Embed Size (px)
Citation preview

Accepted by W.E. Sterrer: 21 Sep. 2011; published: 11 Nov. 2011
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2011 · Magnolia Press
Zootaxa 3096: 27–40 (2011) www.mapress.com/zootaxa/ Article
27
Koinocystididae and Gnathorhynchidae (Platyhelminthes: Rhabdocoela: Kalyptorhynchia) from the Galapagos, with the description of three new species
PATRICK C. REYGEL1, WIM R. WILLEMS & TOM J. ARTOISResearch Group Zoology: Biodiversity & Toxicology, Centre for Environmental Sciences, Hasselt University, Campus Diepenbeek, Agoralaan, Building D, B−3590 Diepenbeek, Belgium1Corresponding author. E-mail: [email protected]
Abstract
Seven species of eukalyptorhynch flatworms from the Galapagos Islands are reported. Six of them belong to the taxonKoinocystididae Meixner, 1924, three of which are new to science. Two of these new species can be attributed to the genusItaipusa Marcus, 1949 based on the construction of the prostate vesicle and the general structure of the female system.Itaipusa biglandula n. sp. is characterised by a very short, muscular, unarmed penis papilla, whereas I. renei n. sp. is char-acterised by the presence of a cirrus armed with minute scales, and two large, blunt hooks in the male atrium. The thirdnew species cannot be placed in any existing taxon and therefore a new genus is erected: Galapagetula annikae n. gen. n.sp.. It is characterised by a straight and rather long cirrus, armed with small spines, and a clearly bipartite bursa in thefemale system. Additional morphological information is given for the three known species of Koinocystididae: I. divaeMarcus, 1949; I. variodentata (Karling, Mack-Fira & Dörjes, 1972) Karling, 1978 and Utelga heinckei (Attems, 1897)Karling, 1954. For I. divae a new locality from Curaçao is also mentioned. The seventh species is a species of Gnatho-rhynchidae: Prognathorhynchus eurytuba Ax & Armonies, 1987, for which new morphological information on the organ-isation of the genital system is given.
Key words: free-living flatworms, Eukalyptorhynchia, taxonomy, biodiversity
Introduction
With in total 64 species recorded, the marine interstitial flatworm fauna of the Galapagos Islands is rather well-known (Ax & Ehlers 1973; Ax & Ax 1974a, b; Ehlers & Ax 1974; Sopott-Ehlers & Schmidt 1974a, b, 1975;Schmidt & Sopott-Ehlers 1976; Ax & Ax 1977; Ehlers & Ehlers 1981; Noldt & Hoxhold 1984; Ehlers & Sopott-Ehlers 1989; Artois & Schockaert 1999, 2000, 2001). This material was collected during intensive sampling cam-paigns by members of the Zoological Institute of the Göttingen University, during a broad-scale study on the inter-stitial fauna of the archipelago. A large number of species of Kalyptorhynchia, free-living flatworms characterisedby the presence of a muscular proboscis, were present in the material. Part of this material was treated in fourpapers: one (Noldt & Hoxhold 1984) dealing with 10 new species of Schizorhynchia, kalyptorhynchs with a splitproboscis, the other three (Artois & Schockaert 1999, 2000, 2001) dealing with in total 14 species of the eukalypto-rhynch taxon Polycystididae Graff, 1905.
In this paper, a total of seven species is reported, six of them belonging to the Koinocystididae Meixner, 1924,while one is a member of the Gnathorhynchidae Meixner, 1929. Three of the koinocystidid species are new to sci-ence and are formally described and their taxonomical position is discussed. The remaining species of Koinocys-tididae, as well as the gnathorhynchid species are reported from the Galapagos for the first time. The material of thespecies of Gnathorhynchidae, Prognathorhynchus eurytuba Ax & Armonies, 1987, is of particular interest, as itreveals some important new morphological details, which could have implications for the taxonomy within thetaxon Gnathorhynchidae.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.28 · Zootaxa 3096 © 2011 Magnolia Press
Material and methods
The animals for this study were collected on the Galapagos Islands by Dr S. Hoxhold (SH) and Dr P. Schmidt, whoalso studied the live animals. We utilised sectioned material (some of which we re-stained) and the drawings andphotographs made by SH. Brief locality data are followed by a number that refers to the detailed descriptions of thelocalities in Ax and Schmidt (1973: 580−586). For Itaipusa divae Marcus, 1949 a new locality outside the Galapa-gos is also mentioned. This material was collected in Curaçao by the last author (TA) and Dr Frank Van Belleghem(Hasselt University) in December 1998–January 1999.
Drawings of live specimens and hard parts are based on initial drawings and notes from SH. Measurementswere made by SH on live material. The measurements of hard parts are taken axially, unless indicated otherwise.As no whole mounts are available, these measures too are based upon drawings and photographs by SH of livespecimens, and provided with a scale bar. Relative positions of structures are expressed as a percentage of the totalbody length, measured from the anterior end. Sections (3 or 4 µm) were stained with Heidenhain’s iron haematox-ylin and eosin as a counter stain.
All material from Galapagos will be deposited in the Zoological Museum of the University of Göttingen(ZMUG). The specimens from Curaçao will be stored in the collections of the research group at Hasselt University(HU).
Abbreviations used in the figures
b, brain; bs, bursal stalk; bu, bursa; cg, caudal glands; cga, common genital atrium; ci, cirrus; de, ejaculatoryduct; ds, spermatic duct; e, eye; fd, female duct; gl, gland; gp, genital pore; h, hook; m, mouth; ma, male atrium;mco, male copulatory organ; ms, muscular septum; od, oviduct; ov, ovary; p, proboscis; pd, prostate ducts, pg,prostate glands; ph, pharynx; pp, penis papilla; prh, proboscis hook; pv, prostate vesicle; s, stylet; sph, sphincter;t, testis; ut, uterus; vd, vitelloduct; vit, vitellarium; vp, vaginal pore; vs, seminal vesicle; x−y, explained in the text.
Taxonomic results
Gnathorhynchidae Meixner, 1929
Prognathorhynchus eurytuba Ax & Armonies, 1987(Fig. 1)
New locality. Santa Cruz Island, Bahia Academy, stations IX5e and IX5f.Known distribution. North West Atlantic: several localities in New Brunswick, Canada and in South Caro-
lina, USA (see Ax & Armonies 1987; Ax 1997).Material. Four individuals studied alive, five sagittally-sectioned specimens (ZMUG 23317-23321).Remarks. The Galapagos specimens can easily be identified as P. eurytuba, based on the overall organisation
of the body, the morphology of the proboscis hooks and that of the stylet. As the original description by Ax andArmonies (1987) and a later account by Ax (1997) were based only on observations on live animals, very little isknown of the detailed morphology, and a reconstruction of the genital system is lacking. The new material from theGalapagos allows a detailed and more complete description, although certain features are still hard to discern, espe-cially concerning details of the female system, which appears to be difficult to study in members of the taxon Prog-nathorhynchus Meixner, 1929 (see Karling 1947; Brunet 1973; Ax & Armonies 1987).
