Embed Size (px)
Citation preview

Lactoferrin, myeloperoxidase, and ceruloplasmin:complementary gearwheels cranking physiologicaland pathological processes
Alexey V. Sokolov • Elena T. Zakahrova •
Valeria A. Kostevich • Valeria R. Samygina •
Vadim B. Vasilyev
Received: 24 February 2014 / Accepted: 30 May 2014
� Springer Science+Business Media New York 2014
Abstract Copper-containing plasma protein cerulo-
plasmin (Cp) forms a complex with lactoferrin (Lf), an
iron-binding protein, and with the heme-containing
myeloperoxidase (Mpo). In case of inflammation, Lf
and Mpo are secreted from neutrophil granules.
Among the plasma proteins, Cp seems to be the
preferential partner of Lf and Mpo. After an intraperi-
toneal injection of Lf to rodents, the ‘‘Cp–Lf’’
complex has been shown to appear in their blood-
stream. Cp prevents the interaction of Lf with
protoplasts of Micrococcus luteus. Upon immunopre-
cipitation of Cp, the blood plasma becomes depleted
of Lf and in a dose-dependent manner loses the
capacity to inhibit the peroxidase activity of Mpo, but
not the Mpo-catalyzed oxidation of thiocyanate in
the (pseudo)halogenating cycle. Antimicrobial effect
against E. coli displayed by a synergistic system that
includes Lf and Mpo–H2O2–chloride, but not thiocy-
anate, as the substrate for Mpo is abrogated when Cp is
added. Hence, Cp can be regarded as an anti-inflam-
matory factor that restrains the halogenating cycle and
redirects the synergistic system Mpo–H2O2–chloride/
thiocyanate to production of hypothiocyanate, which
is relatively harmless for the human organism. Struc-
ture and functions of the ‘‘2Cp–2Lf–Mpo’’ complex
and binary complexes Cp–Lf and 2Cp–Mpo in
inflammation are discussed.
Keywords Ceruloplasmin � Lactoferrin �Myeloperoxidase � Protein–protein interactions �Synergism of antimicrobial proteins � Inflammation �Thiocyanate � Halogenative stress
Abbreviations
Cp Ceruloplasmin
Lf Lactoferrin
Mpo Myeloperoxidase
Introduction
Ceruloplasmin (Cp, ferro:O2–oxidoreductase) is the
copper-containing protein of vertebrate blood plasma.
Human Cp has M *132 kDa. Along with the soluble
A. V. Sokolov (&) � E. T. Zakahrova �V. A. Kostevich � V. B. Vasilyev
N-W Branch of the Russian Academy of Medical
Sciences, Institute for Experimental Medicine, Pavlov
Street 12, Saint Petersburg 197376, Russia
e-mail: [email protected]
A. V. Sokolov � V. B. Vasilyev
Saint-Petersburg State University, Mendeleevskaya Line,
Saint Petersburg 199000, Russia
A. V. Sokolov � V. A. Kostevich
Research Institute of Physico-Chemical Medicine,
ul. Malaya Pirogovskaya 1a, Moscow 119435, Russia
V. R. Samygina
Institute of Crystallography, RAS, Leninsky pr 59,
Moscow 117333, Russia
123
Biometals
DOI 10.1007/s10534-014-9755-2

Cp secreted by hepatocytes into plasma, another form
of this protein is known to be anchored by glycosyl-
phosphoinositol to membranes of some cells in the
nervous, immune, and other systems (Salzer et al.
1998; Vassiliev et al. 2005; Marques et al. 2012). Cp
synthesis is increased in response to hypoxia, iron
deficiency (Mukhopadhyay et al. 2000), and copper
excess (Martin et al. 2005). A similar result is envoked
by the rise of insulin (Seshadri et al. 2002), thrombin
(Yang et al. 2006), estradiol (Voronina and Monakhov
1980), and proinflammatory cytokine (Mazumder
et al. 1997) level. A number of enzymatic and anti-
inflammatory activities are characteristic of Cp as an
acute phase reactant (Gitlin 1988). The distinctive
feature of Cp is its capacity to oxidize Fe2? to Fe3?
(Osaki 1966). However, the physiological roles of this
protein are not likely to be reduced to Fe2? oxidation,
even though deficiencies of the Cp gene in humans
(aceruloplasminemia) are known to provoke the
oxidative stress resulting from accumulation of ferrous
iron in tissues (Vassiliev et al. 2005). Cp as the enzyme
actively precludes the formation and persistence of
free radicals, having the activities of ferroxidase
(Osaki 1966), cuproxidase (Stoj and Kosman 2003),
superoxide dismutase (Vasilyev et al. 1988), glutathi-
one-linked peroxidase (Kim and Park 1998), and NO–
oxidase (Shiva et al. 2006). Plasma concentration of
Cp in inflammation can grow from 0.3 to 0.9 mg/ml,
which allows suggesting its role in the regulation of
inflammatory reactions (Glezer et al. 2007).
In the last 10 years we were the first to characterize
complexes of Cp with cationic proteins of neutrophils,
such as lactoferrin (Lf), myeloperoxidase (Mpo)
(Sokolov et al. 2007a), the members of the serprocidin
family (elastase, cathepsin G, proteinase 3 and
azurocidin) (Sokolov et al. 2007b), matrix metallo-
proteinase 2 and 12 (Sokolov et al. 2009a), and
5-lipoxygenase (Sokolov et al. 2010a). Anionic Cp
(pI 4.7) interacts with cationic proteins in a somewhat
similar manner, yet the complexes formed display
certain diversity.
We succeeded in showing the high affinity of
components within the complexes. For example, the
affinity of Cp to Lf and azurocidin is characterized
by Kd *13 nM (Sokolov et al. 2009b, 2010b). Both
in vitro and in vivo Cp is able to form multimeric
complexes that include Lf and Mpo (Sokolov et al.
2007a; Samygina et al. 2013). Lf increases the
ferroxidase activity of Cp upon forming a complex
with the latter (Sokolov et al. 2005a). Considering
that one of the mechanisms of antimicrobial activity
of Lf is its high-affinity binding of Fe3? that is
essential for bacterial growth (Tenovuo 2002), it
cannot be excluded that formation of complexes
with Cp favors sequestration by Lf of iron from the
milieu.
Interaction between Cp and Mpo results in sup-
pression of the prooxidative activity of the leukocytic
enzyme (Segelmark et al. 1997). Participation of Mpo
in protection of the host organism against pathogenic
bacteria is unquestionable (Panasenko et al. 2013), and
yet the enzyme plays an important role in the
development of the halogenative stress associated
with inflammation. Antimicrobial effect of Mpo is
provided by a cycle of reactions in which Mpo reacts
with hydrogen peroxide and is transformed into
Compound I possessing a high two-electron redox
potential (1.16 V). Being highly reactive, Compound I
oxidizes halogenides (Cl-, Br-) and thiocyanate to
respective (pseudo) hypohalide acids, i.e., HOCl,
HOBr, and HOSCN as the enzyme returns to its
native state. Thiocyanate is the most specific substrate
for Mpo (van Dalen et al. 1997). Along with the
(pseudo)halogenating cycle described above, Mpo is
capable of oxidizing a number of substrates using
single-electron transfer (Panasenko et al. 2013) in
course of its peroxidase cycle (e.g., the chromogenic
substrate ABTS (Sokolov et al. 2008).
Our latest data show that the inhibiting effect of Cp
on Mpo depends on the integrity of the copper protein,
i.e., partially proteolyzed Cp is inefficient as an
inhibitor of the chlorinating activity of Mpo (Pana-
senko et al. 2008; Sokolov et al. 2008). Likewise, it
loses the capacity to inhibit synthesis of leukotrienes
catalyzed by 5-lipoxygenase (Sokolov et al. 2010a).
