Embed Size (px)
Citation preview

LA TEORÍA CELULAR Y EL CONCEPTO DE CÉLULA
S.XVII: A.VanLeewenhoek construyó el primer microscopio
Hooke observó laminillas de corcho y describió lacélula.
1831, Brown demostró la existencia de un corpúsculo en el interior de las células, al que denominó núcleo
1837, Purkinje, introdujo el término de protoplasma
1838-39, por Schleiden&Schwann: las plantas y animales están constituidos por una o más unidades fundamentales: las células.
1. Todo en los seres vivos está formado por células o por sus productos de secreción: unidad anatómica dela materia viva,
2. Todas las células proceden de células preexistentes, por división de éstas (Omniscellula e cellula). Virchow (1858)
3. Las funciones vitales de los organismos ocurren dentro de las células, o en su entorno inmediato: unidad fisiológica de la vida.
4. Cada célula contiene toda la información hereditaria necesaria : unidad genética.
1855, por Virchow: toda célula procede de otra célula.
La teoría celular:1. -La célula es la unidad anatómica y fisiológica de todos los seres vivos, pues todos los organismos están constituidospor una o varias células2. -La célula es la unidad de origen, cada célula procede de otra célula anterior a ella por división de la misma3. -La información genética necesaria para el mantenimiento

de la existencia de la célula y la producción de nuevas células se transmite de una generación a la siguiente4. -Las reacciones químicas que constituyen el metabolismo tienen lugar en las células.
Posteriormente se seguía considerando al tejido nervioso comouna excepción. Fue Ramón y Cajal el que demostró que está formado por células independientes.
1937: construcción del primer microscopio electrónico. observándose los primeros orgánulos,
NIVELES DE ORGANIZACION CELULAR
En el nivel celular es donde las estructuras alcanzan el grado de complejidad necesario para manifestar propiedades vitales
En el nivel celular es donde las estructuras alcanzan el grado de complejidad necesario para manifestar propiedades vitales
Según el grado de complejidad se consideran dos tipos de organización celular: la célula procariota y la eucariota y aún se considera un tercer tipo, formado por los virus, cuyaextraordinaria simplicidad estructural y la carencia de metabolismo propio los convierte en organismos entre lo vivo y lo inerte. Hay organismos formados por una sóla célula: seres unicelulares. Así ocurre con el grupo de los protozoos. A veces, los organismos unicelulares se agrupan formando colonias en las que todos los individuos son iguales,como las Vorticelas.En otras, los individuos son distintos y se especializan en determinadas funciones, como las colonias de Volvox. Frecuentemente los organismos están constituidos por muchas células y dan orígen a los seres pluricelulares .En estos organismos las células se especializan en determinadas funciones dando orígen a los tejidos. Los primeros seres vivos que aparecieron sobre elplaneta hace unos 3500 millones de años fueron unicelulares y

durante más de 2000 millones de años los seres con una célulafueron los únicos pobladores del planeta
LA CÉLULA PROCARIOTA
Su origen evolutivo es anterior al de eucariotas,están normalmente aisladas y dan lugar a organismos unicelulares denominados moneras.Con frecuencia forman colonias o filamentos de células independientes, unidas entresí por sustancias extracelulares.Tamaño : 1-10 mm -Carece de verdadero núcleo aislado por una membrana nuclear.-Material cromosómico formado por una doble hélice de DNA desnudo, que no se asocia con proteínas del tipo de las histonas y se localiza en una región central denominada nucleoide, sin separación del resto.-Protoplasma poco diferenciado y aunque existen un conjunto de sistemas membranosos formados por repliegues internos de la membrana plasmática, no se alcanza un alto grado de compartimentación.-Contiene ribosomas, a veces flagelos, cromatóforos con actividad fotosintética, inclusiones lipídicas y el equipo enzimático que les permite independizarse.-Suele contener pared celular
En bacterias Son microorganismos adaptados a vivir en cualquier ambiente, lo que las convierte en organismos indispensables para el reciclaje de la materia. Forma: bacilos, cocos, estafilococos (arracimados) estreptococos(arrosariados), espirilos (hélice), vibrios (coma)
La membrana está plegada hacia el interior formando los mesosomas, que intervienen en el intercambio de sustancias con el exterior y con la división celular.Contienen además enzimas respiratorios, que en las eucariotas se encuentran en las mitocondrias. La pared celular es fuerte y rígida,
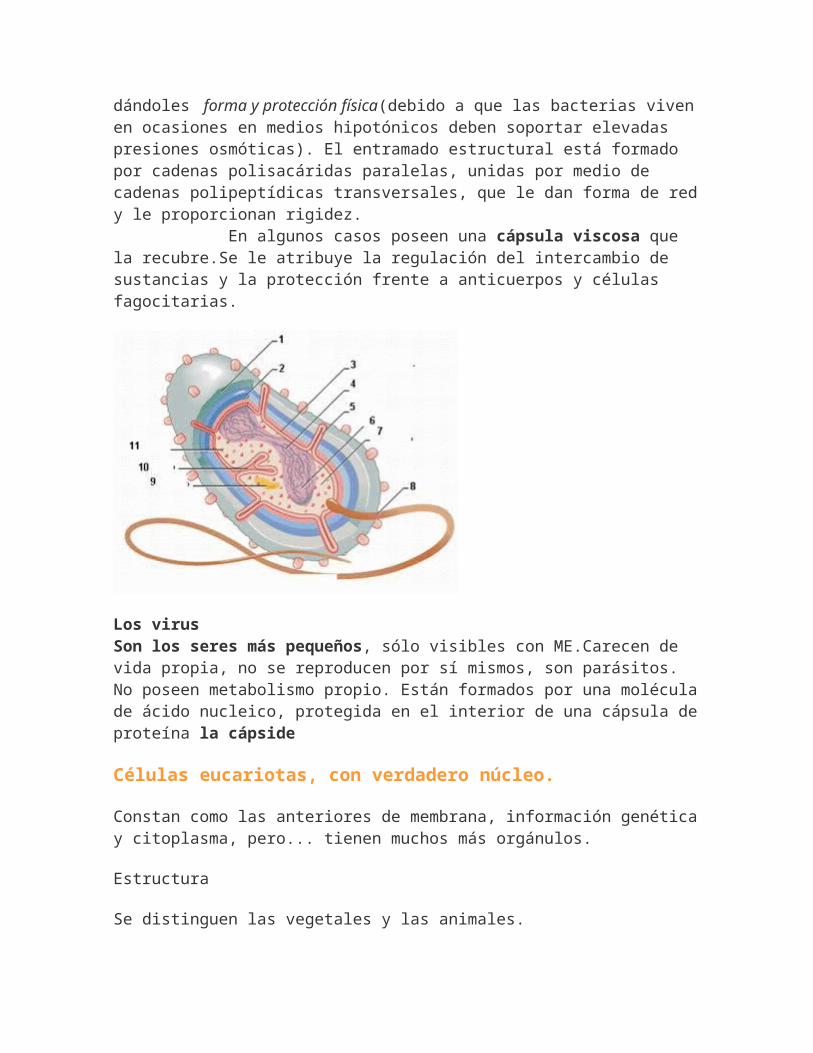
dándoles forma y protección física(debido a que las bacterias vivenen ocasiones en medios hipotónicos deben soportar elevadas presiones osmóticas). El entramado estructural está formado por cadenas polisacáridas paralelas, unidas por medio de cadenas polipeptídicas transversales, que le dan forma de redy le proporcionan rigidez. En algunos casos poseen una cápsula viscosa que la recubre.Se le atribuye la regulación del intercambio de sustancias y la protección frente a anticuerpos y células fagocitarias.
Los virusSon los seres más pequeños, sólo visibles con ME.Carecen de vida propia, no se reproducen por sí mismos, son parásitos. No poseen metabolismo propio. Están formados por una moléculade ácido nucleico, protegida en el interior de una cápsula deproteína la cápside
Células eucariotas, con verdadero núcleo.
Constan como las anteriores de membrana, información genéticay citoplasma, pero... tienen muchos más orgánulos.
Estructura
Se distinguen las vegetales y las animales.

Funciones de reproducción
CÉLULAS EUCARIOTAS: MORFOLOGÍA DE LA CÉLULA ANIMAL; DIFERENCIAS MORFOLÓGICAS ENTRE LA CÉLULA ANIMAL Y LA VEGETAL.-Poseen un núcleo delimitado por una envuelta: la membrana celular-Citoplasma compartimentado por membranas que separan orgánulos celulares, cada uno con una función-Membrana plasmática
FORMA Y TAMAÑO DE LAS CÉLULAS
-En unicelulares: forma semiglobular.oTras, debido a estructura semirígida debido a citoesqueleto de estructura rígida: así amebas y leucocitos que cambian de forma al desplazarse-En pluricelulares, tejidos: neuronas, musculares, vegetales,epiteliales...Tamaño : 10-50, aunque las hay visibles a simple vista: huevos, el de galllina 30 mm y el de avestruz 100 mm.La Acetabularia llega a medir 10 cm
Diferencias célula animal y vegetal-Hoja de células
LA MEMBRANA PLASMÁTICA
Modelo desarrollado por Singer&Nicolson en 1972: "Mosaico fluido"Formada por un mosaico fluido de proteínas, que flotan como icebergs, inmersas en un mar de fosfolípidos; estos se disponen en una bicapa lipídica que contiene, además otros lípidos, como los esfingolípidos y el colesterol.
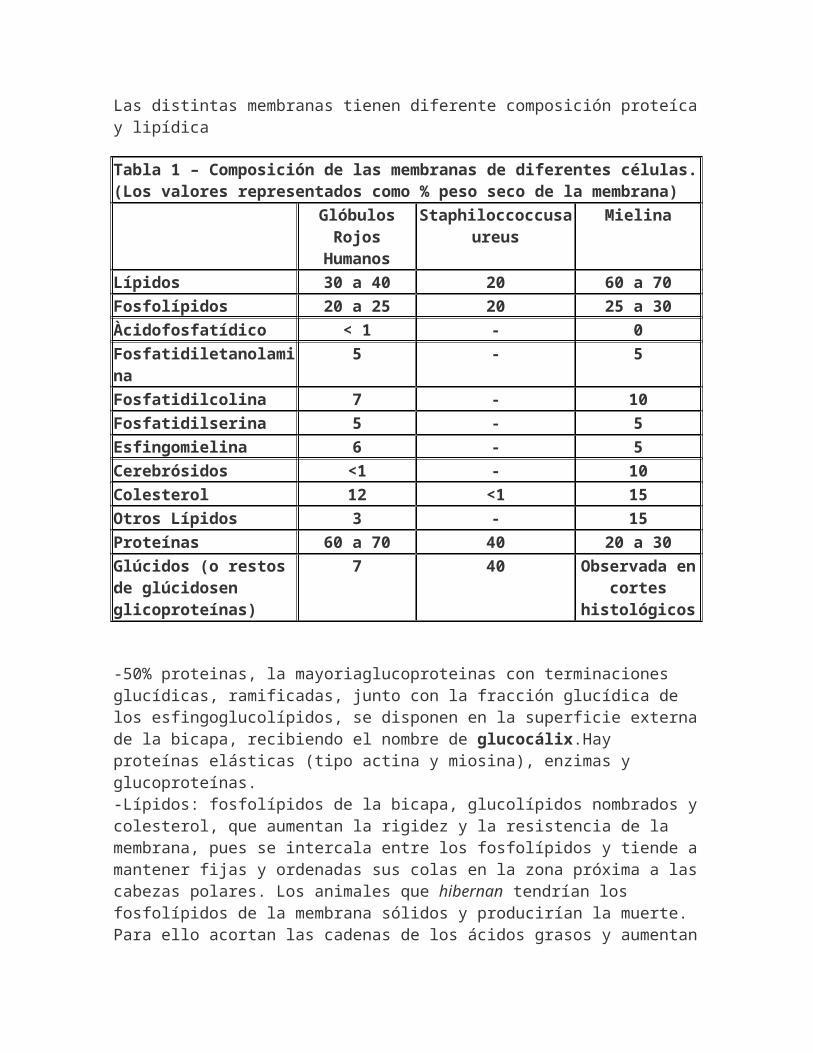
Las distintas membranas tienen diferente composición proteícay lipídica
Tabla 1 – Composición de las membranas de diferentes células.(Los valores representados como % peso seco de la membrana) Glóbulos
RojosHumanos
Staphiloccoccusaureus
Mielina
Lípidos 30 a 40 20 60 a 70Fosfolípidos 20 a 25 20 25 a 30Àcidofosfatídico < 1 - 0Fosfatidiletanolamina
5 - 5
Fosfatidilcolina 7 - 10Fosfatidilserina 5 - 5Esfingomielina 6 - 5Cerebrósidos <1 - 10Colesterol 12 <1 15Otros Lípidos 3 - 15Proteínas 60 a 70 40 20 a 30Glúcidos (o restos de glúcidosen glicoproteínas)
7 40 Observada encortes
histológicos
-50% proteinas, la mayoriaglucoproteinas con terminaciones glucídicas, ramificadas, junto con la fracción glucídica de los esfingoglucolípidos, se disponen en la superficie externade la bicapa, recibiendo el nombre de glucocálix.Hay proteínas elásticas (tipo actina y miosina), enzimas y glucoproteínas.-Lípidos: fosfolípidos de la bicapa, glucolípidos nombrados ycolesterol, que aumentan la rigidez y la resistencia de la membrana, pues se intercala entre los fosfolípidos y tiende amantener fijas y ordenadas sus colas en la zona próxima a lascabezas polares. Los animales que hibernan tendrían los fosfolípidos de la membrana sólidos y producirían la muerte. Para ello acortan las cadenas de los ácidos grasos y aumentan

el grado de insaturación para favorecer la fluidez y evitar la solidificación. Mientras que los seres que aguantan Tª muyaltas (a las que las membranas se podrían disgregar) incorporan ácidos grasos saturados de cadena larga que aumentan la rigidez y los fosfolípidos forman una monocapa enla que las dos zonas hidrófilas se encuentran unidas covalentemente a través de las colas
Los fosfolípidos en un medio acuoso se disponen formando capas. Esta capa es fluida debido a que los lípidos se mueven con entera libertad. Las moléculas pueden cambiar de sitio con respecto a sus vecinas, flexionarse y raramente cambiar de capa.
Existen tres tipos de movimientos posibles en las membranas:
rotación (sobre su propio eje) traslación (o difusión lateral) sobre el plano de la
membrana. flip-flop
El movimiento de flip-flop es el intercambio de fosfolípidos de una monocapa (o hemimembrana) a la otra; esta sumamente restringido, debido a la dificultad que posee la cabeza polarpara atravesar el medio hidrofóbico de la matriz de la membrana. De allí que no sea un movimiento que ocurra de manera espontánea sino que está mediado por enzimas denominadas flipasas.
las membranas son estructuras dinámicas donde los componentespueden desplazarse en todas las direcciones sobre el plano dela bicapa. De ahí que el modelo reciba el nombre de mosaico fluido.

Fig. 4.12 - Movimientos de los fosfolípidos en una bicapa liplídica
Debido a esta fluidez, la membrana es flexible, potenciado esto por el colesterol.-Las sustancias liposolubles atraviesan la membrana con más rapidez que las hidrosolubles.-Las proteínas de membrana ocupan distintas posiciones dependiendo de su solubilidad: Las hidrosolubles están situadas en los bordes de la bicapa y establecen uniones débiles con las cabezas de los fosfolípidos: extrínsecas o periféricas.Las hidrófobas están inmersas más o menos profundamente en las capas lipídicas, estableciendo fuertes uniones con los fosfolípidos. La parte hidrófila suele quedar en el medio extracelular, citoplasma o en ambas: intrínsecas o integrales.-No es simétrica, las proteínas periféricas son distintas en cada una de sus caras ya que las partes glicídicas de las glucoproteínas están en contacto con el medio extracelular
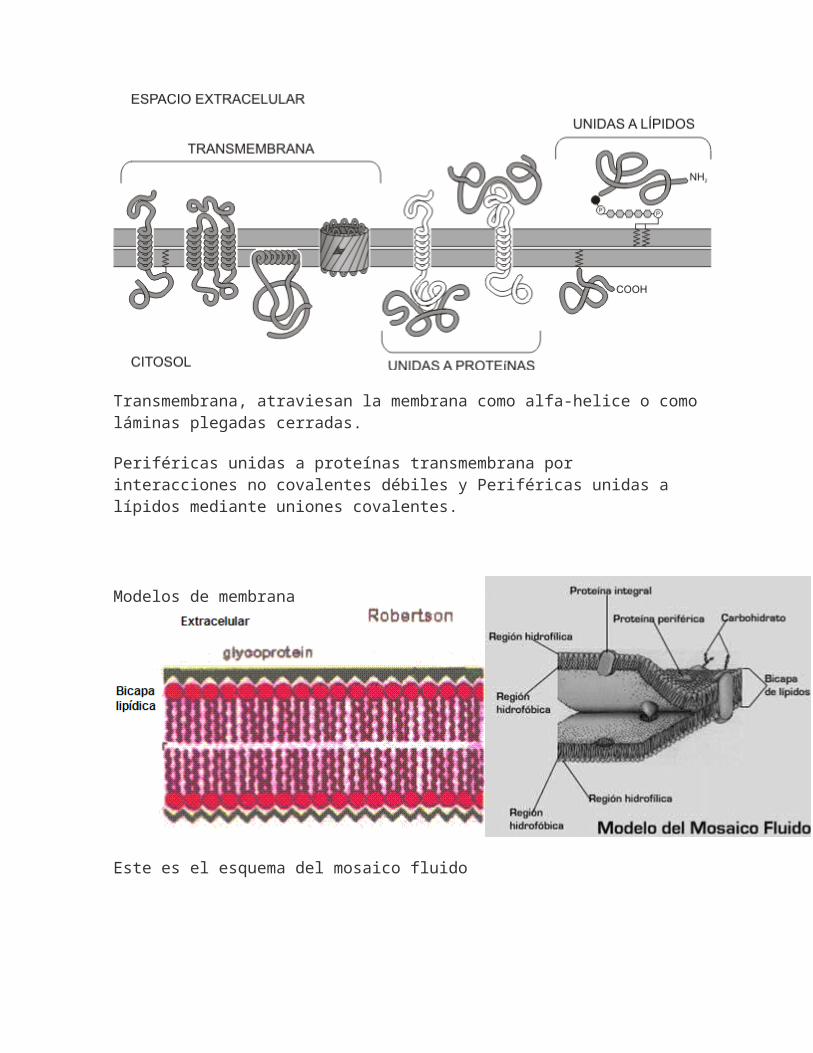
Transmembrana, atraviesan la membrana como alfa-helice o comoláminas plegadas cerradas.
Periféricas unidas a proteínas transmembrana por interacciones no covalentes débiles y Periféricas unidas a lípidos mediante uniones covalentes.
Modelos de membrana
Este es el esquema del mosaico fluido
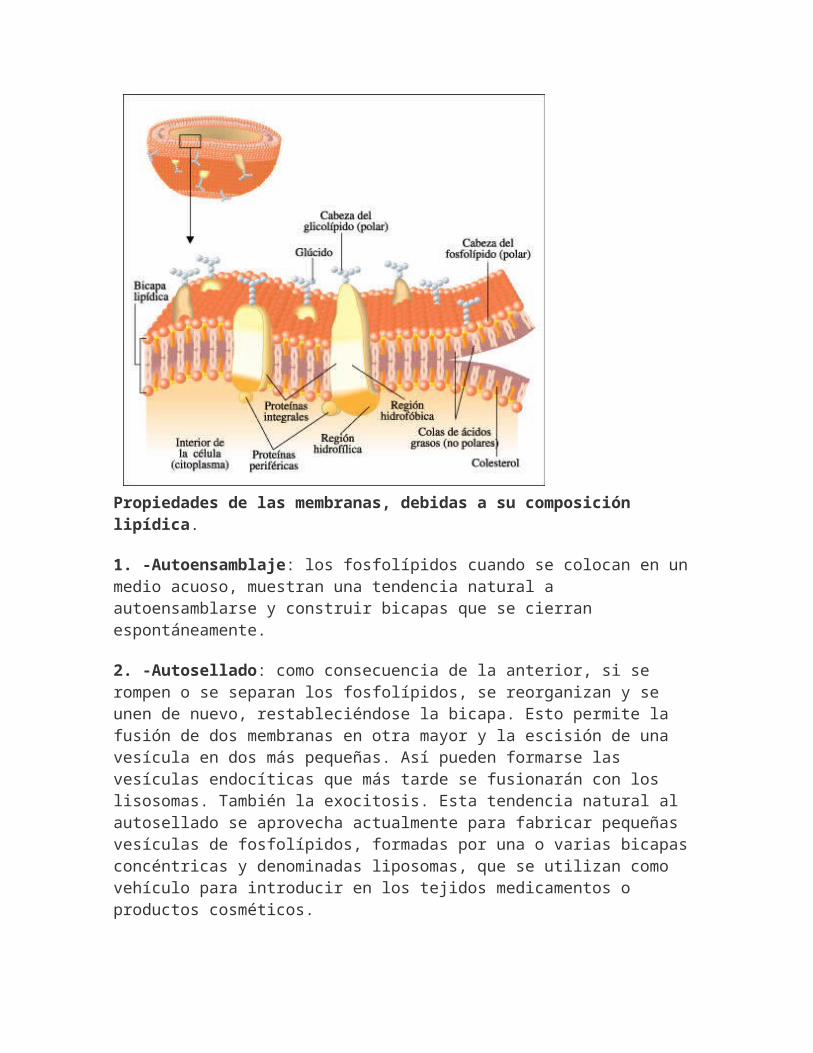
Propiedades de las membranas, debidas a su composición lipídica.
1. -Autoensamblaje: los fosfolípidos cuando se colocan en un medio acuoso, muestran una tendencia natural a autoensamblarse y construir bicapas que se cierran espontáneamente.
2. -Autosellado: como consecuencia de la anterior, si se rompen o se separan los fosfolípidos, se reorganizan y se unen de nuevo, restableciéndose la bicapa. Esto permite la fusión de dos membranas en otra mayor y la escisión de una vesícula en dos más pequeñas. Así pueden formarse las vesículas endocíticas que más tarde se fusionarán con los lisosomas. También la exocitosis. Esta tendencia natural al autosellado se aprovecha actualmente para fabricar pequeñas vesículas de fosfolípidos, formadas por una o varias bicapas concéntricas y denominadas liposomas, que se utilizan como vehículo para introducir en los tejidos medicamentos o productos cosméticos.