Live animals about 0.7−0.8 mm long. The cellular epidermis (± 5 µm thick) is ciliated over the entire body(cilia ± 5 µm long). A few sensory bristles (± 20 µm long) are present around the proboscis opening. The proboscishooks (Fig. 1C) are as described by Ax and Armonies (1987), only slightly larger. The diameter of the basal plate is± 25 µm (Ax & Armonies 1987: 17 µm). The stylet’s (Fig. 1B) shape is as illustrated by Ax and Armonies (1987).
The stylet length in the Galapagos specimens varies from 42−48 µm ( = 45 µm; n = 3), which is longer than inthe Canadian (Ax & Armonies 1987: 32 µm) and the specimens from South Carolina (Ax 1997: 35 µm).
x
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Zootaxa 3096 © 2011 Magnolia Press · 29KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
The common genital pore (Fig. 1A, D: gp) is situated ventrally, very close to the caudal end and opens into atubiform common genital atrium, which is lined with a low, anucleated epithelium and is surrounded by a longitu-dinal muscle layer. Basophilic caudal glands are situated on both sides of the genital atrium. The genital atriumreceives the male system from the dorsal side, and both the female system and the uterus from the rostral side. Theelongated seminal vesicle is surrounded by strong longitudinal muscles with an oblique orientation in the proximalpart. Sperm is discharged through the funnel-shaped opening of the stylet (see also Ax & Armonies 1987 and Ax1997), whereas the globular prostate vesicle, which contains a coarse-grained basophilic secretion and which issurrounded by circular muscles, discharges through the second, smaller opening of the stylet. The stylet lies in theoviform male atrium, which is surrounded by longitudinal muscles and lined with a very low, anucleated epithe-lium. From the single ovary, the female duct, which is lined with a very low, anucleated epithelium and only dis-tally surrounded by longitudinal muscles, runs ventro-caudally towards the common genital atrium. A very largeand vaguely defined bursa contains numerous large nuclei and sperm in different stages of degeneration as well aslive sperm, organized in separate compartments. From one of these compartments a curved spermatic duct (Fig.1D: ds), surrounded by longitudinal muscles, runs towards the ovary, although the connection proper is hard to dis-cern. A vaginal pore (Fig. 1A, D: vp), situated rostrally from the common genital pore, is clearly visible on all sec-tioned animals, but could only be observed in one out of four live specimens. It is surrounded by a strong sphincterand continues into a vaginal duct, which is surrounded by circular muscles and which leads to the bursa.
FIGURE 1. Prognathorhynchus eurytuba Ax & Armonies, 1987 A, general organisation. B, stylet. C, proboscis hooks (A−Cfrom a live specimen). D, reconstruction of the atrial organs from the left hand side.
In the diagnosis for the taxon Gnathorhynchidae by Karling (1947), the presence of only a single genital pore ismentioned. Also Ax and Armonies (1987) and Ax (1997) do not mention a vaginal pore for P. eurytuba. However,a second opening of the genital system (i.e. a vaginal pore as in P. eurytuba) is also observed by Hochberg (2004)in P. busheki Ax, 1997, using confocal laser scanning microscopy. Apart from P. busheki and P. eurytuba at leasttwo other species of Prognathorhynchus have a female bursa functioning as a seminal receptacle: P. parvulus Bru-
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.30 · Zootaxa 3096 © 2011 Magnolia Press
net, 1973 and P. dividibulbosus Ax & Armonies, 1990. As the vaginal pore is often very difficult to see in live spec-imens, and many of the species of Prognathorhynchus were described without sectioned material, it is possible thata vaginal pore is present in more (even all?) species of this taxon, especially in those in which the presence of afemale bursa has been recorded. It is, therefore, apparent that the taxa Prognathorhynchus and Gnathorhynchidaeare highly in need of taxonomic revision.
It should be checked on animals from the Atlantic populations of P. eurytuba whether they indeed are identicalin all details to the population from Galapagos, and hence indeed belong to the same species. The disparate distri-bution, indeed, could indicate a geographical isolation of both populations. At the moment, however, we cannot butconsider the Galapagos population as belonging to the same species, as at this moment no fixed morphological dif-ferences between the two populations can be found.
Koinocystididae Meixner, 1924
Itaipusa biglandula n. sp.(Fig. 2)
Locality. Santa Cruz Island, Bahia Academy, Station IX5g, upper layer of coarse-grained sand (type locality).Material. Several specimens studied alive. A total of 42 serially-sectioned animals, one of which is designated
holotype (ZMUG 23231), 13 others designated paratypes (ZMUG 23232-23244). Voucher specimens deposited asZMUG 23252-23279.
Etymology. Species name refers to the presence of two symmetrical atrial glands. Glandula (Lat.): gland.Diagnosis. Species of Itaipusa Marcus, 1949, with two atrial glands symmetrically located beside the female
duct; male system with a short, unarmed penis papilla; large bursa with resorptive part and muscular bursal stalkopening into female duct.
Description. Live animals ± 1.3 mm long, caudally rounded, with two eyes. The syncytial epidermis is ± 6 µmthick and shows a prominent basement membrane. It contains numerous apically-elongated, oviform, 2−4 µm longrhabdites, which are absent in the head region, from the anterior tip to half the proboscis length. The animal is cili-ated over the entire body (cilia ± 6 µm long) with sensory bristles (± 25 µm long) surrounding the proboscis open-ing.
The proboscis measures almost 1/5 of the body length and shows the characteristic features of a typical koino-cystidid proboscis (see Brunet 1972; Karling 1980). It has a very strong Itaipusa-type proboscis juncture sphincter(see Karling 1980: p. 260).
The pharynx is situated at about 40%, and does not show any peculiarities in comparison with other species ofItaipusa or Koinocystididae in general (see Meixner (1925) and Karling (1954, 1980) for a detailed account). Theoesophageal glands, clearly visible in sectioned material, are not distinguishable on live animals. A pharynx mouthsphincter could not be observed on sectioned specimens.
Male and female gonads are paired. The oval testes lie dorso-laterally, caudally from the pharynx, extending tothe distal part of the ovaries, which are situated at ± 75%. The vitellaria extend from the mouth to the ovaries, theanterior part being situated dorso-laterally, whereas the posterior part lies more central and ventrally to the testes.The common genital pore, situated at ± 90%, opens into the common genital atrium, which is lined with a high,nucleated epithelium and is surrounded by longitudinal muscles. Anteriorly it receives the uterus, and the male ductmore dorsally. The female system opens into the genital atrium from the dorso-caudal side.