When bound to an intact Cp molecule, Mpo makes the
vulnerable interdomain loop inaccessible to protein-
ases, which protects Cp against the attack of trypsin,
elastase, and plasmin, preventing the cleavage of the
Cp molecule between domains 5 and 6. Proteolytic
cleavage of peptide bonds in Cp beyond the region of
protein–protein interaction was also inhibited when it
formed a complex with Mpo. This may be explained
by a trigger effect: the proteinases are known to
hydrolyze peptide bonds in Cp in a certain order.
Therefore, unless the first one is cleaved, splitting of
other bonds does not occur (Sokolov et al. 2008). This
observation was confirmed when antiatherogenic
Biometals
123

properties of Cp were studied, i.e., solely non-prote-
olyzed Cp was capable of efficient protection of low-
density lipoproteins from proatherogenic modification
being the result of the chlorinating activity of Mpo
(Sokolov et al. 2014).
Cp crystal structure obtained at 2.6-A resolution
distinguishes six ß-barrel homologous domains con-
nected by flexible loops (Samygina et al. 2008). Six
tightly bound copper ions, which can be divided into
three types according to their spectral characteristics,
are distributed irregularly among these six domains.
Domains 2, 4, and 6 contain one type I copper each.
Three copper ions (two type III and one type II) form a
trinuclear cluster with ligands provided by domains 1
and 6. Lf (78 kDa) is composed of two highly
homological sequences known as N- and C-lobes.
Each lobe contains one specific metal-binding site in a
deep cleft between two dissimilar domains (Sun et al.
1999). According to a 1.95-A crystal structure (Blair-
Johnson et al. 2001), Mpo is a homodimer of 140 kDa,
each monomer consisting of two polypeptides of 108
a.a. (light chain) and 466 a.a. (heavy chain) and
containing a heme.
Revealing the sophisticated molecular assembly
including three metal-containing proteins (Cp, Lf, and
Mpo) is a prerequisite for a detailed study of their
interaction with reactive oxygen and halogen species
that are formed in inflammation (Samygina et al.
2013). The damage of Cp in reaction with hydrogen
peroxide (Sokolov et al. 2012a), superoxide anion-
radical, and HOCl (Sharonov et al. 1988, 1989) has
been documented.
Here we present data concerning the specificity
of interaction of Cp with Lf and Mpo. In particular,
this communication is focused on the selectivity of
interaction occurring in the bloodstream between
Cp and exo- and endogenous Lf, on the Mpo-
inhibiting potential of plasma Cp, and on the
interaction of the latter with the synergistic anti-
microbial system containing Lf and Mpo–H2O2–
chloride/thiocyanate.
Materials and methods
The following reagents were used: arginine, glycerol,
Coomassie R-250, mercaptoethanol, ammonium per-
sulfate, Tris (Serva, Germany); SDS, NaSCN, KSCN,
neomycin trisulfate, resazurin, phenylmethylsulfonyl
fluoride (PMSF), 4-chloro-1-naphtol (Sigma, USA);
acrylamide, N,N’-methylene-bis-acrylamide,
N,N,N’,N’-tetramethyleneethylenediamine (Labora-
tory MEDIGEN, Russia); heparin (SPOFA, Poland).
All solutions were prepared using apyrogenic deion-
ized water with resistivity 18.2 MX�cm. Cyanogen
bromide was obtained by bromination of KCN in
biphasic system ‘‘water-dichloroethane’’. The
obtained solution of BrCN in dichloroethane was used
to activate Sepharose for immobilization of neomycin
and heparin (Sokolov et al. 2005b). Molecular mass of
proteins was evaluated in PAAG SDS electrophoresis
(Laemmli 1970).
Optical spectra and the changing absorption rates
were registered on a SF-2000-02 spectrophotometer
(OKB-Spectr, Russia). Concentrations of substances
were measured by spectrophotometry using the fol-
lowing extinction coefficients: dimeric Mpo—
e430 = 178,000 M-1 cm-1 (Bakkenist et al. 1978),
Cp—e610 = 9,780 mM-1 cm-1 (Noyer et al. 1980),
apo-Lf—e280 = 87,360 M-1 cm-1 (Zakharova et al.
2000), H2O2—e240 = 43.6 M-1 cm-1 (Beers and
Sizer 1952).
Protein purification
To obtain a stable preparation of monomeric Cp
containing 95 % of non-fragmented protein with
M 132 kDa and A610/A280 [0.049 human plasma to
which PMSF and EDTA were added, respectively, to
1 mM and to 0.1 mM, was subjected to chromatography
on UNOSphere Q and neomycin-agarose (Sokolov et al.
2012b). Lf was purified from breast milk using ion-
exchange chromatography on CM-Sepharose and gel
filtration on Sephadex G-75 Superfine (Zakharova et al.
2000). Lf from cow milk was isolated similarly. Using
chromatography on heparin-Sepharose, phenyl-Sephar-
ose, and gel filtration on Sephacryl S-200 HR, Mpo
preparation was purified from human leukocytes to the
ratio A430/A280 (RZ) = 0.85, which characterizes the
homogeneous protein (Sokolov et al. 2010c).
Revealing heterologous complexes of Lf and Cp
after intraperitoneal injection of Lf
Wistar rats and mice C57Black were narcotized with
ether and injected intraperitoneally, respectively,
20 mg and 1 mg of Lf from cow or breast milk. Blood
Biometals
123

was sampled from the tail vein 15, 0, 60, and 120 min
past the injection. Animal serum (5 ll) was analyzed
by SDS-free disc-electrophoresis (Davis 1964) and
then by Western blotting (Anderson et al. 1982) with
antibodies against CP (rat or mouse) and Lf (bovine or
human).
Evaluation of the effect of Cp on Mpo activity
Electrochemical measurements of the rate of H2O2
concentration decrease were done with planar sensors
constructed on the basis of Prussian blue nanofilms
(Borisova et al. 2009). Our device included the planar
electrode-containing unit placed on a permanently func-
tioning magnetic stirrer, and the potentiostat
P-8 Elins (Chernogolovka, Russia) that registered the
electric current proportional to H2O2 concentration. The
initial current strength was registered upon introducing
H2O2 to 50 lM into the medium containing 100 mM KCl,
10 mM potassium phosphate buffer, pH 6.2, 10 mM Tau.
The reaction was launched by adding an aliquot of Mpo (to
the final concentration 1–5 nM) and the time-dependent
current strength was tracked. The rate of H2O2 concentra-
tion decrease and the turnover number (1/s) for the
catalytic center of Mpo were determined using the linear
part of the plot. Also, the effect on H2O2 utilization
resulting from adding to this system of 0.2 mM KSCN and
of 0.2, 0.4, 0.8 b 1.6 lM Cp was quantified. Graphs
reflecting the effect of Cp on Mpo-catalyzed H2O2
utilization in presence of various substrates were plotted.
Peroxidase activity of Mpo was assayed by oxidation of
the chromogenic substrate sodium 2,20-diasinobis(3-
ethylbenzotriazoline-6-sulphonate) (ABTS) (Sokolov
et al. 2008). Oxidation of the substrate gives origin to the
stable radical ABTS•?, the amount of which is measured
by light absorption. The reaction mixture for this assay
contained 3 nM Mpo, 100 lM H2O2, 1 mM ABTS in
100 mM sodium acetate buffer, pH 5.5. Upon adding
H2O2 to the mixture, the activity of Mpo was assayed at
room temperature as DA414/min having set the ‘‘Kinetics’’
mode in SF 2000–2002. Peroxidase activity of Mpo was
measured upon adding samples of blood plasma contain-
ing various amounts of Cp, after its immunoprecipitation.