3. -Fluidez: no existen enlaces covalentes entre los fosfolípidos, ni con las proteínas, sino que se mantiene por la acción de enlaces débiles, lo que les confiere una gran fluidez y flexibilidad, ya que todos los componentes se pueden desplazar lateralmente.
4. -Impermeabilidad: frente a moléculas hidrófilas, sobre todo si tienen un tamaño considerable. Lo que hace que actúe como una barrera de contención.Pero a través de la membrana plasmática se realizan todos losintercambios de materia y energía con el ambiente externo. Por ello se desarrollan sistemas de transporte que permiten el paso.
FUNCIONES BIOLOGICAS
-Mantener cerrada a la célula-Mantener una permeabilidad selectiva:. Separa dos medios acuosos el externo del citosol. Mantiene el medio interno conuna elevada concentración molecular, además de una carga eléctrica interna de signo negativo.Las células requiere nutrientes del exterior y también necesitan eliminar sustancias de desecho. Han de mantener su medio interno estable, regulando la concentración interna para lo que transportan a través de su membrana,. Agua y solutos.
+procesos de ósmosis, se trata de una difusión especial. El agua pasa a través de membrana semipermeable, dela menos concentrada a la más concentrada. Al medio con mayorconcentración, se le llama hipertónico+transporte pasivo.-el paso de moléculas e iones a través de la membrana sin consumo de energía a favor de gradiente, de concentraciones y eléctrico, en las soluciones que contienen moléculas, iones que poseen cargas eléctricas.
1. Difusión simple: la sustancia a transportar debe ser soluble en la membrana. Es más rápido cuanto mayor sea el gradiente de concentraciones y en él no intervienen proteínas transportadoras O2, CO2, glucosa, aminoácidos

2. Difusión facilitada utiliza proteínas transportadoras: permeasas que se unen a las moléculas atransportar y las liberan de nuevo en el otro lado de lamembrana
3. Transporte activo. Se realiza contra gradiente y se requieren proteínas transportadoras específicas y un aporte de energía que se traduce en un consumo de ATP. Mantiene la diferencia de potencial que existe en todas las membranas en las que el interior está cargado negativamente con respecto al exterior. Esto es así por la bomba de sodio -potasio.
4. Transporte de macromoléculas: endocitosis y exocitosis.La célula no puede introducir en su interior sustancias de gran tamaño (macromoléculas, virus, bacterias...) sindañar a la M.P, por ello presentan mecanismos basados enla formación de vesículas membranosas en cuyo interior se sitúan las macromoléculas.Los procesos de introducción de macromoléculas en el interior de vesículas reciben el nombre deendocitosis. Por el contrario la expulsión de macromoléculas transportadas por vesículas recibe el nombre de exocitosis.La endocitosis se inicia cuando un mecanismo de control en la membrana que induce un sistema reticular de clatrina (proteína filamentosa) arrastra dicho sectormembranoso hacia el interior del citoplasma formando unavesícula. Posteriormente las moléculas abandonan la vesícula y vuelven a la membrana.Algunas moléculas al unirse con receptores específicos de la membrana inducen la formación de vesículas que lasengloban.Puede ser de dos tipos:Pinocitosis cuando la célula ingiere líquidos y sustancias disueltas que almacena en pequeñas vesículas.Fagocitosis cuando ingiere grandes partículas, microorganismos en el interior de grandes vesículas o endosomas. Es realizada para obtener alimento del exterior.-Producir, modular y conservar gradientes

electroquímicos entre uno y otro lado-Recibir y transmitir señales-Controlar el desarrollo y división celular-Permitir disposición adecuada de moléculas funcionales-Delimitar compartimentos intracelulares
Diferenciaciones de la membrana plasmática
1. Microvellosidades2. Interdigitaciones laterales3. Invaginaciones basales4. Complejos de unión:
1. Desmosomas2. Uniones estrechas3. Hendiduras o gap-junction
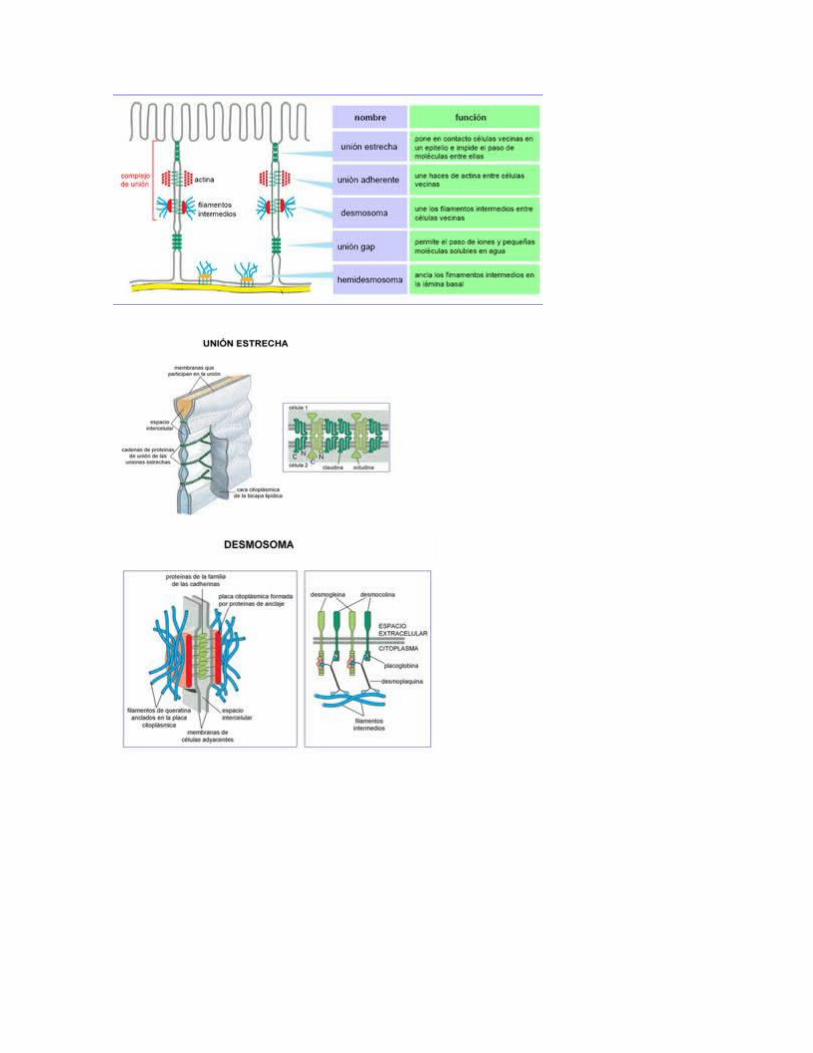
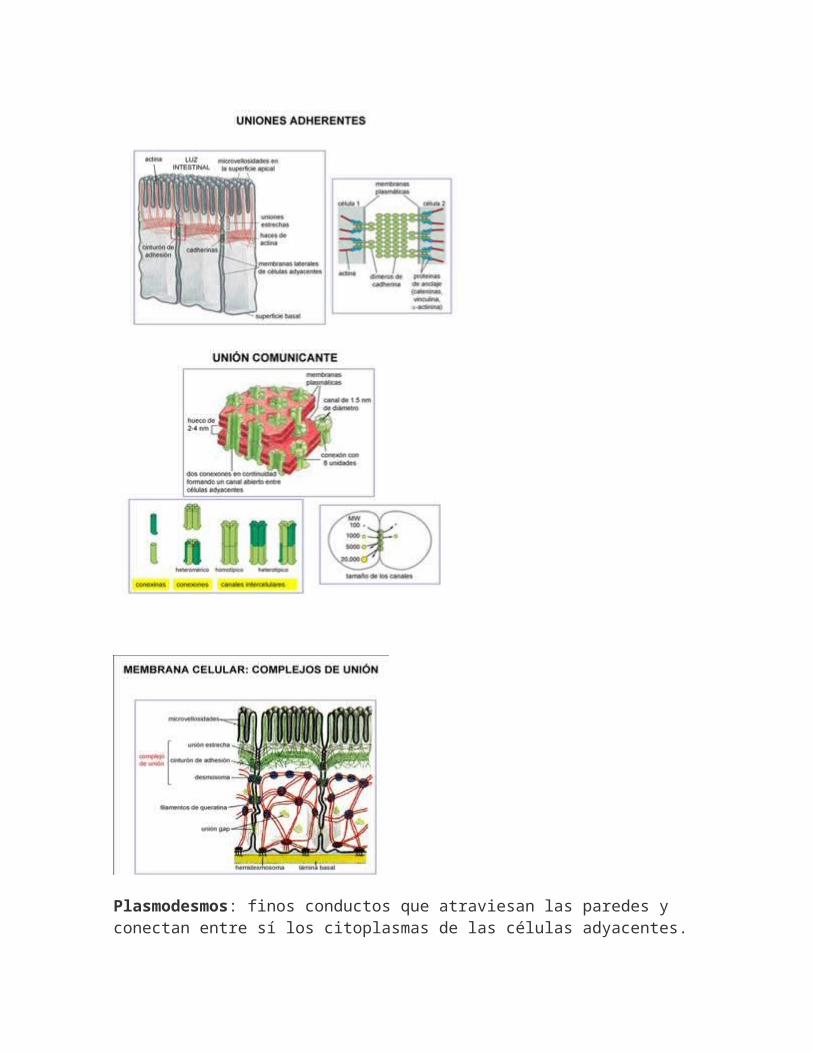
Plasmodesmos: finos conductos que atraviesan las paredes y conectan entre sí los citoplasmas de las células adyacentes.

Están rodeados por una membrana común y presentan un túbulo de retículo endoplasmático, el desmotúbulo.FUNCIONES:-Exosqueleto que protege y da forma.-Unen las células entre sí.-Les permite vivir en medio hipotónico.-La lignificación permite porte erecto y formación de vasos.-Cutinización y suberificación impermeabiliza la superficie.-Barrera para el paso de sustancias.
Punteaduras, que son adelgazamientos de las paredes, Constan de cavidad y membrana formada por lámina y pared primaria muydelgada. Son pares.
GLUCOCÁLIX.-Zona periférica rica en glúcidos de la superficie de células eucariotas (50 nm). Formada por cadenas de oligosacáridos de glicolípidos y glicoproteínas de membrana. También contiene glicoproteínas que han sido segregadas y luego absorbidas sobre la superficie.Funciones:-Protege la superficie celular del daño mecánico y químico.-Reconocimiento celular: la complejidad de oligosacáridos y el hecho de que se encuentren en la superficie sugiere que son los principales marcadores de identidad que actúan en procesos de reconocimiento: -Comunicaciones intercelulares. -Funcionamiento del sistema inmunitario.(rechazo,alergias) -La capacidad patogénica de agentes infecciosos.(amigdalitis, gastroenteritis...)
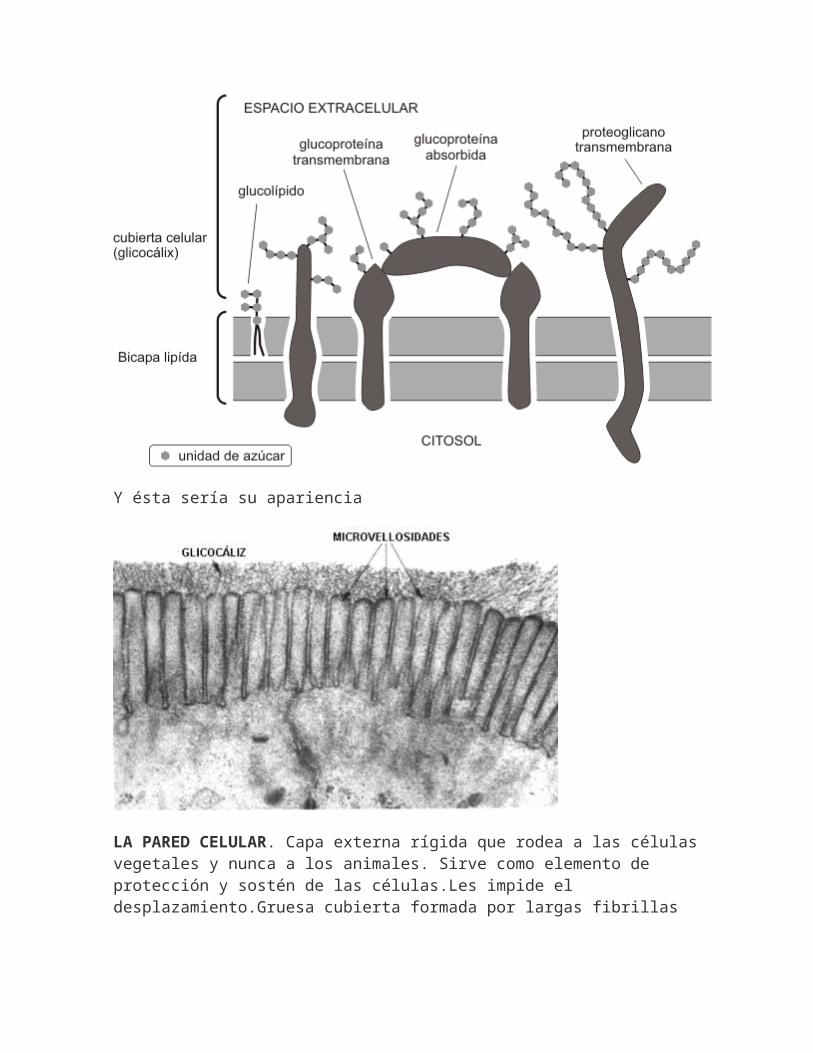
Y ésta sería su apariencia
LA PARED CELULAR. Capa externa rígida que rodea a las célulasvegetales y nunca a los animales. Sirve como elemento de protección y sostén de las células.Les impide el desplazamiento.Gruesa cubierta formada por largas fibrillas

de celulosa unidas ente sí por una matriz de polisacáridos y proteínas.
Composición-Celulosa, glucosa unida por enlaces b(1®4). Entre 60-70 cadenas unidas por puentes de H se agrupan en microfibrillas.Las microfibrillas están englobadas en una matriz entrecruzada de moleculas no celulósicas: polisacáridos, hemicelulosa y pectinas, glicoproteínas, minerales (Ca) y agua. Las células pueden sufrir modificaciones: lignina, paradar rigidez en soporte. La cutina en paredes epidérmicas y sobre ellas se depositan ceras. La suberina en pared secundaria.Capas-Lámina media: formada por pectinas. -Pared primaria: formada por microfibrillas de celulosa en forma reticular y matriz de hemicelulosa, pectinas y glicoproteinas, siendo el 60% agua. La disposición hyace que esta pared se pueda estira y expandir, conforme la célula aumenta de tamaño. Las células que se dividen continuamente sólo contienen paredes primarias-Pared secundaria. Se inicia una vez completado el crecimiento celular. Es la pared más gruesa y está formada por varias capas. La celulosa es más abundante que en la Primaria y están orientas paralelamente a la matriz formada por hemicelulosa, conteniendo un 30% de agua. Es rígida y difícilmente deformable, por lo que impide el crecimiento. Consta de varias capas y suele esta formada por suberina

Origen . Las capas van apareciendo por aposición de sustancias del exterior al interior, siendo por eso las más externas las más antiguas. La primera en aparecer es la lámina media, que es gelationosa y está formada por pectina apartir de las cvesículas del Ap de Golgi)s. Después varias láminas que forman la capa secundaria, que es el verdadero esqueleto, formado por un armazón de celulosa, que puede sufrir modificaciones, como la mineralización si se impregna de sales de calcio o silicio, lignificación, suberificación, cutinización...
EspecializacionesLa pared primaria y la lámina media son permeables a gases y pequeñas moléculas hidrosolubles. El paso de sustancias está favorecido por la presencia de:-Punteaduras, que son adelgazamientos de las paredes, Constande cavidad y membrana formada por lámina y pared primaria muydelgada. Son pares.-Plasmodesmos: finos conductos que atraviesan las paredes y conectan entre sí los citoplasmas de las células adyacentes. Están rodeados por una membrana común y presentan un túbulo de retículo endoplasmático, el desmotúbulo.FUNCIONES:-Exosqueleto que protege y da forma.-Unen las células entre sí.-Les permite vivir en medio hipotónico.-La lignificación permite porte erecto y formación de vasos.-Cutinización y suberificación impermeabiliza la superficie.-Barrera para el paso de sustancias.
GLUCOCÁLIX.-Zona periférica rica en glúcidos de la superficie de células eucariotas (50 nm). Formada por cadenas de oligosacáridos de glicolípidos y glicoproteínas de membrana. También contiene glicoproteínas que han sido segregadas y luego absorbidas sobre la superficie.

