of three hours. This intermittent irrigation was continued forthe rest of the period.
Wells (1950, p.138) described irrigation in the polychaete Arenicola marinawhich resembles the intermittent type observed in P.strombi during oxygendeficiency. He found that intermittent irrigation with short bursts of irrigation isuseful, as Arenicola marina during tidal exposure may detect when favourableconditions return. A testing function of the shortlasting bursts is possbile in P.strombi, too, but it seems more likely that the bursts of irrigation are related toremoval of waste products from the shelter. Exploration of the surroundings canmore easily be made by occasional protrusions of the introvert.
An irrigation in P. strombi of 15 ml per g per hour at 15 to 22 0 C looks rathersmall. Unfortunately, it is impossible from the present knowledge to estimate theamount of oxygen taken up through the trunk, introvert and tentacles, respectively.In addition the oxygen consumption is unknown. The oxygen capacity of thecoelomic fluid is estimated at 0.008 ml oxygen per g, by use of the data fromSipunculus nudus (Florkin, 1933, p.705). Horkin finds that the oxygen content ofthe coelomic fluid is 1.6 %by volume of the coelomic fluid, which makes 50 %ofthe total animal volume. The oxygen content of 15 ml aereated deep sea-water at15 0 C is about 0.087 ml oxygen. The oxygen store of the coelomic fluid couldtheoretically be renewed every five minutes if all oxygen is taken up from the irrigated water, but this is undoubtedly not the case. Polychaetes take up 50 to 60 %of the oxygen in the irrigated water (Dales, 1960, p. 81).
Most probably there is only oxygen enough in the irrigated water for a localsupply of the body wall. It must be taken into consideration that besides P. strombi,the micro flora and fauna of the clay plug, about 30 entoprocts and 10 other smallcohabiting animals in the shelter, take up their oxygen from the water.

112 J. HYLLEBERG KRISTENSEN
In large sipunculid species of some genera (e. g. Sipunculus), coelomic canalsof the integument make the contact between oxygen and coelomic fluid easier.Such canals are not found in P.strombi. The oxygen must pass through therelatively thick body wall to come in contact with the erythrocytes, the numberof which is low. Franzen & Hinge (1962) estimated the cell volume in sipunculidsto a little higher than 3 %of the coelomic fluid. Florkin (1933) mentioned 7 %forSipunculus nudus.
A steep oxygen dissociation curve (Florkin, 1932, p.833), and an absence of aBohr effect (Florkin, 1932), together with a low concentration of erythrocytes anda minimal vascularization, indicate that sipunculids in general have small demandson oxygen. Information given by Theel (1875, p.7) and own observations supportthis for Phascolion strombi. A small irrigation is on this background not surprising.
REFERENCES
AKESSON, B. 1958. A study of the nervous system of the Sipunculoideae. Dissertation Lund.(Unders. over Oresund. 38) 249 pp.
BRUMPT, E. 1897. Quelques faits relatifs a l'histoire du Phascolion strombi. Archs Zool. expo gen.3.Ser., 5: 483-496.
DALES, R. P. 1963. Annelids. Hutchinson Univ. Lib., London, 200 pp.FLORKIN, M. 1932. La courbe de dissociation de l'oxyhemerythrine dans Ie liquide coelomique
du Siponcle. C. r. hebd. Seanc. Acad. Sci., Paris, 195: 832-833.- 1933. Le pouvoir oxyphorique du liquide coelomique et des hematies de Sipuncllilis nlldlls.
C. r. Seanc. Soc. BioI., 112: 705-706.FRANZEN, A. & R. FANGE. 1962. Observations on hemerythrin-containing coelomocytes in in
vertebrates. Report from the third Scandinavian conference on cell research, pp. 31-32.Danish Science Press, Copenhagen.
HAMPSON, G. R. 1964. Redescription of a commensal pelecypod, Rochefortia cuenata. with noteson ecology. Nautilus, 77: 125-129.
KNUDSEN, J. 1944. A gephyrean, a polychaete and a bivalve living together (commensalistically)in the Indo-Malayan Seas. Vidensk. Meddr dansk naturh. Foren., 108: 15-24.
LINDROTH, A. 1941. Atmungsventilation der Polychaten. Z. vergl. Physiol., 28: 485-532.NIELSEN, C. 1964. Studies on Danish entoprocta. Ophelia, 1: 1-76.PEREZ, C. 1924. Le complexe ethologique de la Turritelle et du Phascolion strombi. Bull. Soc. zool.
Fr., 49: 341-343.- 1925. Sur Ie complexe ethologique du Phascolion strombi. Ibid., 50: 74-76.SCHLEIP, W. 1934. Die Regeneration des Riissels von Phascolion strombi Mont. (Sipunculidae).
Z. wiss. Zool., 145: 462-469.THEEL, H. 1875. Recherches sur Ie Phascolion strombi (Mont.), K. svenska VetenskAkad. Handl.,
14: 1-32.WELLS, G.P. 1950. Spontaneous activity cycles in polychaete worms. Symp. Soc. expo BioI., 4:
127-142.- 1951. On the behaviour of Sabella. Proc. R. Soc., B, 138: 278-299.






