Sample dilution providing IC50 was determined.
Immunoprecipitation of Cp from plasma
To achieve immunoprecipitation, we incubated blood
plasma with increasing concentrations of high-affinity
rabbit antibodies against human Cp (Sokolov et al.
2010c), which was followed by precipitation of an
immune complex by goat antibodies against rabbit
immunoglobulins. In 30 min of plasma incubation with
rabbit affinity antibodies against Cp (0.1–2 mg/ml), the
immunoprecipitation of Cp was accomplished. Then
goat antibodies (2 mg/ml) against rabbit immunoglob-
ulins were added with subsequent incubation for
30 min. Samples were centrifuged for 10 min at
15,000 9 g (4 �C). Cp content in plasma after immu-
noprecipitation was assayed by ELISA (Sokolov et al.
2010c), Lf content was assayed by commercial ELISA
(Vector-Best, Russia).
Agglutination of Micrococcus luteus protoplasts
Agglutination of Micrococcus luteus protoplasts was
followed by changes in A450 using a model system
containing 0.5 % suspension of freeze-dried M. luteus,
5 lg/ml lysozyme, 0.5 mg/ml Lf in 0.1 M NaCl,
66 mM sodium acetate buffer, pH 5.4 (Perraudin and
Prieels 1982). Cp was added to molar ratio with Lf 2:1,
1:1, and 1:2, and its effect was studied.
Circular dichroism spectroscopy
Circular dichroism (CD) spectra were registered on a
CD6 dichrograph (Jobin–Yvon, France), calibrated
with (?)-10-D-camphora-sulfonic acid. Measure-
ments were made using dismountable 0.05-cm cuv-
ettes (near-UV and visible regions) and 0.001-cm
cuvettes (far-UV). To carry out these measurements,
5 lM Mpo, 10 lM Cp, 10 lM Lf, and mixtures of
proteins corresponding to complexes 2Cp:1Mpo and
2Lf:2Cp:1Mpo were used.
Antimicrobial activity (MIC50)
The slowing growth of Escherichia coli, strain ML-35p,
was evaluated by absorption spectrum of the metabolic
indicator resazurin, which allowed estimating the
antimicrobial activity of Lf and of the system Mpo–
H2O2–chloride/thiocyanate (Cooper 2013). Cells were
cultured overnight at ?37 �C in 3 % soybean tryptic
hydrolysate. Thus, grown suspension of E. coli culture
was centrifuged at 6,000 9 g for 5 min, then washed
with PBS cooled to ?4 �C, and again centrifuged under
the same conditions. Cell precipitate was resuspended in
PBS, after which the concentration of cells was
Biometals
123

determined by measuring A620, on account that
2.5 9 108 CFU/ml corresponds to one optical density
unit. A 96-well flat-bottom plate was filled with studied
substances in PBS, i.e., the bacterial suspension (final
concentration in a well was 4 9 104 CFU/ml), soybean
tryptic hydrolysate (final concentration 0.18 %), 30 lM
resazurin. Lf and Mpo in the presence of H2O2 (10 lM)
and sodium thiocyanate (10 lM) were tested as anti-
microbial agents. Proteins (Cp, Lf bMpo) were added in
amounts providing the ratio 2Lf:2Cp:1Mpo. The plate
was put on a shaker at ?37 �C. The metabolic activity of
bacteria was evaluated by the growth of A530–A630. This
index was registered every 30 min in a multichannel
spectrophotometer Stat Fax (USA). The antimicrobial
activity was expressed as the protein concentration that
caused a two-fold drop of A530–A630 as compared to the
control cell culture (MIC50).
Interaction between protein regions analyzed using
a 3D model of complex
To reveal the sites of interaction in Cp and Lf previous
3D models obtained in a SAXS study of the 2Lf–2Cp–
Mpo complex (Samygina et al. 2013) was used.
Results
The specificity of interaction between Cp and Lf
To explore the selectivity of interaction between Lf
and Cp we used a heterologous system, when either
bovine or human Lf was injected intraperitoneally to
rats and mice, after which electrophoretic mobility of
Cp and Lf was revealed by Western blotting of SDS-
free PAAG in which samples of serum had been
subjected to disc-electrophoresis (Fig. 1). It is seen
that 15 min after injection of Lf into animals the
immunoreactive band corresponding to rat or mouse
Cp had an altered mobility as compared to the serum
sampled before injection. The novel mobility con-
forms to that of the Cp–Lf complex which is formed
when Lf is added to either rat or mouse serum
(Fig. 1a). Lf (either bovine or human) detected in sera
of rats and mice also migrated with the speed of the
Cp–Lf complex (Fig. 1b). The intensity of the band
corresponding to the heterologous Cp–Lf complex did
not change for 2 hours. No other part of a nitrocellu-
lose membrane bound antibodies to human or bovine
Lf hence, Cp is likely to be the preferable partner of Lf.
We have shown previously that interaction of Cp with
Lf is prevented by polyanionic substances bound to the
N-terminal polycationic cluster in Lf, such as LPS,
DNA, and heparin (Pulina et al. 2002).
A study of Lf-mediated agglutination of M. luteus
protoplasts showed that Cp blocks this process
(Fig. 2). In the presence of Lf, instead of monotonous
decrease of turbidity, a temporary increase of A450 is
observed, which results from agglutination of protop-
lasts. In the presence of Cp that peak goes down in a
dose-dependent manner, so that the curve practically
coincides with that observed when protoplasts are
lyzed essentially by pure lysozyme.
We studied Lf content in plasma samples subjected
to immunoprecipitation of Cp. It appeared that plasma
Lf concentration goes down concomitantly with pre-
cipitation of Cp. Hence, upon precipitation, 98 % of Cp
the plasma became depleted of Lf for about 92 %
(Fig. 3a). Control experiments showed that antibodies
against Cp alone cause no precipitation of Lf.
The specificity of Mpo inhibition by Cp
Our previous study provided evidence that introduc-
tion of human Mpo into the bloodstream of rats also
Fig. 1 Detection of mouse and rat Cp (a), of human and bovine
Lf (b) before and after the intraperitoneal injection into mice and
rats of 1 and 20 mg Lf, respectively. Western blotting of human
serum samples (5 ll) after SDS-free electrophoresis. 1—serum
before injection of Lf, 2–5 to 15, 30, 60, and 120 min after
injecting Lf, 6—blood serum after adding Lf (0.5 lg)
Biometals
123

results in forming a heterologous complex (Sokolov
et al. 2007a). Besides, upon adding excessive amounts
of Mpo to human plasma, it got bound to Cp, including
the incorporation into multimeric LDL/VLDL-con-
taining complexes (Sokolov et al. 2010c).
Cp is likely to be the physiological inhibitor of
Mpo. We studied the effect of immunoprecipitation of
Cp from plasma (adding varying amounts of affinity
antibodies against Cp) on inhibition of Mpo activity by
such plasma. This approach allowed obtaining plasma
preparations varying in Cp content (from 80 nM to
3.4 lM). In every such case, a dilution corresponding
to IC50 for peroxidase activity of Mpo with ABTS was
determined. The direct approximating dependence of
plasma dilution on Cp concentration appeared to get
interpolated to zero (Fig. 3b). This is evidence of Cp
being the major inhibitor of Mpo peroxidase activity in
blood plasma. However, an electrochemical sensor
used to study the effect of added plasma on the rate of
hydrogen peroxide utilization allowed showing that
the rate of H2O2 utilization is not decreased propor-
tionally to the degree of the Mpo peroxidase activity
inhibition provided by that same portion of plasma
(Fig. 3c). When under the same conditions, plasma
preparations containing varying amounts of Cp were
used, no effect on the rate of H2O2 utilization was
observed (data not shown). Therefore, adding plasma
capable of inhibiting Mpo peroxidase activity (by
virtue of interacting with Cp) does not prevent to the
same extent hydrogen peroxide utilization.