Funciones:-Protege la superficie celular del daño mecánico y químico.-Reconocimiento celular: la complejidad de oligosacáridos y el hecho de que se encuentren en la superficie sugiere que son los principales marcadores de identidad que actúan en procesos de reconocimiento:-Comunicaciones intercelulares.-Funcionamiento del sistema inmunitario.(rechazo, alergias)-La capacidad patogénica de agentes infecciosos.(amigdalitis,gastroenteritis...)
CITOPLASMA; espacio celular comprendido entre la membrana citoplasmática y la envoltura nuclear. Constituida por el citosol y orgánulos.El citosol es el medio interno en el que se encuentran los orgánulos celulares y el núcleo.Representa el 55% del volumen celular y es un medio acuoso, entre un 75-85% de agua con gran cantidad de enzimas, nucleótidos, Aa...Es un medio dinámico en el que se producen muchas de las reacciones metabólicas de las células. En él tienen lugar movimientos internos que son los responsables del movimiento de los orgánulos.Dispersas en el citoplasma se encuentran estructuras granulares sin membrana: ribosomas e inclusiones, que son acúmulos de sustancias como gránulos de glucógeno o gotas lipídicas. Posee una consistencia de gel y lleva disueltos iones, Aa, monosacáridos, nucleótidos, coenzimas y metabolitos, RENA y proteínas. También está formado por un conjunto de filamentos proteícos que constituyen el citoesqueleto.
CITOESQUELETO.-Está formado por complejas redes de filamentos proteínicos, que son los responsables de la formade las células, de su movimiento y de su organización interna. Los tres tipos de filamentos están conectados entre sí, lo que les permite coordinar sus funciones.
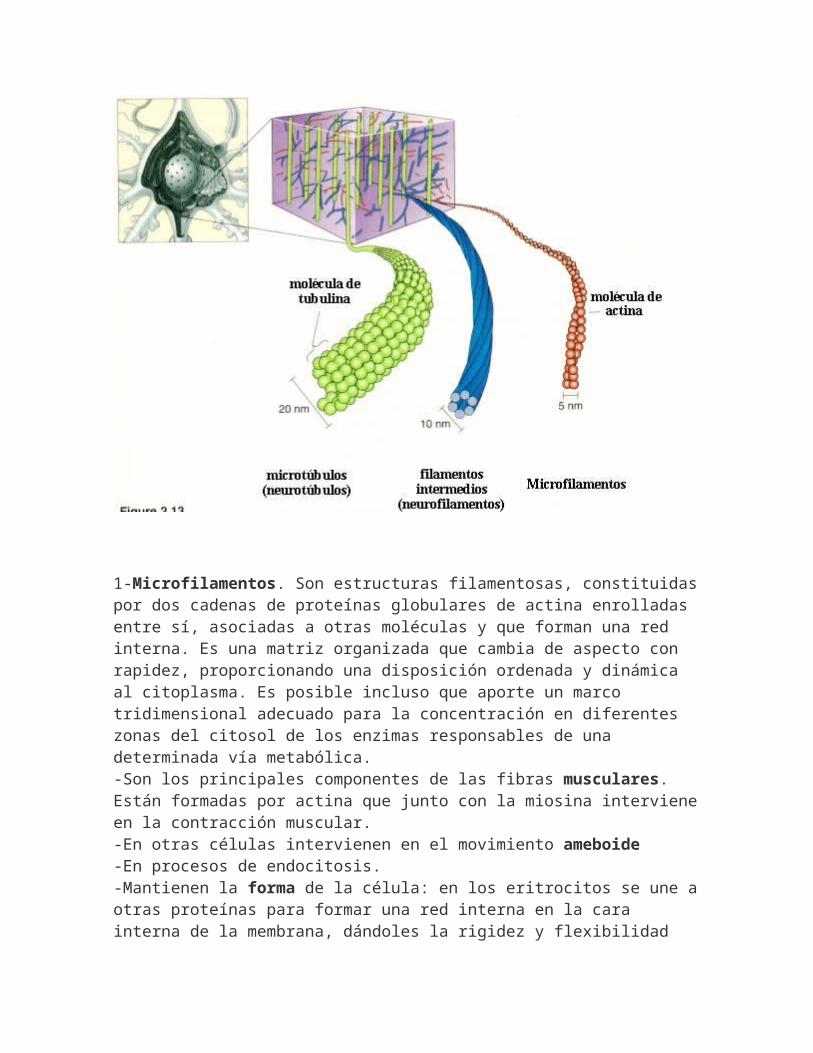
1-Microfilamentos. Son estructuras filamentosas, constituidaspor dos cadenas de proteínas globulares de actina enrolladas entre sí, asociadas a otras moléculas y que forman una red interna. Es una matriz organizada que cambia de aspecto con rapidez, proporcionando una disposición ordenada y dinámica al citoplasma. Es posible incluso que aporte un marco tridimensional adecuado para la concentración en diferentes zonas del citosol de los enzimas responsables de una determinada vía metabólica.-Son los principales componentes de las fibras musculares. Están formadas por actina que junto con la miosina intervieneen la contracción muscular.-En otras células intervienen en el movimiento ameboide -En procesos de endocitosis.-Mantienen la forma de la célula: en los eritrocitos se une aotras proteínas para formar una red interna en la cara interna de la membrana, dándoles la rigidez y flexibilidad

necesarias para atravesar los capilares.-Permite la estabilidad de prolongaciones citoplasmáticas: las microvellosidades-Forman el anillo contráctil en el división.
2-Microtúbulos. Son filamentos tubulares formadospor proteínas, la tubulina, por dos monómeros distintos:α y. Los dímeros se unen formando un protofilamento. Consta de 13 protofilamentos paralelos que forman un cilindro hueco. Sólo se encuentran en células eucariotas y podrían proceder de microorganismos flagelados en simbiosis Se originan a partirde la centrosfera en células animales. Se dividen en:-Lábiles; los del huso acromático, pseudópodos, transporte deneurotransmisores...-Estables: los de centriolos, flagelos y cilios.FUNCIONES:-Principales generadores del movimiento de cilios y flagelos.-Dirigen el transporte de orgánulos en el citoplasma, yha queactúan como guías. (se ha visto en mitocondrias)-Constituyen el huso mitótico.-Determinan la forma y polaridad de la célula.-Disponen el retículo endoplasmático y Golgi en lugares adecuados.-Intervienen en la organización de todos los filamentos del citoesqueleto.

3.-Filamentos intermedios. Fibras proteícas resistentes, parecidas a cuerdas. Muy abundantes en células sometidas a tensiones mecánicas, como epitelios, axones y músculo.Distintos tipos: queratina –proporcionan fuerza mecánica a epitelios.-Neurofilamentos de células nerviosas.-Filamentos de vimentina en conjuntivo y vasos.-Filamentos de lámina nuclear.
CENTRIOLOS.Se presentan de dos en dos orientados perpendicularmente entre sí y que reciben el nombre de diplosomas, en el centro de la célula, que se denomina centrosoma, localizado próximo alnúcleo y que es considerado un centro formador de microtúbulos.El diplosoma se encuentra en el interior de un material denso, que se denomina centrosfera dónde se organizan y de dónde parten radialmente unos microtúbulos que se denominan aster.Cada centriolo consta de nueve grupos de tres microtúbulos que se disponen formando un cilindro. Son orgánulos exclusivos de células animales. Proteínas que mantienen su estructura cilíndrica. Están relacionados con la organizaciónde los microtúbulos del citoesqueleto, en el transporte citoplasmático y de la disposición de los filamentos del husodurante la mitosis.

CILIOS Y FLAGELOS.Son prolongaciones citoplasmáticas dotadas de movimiento, presentan un eje central rodeado por una membrana que es prolongación de la plasmática.CILIOS: con diámetro de 0,2μ y longitud, 5-10μ. Constituidos por 4 zonas: par de microtúbulos en la parte central encerrados en una vaina y rodeados por un conjunto cilíndricode 9 pares de microtúbulos paralelos que se unen al par central mediante ejes radiales. En el cuerpo basal carecen del par de microtúbulos centrales. Permite el desplazamiento celular o crea turbulencias. Los flagelos únicamente difierenen sus dimensiones, longitud de 100μ.
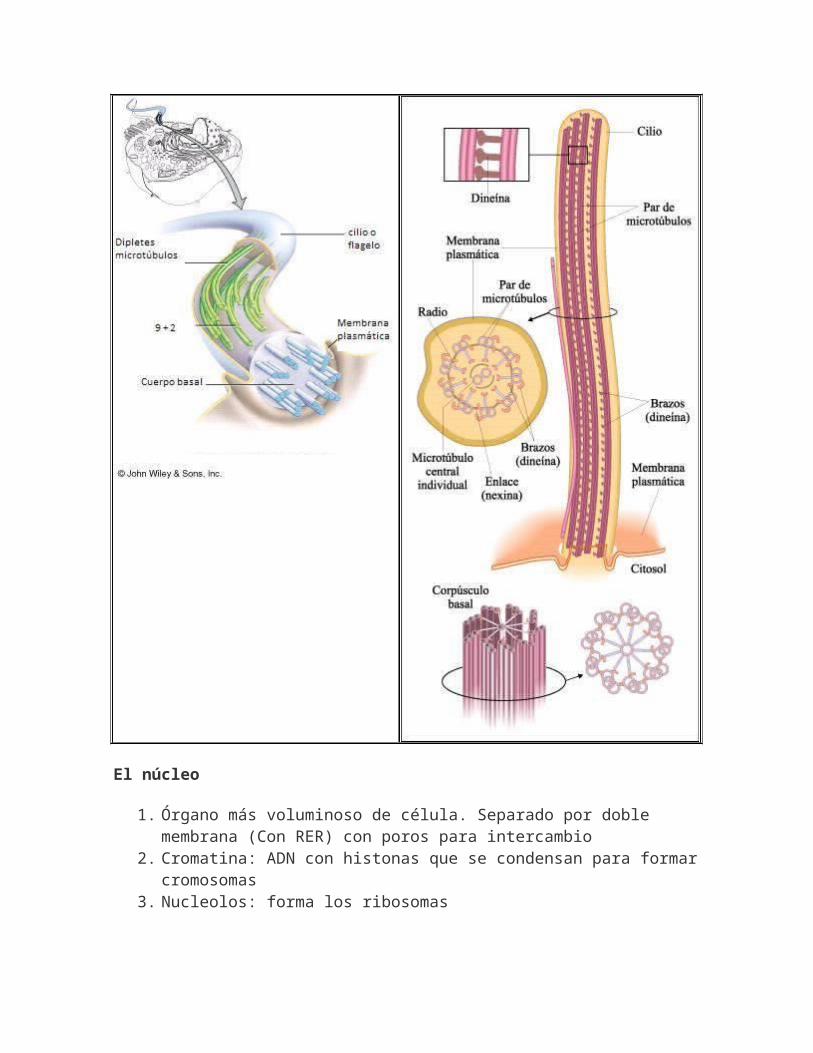
El núcleo
1. Órgano más voluminoso de célula. Separado por doble membrana (Con RER) con poros para intercambio
2. Cromatina: ADN con histonas que se condensan para formarcromosomas
3. Nucleolos: forma los ribosomas
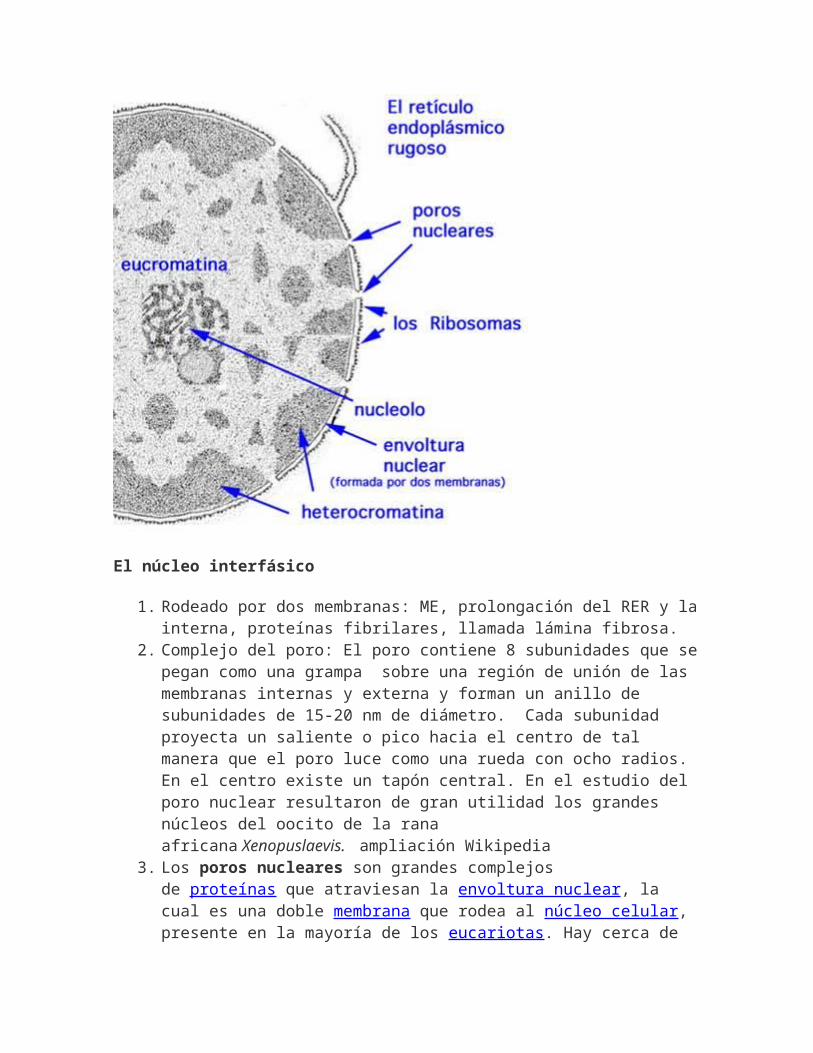
El núcleo interfásico
1. Rodeado por dos membranas: ME, prolongación del RER y lainterna, proteínas fibrilares, llamada lámina fibrosa.
2. Complejo del poro: El poro contiene 8 subunidades que sepegan como una grampa sobre una región de unión de las membranas internas y externa y forman un anillo de subunidades de 15-20 nm de diámetro. Cada subunidad proyecta un saliente o pico hacia el centro de tal manera que el poro luce como una rueda con ocho radios. En el centro existe un tapón central. En el estudio del poro nuclear resultaron de gran utilidad los grandes núcleos del oocito de la rana africana Xenopuslaevis. ampliación Wikipedia
3. Los poros nucleares son grandes complejos de proteínas que atraviesan la envoltura nuclear, la cual es una doble membrana que rodea al núcleo celular, presente en la mayoría de los eucariotas. Hay cerca de

2000 complejos de poro en la envoltura nuclear en la célula de un vertebrado, pero varía dependiendo del número de transcripciones de la célula.
4. Las proteínas que forman los complejos de poro nuclearesson conocidas como nucleoporinas. Cerca de la mitad de las nucleoporinas contienen comúnmente una estructura terciaria alfa solenoide o beta hélice, o en algunos casos ambas como dominios proteicos separados. La otra mitad muestra características estructurales típicas de proteínas "nativamente no dobladas", por ejemplo son proteínas altamente flexibles que carecen de estructura secundaria ordenada[1] . Estas proteínas desordenadas son las nucleoporinas FG, llamadas así por su secuencia aminoacídica que contiene varias repeticiones del péptido fenilalanina-glicina [2] .
5. Los poros nucleares permiten el transporte de moléculas solubles en agua a través de la envoltura nuclear. Este transporte incluye el movimiento de ARN y ribosomas desde el núcleo al citoplasma, y movimiento de proteínas (tales como ADN polimerasa y lamininas), carbohidratos, moléculas de señal y lípidos hacia el núcleo. Es notable que el complejo de poro nuclear (CPN o NPC en inglés) puede conducir activamente 1000 translocaciones por complejo por segundo. Aunque las moléculas pequeñas pasan por difusión simple a través de los poros, las moléculasde mayor tamaño pueden ser reconocidas mediante secuencias de señal específicas y luego difundidas con la ayuda de las nucleoporinas hacia o desde el núcleo. Esto es conocido como el ciclo RAN. Cada una de las ochosubunidades proteicas que rodean el poro verdadero (el anillo externo) proyecta una proteína con forma de radiohacía el canal del poro. El centro del poro muchas vecesparece que tuviera una estructura parecida a un tapón. Aún no se sabe sí esto corresponde a un tapón verdadero o es simplemente carga atrapada durante el transito.

1. Funciones:1. separar al citoplasma del nucleoplasma,2. Mantener separados los procesos metabólicos de
ambos medios.3. Regula el intercambio de sustancias a través de los
poros4. La lámina nuclear permite la unión con las fibras
de ADN para formar los cromosomas.
Nucleoplasma:
1. dispersión coloidal en forma de gel con proteínas relacionadas con la síntesis y empaquetamiento de los ácidos nucleicos.
2. Nucleótidos, ARN, ADN, agua e iones.
Red de proteínas fibrilares similar a las del citoplasma. Su función es ser el seno en el que se produce la síntesis de ARN diferentes y la síntesis del ADN nuclear. Además, evita la formación de nudos en la cromatina.
1. Nucleolo/S: ADN, RNr y enzimas: ribosomas2. Cromatina: ADN + Histonas3. Proporciona la información genética necesaria para que
los orgánulos celulares puedan realizar la transcripcióny síntesis de proteínas; también conservan y transmiten la información genética contenida en el ADN, duplicando el ADN en la reproducción celular.

1. Eucromatina: poco densa y genéticamente activa2. Heterocromatina: ADN empaquetado e inactivo
(1) Hebra simple de ADN. (2) Hebra de cromatina (ADN con histonas, "cuenta de collar"). (3)Cromatina durante la interfase con centrómero. (4) Cromatina condensada durantela profase (Dos copias de ADN están presentes). (5) Cromosoma durante la metafase.
De Wikipedia
Grado empaquetamiento ADN
Los cromosomas
1. Estructuras en la división del núcleo2. ADN más histonas son cromatina condensada.

3. Su número es constante en todas las células de un individuo pero varía según las especies. Esta cantidad de cromosomas se denomina númerodiploide y se simboliza como 2n. Cuando se examina la longitud de tales cromosomas y la situación del centrómero surge el segundo rasgo general: para cada cromosoma con una longitud y una posición del centrómero determinada existe otro cromosoma con rasgos idénticos, o sea, casi todos los cromosomas se encuentran formando parejas. Losmiembros de cada par se denominan cromosomas homólogos.
4. Un cromosoma está formado por dos cromátidas unidas por un centrómero, que permite clasificarlos según la posición del centrómero a lo largo del cromosoma.
5. El cromosoma puede presentar secundarias que se producenen los brazos y originan satélites. Son los telómeros. La palabra telómero procede del griego telos, "final" y meros, "parte". Los telómeros son los extremos de los cromosomas. Son regiones de ADN no codificante, altamente repetitivas, cuya función principal es la estabilidad estructural de los cromosomas en las célulaseucariotas, la división celular y el tiempo de vida de las estirpes celulares. Además están involucradas en enfermedades tan importantes como el cáncer.
6. Alrededor del centrómero, cinetocoro, que organiza los microtúbulos que facilitarán la separación de las dos cromátidas en la división celular.
7.
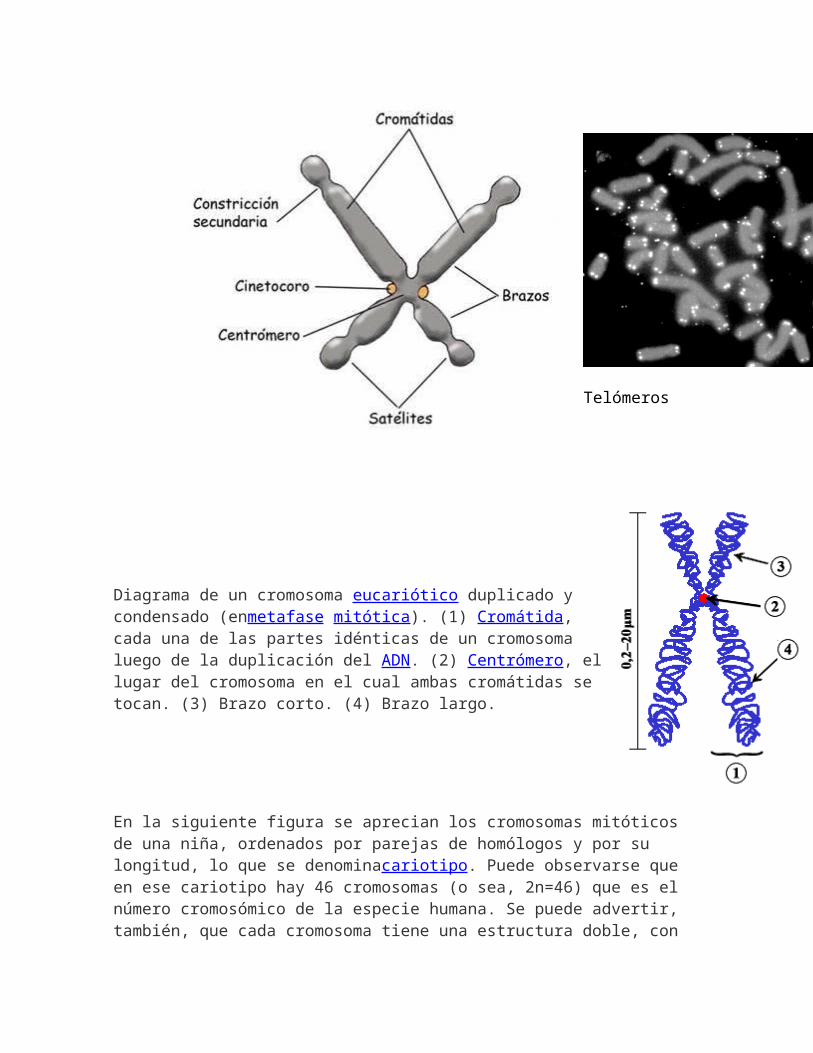
Telómeros
Diagrama de un cromosoma eucariótico duplicado y condensado (enmetafase mitótica). (1) Cromátida, cada una de las partes idénticas de un cromosoma luego de la duplicación del ADN. (2) Centrómero, el lugar del cromosoma en el cual ambas cromátidas se tocan. (3) Brazo corto. (4) Brazo largo.
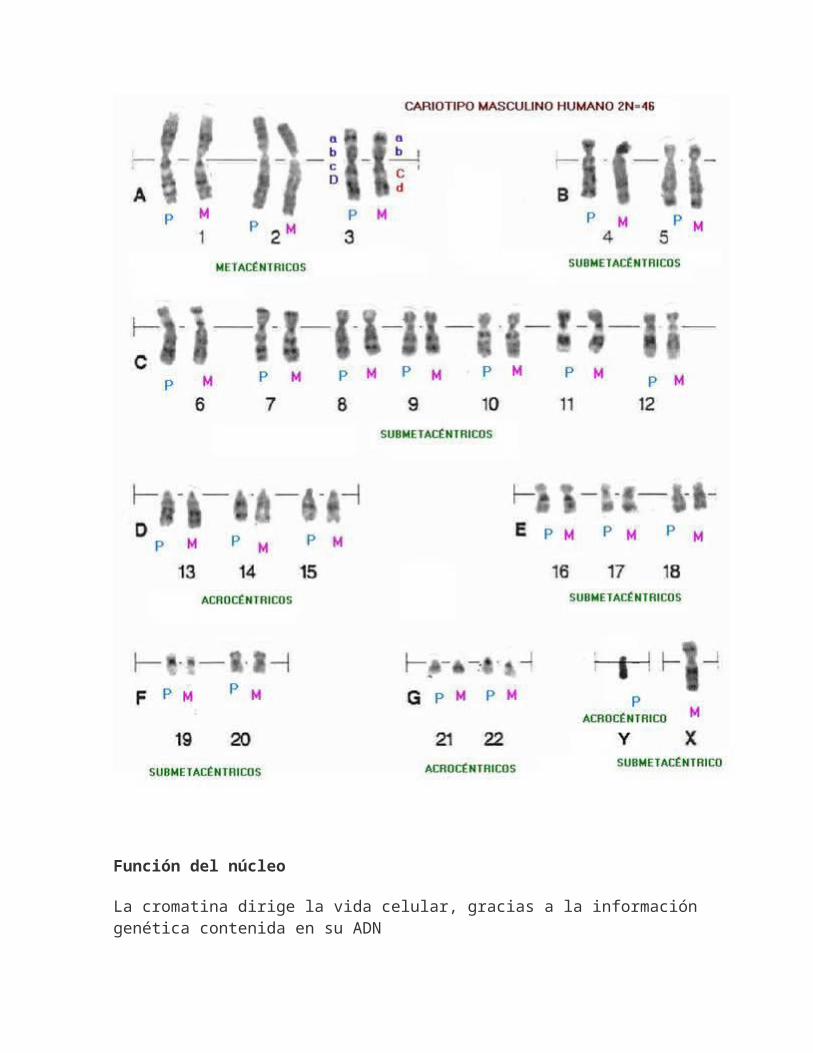
En la siguiente figura se aprecian los cromosomas mitóticos de una niña, ordenados por parejas de homólogos y por su longitud, lo que se denominacariotipo. Puede observarse que en ese cariotipo hay 46 cromosomas (o sea, 2n=46) que es el número cromosómico de la especie humana. Se puede advertir, también, que cada cromosoma tiene una estructura doble, con