The elongated, paired seminal vesicles are lined with a low, nucleated epithelium and surrounded by longitudi-nal muscles. Both vesicles fuse just before entering the prostate vesicle, forming the ejaculatory duct. This ductruns centrally through the prostate vesicle and is surrounded by eight to ten filiform prostate gland necks, calledprostate ducts by Karling (1980). The prostate glands proper are very large, surrounding the seminal vesicles andcontaining one type of basophilic, coarse-grained secretion. The almost globular prostate vesicle bulges into themale genital atrium in a short, muscular penis papilla. The exact course of the ejaculatory duct could not berevealed, except for its distal part that opens into the penis papilla. This part of the ejaculatory duct is surroundedby circular muscles. The male atrium is lined with a low, anucleated epithelium and does not seem to be surroundedby a muscle layer.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Zootaxa 3096 © 2011 Magnolia Press · 31KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
The ovoid ovaries have short oviducts, which join to form the female duct and are lined with a low, anucleatedepithelium. On this bifurcation, a short vitelloduct enters the female duct. This female duct is lined with a low,nucleated epithelium and surrounded by weak, circular muscle fibres, forming a strong sphincter distally. Althoughno sperm could be observed in this duct, it probably acts as a seminal receptacle, indicated by the fact that it isswollen in all specimens. Instead of opening into the common genital atrium, it is connected to a very muscularbursal stalk, which enters the atrium dorso-caudally, together with two symmetrically-placed, glandular complexeswith fine-grained eosinophilic secretion (Fig. 2B: gl). Over its whole length this stalk is surrounded by circularmuscles, which are extremely thick proximally, close to the bursal organ proper, forming two very large sphincters.The bursa acts as a sperm-resorbing organ (resorptive vesicle of Karling 1980). The bursa has a nucleated epithe-lium, which merges with the sperm-resorbing mass more proximally.
The uterus is lined with a high, nucleated epithelium and surrounded by spirally-running muscle fibres. Itopens into the common genital atrium close to the genital pore and is provided with two types of glands. The firsttype contains a fine-grained eosinophilic secretion and is situated near the proximal end, although the exact loca-tion of the opening into the uterus is unclear. More distally a coarse-grained basophilic gland type is present. Theseglands open into the uterus lumen half way along the organ.
Discussion. See the general discussion on Itaipusa following the remarks on I. variodentata (Karling et al.,1972) Karling, 1978.
FIGURE 2. Itaipusa biglandula n. sp. A, general organisation (from a live specimen). B, reconstruction of the atrial organsfrom the left hand side (from the holotype).
Itaipusa divae Marcus, 1949 (Fig. 3)
New localities in the Galapagos. Santa Cruz Island, stations IX6b, IX5b (both rock pools), IX6a (sandy beach);Baltra, station X3 (fine-grained sand); Barrington, stations XI1 and XI2 (fine-grained sand).
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.32 · Zootaxa 3096 © 2011 Magnolia Press
New localities outside the Galapagos. Several localities in Curaçao (Netherlands Antilles): Spaanse Water,Isla Grandi (N 12°04'57.21", W 68°51'21.21"), detritus-rich sample taken from a littoral sea grass bed (Thalassiatestudinum) at the eastern side of the most narrow point of the peninsula (Dec. 14, 1998); Dam di Cabicuchi(between the Caracasbaai and the Spaanse Water, N 12°04'39.57", W 68°51'48.89"), on Thalassia testudinum withsmaller epiphytic algae (Dec. 14 & 30, 1998); Playa Canoa (N 12°10'30.65", W 68°51'55.19"), on small brownalgae taken from the sheltered side of the narrow spit of land that protects the beach (Dec. 28, 1998).
Known distribution. Two localities in Baia de Santos (Brazil) (see Marcus 1949 for details).Material. Galapagos: two individuals studied alive, nine serially-sectioned ones (ZMUG 23280-23288). Cura-
çao: several individuals studied alive, five whole mounts, two serially-sectioned specimens (HU IV.4.11-IV.4.17).Remarks. The general organisation of our specimens corresponds well with the observations by Marcus
(1949), including the fact that the epidermis is syncytial. As mentioned earlier by Karling (1980) the exact courseof the female duct is hard to discern.
In both live and sectioned animals, the numerous small hooks on the sclerotized folds of the cirrus are longerthan the 2 µm mentioned by Marcus (1949). The pictures (see Fig. 3B for one of them) of the two specimens fromGalapagos show hooks with a length of ± 4 µm on all sclerotized folds. This is also the case in the specimens fromCuraçao, with exception of one specimen that shows a few areas of the sclerotized folds with smaller hooks (1−2µm). On the syntype, Karling (1980) observed some areas with rather large hooks (4−7 µm). This observation indi-cates that, possibly depending on the area of cirrus, the hooks may vary in length between 1 and 7 µm.
In the Galapagos specimens studied alive, the course-grained basophilic prostate ducts have a more saccularappearance. In the sectioned material of both the Galapagos and Curaçao, the exact form of the ducts is not wellvisible, except in two specimens from Galapagos where these ducts are filiform, at least in the distal part of theprostate vesicle. Pictures of the Galapagos specimens reveal large (secretion?) granules scattered around the distalpart of the ejaculatory duct, which could not be found again in the sectioned material. Nevertheless, in all sectionedspecimens some ill-defined, fine-grained, eosinophilic secretion could be observed near to the distal part of theprostate ducts.
FIGURE 3. Itaipusa divae Marcus, 1949 A, general organisation. B, male copulatory organ (A and B from a live specimenfrom Galapagos).
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Zootaxa 3096 © 2011 Magnolia Press · 33KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
Itaipusa renei n. sp.(Fig. 4)
The pharynx is situated at ± 50%. It does not show any peculiarities in comparison with other species of Itai-pusa or Koinocystididae in general (for a detailed account see Meixner 1925 and Karling 1954, 1980). A pharynxmouth sphincter could not be observed. Oesophageal glands are prominent.
Male and female gonads are paired. The two oval testes are situated dorso-laterally at both sides of the phar-ynx. The kidney-shaped ovaries lie dorso-laterally at ± 70%. The vitellaria extend from the pharynx to the commongenital atrium. The common genital pore, situated at ± 90%, opens into this atrium, which can be expanded or con-tracted and folded in several directions. It is surrounded by an outer longitudinal and an inner circular muscle layer,the latter one becoming weaker towards the genital pore. The most distal part of the atrium is lined with a nucleatedepithelium, which lowers to become a membranous epithelium more proximally. In all studied specimens spermare present in the proximal part of the atrium. Dorso-caudally the common atrium is connected to a copulatorybursa, which is filled with degenerating sperm. The bursal stalk is lined with a dense and thickened basement mem-brane (pseudocuticle) and filled with sperm. It is surrounded by strong circular muscles, which form a very largesymmetrical sphincter (Fig. 4E: sph2), situated somewhat distally from the entrance of the bursa. In an immaturespecimen, the developing bursa appears as an eosinophilic globular structure packed with many round nuclei andclearly discernable cells. In adult specimens the bursa shows some flattened nuclei and a central area filled with apatchwork of dense and clear areas.