We suggested that Cp cannot counteract the
oxidation of some plasma-contained substrate in the
halogenating cycle of Mpo, e.g., of thiocyanate.
Indeed, when H2O2 utilization by chlorinating Mpo
in the presence of Cp and SCN- was measured, it
appeared that Cp does not preclude the utilization of
hydrogen peroxide in course of Mpo-catalyzed oxi-
dation of SCN- to HOSCN (Fig. 3d).
CD spectra of the 2Cp–Mpo and of the 2Lf–2Cp–
Mpo complexes showed noticeable changes in the
ellipticity of the heme in Mpo, occurring upon forming
of a complex, once the experimental spectra and their
arithmetic sum were compared (Fig. 4). For instance,
a significant decrease of the ellipticity was registered
in the far-UV region with a shift of the minimum from
284 nm in Mpo to 281 nm in the 2Cp–Mpo and the
2Lf–2Cp–Mpo complexes, while no shift of that was
observed in the arithmetic sum of the spectra (Fig. 4a).
In the spectra of Mpo, of its complex with Cp and of
the triple complex the ellipticity in the region of the
Fig. 2 Effect of Cp on Lf-dependent agglutination of 0.5 %
cell suspension of M. luteus in the presence of lysozyme,
evaluated by an increase of the solution transparency (A450). The
system contained 5 lg/ml lysozyme, 0.5 mg/ml Lf in 0.1 M
NaCl, 66 mM sodium phosphate buffer, pH 5.4, and portions of
Cp: 4, 2, and 1 mol per mole of Lf. 1 lysis caused by lysozyme, 2
in the presence of lysozyme and Lf, 3 in the presence of
lysozyme, and Lf with Cp (2:1), 4 in the presence of lysozyme,
and Lf with Cp (1:1), 5 in the presence of lysozyme, and Lf with
Cp (1:2)
Biometals
123
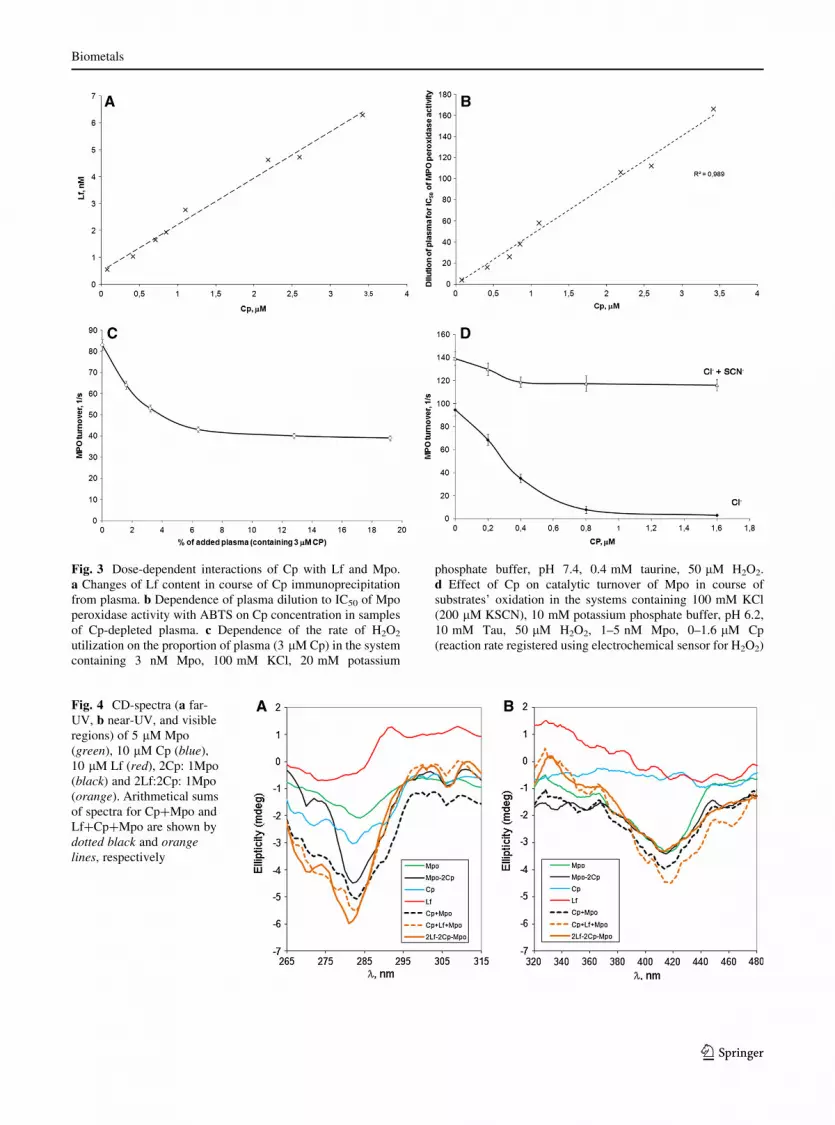
Fig. 3 Dose-dependent interactions of Cp with Lf and Mpo.
a Changes of Lf content in course of Cp immunoprecipitation
from plasma. b Dependence of plasma dilution to IC50 of Mpo
peroxidase activity with ABTS on Cp concentration in samples
of Cp-depleted plasma. c Dependence of the rate of H2O2
utilization on the proportion of plasma (3 lM Cp) in the system
containing 3 nM Mpo, 100 mM KCl, 20 mM potassium
phosphate buffer, pH 7.4, 0.4 mM taurine, 50 lM H2O2.
d Effect of Cp on catalytic turnover of Mpo in course of
substrates’ oxidation in the systems containing 100 mM KCl
(200 lM KSCN), 10 mM potassium phosphate buffer, pH 6.2,
10 mM Tau, 50 lM H2O2, 1–5 nM Mpo, 0–1.6 lM Cp
(reaction rate registered using electrochemical sensor for H2O2)
Fig. 4 CD-spectra (a far-
UV, b near-UV, and visible
regions) of 5 lM Mpo
(green), 10 lM Cp (blue),
10 lM Lf (red), 2Cp: 1Mpo
(black) and 2Lf:2Cp: 1Mpo
(orange). Arithmetical sums
of spectra for Cp?Mpo and
Lf?Cp?Mpo are shown by
dotted black and orange
lines, respectively
Biometals
123

Soret band (maximum at 412 nm) is virtually the same
between 405 and 425 nm (Fig. 4b). However, when
the experimental spectra of complexes are compared
with the arithmetic sum of the proteins’ spectra, the
heme ellipticity in complexes of Mpo with Cp and Lf
appears to be greater than in Mpo alone.