dos cromátidas hermanas, paralelas entre sí y unidas por un único centrómero. Durante la mitosis las cromátidas hermanas,que son idénticas, se separan una de otra hacia dos nuevas células. Las parejas de cromosomas homólogos que se observan en la imagen tienen, además, una semejanza genética fundamental: presentan los mismos genes situados en los mismos lugares a lo largo del cromosoma (tales lugares se denominan locus o loci en plural). Esto indica que cada miembro del par de homólogos lleva información genética para las mismas características del organismo. En organismos con reproducción sexual, uno de los miembros del par de cromosomas homólogos proviene de la madre (a través del óvulo) y el otro del padre (a través del espermatozoide).Cada organismo diploide tiene dos copias de cada uno de los genes, cada una ubicada en uno de los cromosomas homólogos, excepción hecha de los cromosomas sexuales, que en muchas especies no tienen el mismo tamaño, igual situación del centrómero, la misma proporción entre los brazos o, incluso, los mismos loci. En la imagen puede observarse, por ejemplo, que el cromosoma Y (que determina el sexo masculino en humanos) es de menor tamaño y carece de la mayoría de los loci que se encuentran en el cromosoma X
Grado de empaquetamiento
Forma de los cromosomas
La forma de los cromosomas es para todas las células somáticas constante y característica de cada especie. La forma depende fundamentalmente de las constricciones que presente el cromosoma y de su localización en la cromátida.
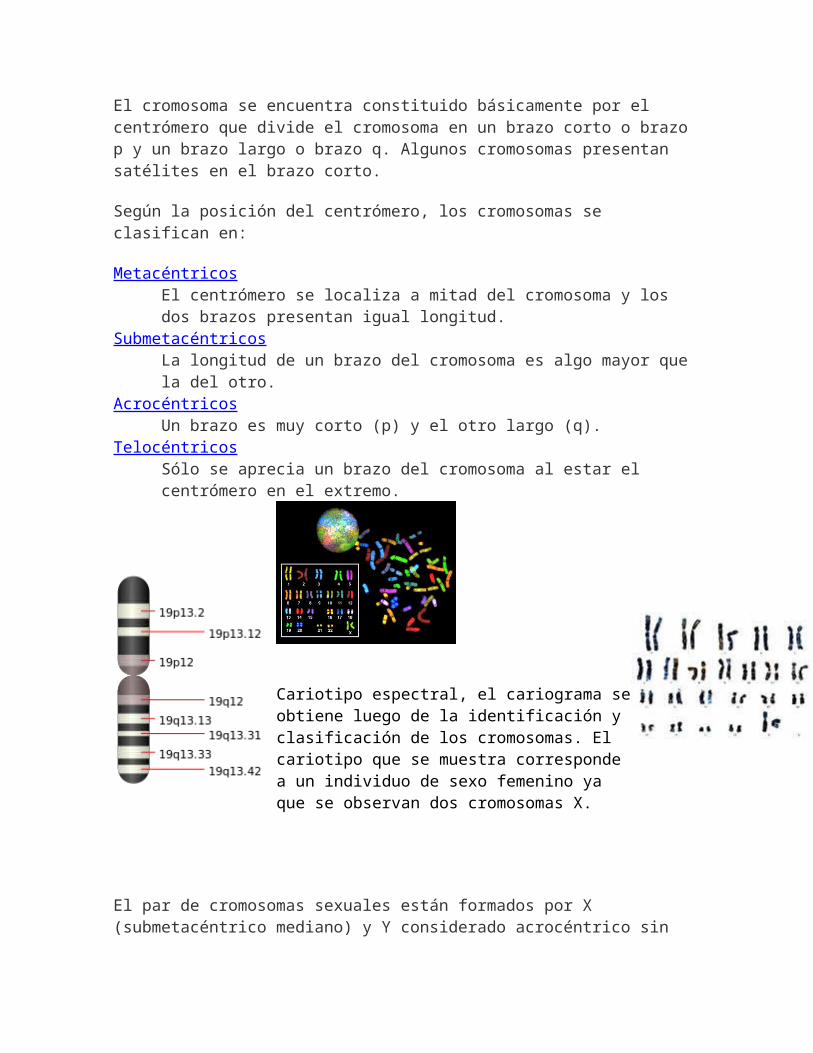
El cromosoma se encuentra constituido básicamente por el centrómero que divide el cromosoma en un brazo corto o brazo p y un brazo largo o brazo q. Algunos cromosomas presentan satélites en el brazo corto.
Según la posición del centrómero, los cromosomas se clasifican en:
MetacéntricosEl centrómero se localiza a mitad del cromosoma y los dos brazos presentan igual longitud.
SubmetacéntricosLa longitud de un brazo del cromosoma es algo mayor que la del otro.
AcrocéntricosUn brazo es muy corto (p) y el otro largo (q).
TelocéntricosSólo se aprecia un brazo del cromosoma al estar el centrómero en el extremo.
Cariotipo espectral, el cariograma seobtiene luego de la identificación y clasificación de los cromosomas. El cariotipo que se muestra corresponde a un individuo de sexo femenino ya que se observan dos cromosomas X.
El par de cromosomas sexuales están formados por X (submetacéntrico mediano) y Y considerado acrocéntrico sin
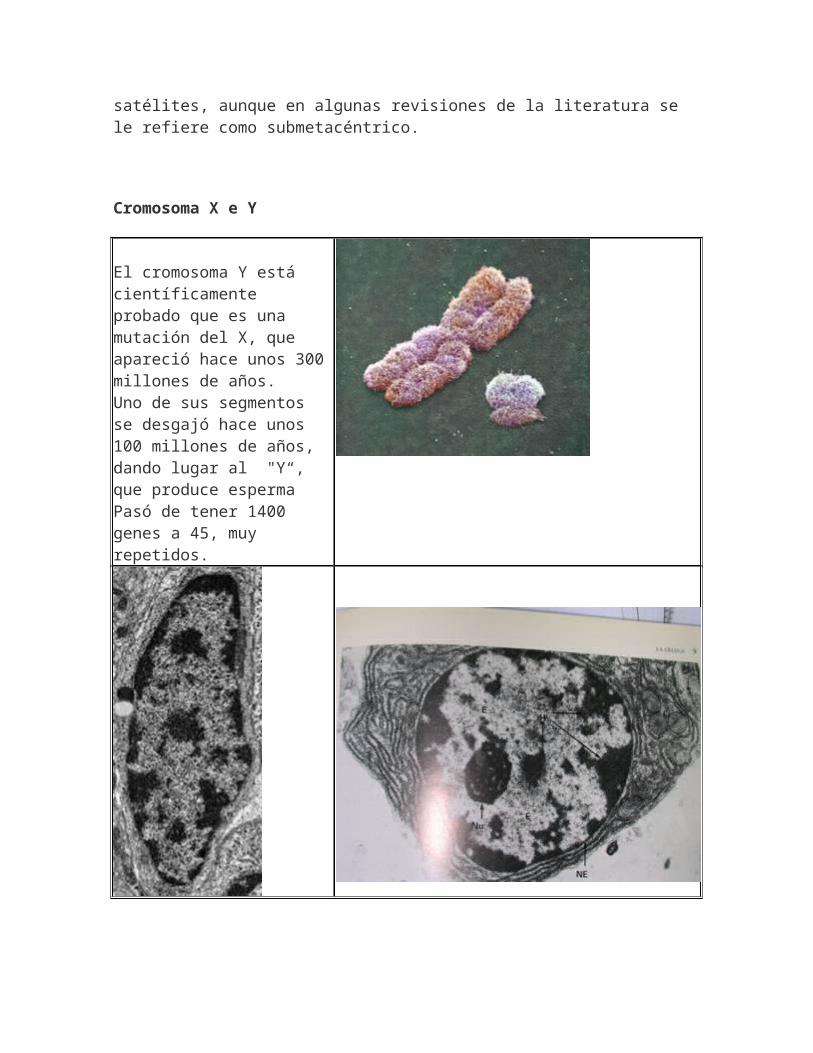
satélites, aunque en algunas revisiones de la literatura se le refiere como submetacéntrico.
Cromosoma X e Y
El cromosoma Y está científicamente probado que es una mutación del X, que apareció hace unos 300millones de años. Uno de sus segmentos se desgajó hace unos 100 millones de años, dando lugar al "Y“, que produce esperma Pasó de tener 1400 genes a 45, muy repetidos.

Función del núcleo
La cromatina dirige la vida celular, gracias a la informacióngenética contenida en su ADN
Debe duplicar ese material y transmititrl a las células hijasen cda división.
El nucleolo es el lugar de síntesis del ARNr y de ribosomas.
El nucleoplasma aporta la materia prima y los enzimas necesarios para todos estos procesos
La membrana, gracias al complejo del poro, regula el intercambio de sustancias con el citoplasma.
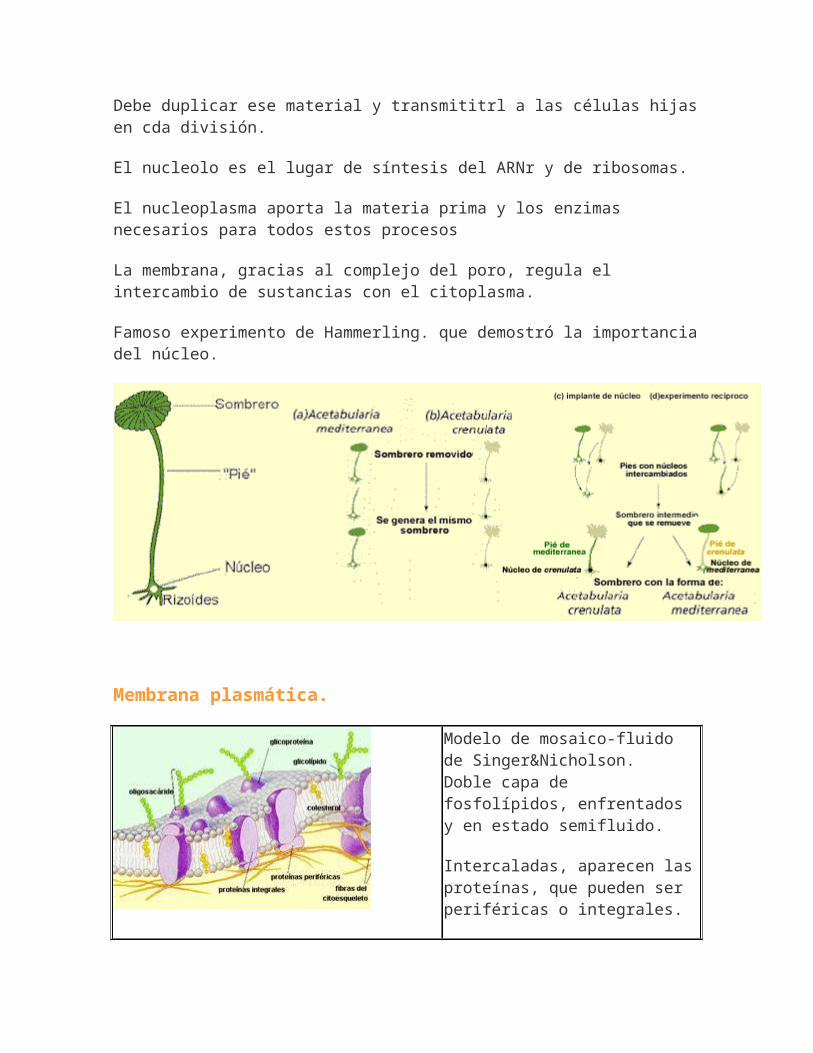
Famoso experimento de Hammerling. que demostró la importanciadel núcleo.
Membrana plasmática.
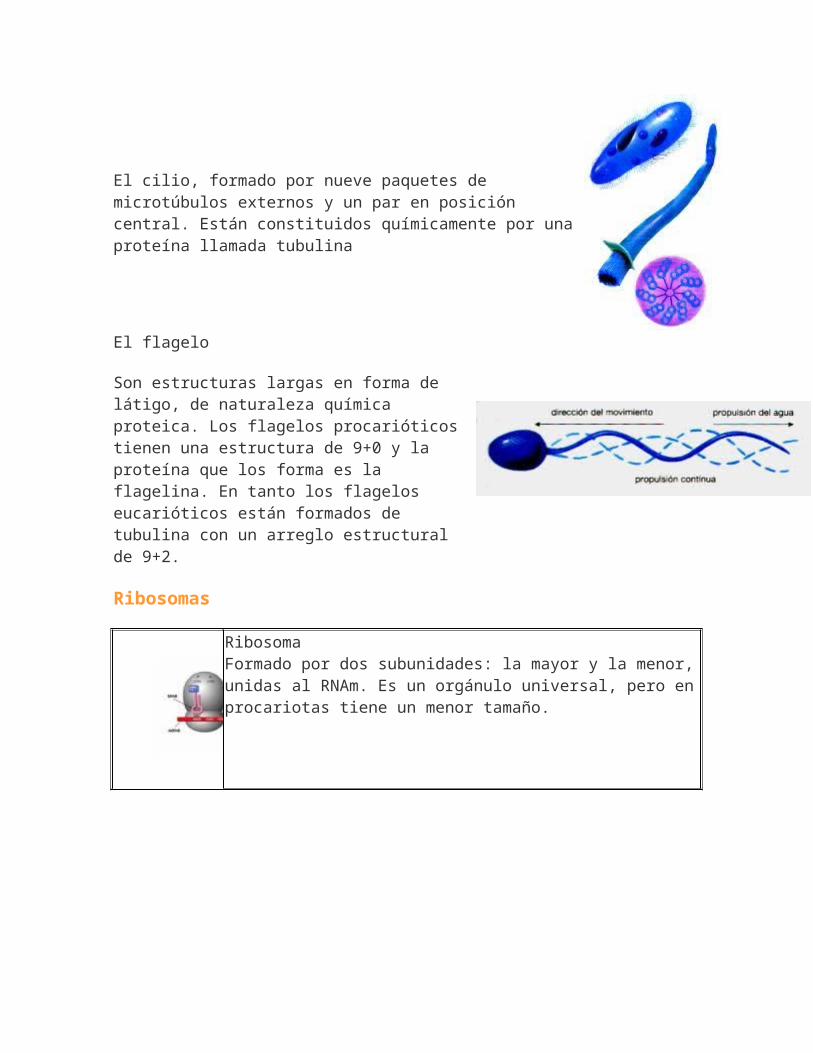
Modelo de mosaico-fluido de Singer&Nicholson.Doble capa de fosfolípidos, enfrentados y en estado semifluido.
Intercaladas, aparecen lasproteínas, que pueden ser periféricas o integrales.

Asociados a ambos, los glúcidos
Función
Aislamiento y relación: permite el paso de determinadas sustancias. Transporte. Reconocimiento celular.
GLICOCÁLIXZona periférica rica en glúcidos de la superficie de células eucariotas (50 nm). Formada por cadenas de oligosacáridos de glicolípidos y glicoproteínas de membrana. También contiene glicoproteínas que han sido segregadas y luego absorbidas sobre la superficie.
-Protege la superficie celular del daño mecánico y químico.
-Reconocimiento celular: la complejidad de oligosacáridos y el hecho de que se encuentren en la superficie sugiere que son los principales marcadores de identidad que actúan en procesos de reconocimiento
Citoplasma
Es el espacio celular comprendido entre la membrana citoplasmática y la envoltura nuclear. Constituida por el citosol y orgánulos.
El citosol es el medio interno en el que se encuentran los orgánulos celulares y el núcleo.Representa el 55% del volumen celular y es un medio acuoso, entre un 75-85% de agua con gran cantidad de enzimas, nucleótidos, Aa...Es un medio dinámico en el que se producen muchas de las

reacciones metabólicas de las células. En él tienen lugar movimientos internos que son los responsables del movimiento de los orgánulos.
Dispersas en el citoplasma se encuentran estructuras granulares sin membrana: ribosomas e inclusiones, que son acúmulos de sustancias como gránulos de glucógeno o gotas lipídicas. También flota en él el citoesqueleto.
Citoesqueleto
Está formado por complejas redes de filamentos proteínicos, que son los responsables de la forma de las células, de su movimiento y de su organización interna. Son tres tipos de filamentos, conectados entre sí, lo que les permite coordinarsus funciones.
1-Microfilamentos.
2-Microtúbulos.
3.-Filamentos intermedios.
LA PARED CELULAR.
Capa externa rígida que rodea a las células vegetales y nunca a los animales.
Sirve como elemento de protección y sostén de las células: es impide el desplazamiento.
Gruesa cubierta formada por largas fibrillas de celulosa unidas ente sí por

una matriz de polisacáridos y proteínas. Las células pueden sufrir modificaciones:lignina, para dar rigidez en soporte. La cutina en paredes epidérmicas y sobre ellas se depositan ceras. La suberina en pared secundaria.
CENTRIOLOS. Se presentan de dos en dos orientados perpendicularmente entre sí y que reciben el nombre de diplosomas, en el centro de la célula, que se denomina centrosoma, localizado próximo al núcleo y que es considerado un centro formador de microtúbulos.Está constituido por un par centriolos, cada uno construido por unanillo de nueve grupos de microtubulos. Sólo aparecen en células animales.
El centrosoma: produce los microtúbulos Durante la división, cada centrosoma se mueve a lugares opuestos del núcleo, y de cada uno crece un haz de tubulos que separarán los cromosomas en las dos celulas hermanas. Las celulas de las plantas tienen centrosomas pero sin centriolos.
CILIOS Y FLAGELOS.
Son prolongaciones citoplasmáticas dotadas de movimiento. Presentan un eje central rodeado por una membranaque es prolongación de la plasmática. Son mecanismos de locomoción y en el caso de los cilios, de captura de alimentos.
El cilio, formado por nueve paquetes de microtúbulos externos y un par en posición central. Están constituidos químicamente por una proteína llamada tubulina
El flagelo
Son estructuras largas en forma de látigo, de naturaleza química proteica. Los flagelos procarióticos tienen una estructura de 9+0 y la proteína que los forma es la flagelina. En tanto los flagelos eucarióticos están formados de tubulina con un arreglo estructural de 9+2.
Ribosomas
RibosomaFormado por dos subunidades: la mayor y la menor,unidas al RNAm. Es un orgánulo universal, pero enprocariotas tiene un menor tamaño.