The male copulatory organ (Fig. 4D, also indicated on Fig. 4E) is situated dorsally in most sectioned specimenswith its proximal part pointing rostrally. It enters the common genital atrium from the left hand side, in between thefemale system and the uterus. The copulatory organ basically consists of an asymmetrical bulb surrounded by amuscular septum that consists of strong inner circular muscles and weak outer longitudinal muscles. The paired,elongated seminal vesicles are lined with a low, nucleated epithelium and surrounded by a circular muscle layerthat runs somewhat spirally. The vesicles fuse just outside the septum, and the ejaculatory duct enters the copula-tory bulb accompanied by several gland necks of the extra-capsular prostate glands. The bulb is partly filled withprostate ducts with coarse-grained basophilic secretion. This secretion is discharged into a flattened cup-like struc-ture, surrounding the pore of the ejaculatory duct, which is surrounded by weak longitudinal muscles. This cup-likesecretion area (Fig. 4D: y) is surrounded by circular muscles and opens through an orifice into the male genitalatrium. Next to this orifice, the male atrium forms a large diverticulum, which is armed with many, small (± 3 µmlong), pointed spines. This diverticulum, therefore, can be considered as an armed cirrus, although it is not directlyconnected to the ejaculatory duct. At one side, this cirrus incorporates a fine-grained eosinophilic secretory area(Fig. 4D: x). Its small gland necks are continuous with the basophilic prostate ducts surrounding the ejaculatoryduct. They penetrate the wall of the cirrus at the side facing the centre of the copulatory bulb, forming an eosino-philic secretion structure, which bulges out of the wall. This secretory area extends from the proximal point of thecirrus towards the cup-like structure mentioned above. The circular muscles of the copulatory bulb continue in thecircular muscles of the diverticulum, which also shows an outer layer of prominent longitudinal muscles on its dor-sal side. The most distal part of the male genital duct is surrounded by inner circular and outer longitudinal mus-cles, and lined with a pseudocuticle. It also contains two hollow, smoothly-curved, blunt hooks (± 50 and 80 µmlong; Fig. 4B, 4D: h). The smaller hook possesses a well-developed basal ring. The exact position of the hookscould not be determined in the sectioned specimens. They are not connected to any muscular structure.
Two short oviducts, which are lined with a low, anucleated epithelium and surrounded by longitudinal muscles,connect the ovaries to the female duct. The oviducts can be closed by a weak distal sphincter. In between the twooviducts the common vitelloduct opens into the female duct through a weak sphincter. This female duct is swollenand filled with sperm, functioning as a seminal receptacle. It is lined with a nucleated epithelium and surroundedby circular muscles over its whole length, accompanied by an external layer of longitudinal muscles at its most dis-tal part. The duct opens into the common genital atrium from the dorsal side and can be closed by a strong symmet-rical sphincter (Fig. 4E: sph1). Close to the opening of the female duct into the atrium, a large bundle of glands ispresent, containing small, fine-grained basophilic glands surrounded by larger, fine-grained, eosinophilic ones.
The uterus is lined with a high, nucleated epithelium and is surrounded by longitudinal muscle fibres. In itsproximal part a small, weak sphincter is present. It is provided with two types of glands. The first type contains afine-grained eosinophilic secretion, whereas the second type is situated just distally of the first type and contains acoarse-grained basophilic secretion.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.34 · Zootaxa 3096 © 2011 Magnolia Press
FIGURE 4. Itaipusa renei n. sp. A, general organisation (from a live specimen). B, hooks (from a live specimen). C, habitus ofretracted individual. D, reconstruction of the male genital system (from the holotype). E, reconstruction (from the holotype) ofthe atrial organs from the right hand side; the position of the male system is indicated in dashed lines.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Zootaxa 3096 © 2011 Magnolia Press · 35KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
Itaipusa variodentata (Karling, Mack-Fira & Dörjes, 1972) Karling, 1978
New locality. Santa Cruz Island, stations IX5b, IX5d, IX6b, rock pools.Known distribution. Hawaii (Karling et al. 1972).Material. Four specimens studied alive, 28 specimens serially-sectioned (ZMUG 23289-23316). Remarks. The complex structure and orientation of the organs in the specimens studied alive are in accordance
with the observations of Karling et al. (1972) on specimens from Hawaii. On sections it is clear that the epidermis iscellular, showing distinctly-delimited epidermal cells as was mentioned in the original description (Karling et al.1972). The Galapagos specimens (3 mm long) are, however, clearly larger than those from Hawaii (Karling et al.1972: 2 mm). The two hooks of the copulatory organ clearly differ in size in comparison with those from Hawaii.
The smaller hook measures 52−108 µm ( = 86 µm; n = 4), whereas the larger one measures 75−140 µm ( = 102
µm; n = 4). This is significantly larger than what was observed by Karling et al. (1972): 30 µm for the small hookand 37 µm for the large hook. However, since no other differences could be observed and since molecular data arelacking in order to back up a splitting of the species, we deliberately choose not to describe the population from theGalapagos as a separate species for the time being.
Discussion on Itaipusa
Both new species of Itaipusa, I. biglandula and I. renei possess all diagnostic characters of the taxon Itaipusa asgiven by Karling (1980): muscular copulatory bulb with an irregular (spiny) cirrus or (spiny) papilla, often com-bined with strong hooks; filiform prostate ducts; seminal receptacle (i.e. female duct) opening distally in a caudally-located bursa; testes at about half body length; strong proboscis juncture sphincter of the Itaipusa-type and a phar-ynx without a closed mouth sphincter.
As to the bursal organs in koinocystidids, Karling (1980) distinguishes between a (primary) bursa, which ismostly surrounded by strong circular muscles, and a resorptive vesicle, which is not muscular, and which is proxi-mally connected to the muscular primary bursa. Additionally, he recognises a “receptacle bursa” which is a swollen,sperm-containing duct between the (stalk of the) primary bursa and the distal sphincter of the seminal receptacle.This terminology is mainly based on conjectures regarding the evolutionary trends in the origin of these structures(for details: see Karling 1980: p. 264−265). However, to allow comparison with other kalyptorhynchs we prefer herea more traditional terminology in which the distal, sperm-resorbing part is called a bursa (the “resorptive vesicle” ofKarling 1980) and the muscular duct that connects it to the common genital atrium a bursal stalk. This bursal stalkcan be swollen and contain sperm (the “primary bursa” of Karling 1980). The “receptacle bursa” of Karling (1980),occurring in a number of species (e.g. I. ruffinjonesi Karling, 1978), on the other hand is typical of koinocystididsand is not found in any other kalyptorhynch taxon.