Effect of Cp on antimicrobial activity of Lf
and Mpo
The results of assaying the antimicrobial activity of Lf,
of the system Mpo–H2O2–chloride/thiocyanate, of
their combined effect, and of the influence of Cp, are
summarized in Table 1. It is seen that Cp blocks the
antimicrobial effect of Mpo once chloride becomes the
substrate of the latter. Besides, in the presence of Cp,
the synergizing effect of such a system with Lf goes
down to the activity of Lf acting separately. These
results are in good agreement with the data on
inhibited chlorinating activity of Mpo obtained by
measuring the luminol-dependent chemiluminescence
when the enzyme forms a complex with other proteins
(Panasenko et al. 2008). On the other hand, Cp had no
effect on the system Mpo–H2O2–thiocyanate and did
not alter the synergism of Mpo and Lf in the presence
of thiocyanate as the substrate of the (pseudo)haloge-
nating cycle of Mpo. These results are in full
agreement with the incapacity of Cp to inhibit the
Mpo-dependent production of hypothiocyanate.
Discussion
Our results allow concluding that Cp, Lf, and Mpo
function in concord like tooth-wheels in a chain gear,
and the functions of each metal-containing protein
composing the complex are interactively aimed at
decreasing the oxidative/halogenative stress accom-
panying inflammation (Fig. 5). The interaction of Cp
with Mpo and Lf was discovered only at the turn of the
XX and XXI centuries. However, of all the antioxidant
proteins of blood plasma, Cp was shown to possess the
highest potential as the scavenger of superoxide anion
radical and hypochlorous acid which are known,
respectively, as the substrate and the product of
reaction catalyzed by Mpo (Sharonov et al. 1988,
1989). For instance, Cp was identified as the protein
adsorbed on immobilized Mpo and capable of sup-
pressing its peroxidase activity (Segelmark et al.
1997). Direct interaction of Cp and Lf was docu-
mented in our laboratory when studying the properties
of Cp in breast milk. It turned out that Lf is retained on
immobilized Cp during the chromatography of breast
milk and once these two proteins are mixed, they form
a complex with the same electrophoretic mobility as
the abnormal mobility of Cp in breast milk (Zakharova
et al. 2000; Sokolov et al. 2006). We were the first to
show that after injection of Lf and Mpo into the rat
bloodstream, these proteins form heterologous com-
plexes with the host Cp (Zakharova et al. 2000;
Sokolov et al. 2007a).
The Cp–Lf complex was isolated from breast milk
and lacrimal fluid of healthy donors (Sokolov et al.
2006, 2013). Complexes composed of Cp, Lf, and
Mpo at a time can be formed in vitro upon mixing the
purified proteins, but also are found in biological fluids
obtained from patients with inflammatory diseases
(Sokolov et al. 2007a). Both Lf and Mpo were shown
to interact with a number of other plasma proteins,
such as albumin (Lampreave et al. 1990; Tiruppathi
et al. 2004). Considering these data, we analyzed the
selectivity of interaction of Cp with Lf after intraperi-
toneal injections of human and bovine LF to mice and
rats (Fig. 1).
We have observed previously the peculiar changes in
electrophoretic mobility of Cp upon adding Lf to a
Table 1 Effect of Cp on antimicrobial activity of Lf and of the
system Mpo–H2O2–chloride/thiocyanate with respect to E. coli
System under study MIC50a , nM
Cp Cannot be determined
apo-Lf 980 ± 40
apo-Lf: Cp 990 ± 30
Mpo (H2O2–chloride) 45 ± 12
2Cp: Mpo (H2O2–chloride) Cannot be determined
2apo-Lf: 1Mpo (H2O2–chloride) 4 ± 1
2: Cp: 2apo-Lf: 1Mpo
(H2O2–chloride)
860 ± 30
Mpo (H2O2–thiocyanate) 38 ± 11
2Cp: Mpo (H2O2–thiocyanate) 42 ± 9
2apo-Lf: 1Mpo (H2O2–thiocyanate) 7 ± 2
2: Cp: 2apo-Lf: 1Mpo
(H2O2–thiocyanate)
8 ± 2
a Expressed as concentration of the protein that was present in
the system at a lower concentration as compared to other
components, which resulted in a two-fold decrease of A530–
A630 from the control level
Biometals
123

sample of blood plasma, however, such observations
provided no evidence of the extent to which Lf is engaged
in complexes with Cp and other plasma components. In
this study we revealed no other Lf-positive electropho-
retic band except its complex with either mouse or rat Cp.
The absence of strict species specificity in interaction of
these proteins indicates certain evolutionary conserva-
tism of such complex. Similar results were obtained in
our study of interaction of human and canine Mpo with
Cp of a number of mammalian species (Sokolov et al.
2007a).
Our observation that practically all the Lf of plasma
co-immunoprecipitates with Cp (concentration of the
former drops from 6.3 to 0.55 nM, see Fig. 3a)
indicates that interaction of these two proteins can
take place in plasma under normal conditions. This
means that Kd 13 nM determined in a model system
with Sepharose-immobilized Cp (Sokolov et al.
2009b) in reality can be even lower.
Structural studies of complexes formed by Cp with
Lf and Mpo indicate the stoichiometry 2Cp–2Lf–Mpo
(Sokolov et al. 2009c; Samygina et al. 2013). Mean-
while, such methods as SAXS, laser correlation
spectroscopy, and fluorescence studies showed the
presence of a complex with 1:1 stoichiometry in
solution (Sabatucci et al. 2007; Sokolov et al. 2009c;
Ha-Doong et al. 2010). Lf participates in forming the
complex 2Lf–2Cp–Mpo and, using a number of
mechanisms, in this way restricts the production of
reactive oxygen species. Firstly, it enhances the
oxidation of Fe2? by Cp and thus decreases the
oxidative potential of the iron pool (Sokolov et al.
2005a, 2009b). Secondly, it binds Cu2?, which
precludes the production of hydroxyl radicals in
course of H2O2-induced degradation of Cp (Sokolov
et al. 2012a). Lastly, Lf does not hamper the inhibition
by Cp of the chlorinating activity of Mpo (Panasenko
et al. 2008) and binds Fe3?; on the whole this
decreases the production of hydroxyl radicals in
reaction of HOCl with Fe2?.
The physiological role of Cp as the inhibitor of Mpo
activity is beyond doubt (Segelmark et al. 1997;
Sokolov et al. 2008; Chapman et al. 2013). Cp inhibits
Mpo even in the presence of C-reactive protein, which
also interacts with Mpo (Xu et al. 2013).
The activity of Mpo provoking the halogenative
stress in inflammation (Panasenko et al. 2008) is
important for antimicrobial protection of an organism.
However, the role of this enzyme does not seem to be
limited exclusively to the antibacterial defense. Indi-
viduals with autoantibodies against Mpo (ANCA)
causing dissociation of its complex with Cp suffer
from systemic vasculitis (Griffin et al. 1999; Xu et al.
2012). On the other hand, hereditary deficiency of
Mpo results in the development of candidosis (Lehrer
and Cline 1969).
Fig. 5 Scheme of influence
of Cp (blue arrows), Mpo
(green arrows), Lf (red
arrows) on functions of each
other due to interactions
Biometals
123

Mpo has no effect on the activity of Cp (Park et al.
2000), except for the capacity to enhance its oxidase
activity in reaction with p-phenylene diamine (Soko-
lov et al. 2008). As shown by an X-ray study of the
2Cp–Mpo complex, in this case the effect of Mpo
results from its direct contact with the binding site of
p-phenylene diamine in Cp (Samygina et al. 2013).
Each of the two protomers of Mpo interacts with one
Cp molecule in the 2Cp–Mpo complex.
This study has shown that upon elimination of Cp
by immunoprecipitation blood plasma loses much of
its capacity to inhibit the peroxidase activity of Mpo
(Fig. 3b), but retains the ability to utilize hydrogen
peroxide in Mpo-catalyzed oxidation of substrates that
can be regarded as more physiological (Fig. 3c).