Función.-La síntesis de proteínas. Los aminoácidos son transportados por el RNA de transferencia (tRNA), específico para cada uno de ellos, y son llevados hasta el RNA mensajero(mRNA), dónde se aparean el codón de éste y el anticodón del RNA de transferencia, por complementariedad de bases, y de ésta forma se sitúan en la posición que les corresponde
Orgánulos membranosos.
RETÍCULO ENDOPLASMÁTICO
Conjunto de membranas que delimitan un espacio interno, que es el lumen, que unas veces aparece fragmentado en forma de cavidades planas interconectadas y otras en forma de cisternas, túbulos y vesículas, aunque se cree que existe unaúnica lámina membranosa y continua que envuelve el lumen, como un saco replegado una y otra vez. Las membranas del retículo presentan asimetría respecto a los enzimas prinicipales en sus dos caras
Existe una zona del RE que contiene ribosomas asociados a su cara, que es el rugoso y que continúa con la membrana externa
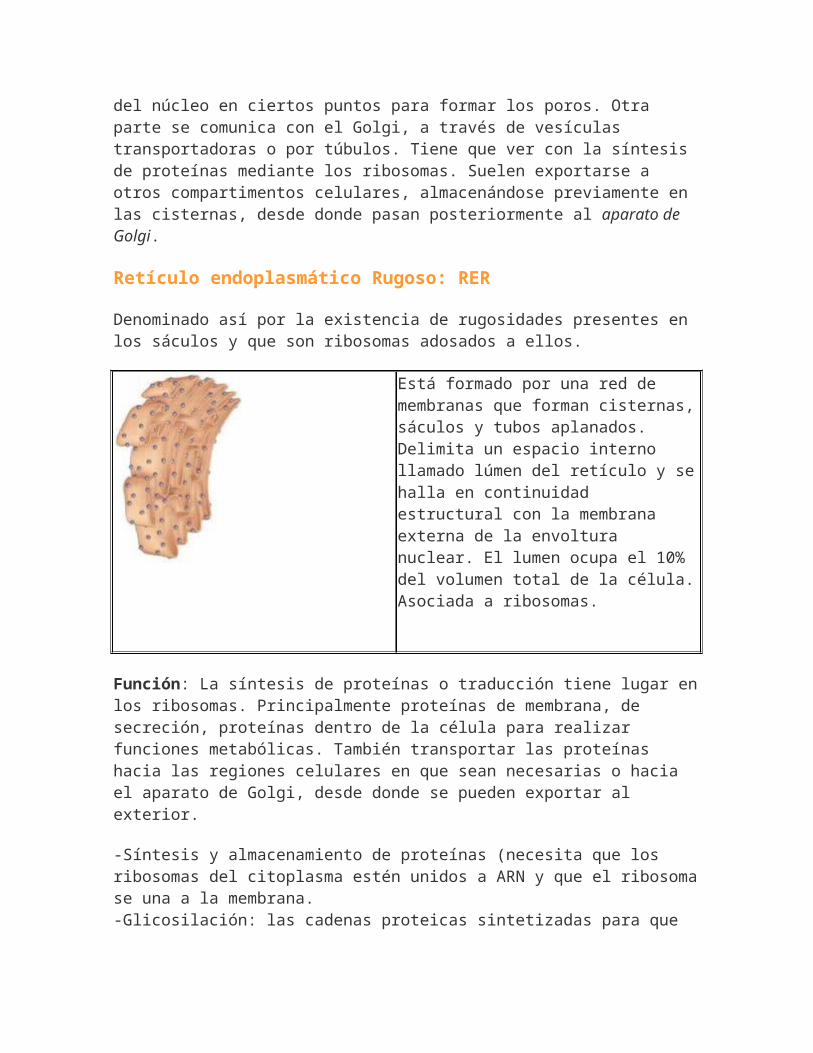
del núcleo en ciertos puntos para formar los poros. Otra parte se comunica con el Golgi, a través de vesículas transportadoras o por túbulos. Tiene que ver con la síntesis de proteínas mediante los ribosomas. Suelen exportarse a otros compartimentos celulares, almacenándose previamente en las cisternas, desde donde pasan posteriormente al aparato de Golgi.
Retículo endoplasmático Rugoso: RER
Denominado así por la existencia de rugosidades presentes en los sáculos y que son ribosomas adosados a ellos.
Está formado por una red de membranas que forman cisternas,sáculos y tubos aplanados. Delimita un espacio interno llamado lúmen del retículo y sehalla en continuidad estructural con la membrana externa de la envoltura nuclear. El lumen ocupa el 10% del volumen total de la célula.Asociada a ribosomas.
Función: La síntesis de proteínas o traducción tiene lugar enlos ribosomas. Principalmente proteínas de membrana, de secreción, proteínas dentro de la célula para realizar funciones metabólicas. También transportar las proteínas hacia las regiones celulares en que sean necesarias o hacia el aparato de Golgi, desde donde se pueden exportar al exterior.
-Síntesis y almacenamiento de proteínas (necesita que los ribosomas del citoplasma estén unidos a ARN y que el ribosomase una a la membrana.-Glicosilación: las cadenas proteicas sintetizadas para que
puedan ser transportadas a otros orgánulos o al exterior de la célula han de estar glicosiladas.
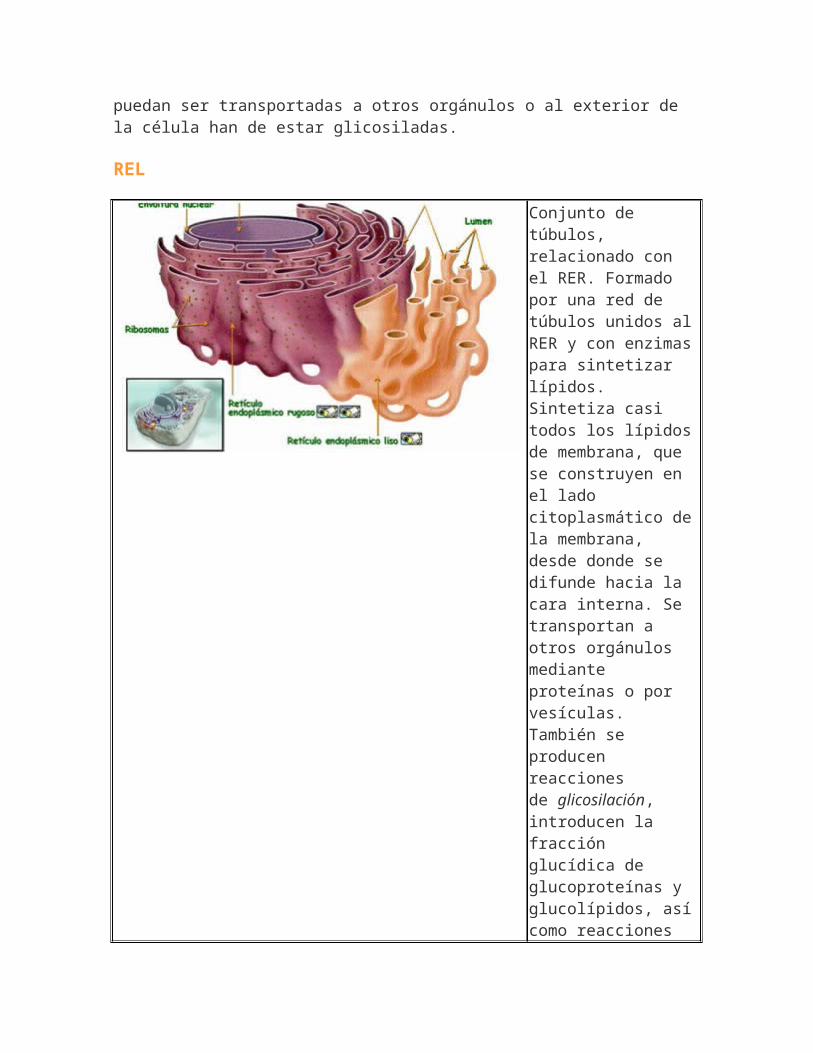
REL
Conjunto de túbulos, relacionado con el RER. Formado por una red de túbulos unidos alRER y con enzimaspara sintetizar lípidos. Sintetiza casi todos los lípidosde membrana, que se construyen en el lado citoplasmático dela membrana, desde donde se difunde hacia la cara interna. Se transportan a otros orgánulos mediante proteínas o por vesículas. También se producen reacciones de glicosilación, introducen la fracción glucídica de glucoproteínas y glucolípidos, asícomo reacciones

de elongación y formación de insaturaciones enlos ácidos grasos.
Síntesis de lípidos
Las membranas constituyen un ambiente lipídico, idóneo para la síntesis de sustancias apolares insolubles en el ambiente acuoso del citosol Además en las membranas se localizan sistemas enzimáticos que introducen grupos hidroxilo... en las moléculas extrañas para aumentar su hidrosolubilidad y facilitar su eliminación.En las células de la musculatura estriada se denomina retículo sarcoplásmico y acumula iones Ca para la contracción.Ambos retículos se intercomunican entre si, observándose conectes con la envoltura nuclear externa y la MP. Se pensó que el retículo formaba un canal de transporte que discurría por toda la célula y le comunicaba con el exterior: las gotaslipídicas en la absorción de grasas en enterocitos pasan por endocitosis a través de las microvellosidades al RER y al REL y a la envoltura nuclear (no es universal este sistema)Funciones -Síntesis y almacenaje y transporte de lípidos o esteroides, principalmente fosfolípidos y colesterol que constituyen el mayor porcentaje de lípidos de membrana.-Proceso de detoxificación de sustancias exógenas: insecticidas, herbicidas, conservadores de alimentos, son transformadas en otras menos tóxicas y fácilmente eliminadas por el REL mediante reacciones de oxidación y conjugación.-En la conducción de impulsos en el músculo estriado.-Metabolismo de minerales, producción de pigmentos y glucogenolisis.
Funciones comunes a ambos
-Soporte mecánico del citoplasma debido a la red de canalículos que se dispersan en el citoplasma.-Interviene en la formación de lipoproteínas: las proteínas sintetizadas en el RER pasan a los túbulos del REL uniéndose a los lípidos allí formados, luego son transportados por vesículas al Golgi que los llevará a su destino.-Transporta y almacena sustancias extracelulares (procedentesde endocitosis) o sintetizadas pro la célula.
Aparato de Golgi
Conjunto de membranas aplanadas, sáculo y túbulos (cisternas) asociados con vesículas de secreción . Cuando una serie de sáculos se apilan, forman un dictiosoma. Además, contiene vesículas a ambos lados y entre los sáculos. El conjunto de todos los dictiosomas y vesículas constituye el aparato de Golgi. Normalmente asociado a RER y REL
Función.-Es el organizador de la circulación molecular. Pasan gran número de moléculas procedentes del Re que sufren maduración a lo largo del recorrido. -Transporte, maduración y acumulación de proteínas del RER. Las sustancias pasan a través de las vesículas de transición y el contenido va avanzando de sáculoen sáculo mediante vesículas hacia la cara trans, en donde es

liberado en vesículas de secreción, liberándose por exocitosis. Otras vesículas se convierten en lisosomas primariosque vierten sus contenidos en los lisosomas secundarios. Funciona como una planta empaquetadora. -Reciclaje de la membrana, por la fusión de las vesículas secretoras con ella, se permite reponer los fragmentos que se pierden en la endocitosis tras la formaciónde las vesículas. -Síntesis de los componentes de la pared de células vegetales. -Modificación de la fracción glucídica de las glucoproteínas y glucolípidos sintetizados en el RE.Se eliminan azúcares y se adicionan otros nuevos. -Glucosilación: los productos elaborados en el REsufren nuevas transformaciones.
-Formación del tabique telofásico en vegetales, formación de la pared celular, formación acrosoma. Formación de nuevas membranas.
VACUOLAS.
Son orgánulos característicos de células vegetales relacionados con los lisosomas, funcional y estructuralmente.Suelen ser más abundantes y más voluminosos, pudiendo llegar a ocupar prácticamente todo el volumen celular, recibiendo elnombre de vacuola central.
Una vacuola es una vesícula muy grande, llena de líquido y rodeada por una membrana. La membrana de la vacuola se llama tonoplasto. Se forman por fusión de vesículas procedentes del golgi. El nº y tamaño varía, pudiendo alcanzar en la célula madura el 90%. También varía su
composición, normalmente agua, sales, azúcares y proteínas. Aveces están tan concentradas que forman cristales.
FUNCIÓN -En las vacuolas más pequeñas se acumulan sustancias de reserva, proteínas e hidrolasas inhibidas mientras no sean necesarias. -Algunos vegetales almacenan aceites y taninos. -En las semillas se forman los granos de aleurona, que es la forma en que la semilla mantiene en estado inactivo las reservas de materia que necesitará para su germinación. En ella, los granos de aleurona se hidratan hasta constituir las grandes vacuolas típicas.y los enzimas hidrolíticos desdoblan las moléculas de reserva en otras más sencillas que son utilizadas durante el crecimiento. -Productos de desecho tóxico (nicotina) como sistema de reserva. -Pigmentos antociánicos. -Medio de transporte entre orgánulos citoplasmáticos, o con el medio -Regulan la presión osmótica. -Actividad digestiva, por enzimas hidrolíticos.
Lisosomas
Los lisosomas son pequeños orgánulos de forma irregular que contienen reservas de enzimas necesarias para la digestión celular de numerosas moléculas. Normalmente indeseables. Muy teñidos y congruesas paredes para evitar la autodigestión.
Digestión celular: se encarga de romper moléculas orgánicas yfacilitar su aprovechamiento. Constituyen el aparato digestivo de la célula y se caracterizan por poseer

hidrolasas, que se encargan de la digestión de macromoléculas, procedentes del medio externo o de la propia célula. Poseen una membrana recubierta de glucoproteínas,que impiden que las enzimas ataquen a la membrana del lisosoma.
PEROXISOMAS Presentes en todas las células, casi. Son compartimentos que poseen enzimas, pero que intervienen en reacciones enzimáticas en las que se consume gran cantidad de oxígeno.
RH2 + O2 ---------R + H2O2 H2O + O2
Se forma H2O2, que es un veneno para la célula, pero los peroxisomas contienen gran cantidad de catalasa que cataliza la reacción. Así desaparece en el mismo orgánulo en el que seha producido.Utilizan oxígeno molecular para eliminar átomos de hidrógeno de sustratos orgánicos (Aa, ácidos grasos...). Y luego la catalasa utiliza el peróxido para oxidar diversas moléculas orgánicas pequeñas.Funciones:-Llevan a cabo reacciones ocidativas de degradación de ácidosgrasos y aminoácidos. No proporcionan a la célula energía, pero producen calor.-Intervienen en destoxificación (etanol)-En plantas están los glioxisomas, que transforman las grasasalmacenadas en las semillas en azúcares, necesarios para el desarrollo del embrión.
Orgánulos de doble membrana
<Además poseen ADN y ribosomas, es decir, la maquinaria necesaria para autoreplicarse. Se explica gracias a la teoríaendosimbiótica elaborada por Lynn Margulis. La teoría endosimbiotica afirma que las mitocondrias, cloroplastos y quizás también los centríolos y los flagelos surgieron de
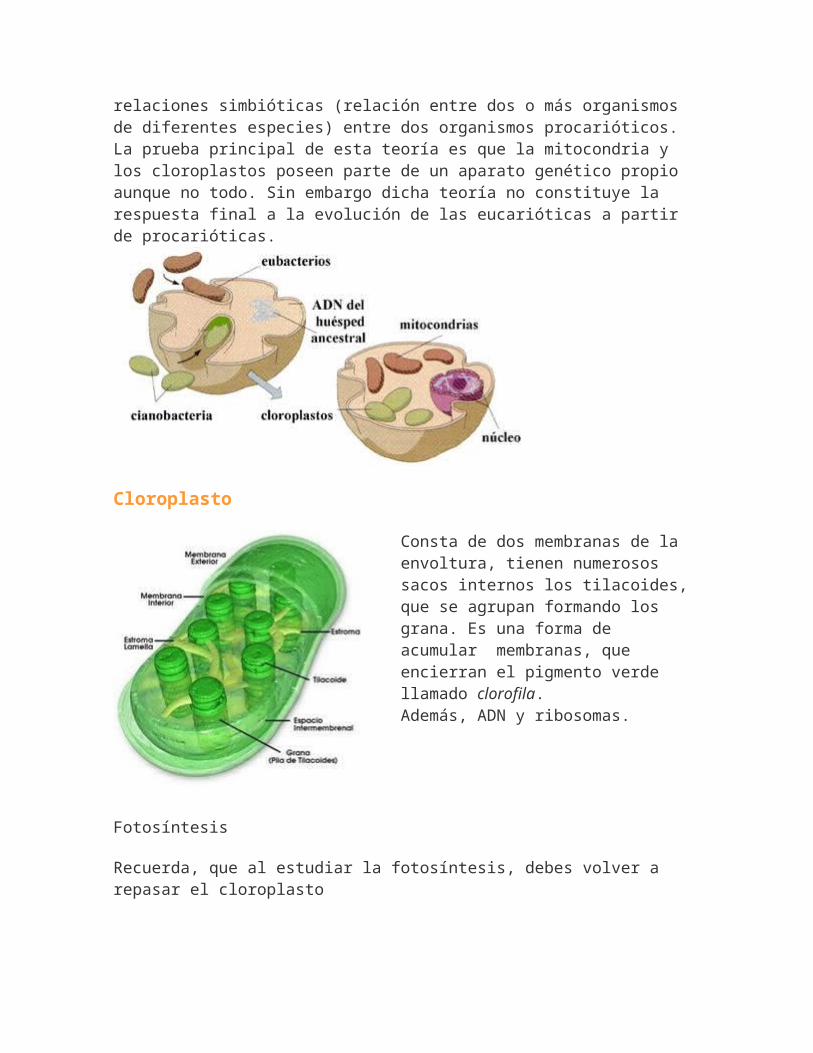
relaciones simbióticas (relación entre dos o más organismos de diferentes especies) entre dos organismos procarióticos. La prueba principal de esta teoría es que la mitocondria y los cloroplastos poseen parte de un aparato genético propio aunque no todo. Sin embargo dicha teoría no constituye la respuesta final a la evolución de las eucarióticas a partir de procarióticas.
Cloroplasto
Consta de dos membranas de la envoltura, tienen numerosos sacos internos los tilacoides, que se agrupan formando los grana. Es una forma de acumular membranas, que encierran el pigmento verde llamado clorofila.Además, ADN y ribosomas.
Fotosíntesis
Recuerda, que al estudiar la fotosíntesis, debes volver a repasar el cloroplasto