Whereas the epidermis of I. biglandula is syncytial, the epidermis of I. renei is cellular. Whether the epidermisis cellular or not is an important taxonomical marker in kalyptorhynch systematics. Karling (1980) considers the cel-lular epidermis, consisting of “distinctly delimited cells”, to be a putative symplesiomorphy not only of species ofKoinocystididae, but of rhabdocoels in general. However, he does not elaborate on the distribution of this characterstate in members of the family. In many cases, species descriptions in literature do not mention the nature of the epi-dermis, but it is clear that both a cellular and a syncytial epidermis occur within the taxon Koinocystididae, and evenspecies of the same genus may differ as to this character. Apparently, all species of Utelga Marcus, 1949 have a cel-lular epidermis (Ax 1959), whereas all species of Koinocystis Meixner, 1924 have a syncytial one (Meixner 1924;Karling 1954). For the latter genus, this conclusion is mainly based on the statement by Meixner (1925: p. 261) thatall species of kalyptorhynchs posses a syncytial epidermis. Within the genus Itaipusa, the epidermis can be syncytialor cellular, depending upon the species considered. Our study shows that the epidermis of I. divae is indeed syncy-tial, as was already mentioned by Marcus (1949), who described the epidermal cell borders as “generally indistinct”(Marcus 1949: p.28: os limites inter-celulares, geralmente indistintos). Brunet (1972) also mentions a syncytial epi-dermis for I. similis (Brunet, 1972). I. biglandula, therefore, is the third species for which it is sure that the epidermisis syncytial. Apart from in I. renei, a cellular epidermis is also found in I. acerosa (Brunet, 1972) Karling, 1980 (seeBrunet, 1972), I. scotica (Karling, 1954) Karling, 1978 (see Karling 1954), I. sophiae (Graff, 1905) Karling, 1978(see Ax 1959) and I. variodentata (see Karling et al. 1972; confirmed in this study). Data are lacking for all other
x x
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.36 · Zootaxa 3096 © 2011 Magnolia Press
species of Itaipusa. From the above it is clear that at the moment it is not easy to assess which situation is plesiomor-phic within Koinocystididae. Only a thorough phylogenetic analysis can elucidate this question.
At present 13 species of Itaipusa have been described. Five species (I. bispina Karling, 1980, I. curvicirra Kar-ling, 1980, I. evelinae (Marcus, 1954) Karling, 1980, I. riegeri Karling, 1978 and I. ruffinjonesi) have a male copu-latory organ combining a cirrus armed with small spines and some larger hooks (Karling 1980). This combination isalso found in I. renei, although the spines in this species are considerably smaller and the hooks are not pointed as inthe above-mentioned species. A tubiform penis papilla, as in I. evelinae (see Marcus 1954; Karling 1980) or anaccessory cirrus as in I. riegeri and I. variodentata are missing in I. renei. In I. bispina the hooks are located in sidepockets of the male atrium (Karling 1980), whereas they are situated in the male atrium proper in I. renei and theother four species (Marcus 1954; Karling et al. 1972; Karling 1978, 1980). Another striking feature in this speciesgroup is the detailed structure of the female system and the bursa. In I. evelinae and I. ruffinjonesi, a so-called recep-tacle bursa (the most distal part of the female duct, which is swollen and highly muscular and is separated from theproximal part of the female duct by a strong sphincter; see above) is present (Marcus 1954; Karling 1978, 1980). Inall other species of the group, including I. renei such a receptacle bursa is missing. In I. curvicirra, I. renei, I. riegeriand I. variodentata, the bursa is non-muscular and connected to the genital atrium by a muscular bursal stalk. In I.bispina also the bursa proper is muscular (Karling et al. 1972; Karling 1978, 1980).
The presence of an unarmed penis papilla as in I. biglandula is unique within the taxon Itaipusa. I. scotica; I.sophiae and I. evelinae have a penis papilla, but in these species it is provided with spines, and therefore armed. Thissituation is referred to as a permanently-everted cirrus (Karling 1980). Even within the Koinocystididae an unarmedpenis papilla is very uncommon, and only found in four species: Neoutelga inermis Karling, 1980, Sekerana stolzi(Sekera, 1912) Strand, 1914; Tenerrhynchus magnus Brunet, 1972 and Parautelga biloi Karling, 1964 (see Karling1980). However, these four species differ in many characters from the species of Itaipusa (see Meixner 1925; Kar-ling 1964, 1980; Brunet 1972), whereas the new species from Galapagos shows all diagnostic features of this taxon.Hence we decide to keep the new species within Itaipusa. Another unique feature of I. biglandula is the presence ofthe two globular complexes of glands, surrounding the opening of the female duct into the common genital atrium.
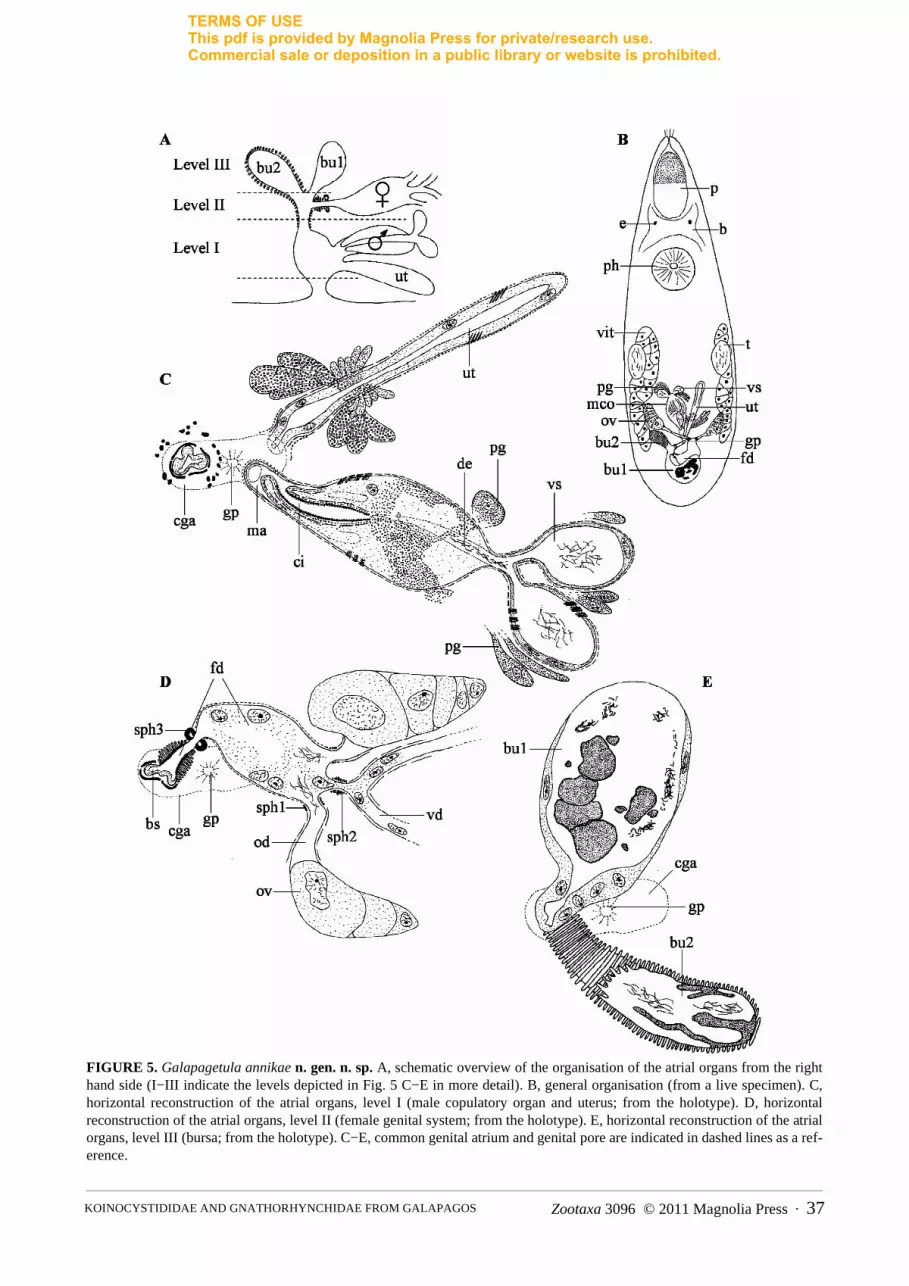
Galapagetula annikae n. gen. n. sp. (Fig. 5)
Locality. Santa Cruz Island, station IX6b, rock pools (type locality).Material. One specimen studied alive and one serially-sectioned specimen designated holotype (ZMUG
23251).Etymology. The genus name is a contraction of “Galapagos”, and “Getula”, a resembling taxon. Species name