Several substrates of Mpo-catalyzed peroxidase reac-
tion are known to be present in blood plasma, e.g.,
tyrosine, urate, ascorbate, and nitrite (Vlasova et al.
2012). Along with those, however, the most specific
substrate of the halogenating cycle of Mpo can be
found, which is thiocyanate (van Dalen et al. 1997).
Yet, in our experiments, the presence of Cp had no
inhibitory effect on Mpo-catalyzed oxidation of thio-
cyanate (Fig. 3d) performed with the minimum Km
and the highest specificity for the enzyme (van Dalen
et al. 1997). It has been suggested that inhibition of
Mpo by Cp is realized via direct contact of the peptide
loop of the latter (amino acids 883–892) with the heme
pocket of Mpo (Samygina et al. 2013). This means a
competition between Cp and various substrates for the
access to the active center of Mpo. Taking into account
the results described above, SCN- seems to be a more
successful competitor than Cp. We have shown
previously that the efficiency of inhibition by Cp of
Mpo peroxidase activity depends on the dimensions of
a peroxidized substrate, but also on the integrity of the
peptide loop in Cp (a.a. 883–892) connecting domains
5 and 6 (Sokolov et al. 2008). Properly this loop, as
judged by the X-ray studies, directly contacts with the
entrance into the heme pocket of Mpo, and the
synthetic peptide RPYLKVFNPR mimicking its
amino acid sequence efficiently inhibits the Mpo
peroxidase activity (Samygina et al. 2013).
CD spectra showed that when Mpo forms com-
plexes with Cp and Lf, the ellipticity of its heme
changes (Fig. 4). This observation supports the notion
of interaction of Cp with the entrance into the heme
pocket in Mpo, while Lf does not prevent Cp from
inhibiting the chlorinating activity of Mpo (Panasenko
et al. 2008). In line with this observation are the data
on the Mpo heme ellipticity changes occurring upon
forming a complex with low-density lipoproteins,
which increases the chlorinating activity of Mpo
(Delporte et al. 2014).
A competitive mechanism of Mpo inhibition
seems to be inefficient when such a specific substrate
as thiocyanate is oxidized. It should be noted that Km
for hydrogen peroxide does not change in the presence
of Cp (Sokolov et al. 2008). Another important notion
is that the catalytic turnover of Mpo (ca. 90/s) is two
orders higher than that of Cp (0.5/s) when it oxidizes
its most specific substrate, i.e., Fe2? (Stoj and Kosman
2003). In view of all of this, Cp has poor chances to
reduce the highly reactive Compound I, contrary to the
suggestion of Chapman et al. (2013).
It seems important that by redirecting the activity of
Mpo at oxidizing thiocyanate, but not chloride, Cp
favors the production of hypothiocyanate, an efficient
antimicrobial agent, though relatively harmless spe-
cies for the host organism (Chandler and Day 2012).
Antimicrobial synergism of Lf and Mpo function-
ing in the (pseudo)halogenating cycle is well known
(Kokryakov 1999; Tenovuo 2002). Cp interfered with
the antimicrobial effect of Mpo only when chloride
was the substrate of the halogenating cycle. However,
it did not suppress the antimicrobial activity of the
system when, in PBS buffer, production of hypothi-
ocyanate was possible in the (pseudo)halogenating
cycle of Mpo (Table 1). The fact that Cp does not
interfere with the antimicrobial synergism of Lf and
Mpo and redirects the system towards synthesis of the
relatively harmless antimicrobial agent, i.e., hypoth-
iocyanate (Table 1), favors the notion of participation
of the complex 2Cp–2Lf–Mpo in the protection of an
organism against the halogenative stress developed in
inflammation.
Considering the results of Lf displacement from its
complex with Cp by polyanionic structures (DNA, LPS,
heparin) and by peptides mimicking the N-terminal
cationic cluster of Lf (RRRR), it can be concluded that
the site of interaction with Cp is located at the
N-terminus of Lf (Pulina et al. 2002; Sokolov et al.
2006). However, anionic peptides homologous to amino
acid stretches in Cp, such as DQVDKEDEDFQE
(586–597), EVEWDYSPQREWE (721–734), and
DENESWYLDD (905–914) did not displace this pro-
tein from its complex with Lf (Sokolov et al. 2006).
These results are at variance with the data on interaction
Biometals
123

of Lf with two similar peptides, i.e., YYIAAVEVEW-
DYS (715–727) and FDENESWYLDDNI (904–916),
obtained in experiments with absorption of labeled Lf on
peptide library of Cp (White et al. 2012). In our recent
model of the Cp–Lf complex (Samygina et al. 2013),
these two stretches are not included in the contact area
with Lf (Fig. 6a). Cp contacts Lf using the amino acid
stretches from its domains 1 and 6, which contain
ligands for copper ions of trinuclear cluster in Cp
(Fig. 6b). As shown by SAXS studies, there is no direct
contact between Lf and Mpo within the ternary com-
plex. On the one hand, this is in line with the observation
that Lf does not affect the activity of Mpo (Panasenko
et al. 2008), and on the other hand, it shows that the
enzyme’s activity is likely to be inhibited by the
N-terminal peptide of Lf (1–11) only if the latter is
proteolyzed, but not intact Lf (van der Does et al. 2012).
Protective effect of the Cp–Lf complex was doc-
umented when both proteins were applied as antiox-
idants in treatment of patients with malignancies
(Edeleva et al. 2001). Experimental data on involution
of mammary gland in mice provided evidence that
genes encoding Cp and Lf become activated at that
period (Nakamura et al. 2006). The Cp–Lf complex is
found in breast milk, which is more evidence of the
protective effect of these two proteins (Sokolov et al.
2006).
Our recent study (Zakharova et al. 2012) showed
that apo-Lf has a pronounced anti-hypoxic effect and
possesses the properties of a physiological mimetic of
hypoxia as it stimulates the synthesis of Cp and
erythropoietin by stabilizing the hypoxia-inducible
factor 1-alpha (Zakharova et al. 2012). Iron-saturated
Lf has no such features, which allows suggesting a
negative feedback in regulation of the system that
includes Cp and apo-Lf. Firstly, apo-Lf increases the
ferroxidase activity of Cp and becomes saturated with
Fe3? (Sokolov et al. 2005a, 2009b). Secondly, apo-Lf
triggers the synthesis of Cp and erythropoietin, which
favors an increase of the plasma ferroxidase activity
(egress of iron from tissue storages) and stimulates
erythropoiesis. These two mechanisms provide a good
explanation of the anti-anemic properties of apo-Lf
(Pulina et al. 2010; Zakharova et al. 2012). Moreover,
once saturated with iron, Fe2–Lf is unable to activate
the hypoxia-inducible synthesis of Cp and erythropoi-
etin (Zakharova et al. 2012).
Under conditions of focal inflammation and poor
oxygenation, Cp can become a factor that favors iron
binding by apo-Lf with further realization of the
antimicrobial function of the latter. The selectivity of
interaction of the three metal-containing proteins, i.e.,
Cp, Lf, and Mpo, which participate in inflammatory
reactions and antimicrobial defense of an organism,
does not seem accidental, since Cp is the preferred
Fig. 6 Details of the interaction sites in Cp and Lf. a Labile
Fe(II) binding sites (LS1 and LS2, red), anionic peptides
715–727 and 904–916 (blue) in Cp and Fe(III) binding sites in
Lf (violet). b N-terminal cationic peptides 2–5 and 28–32
(magenta) in Lf and amino acid 50–109 and 929–1,012 stretches
(blue) containing ligands for copper ions of trinuclear cluster
(yellow triangle) in Cp
Biometals
123

partner of Lf among other plasma proteins and is
capable of specific inhibition of the peroxidase and
chlorinating activities of Mpo, leaving unaffected the
production of hypothiocyanate, an antimicrobial
agent.