Las mitocondrias
Las mitocondrias son orgánulos granulares y filamentosos que se encuentran como flotando en el citoplasma de todas las células eucariotas. Aunque su distribución dentro de la célula es generalmente uniforme, existen numerosas excepciones. Por otro lado, las mitocondrias pueden desplazarse de una parte a otra de la célula. El tamaño también variable, pero es frecuente que la anchura sea de media micra, y de longitud, de cinco micras o más. En promedio, hay unas 2000 mitocondrias por célula, pero las células que desarrollan trabajos intensos, como las musculares, tienen un número mayor que las poco activas, comopor ejemplo las epiteliales.
Una mitocondria está rodeada por una membrana mitocondrial externa, dentro de la cual hay otra estructura membranosa, la
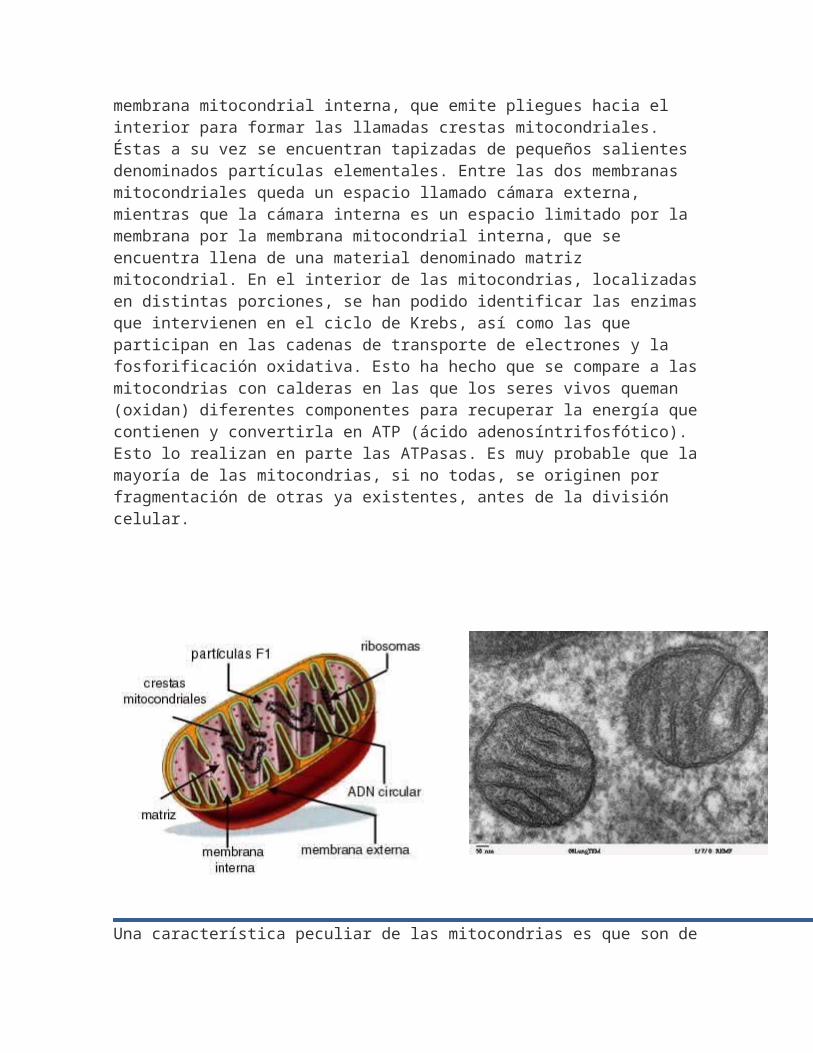
membrana mitocondrial interna, que emite pliegues hacia el interior para formar las llamadas crestas mitocondriales. Éstas a su vez se encuentran tapizadas de pequeños salientes denominados partículas elementales. Entre las dos membranas mitocondriales queda un espacio llamado cámara externa, mientras que la cámara interna es un espacio limitado por la membrana por la membrana mitocondrial interna, que se encuentra llena de una material denominado matriz mitocondrial. En el interior de las mitocondrias, localizadasen distintas porciones, se han podido identificar las enzimasque intervienen en el ciclo de Krebs, así como las que participan en las cadenas de transporte de electrones y la fosforificación oxidativa. Esto ha hecho que se compare a lasmitocondrias con calderas en las que los seres vivos queman (oxidan) diferentes componentes para recuperar la energía quecontienen y convertirla en ATP (ácido adenosíntrifosfótico). Esto lo realizan en parte las ATPasas. Es muy probable que lamayoría de las mitocondrias, si no todas, se originen por fragmentación de otras ya existentes, antes de la división celular.
Una característica peculiar de las mitocondrias es que son de

origen materno, ya que sólo el óvulo aporta las mitocondrias a la célula original, y cómo la mitocondria posee ADN , podemos decir que esta información va pasando a las generaciones exclusivamente a través de las mujeres. Por eso,cuando se hace la prueba de ADN se utiliza el ADN mitocondrial, poque sufre menos variaciones, llamado El ADN de Eva, Recuerda la teoría endosimbiótica que intenta explicar estas peculiaridades.
La científica estadounidense Lynn Margulis, junto con otros científicos, recuperó en torno a 1980 una antigua hipótesis, reformulándola como teoría endosimbiótica. Según esta versiónactualizada, hace unos 1.500 millones de años, una célula procariota capaz de obtener energía de los nutrientes orgánicos empleando el oxígeno molecular comooxidante, se fusionó en un momento de la evolución con otra célula procariota o eucariota primitiva al ser fagocitada sinser inmediatamente digerida, un fenómeno frecuentemente observado. De esta manera se produjo una simbiosis permanenteentre ambos tipos de seres: la procariota fagocitada proporcionaba energía, especialmente en forma de ATP y la célula hospedadora ofrecía un medio estable y rico en nutrientes a la otra. Este mutuo beneficio hizo que la célula invasora llegara a formar parte del organismo mayor, acabando por convertirse en parte de ella: la mitocondria. Otro factor que apoya esta teoría es que las bacterias y las mitocondrias tienen mucho en común, tales como el tamaño, la estructura, componentes de su membrana y la forma en que producen energía, etc.
Esta hipótesis tiene entre sus fundamentos la evidencia de que las mitocondrias poseen su propio ADN y está recubierta por su propia membrana. Otra evidencia que sostiene esta hipótesis es que el código genético del ADN mitocondrial no suele ser el mismo que el código genético del ADN nuclear.[2] A lo largo de la historia común la mayor parte de los genes mitocondriales han sido transferidos al núcleo, de tal manera que la mitocondria no es viable fuera de la célulahuésped y ésta no suele serlo sin mitocondrias.

Fuente: Wikipedia
Proceso de catabolismo: la respiración celular
Respiración Aerobia:
C6H12O6 + 6 O2 6CO2 + 6 H2O + 38 ATP
Fermentación
C6H12O6 2CO2 + 2 CH2OH-CH3 + 2 ATP
LA CÉLULA PROCARIOTA
Su origen evolutivo es anterior al de eucariotas,están normalmente aisladas y dan lugar a organismos unicelulares denominados moneras. Con frecuencia forman colonias o filamentos de células independientes, unidas entresí por sustancias extracelulares. Tamaño : 1-10 -Carece de verdadero núcleo aislado por una membrana nuclear.-Material cromosómico formado por una doble hélice de DNA desnudo, que no se asocia con proteínas del tipo de las histonas y se localiza en una región central denominada nucleoide, sin separación del resto.-Protoplasma poco diferenciado y aunque existe un conjunto desistemas membranosos formados por repliegues internos de la membrana plasmática, no se alcanza un alto grado de compartimentación.-Contiene ribosomas, a veces flagelos, cromatóforos con actividad fotosintética, inclusiones lipídicas y el equipo enzimático que les permite independizarse.-Suele contener pared celular
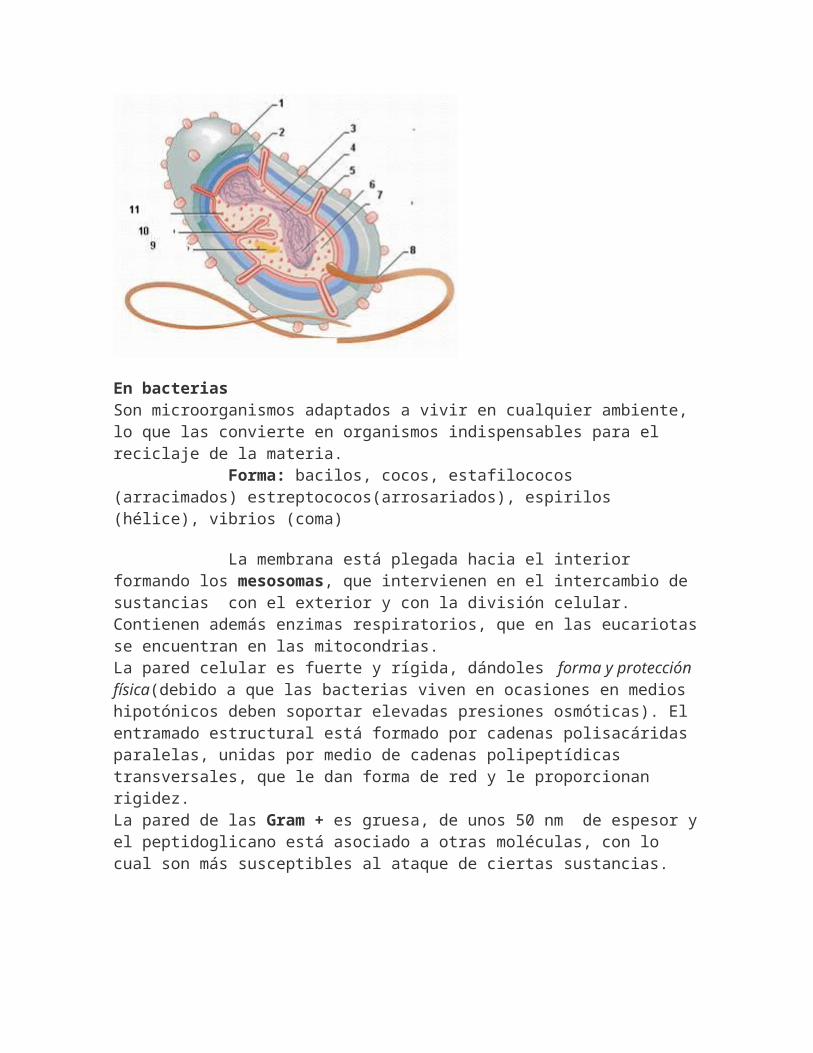
En bacterias Son microorganismos adaptados a vivir en cualquier ambiente, lo que las convierte en organismos indispensables para el reciclaje de la materia. Forma: bacilos, cocos, estafilococos (arracimados) estreptococos(arrosariados), espirilos (hélice), vibrios (coma)
La membrana está plegada hacia el interior formando los mesosomas, que intervienen en el intercambio de sustancias con el exterior y con la división celular. Contienen además enzimas respiratorios, que en las eucariotasse encuentran en las mitocondrias.La pared celular es fuerte y rígida, dándoles forma y protección física(debido a que las bacterias viven en ocasiones en medios hipotónicos deben soportar elevadas presiones osmóticas). El entramado estructural está formado por cadenas polisacáridas paralelas, unidas por medio de cadenas polipeptídicas transversales, que le dan forma de red y le proporcionan rigidez.La pared de las Gram + es gruesa, de unos 50 nm de espesor yel peptidoglicano está asociado a otras moléculas, con lo cual son más susceptibles al ataque de ciertas sustancias.

Comparación de las envolturas celulares bacterianas. Arriba:Bacteria Gram-positiva.1-membrana citoplasmática, 2-peptidoglicano, 3-fosfolípidos, 4-proteínas, 5-ácido lipoteicoico. Abajo: Bacteria Gram-negativa. 1-membrana citoplasmática (membrana interna), 2-espacio periplasmático, 3-membrana externa, 4-fosfolípidos, 5-peptidoglicano, 6-lipoproteína, 7-proteínas, 8-lipopolisacáridos, 9-porinas.
En las gram - es biestratificada, doble membrana plasmática con una capa de peptidoglicano mureina y sobre ella, otra formada fundamentalmente por lípidos, con lipopolisacáridos que son responsables de su resistencia a agentes bactericidas.-Es responsable de la virulencia de muchas bacterias-En ella residen los Ac bacterianos-Preserva a la bacteria de la acción de antibióticos.En algunos casos poseen una cápsula viscosa que la recubre. Se le atribuye la regulación del intercambio de sustancias y la protección frente a anticuerpos y células fagocitarias. Pueden o no formarla dependiendo de las condiciones del cultivo.-Ribosomas de 70S y DNA de doble hélice y circular-Exixtencia de flagelos y de fimbrias para la adherencia a los sustratos y el intercambio de fragmentos de DNA durante la conjugación.Funciones de relación -responden a gran número de estímulos ambientales, mediante modificaciones de su actividad metabólica o de comportamiento.

Forma
Formación de esporas de resistencia: endosporas
Frente a variaciones del medio se forman esporan, desarrollanprocesos en los que protegen su DNA y entran en metabolismo reducido. La más típica es la endospora que soporta condiciones de sequedad, Tª de 80º, agentes químicos y radiaciones hasta siglos.Para ello emigran el nucleoide haciauno de los polos. A partir de la membrana crece un mesosoma que formará una doble membrana alrededor del nucleoide separándolo del resto. La mayor parte degenera pèro las membranas y pared se convierten en las cubiertas externas.

-Movimientos de acercamiento o distancia.
FISIOLOGÍASegún la fuente de C que utilizan se dividen en autótrofos y heterótrofos. Según la fuente de E en fotótrofos y quimiótrofos.
Autótrofas: CO2 Heterótrofas: m. orgánicas
FotótrofasUtilizan la luz
Fotoautótrofas o fotosintéticasPurpúreas del S y verdes del S
FotoheterótrofasPurpúreas no S
QuimiótrofasUsan oxidaciones
QuimiolitotrofosFuente de E por oxidación de inorgánicos sencillos. Son las bacterias oxidantes del H, Nitrito, Fe++
Las cultivadas en laboratorio, las patógenas de plantas y animales:E. Coli, Cólera, tuberculosis...
Según utilicen el oxígeno o no:-Aerobias: usan el oxígeno como aceptor terminal de electrones: Pseudomonas(suelo y agua) y Bacillus(suelos)-Microaerófilas que respiran O pero no resisten concentraciones normales del aire.

-Anaerobias facultativas: como E. Coli y el vibrión del cólera, que no necesitan O.-Anaeróbias estrictas. han de vivir sin oxígeno, como las especies que viven en el intestino: clostridium-Anaeróbiasaerotolerantes: no uilizan el O2 pero no mueren ensu presencia: Lactobacillus y Streptococcus.Los microorganismos anaeróbios que usan biomoléculas orgánicas como aceptores terminales de electrones reciben el nombre de fermentadores. Funciones de reproducción
Las células bacterianas se multiplican asexualmente por división binaria transversa. Las células hijas formadas son genéticamente idénticas a la célula progenitora. Así, por sucesivas divisiones se formarán colonias de células iguales,que reciben el nombre de clones..
-Presentan también mecanismos que permiten un cierto intercambio de material genético, mediante el paso de fragmentos del DNA bacteriano o plásmidos de una célula a otra.ConjugaciónLas bacterias de tipo dador poseen una pequeña molécula de DNA o plásmido que recibe el nombre de factor F (de fecundidad), y se les llama F+. Las receptoras no la tienen y se les llama F—El factor F aparece normalmente separado de la molécula de DNA portadora de los genes y se duplica en cada multiplicación.Durante la conjugación se aproximan entre sí dos bacterias y se establece un puente citoplasmático. Puede ocurrir que una bacteria F- sea transformada en F+. En ocasiones el F y la molécula de DNA portadora de genes se unen en un solo mecanismo por un fenómeno parecido a entrecruzamiento. Se denominan Hfr. A continuación empieza a pasar a F- a través del puente. Normalmente no pasa todo. Así la bacteria F- aparece con genes nuevos. Muchas bacterias poseen plásmidos que se replican independiente del DNA bacteriano y son portadores de genes.Transformación bacteriana: un frgmento de DNA de una bacteriamuerta y rota, penetra en otra bacteria incorporádose al DNA de la 2ª. Se suele dar en cultivo

Transducción: los virus infectan una bacteria, integran su DNA en el bacteriano.
Coco Bacilo Vibrión Espirilo
FUENTE DE CARBONO ENERGÍA UTILIZADA
Autótrofas: la fuente de carbono es inorgánica (CO2)
Fotolitotrofas: la energía utilizada es la luz.(Ejemplo: bacterias purpúreas del azufre).Quimiolitotrofas: la energía utilizada es la liberada en reacciones químicas. (Ejemplo: bacterias incoloras del azufre).
Heterótrofas: la fuente de carbono es inorgánica.
Fotoorganotrofas: la energía utilizada es la luz.Quimioorganotrofas: la energíautilizada es la liberada en reacciones químicas. A este grupo pertenecen la mayoría delas bacterias.
La relación celular
Las funciones de relación son aquellas que permiten a la célula recoger información del medio en forma de señales o estímulos y responder a ellos adecuadamente. Se puede afirmarque son las funciones que confieren a las células su

sensibilidad e irritabilidad.2.1. Tipos de estímulos y respuestasLos estímulos que capta una célula pueden ser cualquier tipo de variación de las condiciones ambientales, pudiendo tener naturaleza muy diversa: luminosos, térmicos, mecánicos, químicos, magnéticos, gravitatorios, eléctricos, etc. Los de tipo químico son los que mayor variedad y complejidad presentan.Las respuestas también pueden tener una amplia gama demodalidades: secreción de sustancias, activación o desactivación delmetabolismo y de la división celular, formación de paredes protectoras (enquistamiento), emisión deluz (bioluminiscencia), etc. Pero, posiblemente, la forma de respuesta más extendida es el movimiento.Elmovimiento celular suele clasificarse según determine o no el desplazamiento de la célula y según afecte a su interior osolo a la superficie celular, aunque en general todos ellos tienen su origen en las estructuras del Citoesqueleto o derivados de lasmismas. Elmismo tipo demovimiento puede ser de locomoción en células libres y no serlo en células de tejidos (por ejemplo, los cilios del epitelio del árbol respiratorio tiene una función protectora).Cuando elmovimiento implica locomoción en organismos unicelulares y en algunas células libres de los pluricelulares, se llama tactismo, taxis o taxia, dependiendolos distintos nombres de la fuente de estimulo (foto tactismo, quimiotactismo, etc.). Se dice que es positivo o negativo según se dirija hacia el estimulo o se aleje de él, respectivamente. Entre los movimientos celulares más representados citaremos:• Ciclosis o corrientes endocelulares. Son frecuentes en células vegetales. Estas corrientes redistribuyen orgánulos celulares en respuesta a ciertos estímulos, como cambios de temperatura o de pH.• Contracción. Los provocan las estructuras fibrilares del Citoesqueleto, como, por ejemplo, los filamentos de actina y miosina de la célulamuscular, que se deslizan entre si y producen el acortamiento o estiramiento de toda la célula, loque conduce a la contracción de todo el tejido y del musculo

en su conjunto.• Ameboide o por pseudópodos. Es típico de amebas y otros protozoos, así como de células como los fagocitos de la sangre y de los tejidos. Recordemos que las prolongaciones citoplasmáticas que constituyen los pseudópodos sirven también para la captura de partículas. La base molecu¬lar de este movimiento implica la participación de filamentos de actina que producen la tracción del citoplasma y la formaciónde contactos mediante moléculas de adhesión con el sustrato. Córtex de actina córtex sometido a tensión polimeración de laactina en los extremos, movimiento de la actina no polimerizadaRetraimiento Movimiento celular Ameboide.
• Por cilios y flagelos. Los primeros producen un movimiento vibrátil sincronizado conotros muchos cilios. Cada cilio se mueve de forma similar a como lo hace el brazo de unnadador. Los segundos tienen un movimiento ondulatorio, como el de un látigo, que recorretoda su longitud. La basemoleculardelmovimiento de cilios y flagelos es el deslizamientode los brazos de dineina que se intercalan entre los microtúbulos, haciendo que
estos se flexionen.