dedicated to Annika Willems, daughter of the second author.Diagnoses. Galapagetula n. gen. Koinocystididae with proboscis of the typical koinocystidid construction
without proboscis juncture sphincter; pharynx without a mouth sphincter; testes at mid-body; copulatory organ pear-shaped, distally cylindrical with a long, folded cirrus, armed with small spines; female duct proximally widened,functioning as seminal receptacle, distally highly muscular with strong sphincter; muscular bursa and resorptivebursa opening separately into the female duct.
Type species (by original monotypy): Galapagetula annikae n. sp., provisionally with the same diagnosis as thegenus.
Description. Live specimen grey, spindle shaped, with two eyes. It is ± 1.2 mm long. The syncytial epidermis(± 10 µm thick) has a prominent basement membrane and contains numerous oviform rhabdites (2−4 µm long).They are absent from the anterior tip to half way along the proboscis. The animal is ciliated over its entire body sur-face (cilia ± 6 µm long), with some sensory bristles around the proboscis opening. The proboscis is about 1/5 of thebody length. It has the characteristic traits of a koinocystidid proboscis (see Brunet 1972; Karling 1980). As far ascould be derived from the sections, a proboscis juncture sphincter is absent. The pharynx is situated at 35% andshows no distinct pharynx mouth sphincter. Unfortunately, the material does not allow a detailed reconstruction ofthe pharynx and the proboscis. Observations indicate that the pharynx contains a fine-grained eosinophilic andcoarse-grained basophilic secretion. The lumen is lined with a membranous epithelium.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Zootaxa 3096 © 2011 Magnolia Press · 37KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
FIGURE 5. Galapagetula annikae n. gen. n. sp. A, schematic overview of the organisation of the atrial organs from the righthand side (I−III indicate the levels depicted in Fig. 5 C−E in more detail). B, general organisation (from a live specimen). C,horizontal reconstruction of the atrial organs, level I (male copulatory organ and uterus; from the holotype). D, horizontalreconstruction of the atrial organs, level II (female genital system; from the holotype). E, horizontal reconstruction of the atrialorgans, level III (bursa; from the holotype). C−E, common genital atrium and genital pore are indicated in dashed lines as a ref-erence.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.38 · Zootaxa 3096 © 2011 Magnolia Press
Male and female gonads are paired and situated in the posterior half of the body. The oval testes lie dorso-later-ally, at the beginning of the second half of the body. The spindle-shaped ovaries are situated at ± 75%. The shortvitellaria lie ventro-laterally and extend from the level of the testes to the ovaries. The common genital pore is situ-ated at ± 85% and opens into the globular common genital atrium, which is lined with a nucleated epithelium andsurrounded by an outer longitudinal and a less prominent, inner circular muscle layer. It receives the uterus and themale and female genital system at three different “levels” (see schematic overview in Fig. 5B). The elongateduterus enters the common genital atrium rostro-ventrally, with the male copulatory organ situated just dorsally tothe uterus and also opening into the atrium from the rostral side (Fig. 5A: Level I; Fig. 5C). The female systementers the atrium from the dorsal side (Fig. 5A: Level II; Fig. 5D). The most distal part of the female duct, which ishighly muscular, receives two different bursal organs Fig. 5A: Level III; Fig. 5E).
The male genital system consists of an oval to pear-shaped copulatory organ with an armed cirrus. The seminalvesicles are almost globular and lined with a low, nucleated epithelium and surrounded by a spirally–running,almost circular muscle layer. Distally from the seminal vesicles, the vasa deferentia fuse just before entering theprostate vesicle, forming an ejaculatory duct that contains sperm in its proximal part. This duct runs centrallythrough the prostate vesicle and is surrounded by several prostate ducts with coarse-grained basophilic secretion.The cell bodies of the prostate glands are visible beside the seminal vesicles and the prostate vesicle (Fig. 5C: pg).The exact course of the prostate ducts is difficult to discern, but there are indications that they enter the prostatevesicle a little distally from the ejaculatory duct. The muscle layer surrounding the seminal vesicles continuesaround the prostate vesicle. In contrast to observations on the live animal, the trajectory of the individual prostateducts and the ejaculatory duct are not clearly visible on the sectioned material. However, they most probably enterthe proximal part of the cirrus, which is situated in the most distal half of the male copulatory organ. The cirrus islined with a thick basement membrane, which forms numerous minute, scale-like hooks (0.1 µm or less). The cir-rus proper is surrounded by an inner layer of circular muscles and an outer layer of longitudinal muscles. It isfolded, making its course very hard to reveal. At its distal end it forms a penis papilla-like structure, which pro-trudes into the male duct.