Acknowledgments This study was supported by RFBR grants
§ 12-04-00301; 13-04-01191, MK-6062.2014.4 and by the
Program ‘‘Human Proteome’’. The authors are grateful to
Professor V. N. Kokryakov for generously providing leukocytes
of healthy donors, to Dr. M. N. Berlov for kind assistance in
mastering the evaluation of antimicrobial activity of proteins, to
Dr. M. O. Pulina and Dr. A. N. Skvortsov for CD-spectra
measurement.
References
Anderson NL, Nance SL, Pearson TW et al (1982) Specific
antiserum staining of two-dimensional electrophoretic
patterns of human plasma proteins immobilized on nitro-
cellulose. Electrophoresis 3:135–142
Bakkenist AR, Wever R, Vulsma T et al (1978) Isolation pro-
cedure and some properties of myeloperoxidase from
human leucocytes. Biochim Biophys Acta 524:45–54
Beers RF Jr, Sizer IW (1952) A spectrophotometric method for
measuring the breakdown of hydrogen peroxide by cata-
lase. J Biol Chem 195:133–140
Blair-Johnson M, Fiedler T, Fenna R (2001) Human myelo-
peroxidase: structure of a cyanide complex and its inter-
action with bromide and thiocyanate substrates at 1.9 A
resolution. Biochemistry 4:13990–13997
Borisova AV, Karyakina EE, Cosnier S et al (2009) Current-free
deposition of Prussian blue with organic polymers: towards
improved stability and mass production of the advanced
hydrogen peroxide transducer. Electroanalysis 21:409–414
Chandler JD, Day BJ (2012) Thiocyanate: a potentially useful
therapeutic agent with host defense and antioxidant prop-
erties. Biochem Pharmacol 84:1381–1387
Chapman AL, Mocatta TJ, Shiva S et al (2013) Ceruloplasmin is
an endogenous inhibitor of myeloperoxidase. J Biol Chem
288:6465–6477
Cooper RA (2013) Inhibition of biofilms by glucose oxidase,
lactoperoxidase and guaiacol: the active antibacterial
component in an enzyme alginogel. Int Wound J 10:
630–637
Davis BJ (1964) Disc electrophoresis. II. Method and applica-
tion to human serum proteins. Ann N Y Acad Sci 121:
404–427
Delporte C, Zouaoui Boudjeltia K, Noyon C et al (2014) Impact
of interaction between myeloperoxidase and low-density
lipoprotein on the specific activity of the enzyme and
subsequent post-translational oxidative modifications of
apolipoprotein B-100. J Lipid Res 55:747–757
Edeleva NV, Sergeeva TV, Nemtsova ER et al (2001) Antiox-
idants ceruloplasmin and lactoferrin in the prevention and
treatment of postoperative complications in cancer
patients. Anesteziol Reanimatol 5:61–64
Gitlin JD (1988) Transcriptional regulation of ceruloplasmin gene
expression during inflammation. J Biol Chem 263:6281–6287
Glezer I, Chernomoretz A, David S et al (2007) Genes involved
in the balance between neuronal survival and death during
inflammation. PLoS ONE 2:e310
Griffin SV, Chapman PT, Lianos EA et al (1999) The inhibition
of myeloperoxidase by ceruloplasmin can be reversed by
anti-myeloperoxidase antibodies. Kidney Int 55:917–925
Ha-Doong NT, Eid C, Hemadi M et al (2010) In vitro interaction
between ceruloplasmin and human serum transferring.
Biochemistry 49:10261–10263
Kim IG, Park SY (1998) Requirement of intact human cerulo-
plasmin for the glutathione-linked peroxidase activity.
FEBS Lett 437:293–296
Kokryakov VN (1999) Biology of antibiotics of animal origin
[in Russian]. Nauka, St. Petersburg
Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:
680–685
Lampreave F, Pineiro A, Brock JH et al (1990) Interaction of
bovine lactoferrin with other proteins of milk whey. Int J
Biol Macromol 12:2–5
Lehrer RI, Cline MJ (1969) Leukocyte myeloperoxidase defi-
ciency and disseminated candidiasis: the role of myelo-
peroxidase in resistance to Candida infection. J Clin Invest
48(8):1478–1488
Marques L, Auriac A, Willemetz A et al (2012) Immune cells
and hepatocytes express glycosylphosphatidylinositol-
anchored ceruloplasmin at their cell surface. Blood Cells
Mol Dis 48(2):110–120
Martin F, Linden T, Katschinski DM et al (2005) Copper-
dependent activation of hypoxia-inducible factor (HIF)-1:
implications for ceruloplasmin regulation. Blood 105:
4613–4619
Mazumder B, Mukhopadhyay CK, Prok A et al (1997) Induction
of ceruloplasmin synthesis by IFN-gamma in human
monocytic cells. J Immunol 159:1938–1944
Mukhopadhyay CK, Mazumder B, Fox PL (2000) Role of
hypoxia-inducible factor-1 in transcriptional activation of
ceruloplasmin by iron deficiency. J Biol Chem 275:21048–
21054
Nakamura M, Tomita A, Nakatani H et al (2006) Antioxidant
and antibacterial genes are upregulated in early involution
of the mouse mammary gland: sharp increase of cerulo-
plasmin and lactoferrin in accumulating breast milk. DNA
Cell Biol 25:491–500
Noyer M, Dwulet FE, Hao YL et al (1980) Purification and
characterization of undegraded human ceruloplasmin.
Anal Biochem 102:450–458
Osaki S (1966) Kinetic studies of ferrous ion oxidation with
crystalline human ferroxidase (ceruloplasmin). J Biol
Chem 241:5053–5059
Panasenko OM, Gorudko IV, Sokolov AV (2013) Hypochlorous
Acid as a precursor of free radicals in living systems.
Biochemistry (Mosc) 78:1466–1489
Panasenko OM, Chekanov AV, Vlasova II et al (2008) A study
of the effect of ceruloplasmin and lactoferrin on the chlo-
rination activity of leukocytic myeloperoxidase using the
chemiluminescence method. Biofizika 53:573–581
Park YS, Suzuki K, Mumby S et al (2000) Antioxidant binding
of ceruloplasmin to myeloperoxidase: myeloperoxidase is
Biometals
123

inhibited, but oxidase, peroxidase and immunoreactive
properties of ceruloplasmin remain intact. Free Radic Res
33:261–265
Perraudin JP, Prieels JP (1982) Lactoferrin binding with lyso-
zyme-treated Micrococcus luteus. Biochim Biophys Acta
718:42–48
Pulina MO, Zakharova ET, Sokolov AV et al (2002) Studies of
the ceruloplasmin-lactoferrin complex. Biochem Cell Biol
80:35–39
Pulina MO, Sokolov AV, Zakharova ET et al (2010) Effect of
lactoferrin on consequences of acute experimental hem-
orrhagic anemia in rats. Bull Exp Biol Med 149:219–222
Sabatucci A, Vachette P, Vasilyev VB et al (2007) Structural
characterization of the ceruloplasmin: lactoferrin complex
in solution. J Mol Biol 371:1038–1046
Salzer JL, Lovejoy L, Linder MC et al (1998) Ran-2, a glial
lineage marker, is a GPI-anchored form of ceruloplasmin.
J Neurosci Res 54:147–157
Samygina VR, Sokolov AV, Pulina MO et al (2008) X-ray
diffraction study of highly purified human ceruloplasmin.