2.2. La comunicación celularLas funciones de relación de la célula incluyen, entre las señales captadas del medio ambiente,aquellas que proceden de otras células y que permiten establecer la comunicación celular. Esta existe, por supuesto, en los organismos unicelulares, sobre todo en los que se agrupan para formar colonias (incluso entre simples bacterias), pero en los organismos pluricelulares llega a serindispensable, hasta el extremo de que debió de ser la dificultad para establecer esta comunicación celular una de las causas por las que se retardo el paso de los seres unicelulares a pluricelulares en el transcurso de la evolución.El estado pluricelular requiere que las células estén comunicadas mediante un complejosistema de señales, principalmente químicas, que permita coordinar su actividad comoun todo en beneficio del organismo. Distintos grupos celulares se han especializado solo en algunos aspectos de lacomunicación para satisfacer la relación del organismo, tantocon su entorno exterior como con su medio interno: la captación de señales por las células de los órganos de los sentidos, la transmisión de información por las neuronas o por las hormonas de la sangre, las respuestas mediante la contracción muscular, etc.Pero, en general, cada célula cuenta con un elaborado sistemade comunicación, que sereduce, en el nivel molecular, a toda una serie de proteínas que le permiten reconocer yresponder a señales procedentes de otras células.Por otra parte, ya hemos observado como la comunicación celular rebasa las funcionesde relación en sen.do estricto y permite que las células puedan responder a cues.onestales como determinar su posi.on espacial y funcional en el organismo, o asegurar quese dividirán en el momento oportuno de acuerdo con sus vecinas.Elementos de la comunicación celular

El sistema de comunicación o serializacion celular consta de los siguientes elementos:• Una célula señal o emisora de señales.• La molécula señal o mensajero químico.• La célula diana o receptora, que posee el receptor y el sistema de transducción de la señal.La célula emisora produce la molécula señal que, al unirse alreceptor, actúa como es.-mulo para generar la secuencia de reacciones intracelulares que hemos llamado transducciónde la señal, que conducen a la respuesta fisiológica de la célula diana.Célula emisora molécula señal célula diana
Elementos de señalizacion o comunicación.Observa que La comunicación celular en los animales es la quealcanza mayor grado de sofisticación y la que mejor se ha estudiado, pero no olvidemos que existe también en otros grupos, aunque menos desarrollada...Más informaciónConocemos ya algunos elementos del sistema de señales hemos tratado la transferencia de información como función de las membranas y hemos estudiado algún ejemplo de transducción de señales en el interior celular, a propósito del control del ciclo celular y la apoptosis
La célula señal y la emisión de señalesLa célula emisora de señales puede tener la molécula señal fija a su membrana y unirse con la célula diana a través de

su receptor, pero es más frecuente que segregue la señal en forma de mensajero químico para hacerla llegar a la célula diana. Se pueden presentar tres modalidades de secreción o formas de señalización:• Paracrina, si la molécula señal, llamada mediador local, sesegrega al medio intercelularpara ser captada de inmediato por los receptores de las células vecinas.• Endocrina, si la señal, llamada ahora hormona, se segrega ala sangre (en los animales), o ala savia (en las plantas), por donde se transportan hasta alcanzar los receptores de las células diana repartidas por el cuerpo.• Sináptica, si la señal, llamada neurotransmisor, es segregada por una neurona en su membrana presináptica y, a través de la brecha sináp.ca, alcanza los receptores de su célula diana.Las moléculas señal Los mensajeros químicos pueden ser sustancias hidrofilias (proteínas, pép.dos y derivados de aminoácidos), como son muchas hormonas, neurotransmisores y factores de crecimiento,que no pueden atravesar la membrana, por lo que sus receptores están en la superficie celular.Otras son sustancias hidrofóbicas, como hormonas esteroides, hormonas .roideas, vitamina D, etc.,que son transportadas en el medio interno por proteínas especificas hasta alcanzar las membranas, que atraviesan con facilidad, para unirse a receptores intracelulares. Algunos gases, como el oxido nítrico y el mon6xido de carbono, actúancomo mensajeros que atraviesan la membrana y actúan directamente.Los receptores y la transducción de señalesLos receptores de las células diana pueden estar en la superficie celular o en su interior.• Los receptores de superficie unen mensajeros hidrolicos y pueden agruparse en tres tipos ofamilias. Cada tipo da lugar a formas de transducción de señales diferentes:— Receptores asociados a canales iónicos. El receptor es un

canal iónico regulado por ligando.— Receptores asociados a proteínas G. El receptor, al unirse con la señal o "primer mensajero",actúa sobre un intermediario, la proteína G (llamada así porque se une al GTP), que a su vez,actúa sobre una enzima de la cara interna de la membrana. Esta ac.va un segundo mensaje-ro intracelular, que inicia la transducción de la señal hacia el interior de la célula, modificando otras proteínasque, finalmente, alteran el comportamiento de la célula y dancomo resul¬tado la respuestafisiológica concreta según cada .po celular, o bien llegan alADN nuclear y activan su duplicación o su transcripción.— Receptores asociados a enzimas. Actúan directamente como enzimas que se ac.van alunirse a la señal. Los más conocidos .enenac.vidad quinasa o están asociados a otras enzimasquinasas o fosfatasas, que a su vez fosforilan o desfosforilanrespec.vamente a otras proteínas en la cadena dereacciones de la transducción de señal. Finalmente, alteran también la expresión de los genes uniéndose al ADN nuclear.• Los receptores intracelulares pueden estar en el citoplasmao en el núcleo. Por ejemplo, elreceptor de la hormona Cor.sol es citoplasmá.co y, una vez formado el complejo de unión entreambos, penetra en el núcleo para unirse al ADN y modificar laexpresión gené.ca, con el consiguiente cambio de comportamiento o ac.vidad celular.Así, la comunicación celular recuerda una carrera de relevos en la que cada corredor deja el testigo (la señal) a otro y así sucesivamente, con la complicación añadida de que una misma serial inicial desencadena en cada tipo celular una respuesta distinta con resultado final diferente y de que la señal se va amplificando en cada relevo, implicando a mas mediadores.
Metabolismo celular

•Metabolismo: conjunto de reacciones químicas que se producen en interior celular, para mantener su composición, para realizar la transcripción, síntesis de proteínas, hidrólisis de grasas, ruptura de almidón o glucógeno...
Se produce gracias a la presencia de enzimas.Clasificación:
Anabolismo, que son reacciones constructivas, de síntesis, endoergónicas, en las que se requiere energía.¿Quién se la proporciona?
Catabolismo, que son reacciones de degradación de moléculas complejas que pasan a convertirse en moléculas sencillas.La liberación de energía química no se pierde, sino que se conserva en forma de enlaces fosfato del ATP, con lo que la energía química liberada se transfiere a las reacciones anabólicas.
Anabolismo y catabolismo se producen simultáneamente en las células, y están catalizadas enzimáticamente. Las reacciones que constituyen el catabolismo sonconvergentes, y las del anabolismo, divergentes (a partir de pocas moléculas se sintetizan En la última fase se producen moléculas que

pueden utilizarse en el catabolismo para su degradación completa hasta CO2, agua y amoníaco, o bien ser moléculas precursoras para la síntesis de otras moléculas
REQUERIMIENTOS ENERGÉTICOS DE LA CÉLULA.NUTRICIÓN AUTOTROFA Y HETEROTROFA
-Todos los seres vivos utilizamos energía para realizar nuestro metabolismo-El tipo de energía es para todos el mismo :la energía química encerrada en las moléculas en forma de enlace.Todos los seres vivos necesitamos esta energía.Pensamos que los vegetales utilizan la energía lumínica para realizar sus funciones, pero fabrican moléculasorgánicas a partir de la cuales realizan su metabolismo.1.Según la fuente de carbono que utilicen para sintetizar susmoléculas orgánicas:-Autótrofos-que utilizan el CO2 como única fuente de carbono.Vegetales y algunas bacterias.-Heterótrofos que necesitan moléculas más complejas, como la glucosa, como fuente de carbono (elaboradas por los autótrofos).Además, hongos y microorganismos.2.-Según la fuente de energía Los autótrofos necesitan una fuente de energía para la conversión de moléculas, ya que la formación de nuevos enlaces requiere un aporte.Según esta fuente, se clasifican enfotosintéticos si la energía la obtienen de la luz, y quimiosintéticos si procede de reacciones exergónicas en las que intervienen moléculas inorgánicas sencillas, como el amoníaco, el azufre o el hierro.Muy importantes en el caso debacterias fijadoras de nitrógeno atmosférico, o las que transforman el amoníaco de suelo en nitritos, que por otras bacterias nitrificantes, se convierten en nitratos, que es laforma en que puede ser asimilado el nitrógeno por las raíces de las plantas.
TIPO DE ORGANISMO FUENTE DE C FUENTE DE E EJEMPLOSFOTOLITOTROFO CO2 LUZ VEGETALES
CIANOBACTERIB.FOTOSINT

FOTOORGANOTROFO COMPUESTOS ORGANICOS
LUZ BACTERIAS PÚRPURA NO S
QUIMIOLITOTROFO CO2 REACCIONES REDOX
B.DESNITRIFICANTES S, FE
QUIMIORGANOTROFO COMPUESTOS ORGÁNICOS
REACCIONES REDOX
SUPERIORES, HONGOS.MICRO
EL ATP Y LOS INTERCAMBIOS DE ENERGÍA EN EL METABOLISMO Hay procesos que liberan energía y otros que la consumen.La liberación y el consumo no tienen porque ocurrir al mismo tiempo, ni en el mismo lugar de la célula.Debe existir un mecanismo que almacene y transporte esta energía desde los lugares donde se produce hasta donde se consume.Se basa en laformación y ruptura de enlaces químicos que acumulan y liberan gran cantidad de energía.Se suele usar el ATP.
•El ATP es intermediario energético celular: por enlaces P-P.La hidrólisis libera 31 K J /mol.•Se forma y se consume continuamente. Es molécula de transferencia, no de reserva E.1.-Utilización de la energía libre almacenada en el ATP.La hidrólisis es un proceso espontáneo que permite acoplarlo a procesos que no son posibles sin aporte energético.El acoplamiento se hace mediante enzimas.2.-Utilización del ATP para almacenar energía libre. La fosforilación es un proceso endoergónico que requiere un aporte energético.Esta reacción tiene lugar en el interior delas células, acoplada a otros procesos exergónicos-En la célula se utilizan dos mecanismos distintos para sintetizar ATP.

Fosforilación a nivel de sustrato.Primero se forma un sustrato intermedio "rico en energía" y en la segunda se utiliza la energía liberada por la hidrólisis de este compuesto para la fosforilación del ADP a ATP Fosforilación en el transporte de electrones.Las células utilizan el transporte de electrones a través de proteínas ubicadas en la membrana de las mitocondrias o cloroplastos, libera energía que es utilizada por una enzima,la ATP-sintetasa, para acoplar la fosforilación del ADP a ATP.Se denomina fosforilaciónfotosintéticasi se produce en elcloroplasto y fosforilación oxidativa, si tiene lugar en las mitocondria.(Hay también otros nucleótidos que cumplen funciones similares)3.-Los electrones: otro vehículo para transferir energía.Muchas de olas reacciones del catabolismo suponen la oxidación de un sustrato, lo que libera electrones, mientras que por el contrario, la biosíntesis de moléculas ricas en hidrógeno, como los ácidos grasos, requiere electrones.Los electrones son transportados enzimáticamente desde las reacciones catabólicas de oxidación en que son liberados, hasta las reacciones anabólicas de reducción que precisan de ellos. Para ello se utilizan coenzimas transportadores de e como el NADP, que llevan a estos desde un punto a otro de la célula, de forma similar a como lo hace el ATP con la energía.
En definitiva, en el metabolismo se producen de forma continua reacciones redox, de oxidación-reducción
Cuando los grupos fosfatos se transfieren al ADP para formar ATP, se está almacenando energía. Otra forma es transferir electrones (e-), las reacciones se denominan de oxidorreducción o reacciones redox.
La ganancia de uno o más e- por un átomo, ión o molécula --> REDUCCIÓNla pérdida de uno o más e- por un átomo, ión o molécula --> OXIDACIÓN

Hay que tener en cuanta que una molécula se oxida o se reduceno solamente cuando intercambia e-, sino también cuando intercambia átomos de Hidrógeno (no iones H), ya que involucra transferencia de electrones: H = H+ + e- . Por ello una oxidación siempre ocurre simultáneamente con unareducción. Cuando un material se oxida, los e- perdidos se transfieren a otro material, reduciéndolo.
-Concepto y finalidad del metabolismo.El metabolismo es el conjunto de reacciones químicas que se producen en el interior de las células y que conducen a la transformación de unas biomoléculas en otras. Las distintas reacciones químicas del metabolismo se denominan vías metabólicas y las moléculas que intervienen son metabolitos.Todas las reacciones del metabolismo están reguladas por enzimas que son específicas para cada metabolito inicial o sustrato y para cada tipo de transformación. Las sustanciasfinales de una vía metabólica se denominan productos . Las conexiones exitentes entre diferentes vías metabólicas reciben el nombre de metabolismo intermediario. Se pueden considerar dos fases.-Fases metabólicas: catabolismo y anabolismo: concepto, finalidad y relación entre ambas.El catabolismo es la transformación de moléculas orgánicas complejas en otras más sencillas, proceso en el que se liberaenergía que se almacena en los enlaces fosfato del ATP.El anabolismo es la síntesis de moléculas orgánicas complejasa partir de otras biomoléculas más senicllas, para lo cual senecesita suministrar energía, proporcionada por los enlaces

fosfato del ATP. Las moléculas del ATP pueden proceder de lasreacciones catabólicas, de la fotosíntesis o de la quimiosíntesis.Anabolismo y catabolismo se producen simultáneamente en las células, y están catalizadas enzimáticamente. Las reacciones que constituyen el catabolismo sonconvergentes, y las del anabolismo, divergentes (a partir de pocas moléculas se sintetizan En la última fase se producen moléculas que pueden utilizarse en el catabolismo para su degradación completa hasta CO2, agua y amoníaco, o bien ser moléculas precursoras para la síntesis de otras moléculas
-Aspectos fundamentales, energéticos y de regulación de las reacciones metabólicas.En las reacciones metabólicas se forman nuevas sustancias-losproductos- a partir de reactivos y en segundo lugar, durante la reacción hay un intercambio de energía entre los compuestos reaccionantes y el medio que las rodea. Además:-Las reacciones suceden en un medio acuoso (reactivos y productos se encuentran en disolución) y el agua puede ser reactivo o producto.-La reacciones están encadenadas, de forma que el producto de

una reacción es el sustrato o metabolito de la siguiente. La mayoría están acopladas de manera que la energía liberada en una de las reacciones, exergónica es captada por otra reacción-endergónica que requiere energía.
Características reacciones metabólicas
•Son reacciones de oxidación-reducción•Son reacciones acopladas: anabolismo, endergónicas, con catabolismo, exergónicas.•Están catabolizadas por enzimas.
Los enzimas
•Término propuesto por Kühne en 1878 (zyme = levadura)
-Cada reacción metabólica tiene su catalizado o enzima. En general. Son sustancias que aceleran las reacciones químicas porque disminuyen la energía de activación de la reacción.-En las reacciones acopladas, una misma enzima cataliza las dos reacciones a la vez.En toda reacción química es necesaria la aportación de una energía inicial o de activación para que se inicie la reacción, independientemente de que la reacción sea espontánea y vaya acompañada de una liberación de energía en forma de calor (exotérmica) o de absorción de calor(endotérmica)
En general todas las reacciones metabólicas están regulas porlos enzimas, unas proteínas globulares que actúan como catalizadores, aumentando la velocidad de aquellas reaccionesque son energéticamente posibles. Las enzimas permiten las

reacciones en las condiciones de Tª. P y pH propios del mediointracelular, reduciendo la energía de activación necesaria para que se produzca la reacción.
1.-Definición.
-Los enzimas ejercen su acción biológica uniéndose selectivamente a determinadas moléculas: SUSTRATOS-Inducen modificaciones químicas en los sustratos por ruptura, formación o redistribución de sus enlaces covalenteso por introducción o pérdida de grupos funcionales.-Son biocatalizadores de las reacciones que constituyen el metabolismo S + E -------P-Intervienen a concentraciones muy bajasAceleran las reacciones sin sufrir modificaciones.
•Hacen que las reacciones se canalicen hacia rutas que sean útiles en lugar de hacia reacciones colaterales que despilfarren energía• Pueda regularse la producción de distintas sustancias segúnlas necesidades
ENERGÍA Y REACCIONES QUÍMICAS
TIPOS DE REACCIONES
1.-EXERGóNICAS O EXOTÉRMICAS.-las moléculas que van a reaccionar poseen una energía interna mayor que los productosque se desprenden de la misma : se libera energía
|| AB -Transcurren de forma espontánea|----- (ej. agua de un depósito en el | tejado, a cualquier piso) | A+B| -----|

|_______________________Reacción
2.-ENDERGONICAS O ENDOTERMICAS: es necesaria la aportación deenergía.Los reactivos tienen menor energía que los productos que se aporta en forma de nuevos enlaces.
Energía
|| CD -Transcurren de forma no espontánea| ----- (ej. agua de un depósito en el | sótano a cualquier piso) |C+D |---- ||_______________________Reacción
En las células:
Se producen ambas de forma espontánea, son Reacciones acopladas.La reacción endergónica, no espontánea, se asocia a la exergónica, en la que se produce el paso de ATP a ADP.El ATP es una molécula móvil que actúa como transportadora deenergía química.Si el P se une a moléculas orgánicas P! y es un enlace rico en energíaCuando en la célula se producen exergónicas (de degradación) la energía liberada es aprovechada por el ADP que lo acumula en forma de ATP.
AB --------------------A + B
ADP + Pi ATP
CD¬--------------------C+D

La espontaneidad no es inmediata: Ninguna reacción se produce de forma espontánea, sin aporte de energía previo, necesario para que se inicie la reacción.Las moléculas que reaccionarán deben alcanzar el estado de transición: activación transitoria, deben " animarse" para colisionar y poder reaccionar.Estas energías que requieren es la energía de activación: a menor energía de activación, más fácilmente se produce la reacción (ej.la combustión de madera es espontánea, exergónica, pero no empieza a arder por sí misma) LAS ENZIMAS CATALIZAN TODAS LAS REACCIONES...¿Cómo?
FUNCION DE LOS CATALIZADORES Reducen la energía necesaria para que las moléculas estén en estado de transición, es decir, rebajan laenergía de activación, son mucho más eficaces que los catalizadores químicos, rebajan mucho más
Esto se traduce en un aumento de velocidad. Ej, digestión defilete sin enzimas sería infinito, con catálisis Qº: 3 meses,y con enzimas dura 2 horas
Catalizador
•La reacción E+S requiere aporte inicial de E: E de activación•Incrementa la Ec de las moléculas que reaccionan, permitiendo que mayor número de ellas choquen, y debiliten los enlaces
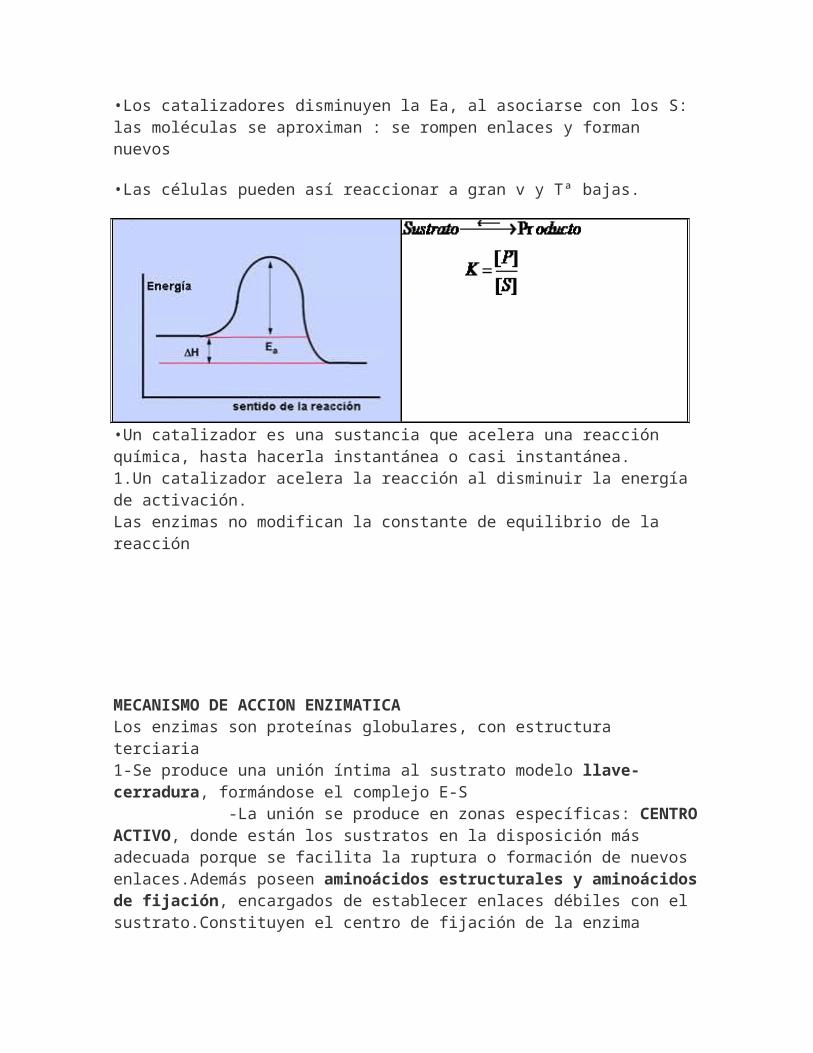
•Los catalizadores disminuyen la Ea, al asociarse con los S: las moléculas se aproximan : se rompen enlaces y forman nuevos
•Las células pueden así reaccionar a gran v y Tª bajas.
•Un catalizador es una sustancia que acelera una reacción química, hasta hacerla instantánea o casi instantánea.1.Un catalizador acelera la reacción al disminuir la energía de activación.Las enzimas no modifican la constante de equilibrio de la reacción
MECANISMO DE ACCION ENZIMATICA Los enzimas son proteínas globulares, con estructura terciaria1-Se produce una unión íntima al sustrato modelo llave-cerradura, formándose el complejo E-S -La unión se produce en zonas específicas: CENTROACTIVO, donde están los sustratos en la disposición más adecuada porque se facilita la ruptura o formación de nuevos enlaces.Además poseen aminoácidos estructurales y aminoácidosde fijación, encargados de establecer enlaces débiles con el sustrato.Constituyen el centro de fijación de la enzima

2-Modelo adaptación inducida. el centro activo no es rígido yla unión con el sustrato induce un cambio conformacional en el centro activo, que sitúa a los Aa catalíticos en la posiciópn correcta para catalizar la reacción
-Se producen reacciones con rapidez, se liberan productos y el enzima queda disponible
CARACTERISTICAS DE ENZIMAS
La característica más sobresaliente de los enzimas es suelevada especificidad.

o Especificidad de sustrato. El sustrato (S) es la molécula sobre la que el enzima ejerce su acción catalítica.
o Especificidad de acción. Cada reacción está catalizada por un enzima específico
Especificidad: si no tiene configuración adecuada, no ofrecerá la posibilidad de que se una a su centro activoy ofrecerá la posiblidad de que se una a su centro activo y no se formará complejo E-S que facilitaría la reacción.El centro activo es una zona de la proteína formada por una serie de Aa: pueden estar muy alejados entre sí en la estructura 1ª, pero cercanos en la 3ª, con lo cual la desnaturalización = inactivación, pierde su forma. Es variable, hay enzimas que llegan a distinguir formas L o D, actuándo específicamente con una u otra, esespecificidad absoluta , otras más generales, de grupo como la sacarasa, que actúa frente a la sacarosa ya la maltosa y de clase, cuando no depende de la molécula sino del enlace: fosfatasas.El resto de Aaayudan a mantener la estructura tridimensional del centro activo.
2.-No forman parte del producto
3.-No se consumen
4.-Son necesarios en pequeña cantidad
5.-La acción enzimática se caracteriza por la formación de un complejo que representa el estado de transición.