The elongated, almost spindle-shaped ovaries are connected to the female duct by short oviducts, which aresurrounded by longitudinal muscles and are provided with a weak distal sphincter (Fig. 5D: sph1). An epitheliumcould not be observed. Both vitelloducts fuse into a very short single duct, which is provided with a strong sphinc-ter, just before entering the female duct in between the two oviducts (see Fig. 5D: sph 2). The most proximal part ofthe female duct is surrounded by circular muscles. As for the oviducts, an epithelium cannot be seen. The femaleduct is clearly widened and contains many vitellocytes and some sperm close to the oviducts, functioning as a sem-inal receptacle. More distally it narrows and can be closed by a very strong sphincter (see Fig. 5D: sph 3). Fromthis point the female duct is lined with pseudocuticle (dense and thickened basement membrane not covered by epi-thelium) and surrounded by a very thick circular muscle layer, before entering the muscular bursal stalk. The bursalsystem consists of a muscular bursal stalk, which proximally ends in a large bursa (Fig. 5A, B, E: bu1). The bursais lined with a nucleated epithelium and filled with a mass of different density, wherein several nodular structuresand clusters of sperm can be recognised. The bursal stalk also forms a very muscular diverticulum which containssome sperm and is lined with a thick irregularly-folded pseudocuticle: the accessory bursa (Fig. 5A, B, E: bu2). It issurrounded by strong circular muscles that stain rather weakly compared to those of the female duct.
The uterus is lined with a nucleated epithelium and surrounded by spirally-running muscle fibres, distally con-tinuing as longitudinal muscles. At the distal end a second outer circular muscle layer is present around the stalk.Two types of uterus glands are present: fine-grained eosinophilic ones proximally and coarse-grained basophilicones distally.
Discussion. The new species shows all features mentioned in the diagnosis of the taxon Koinocystididae,given by Karling (1980). However, the specific combination of features shown by this species is unique, and doesnot allow its placement in any existing genus. The only other taxa without a proboscis juncture sphincter withinKoinocystididae are Axiutelga Karling, 1980, Getula Karling, 1980, Itaipusina Karling, 1980, Leguta Karling,1980, Parautelga Karling, 1964, Pontaralia Mack-Fira, 1968, Rhinolasius Marcus, 1951 and Utelga. G. annikaeclearly differs from all these taxa, and the other koinocystidids, in the presence of the accessory bursa. In a numberof species (e.g. Itaipusa ruffinjonesi) the bursal stalk is enlarged towards the entrance of the seminal receptacle, anenlargement called “receptacle bursa” by Karling (1980). The position of the accessory bursa in G. annikae is, how-ever, clearly different from that of the “receptacle bursa” described by Karling (1980).
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Zootaxa 3096 © 2011 Magnolia Press · 39KOINOCYSTIDIDAE AND GNATHORHYNCHIDAE FROM GALAPAGOS
The fact that the epidermis is syncytial is not uncommon within the Koinocystididae. For a discussion on thistopic, see the discussion on Itaipusa.
A cirrus provided with uniform, small spines is also rather exceptional within Koinocystididae. Either the cir-rus is unarmed, or the spines are not uniform, or additional large hooks are present. Basically, only in Axiutelgadoes the cirrus show uniform, albeit much larger spines than in G. annikae (7−10 µm and 0.1 µm respectively). Theunique construction of the female system combined with a uniformly-spined cirrus warrants the erection of a newgenus for G. annikae.
Utelga heinckei (Attems, 1897) Karling, 1954
Locality. Santa Cruz Island, Station IX6b, rock pools.Known distribution. North Atlantic: West Coast of USA, North Sea, Skagerrak, Irish Sea, Irish Atlantic coast
(see Karling 1980 and references therein; Willems et al. 2007).Material. One individual studied alive, one sagittally-sectioned specimen (ZMUG 23322). Remarks. The internal structure is in accordance with the observations and remarks by Karling (1954, 1980).
The epidermis is clearly cellular, as was mentioned by Karling (1954). One of the hooks associated with the copu-latory organ is 40 µm long, slightly larger than the other two (35 µm). These two smaller hooks also differ slightlyin shape from the third, larger hook. Their basal plate is smaller and the gutter-shape of the spine-like part is lessprominent. Although the measurements are slightly larger, they correspond well with earlier data (e.g. Karling1954: 22.5−28 µm; Karling 1980: 22−35 µm; Willems et al. 2007: 10−23 µm). Difficulties with interpretation ofshape and size of the hooks are mentioned by Willems et al. (2007).
Acknowledgements
The authors would like to thank Prof. Dr Peter Ax for sending us the sectioned material and the drawings and pho-tographs made by Dr Siegmar Hoxhold. The Galapagos Project was financially supported by the StiftungVolkswagenwerk. Furthermore, we want to thank Mrs Natasha Steffanie (Hasselt University, Belgium) for thepreparation of the serially-sectioned material from Curaçao and for restaining part of the Galapagos material. Weare also indebted to Dr Frank Van Belleghem (Hasselt University) for his assistance in collecting specimens onCuraçao. Dr Nikki Watson is thanked for her most valuable linguistic comments. The research was partly fundedby a project G.0208.08 of FWO-Vlaanderen.
References
Artois, T.J. & Schockaert, E.R. (1999) Interstitial fauna of the Galapagos: Porrocystidinae (Plathelminthes, Polycystididae).Tropical Zoology, 12, 309−324.
Artois, T.J. & Schockaert, E.R. (2000) Interstitial fauna of the Galapagos: Typhlopolycystidinae (Plathelminthes, Polycystidi-dae). Tropical Zoology, 13, 141−158.
Artois, T.J. & Schockaert, E.R. (2001) Interstitial fauna of the Galapagos: Duplacrorhynchinae, Macrorhynchinae, Polycystidi-nae, Gyratricinae (Plathelminthes, Polycystididae). Tropical Zoology, 14, 63−85.
Attems, C.G. (1897) Beitrag zur Kenntnis der rhabdocoelen Turbellarien Helgolands. Wissenschaftliche Meeresuntersuchungender Kommission Abteilung Kiel, 2, 219−232.
Ax, P. (1959) Zur Systematik, Ökologie und Tiergeographie der Turbellarienfauna in den ponto-kaspischen Brackwasser-meeren. Zoologische Jahrbücher (Abteilung für Systematik, Ökologie und Geographie der Tiere), 87, 43−184.
Ax, P. (1997) Two Prognathorhynchus species (Kalyptorhynchia, Plathelminthes) from the North Inlet Salt marsh of HobcawBarony, South Carolina, USA. Microfauna Marina, 11, 317−320.
Ax, P. & Armonies, W. (1987) Amphiatlantic identities in the composition of the boreal brackish water community of Plathel-minthes. A comparison between the Canadian and European Atlantic coast. Microfauna Marina, 3, 7−80.
Ax, P. & Armonies, W. (1990) Brackish water plathelminthes from Alaska as evidence for the existence of a boreal brackishwater community with circumpolar distribution. Microfauna Marina, 6, 7−109.