Crystallogr Rep 53:655–662
Samygina VR, Sokolov AV, Bourenkov G et al (2013) Ceru-
loplasmin: macromolecular assemblies with iron-contain-
ing acute phase proteins. PLoS ONE 8:e67145
Segelmark M, Persson B, Hellmark T et al (1997) Binding and
inhibition of myeloperoxidase (MPO): a major function of
ceruloplasmin? Clin Exp Immunol 108:167–174
Seshadri V, Fox PL, Mukhopadhyay CK (2002) Dual role of
insulin in transcriptional regulation of the acute phase
reactant ceruloplasmin. J Biol Chem 277:27903–27911
Sharonov BP, Govorova NJu, Lyzlova SN (1988) A compara-
tive study of serum proteins ability to scavenge active
oxygen species: O2-. and OCl-. Biochem Int 17:783–790
Sharonov BP, Govorova NJu, Lyzlova SN (1989) Serum protein
degradation by hypochlorite. Biochem Int 19:27–35
Shiva S, Wang X, Ringwood LA et al (2006) Ceruloplasmin is a
NO oxidase and nitrite synthase that determines endocrine
NO homeostasis. Nat Chem Biol 2:486–493
Sokolov AV, Pulina MO, Zakharova ET et al (2005a) Effect of
lactoferrin on the ferroxidase activity of ceruloplasmin.
Biochem (Mosc) 70:1015–1019
Sokolov AV, Zakharova ET, Shavlovskiı MM et al (2005b)
Isolation of stable human ceruloplasmin and its interaction
with salmon protamine. Bioorg Khim 31:269–279
Sokolov AV, Pulina MO, Zakharova ET et al (2006) Identifi-
cation and isolation from breast milk of ceruloplasmin-
lactoferrin complex. Biochem (Mosc) 71:160–166
Sokolov AV, Pulina MO, Ageeva KV et al (2007a) Interaction
of ceruloplasmin, lactoferrin, and myeloperoxidase. Bio-
chem (Mosc) 72:409–415
Sokolov AV, Pulina MO, Ageeva KV et al (2007b) Identifica-
tion of leukocyte cationic proteins that interact with ceru-
loplasmin. Biochem (Mosc) 72:872–877
Sokolov AV, Ageeva KV, Pulina MO et al (2008) Ceruloplas-
min and myeloperoxidase in complex affect the enzymatic
properties of each other. Free Radic Res 42:989–998
Sokolov AV, Ageeva KV, Pulina MO et al (2009a) b Effect of
lactoferrin on oxidative features of ceruloplasmin. Bio-
metals 22:521–529
Sokolov AV, Pulina MO, Ageeva KV et al (2009b) Identifica-
tion of complexes formed by ceruloplasmin with matrix
metalloproteinases 2 and 12. Biochem (Mosc)
74:1388–1392
Sokolov AV, Prozorovski VN, Vasilyev VB (2009c) Study of
interaction of ceruloplasmin, lactoferrin and myeloperox-
idase by photon correlation spectroscopy. Biochem (Mosc)
74:1225–1227
Sokolov AV, Ageeva KV, Cherkalina OS et al (2010a) c Iden-
tification and properties of complexes formed by myelo-
peroxidase with lipoproteins and ceruloplasmin. Chem
Phys Lipids 163:347–355
Sokolov AV, Golenkina EA, Kostevich VA et al (2010b) a
Interaction of ceruloplasmin and 5-lipoxygenase. Biochem
(Mosc) 75:1464–1469
Sokolov AV, Ageeva KV, Kostevich VA et al (2010c) Study of
interaction of ceruloplasmin with serprocidins. Biochem
(Mosc) 75:1361–1367
Sokolov AV, Kostevich VA, Romanico DN et al (2012a) b Two-
stage method for purification of ceruloplasmin based on its
interaction with neomycin. Biochem (Mosc) 77:631–638
Sokolov AV, Solovyov KV, Kostevich VA et al (2012b) Pro-
tection of ceruloplasmin by lactoferrin against hydroxyl
radicals is pH dependent. Biochem Cell Biol 90:397–404
Sokolov AV, Pulina MO, Runova OL et al (2013) Complex of
ceruloplasmin and lactoferrin in human lacrimal fluid. Med
Acad J (Russ) 13:39–43
Sokolov AV, Kostevich VA, Runova OL et al (2014) Proath-
erogenic modification of LDL by surface-bound myelo-
peroxidase. Chem Phys Lipid 180:72–80
Stoj C, Kosman DJ (2003) Cuprous oxidase activity of yeast
Fet3p and human ceruloplasmin: implication for function.
FEBS Lett 554:422–426
Sun XL, Baker HM, Shewry SC et al (1999) Structure of
recombinant human lactoferrin expressed in Aspergillus
awamori. Acta Crystallogr D Biol Crystallogr 55:403–407
Tenovuo J (2002) Clinical applications of antimicrobial host
proteins lactoperoxidase, lysozyme and lactoferrin in
xerostomia: efficacy and safety. Oral Dis 8:23–29
Tiruppathi C, Naqvi T, Wu Y et al (2004) Albumin mediates the
transcytosis of myeloperoxidase by means of caveolae in
endothelial cells. Proc Natl Acad Sci U S A 101:7699–7704
van Dalen CJ, Whitehouse MW, Winterbourn CC et al (1997)
Thiocyanate and chloride as competing substrates for
myeloperoxidase. Biochem J 327:487–492
van der Does AM, Hensbergen PJ, Bogaards SJ et al (2012) The
human lactoferrin-derived peptide hLF1-11 exerts immu-
nomodulatory effects by specific inhibition of myeloper-
oxidase activity. J Immunol 188:5012–5019
Vasilyev VB, Kachurin AM, Soroka NV (1988) Dismutation of
superoxide radicals by ceruloplasmin–details of themechanism. Biokhimiya 53:2051–2058
Vassiliev V, Harris ZL, Zatta P (2005) Ceruloplasmin in neuro-
degenerative diseases. Brain Res Brain Res Rev 49:633–640
Vlasova II, Sokolov AV, Arnhold J (2012) The free amino acid
tyrosine enhances the chlorinating activity of human
myeloperoxidase. J Inorg Biochem 106:76–83
Voronina OV, Monakhov NK (1980) Estradiol-induced for-
mation of the polyribosomal complex synthesizing ceru-
loplasmin in rats. Biokhimiia 45:1010–1016
White KN, Conesa C, Sanchez L et al (2012) The transfer of iron
between ceruloplasmin and transferrins. Biochim Biophys
Acta 1820:411–416
Biometals
123

Xu PC, Chen M, Zhao MH (2012) High potential to reverse the
inhibition of myeloperoxidase by ceruloplasmin of anti-
myeloperoxidase autoantibodies of IgG3 subclass. Auto-
immunity 45:218–225
Xu PC, Li ZY, Yang XW et al (2013) Myeloperoxidase influ-
ences the complement regulatory function of modified
C-reactive protein. Innate Immun 20:440–448
Yang S, Hua Y, Nakamura T et al (2006) Up-regulation of brain
ceruloplasmin in thrombin preconditioning. Acta Neuro-
chir Suppl 96:203–206
Zakharova ET, Shavlovski MM, Bass MG et al (2000) Inter-
action of lactoferrin with ceruloplasmin. Arch Biochem
Biophys 374:222–228
Zakharova ET, Kostevich VA, Sokolov AV et al (2012) Human
apo-lactoferrin as a physiological mimetic of hypoxia
stabilizes hypoxia-inducible factor-1 alpha. Biometals
25:1247–1259
Biometals
123





