Bases de la reacción enzimáticaPara que se dé una reacción química tienen que verificarse 3 condiciones: 1.- Los reactivos, llamados sustratos en enzimología, deben colisionar 2.- La colisión molecular tiene que ocurrir con una orientación adecuada (las enzimas aumentan la probabilidad) 3.- Los reactivos deben poseer suficiente energía para alcanzar el estado de transición. Esta energía se llama energía de activación. Los enzimas hacen que la reacciónvaya más rápida
En las reacciones bioquímicas
o Energía de activación: barrera de energía que hay que superar para que se produzca la reacción. Es laenergía necesaria para:
o Alinear grupos reactivoso Formar cargas inestables transitoriaso Reordenar enlaceso Superada esta barrera, se llega a un estado
activado o de transición en el que se produce la orientación y condiciones adecuadas para la reacción.
Acción enzimática
o Una enzima une la molécula de sustrato de forma específica en una región denominada CENTRO ACTIVO

suele ser un bolsillo o una hendidura que se forma entre las cadenas laterales de los aminoácidos.
o Estas cadenas: Facilitan la unión del sustrato
(especificidad de sustrato) Intervienen en la catálisis (centro
catalítico)
El centro activo1.- Supone una porción relativamente pequeña del volumentotal de la enzima 2.- Es una entidad tridimensional 3.- Se unen a los sustratos por fuerzas relativamente débiles 4.- Son hoyos o hendiduras de las que suele quedar excluida el agua, salvo que sea un componente de la reacción 5.- La especificidad del enlace depende de la disposición de los átomos del centro activo 6.-Región que se une al sustrato y contribuye con los residuos que participan en la formación
Ejemplos

ESTRUCTURA DE LOS ENZIMAS
Hay enzimas estrictamente proteicas, holoproteínas y otras qye necesitan moléculas no proteícas para realizarsu función:holoenzimas, que son enzimas constituidos porla asociación de una fracción polipeptídica o apoenzima y de una fracción no polipeptídica o cofactor
HOLOENZIMA =APOENZIMA + COFACTOR
Los cofactores pueden ser:
o Inorgánicos : metales que se encuentran en pequeñas

cantidades, como el Zn, Mg, K...OligoelementosSe pueden integrar directamente en el centro activoSirven de puente E-S
o Orgánicos : +Grupos prostéticos, que están fuertemente unidos por enlace covalente : hemo...+Coenzimas que son moléculas tipo vitaminas. No son específicos del enzima, pero sí del tipo de reacción, transportando grupos funcionales:-El H : NAD, NADP, FAD, FMN (sobre DH)-Grupo acetilo : Co A-Fosfatos : ATP
Vamos a ver cómo funcionan los coenzimas
•Especializados en el transporte de electrones y protones, reduciéndose y oxidándose permitiendo que se den las reacciones metabólicas:N A D + + 2 H + + 2 e- N A D H + H + N A D P + + 2 H + + 2 e- N A D P H + H + FA D + + 2 H + + 2 e- FA D H 2
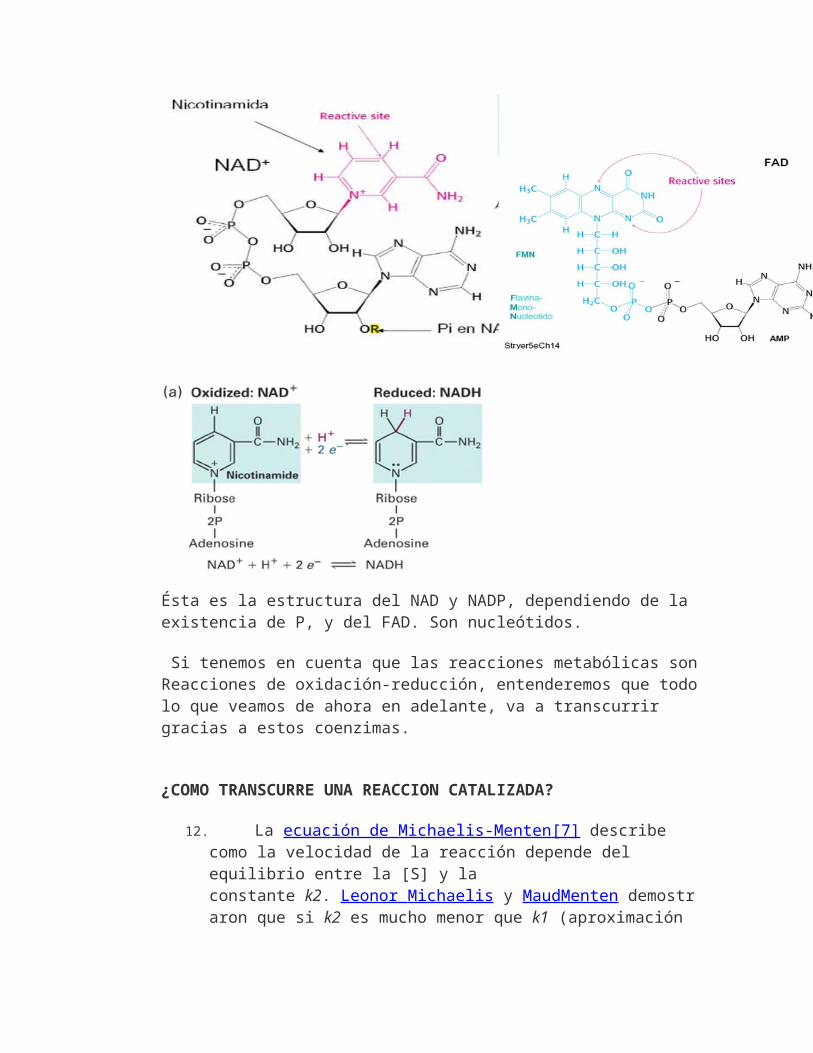
Ésta es la estructura del NAD y NADP, dependiendo de la existencia de P, y del FAD. Son nucleótidos.
Si tenemos en cuenta que las reacciones metabólicas sonReacciones de oxidación-reducción, entenderemos que todolo que veamos de ahora en adelante, va a transcurrir gracias a estos coenzimas.
¿COMO TRANSCURRE UNA REACCION CATALIZADA?
12. La ecuación de Michaelis-Menten [7] describe como la velocidad de la reacción depende del equilibrio entre la [S] y la constante k2. Leonor Michaelis y Maud Menten demostraron que si k2 es mucho menor que k1 (aproximación
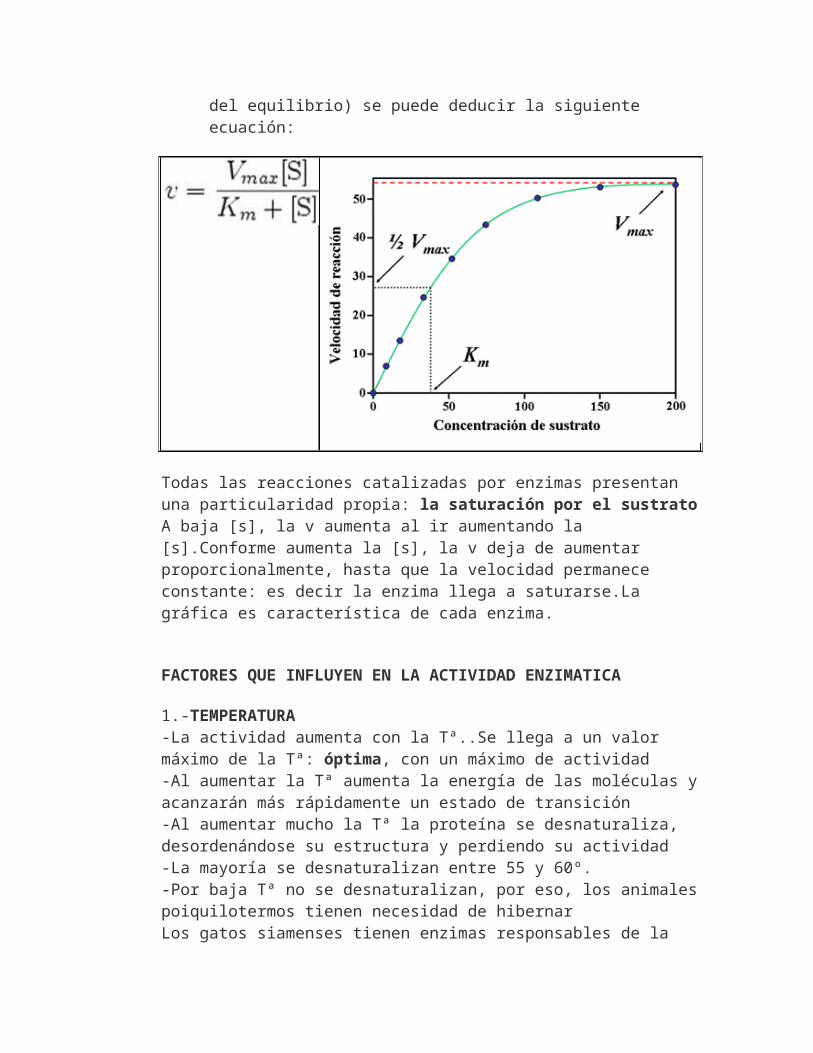
del equilibrio) se puede deducir la siguiente ecuación:
Todas las reacciones catalizadas por enzimas presentan una particularidad propia: la saturación por el sustratoA baja [s], la v aumenta al ir aumentando la [s].Conforme aumenta la [s], la v deja de aumentar proporcionalmente, hasta que la velocidad permanece constante: es decir la enzima llega a saturarse.La gráfica es característica de cada enzima.
FACTORES QUE INFLUYEN EN LA ACTIVIDAD ENZIMATICA
1.-TEMPERATURA-La actividad aumenta con la Tª..Se llega a un valor máximo de la Tª: óptima, con un máximo de actividad-Al aumentar la Tª aumenta la energía de las moléculas yacanzarán más rápidamente un estado de transición-Al aumentar mucho la Tª la proteína se desnaturaliza, desordenándose su estructura y perdiendo su actividad-La mayoría se desnaturalizan entre 55 y 60º.-Por baja Tª no se desnaturalizan, por eso, los animalespoiquilotermos tienen necesidad de hibernarLos gatos siamenses tienen enzimas responsables de la

pigmentación del pelo que funcionan con actividad máximaa Tª inferiores al resto del cuerpo
2.-pH.-La actividad enzimática es máxima para ciertos valores: pH óptimo. siendo éste muy variable de unos a
otros (1,5-10)
El centro activo puede contener aminoácidos con grupos ionizados que pueden variar con el pH.
13. La ionización de aminoácidos que no están en el centro activo puede provocar modiicaciones en laconformación de la enzima.
14. El sustrato puede verse afectado por las variaciones del pH.
o Algunos enzimas presentan variaciones peculiares. La pepsina del estómago, presenta un óptimo a pH=2,y la fosfatasa alcalina del intestino un pH= 12
-Las cadenas laterales de algunos Aa presentan grupos funcionales que a determinados pH presentan carga eléctrica.Estos pueden relacionarse con otros mediante interacciones no covalentes y contribuyen a la estructura tridimensional de la proteína o del centro activo.Si varía el pH determinadas interacciones no

pueden ocurrir, lo que alteraría desde la estructura delcentro activo hasta la interacción con el sustrato.Pequeñas variaciones en el pH ocasionan grandescambios en la actividad del enzima.-El pH óptimo varía para los distintos enzimas: la pepsina a pH ácido y la tripsina a pH básico.
3.-Concentración de enzima y sustrato:
o Exceso: se incrementa
3.-INHIBICION ENZIMATICA
Un inhibidor es una molécula capaz de emular específicamente la actividad enzimática.-En muchos casos para poder establecer el sustrato correcto de una enzima se han utilizado moléculas de estructura química muy parecida que pudieran competir con ella.Con estos inhibidores ha sido posible descubrirla naturaleza del centro activo e identificar los gruposfuncionales responsables de la acción.
3.-INHIBIDORES IRREVERSIBLES: son sustancias que disminuyen la actividad y eficacia de un enzima.Los hay de dos tipos
3.1.IRREVERSIBLES: son compuestos que se unen irreversiblemente a determinados grupos del centro activo y anulan su capacidad catalítica.Ej: venenos y toxinas

3.2.REVERSIBLES : no se inutiliza el centro activo, sólo se impide temporalmente su funcionamiento.
3.2.3.Competitiva: presencia de inhibidores , que son moléculas muy similares al sustrato.Si se fija el inhibidor, la enzima no cataliza.
Características
o Ocupando temporalmente el centro activo.
•Es reversible
•Depende del nº de moléculas de inhibidor y de sustrato.
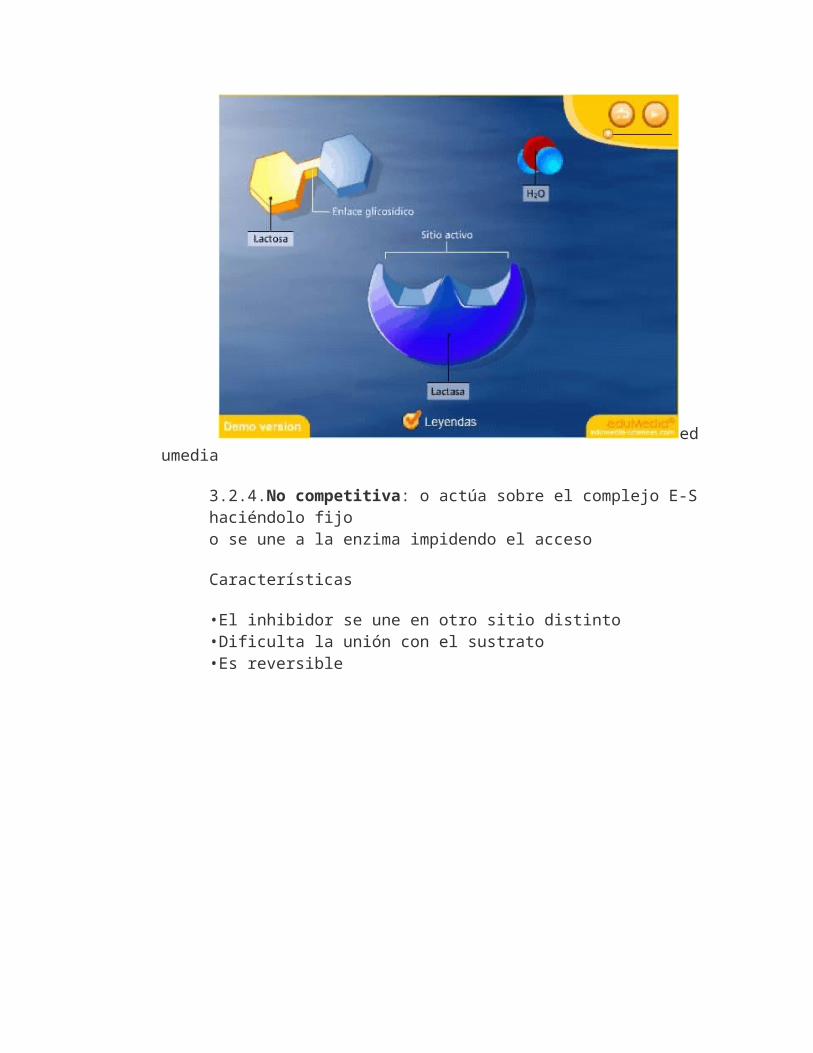
edumedia
3.2.4.No competitiva: o actúa sobre el complejo E-Shaciéndolo fijoo se une a la enzima impidendo el acceso
Características
•El inhibidor se une en otro sitio distinto•Dificulta la unión con el sustrato•Es reversible

4-CONCENTRACION DE SUSTRATO Al aumentar la concentración, aumenta la Probabilidad deencuentroSi aumenta mucho, se produce saturación de enzimas
KM depende de la afinidad enzima sustrato y es la [s] enque la v= 1/ Vmáx5.-ACTIVADORES: algunos iones favorecen la unión E-S.Asíla fosforilasa que regula la formación de ATP y se ve activada por la presencia de Mg2+
ENZIMAS ALOSTÉRICOS Están formados por varias subunidades o protómeros.Cadaprotómero posee dos centros: el regulador y el catalítico Al unirse el activador al centro regulador, la conformación varía haciéndose funcional el centro catalítico.Así pasa del estado inhibido (T) al

catalítico (R).La variación en la conformación se transmite a otros protómeros haciéndolos activos: Transmisión alostérica.
Existen enzimas que adoptan al menos dos conformaciones distintas y estables: activa e inactiva.El paso de una a otra está inducida por la unión de los ligandos a determinados lugares de la superficie enzimática:centros reguladores
Unos se encuentran inhibidos y requieren un activador (el S) para pasar a estado catalítico.Otros, en estado catalítico y la unión del producto con el regulador induce a un estado inhibido.Es el efecto feed-back y supone un ahorro energético
|-------E1 E2 E3Sustrato ------ A -----------B ---------- C
E1 E2 E3Sustrato ------ A -----------B ---------- C |________________________________|
Alosterismo
18. En puntos estratégicos: principio o ramificación

19. Puede darse:
o Sustrato como activadoro Producto final: inhibición por retroalimentación
NOMENCLATURA Y CLASIFICACION
22. Oxidoreductasa: transferencia de electrones23. Transferasa: reacciones de transferencia de
grupos, siempre que no se trate de agua24. Hidrolasa: reacciones de hidrólisis, de
reacción de una sustancia con agua. Ejemplo:25. Liasa: adición de grupos a un doble enlace, o
formación de un doble enlace por eliminación de grupos
26. Isomerasa: tansferencia de grupos dentro de lamisma molécula para dar isómeros.
27. Ligasa: formación de enlaces C-C, C-S, C-O y C-N por reacciones de condensación acoplados a hidrólisis de ATP
El nombre de cada enzima puede ser identificado por:
o Un código numérico encabezado por las letras EC (comisión enzima)
o Cuatro números separados por puntos. El primer número indica a cuál de los seis grupos pertenece

el enzima, el segundo a la subclase dentro de la clase, y el tercero y cuarto, a los grupos químicosespecíficos que intervienen en la reacción.





