Ax, P. & Ax, R. (1974a) Interstitielle Fauna von Galapagos 5. Otoplanidae (Turbellaria, Proseriata). Mikrofauna des Meeres-bodens, 27, 573−598.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

REYGEL ET AL.40 · Zootaxa 3096 © 2011 Magnolia Press
Ax, P. & Ax, R. (1974b) Interstitielle Fauna von Galapagos 7. Nematoplanidae, Poystyliphoridae, Coelogynoporidae (Turbel-laria, Proseriata). Mikrofauna des Meeresbodens, 29, 613−638.
Ax, P. & Ax, R. (1977) Interstitielle Fauna von Galapagos 19. Monocelididae (Turbellaria, Proseriata). Mikrofauna des Meeres-bodens, 64, 397−438.
Ax, P. & Ehlers, U. (1973) Interstitielle Fauna von Galapagos 3. Promesostominae (Turbellaria, Typhloplanoida). Mikrofaunades Meeresbodens, 23, 203−216.
Ax, P. & Schmidt, P. (1973) Interstitielle Fauna von Galapagos 1. Einführung. Mikrofauna des Meeresbodens, 20, 563−598.Brunet, M. (1972) Koinocystididae de la région de Marseille (Turbellaria, Kalyptorhynchia). Zoologica Scripta, 1, 157−174.Brunet, M. (1973) Turbellaries Calyptorhynques de la région marseillaise. Les familles des Placorhynchidae et Gnathorhynchi-
dae. Bulletin de la Société Zoologique de France, 98, 121−135.Ehlers, U. & Ax, P. (1974) Interstitielle Fauna von Galapagos 8. Trigonostominae (Turbellaria, Typhloplanoida). Mikrofauna
des Meeresbodens, 30, 641−671.Ehlers, U. & Ehlers, B. (1981) Interstitielle Fauna von Galapagos 27. Byrsophlebidae, Promesostomidae, Brinkmanniellinae,
Kytorhynchidae (Turbellaria, Typhloplanoida). Mikrofauna des Meeresbodens, 83, 83−115.Ehlers, U. & Sopott-Ehlers, B. (1989) Interstitielle Fauna von Galapagos 38. Haloplanella Luther und Pratoplana Ax (Typhlo-
planoida, Plathelminthes). Microfauna Marina, 5, 189−206.Hochberg, R. (2004) Reproductive anatomy of Prognathorhynchus busheki Ax (Platyhelminthes, Kalyptorhynchia) revealed by
confocal laser scanning microscopy. Meiofauna Marina, 13, 29−36.Karling, T.G. (1947) Studien über Kalyptorhynchien (Turbellaria) 1.Die Familien Placorhynchidae und Gnathorhynchidae.
Acta Zoologica Fennica, 50, 1−65.Karling, T.G. (1954) Einige marine Vertreter der Kalyptorhynchien-Familie Koinocystididae. Arkiv för Zoologi, 7, 165−183.Karling, T.G. (1964) Über einige neue und ungenügend bekannte Turbellaria Eukalyptorhynchia. Zoologischer Anzeiger, 172,
159−183.Karling, T.G. (1978) Anatomy and systematics of marine turbellaria from Bermuda. Zoologica Scripta, 7, 225−248.Karling, T.G. (1980) Revision of Koinocystididae (Turbellaria). Zoologica Scripta, 9, 241−169.Karling, T.G., Mack-Fira, V. & Dorjes, J. (1972) First report on marine microturbellarians from Hawaii. Zoologica Scripta, 1,
251−269.Mack-Fira, V. (1968) Pontarallia beklemichevi n. gen. n. sp., un kalyptorhynque relicque du bassin Ponto-aralo-caspien.
Travaux du Museum d'histoire naturelle Grigore Antipa, 8, 333−341.Marcus, E. (1949) Turbellaria Brasileiros (7). Boletins da Faculdade de Filosofia Ciencias e Letras, Universidade de São
Paulo Zoologia, 14, 7−155.Marcus, E. (1951) Turbellaria Brasileiros (9). Boletins da Faculdade de Filosofia, Ciencias e Letras, Universidade de São
Paulo Zoologia, 16, 1−215.Marcus, E. (1954) Turbellaria Brasileiros (11). Papéis avulsos, 11, 419−489.Meixner, J. (1924) Studien zu einer Monographie der Kalyptorhynchia und zum System der Turbellaria Rhabdocoela. Zoolo-
gischer Anzeiger, 60, 89−105.Meixner, J. (1925) Beitrag zur Morphologie und zum System der Turbellaria-Rhabdocoela: 1. Die Kalyptorhynchia. Zeitschrift
fur Morphologie und Ökologie der Tiere, 3, 255−343. Meixner, J. (1929) Morphologisch-ökologische Studien an neuen Turbellarien aus dem Meeressande der Kieler Bucht.
Zeitschrift fur Morphologie und Ökologie der Tiere, 14, 765−791.Noldt, U. & Hoxhold, S. (1984) Interstitielle Fauna von Galapagos - 34. Schizorhynchia (Plathelminthes, Kalyptorhynchia).
Microfauna Marina, 1, 199−256.Schmidt, P. & Sopott-Ehlers, B. (1976) Interstitielle Fauna von Galapogos - 15. Macrostomum O. Schmidt, 1848 und Sicco-
macrostomum triviale nov.gen. nov.spec. (Turbellaria, Macrostomida). Mikrofauna des Meeresbodens, 57, 363−404.Sekera, E. (1912) Studien über Turbellarien. Sitzungsberichte der Königlichen Böhmischen Gesellschaft der Wissenschaften,
13, 1−38.Sopott-Ehlers, B. & Schmidt, P. (1974a) Interstitielle Fauna von Galapagos - 9. Dolichomacrostomidae. Mikrofauna des
Meeresbodens, 34, 103−120.Sopott-Ehlers, B. & Schmidt, P. (1974b) Interstitielle Fauna von Galapagos - 12. Myozona Marcus (Turbellaria, Macrosto-
mida). Mikrofauna des Meeresbodens, 46, 373−389.Sopott-Ehlers, B. & Schmidt, P. (1975) Interstitielle Fauna von Galapagos 14. Polycladida (Turbellaria). Mikrofauna des
Meeresbodens, 54, 193−222.Strand, E. (1914) Neue Namen verschiedener Tiere. Archiv für Naturgeschichte, 80, 163−164.von Graff, L. (1905) Marine Turbellarien Orotavas und der Küsten Europas. Zeitschrift fur wissenschaftlige Zoologie, 83,
68−148.Willems, W.R., Sandberg, M.I. & Jondelius, U. (2007) First report on Rhabdocoela (Rhabditophora) from deep parts of Skager-
rak, with the description of four new species. Zootaxa, 1616, 1−21.
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.





























